
The Activin Branch Ligand Daw Regulates the Drosophila melanogaster Immune Response and Lipid Metabolism against the Heterorhabditis bacteriophora Serine Carboxypeptidase

{kind=link}
{kind=link}
{kind=link}
Abstract
1. Introduction
2. Results
2.1. H. bacteriophora Recombinant Serine Carboxypeptidase (rSCP) Induces Activin and Imd Signaling Activity in Drosophila
2.2. H. bacteriophora Recombinant Serine Carboxypeptidase Alters the Cellular Immunity and Survival Ability of Activin-Deficient Drosophila
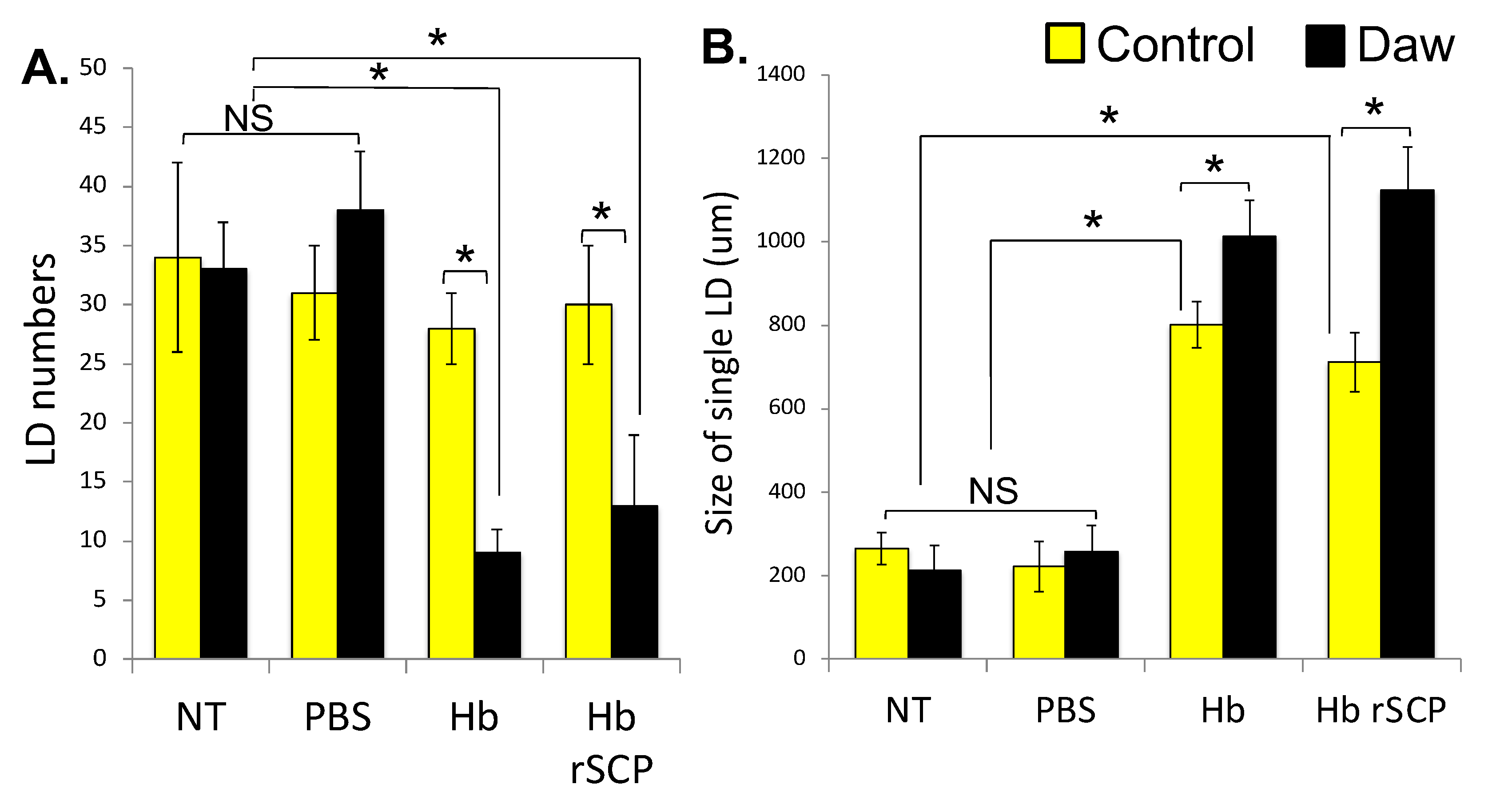
2.3. H. bacteriophora Infection of Activin-Deficient Drosophila Flies Results in Perturbed Intestinal Lipid Homeostasis
3. Discussion
4. Materials and Methods
4.1. Fly Stocks
4.2. Nematode and Bacterial Stocks
4.3. Production of Nematode Recombinant Serine Carboxypeptidase
4.4. Fly Infections
4.5. Gene Expression Analysis
4.6. Hemocyte Count Estimation
4.7. Lipid Droplet Count and Size Estimation
4.8. Statistical Analysis
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Brivio, M.F.; Mastore, M.; Pagani, M. Parasite-host relationship: A lesson from a professional killer. Invertebr. Surv. J. 2005, 2, 41–53. [Google Scholar]
- Castillo, J.C.; Reynolds, S.E.; Eleftherianos, I. Insect immune responses to nematode parasites. Trends Parasitol. 2011, 27, 537–547. [Google Scholar] [CrossRef] [PubMed]
- Eleftherianos, I.; Joyce, S.; Ffrench-Constant, R.H.; Clarke, D.J.; Reynolds, S.E. Probing the tri-trophic interaction between insects, nematodes and Photorhabdus. Parasitology 2010, 137, 1695–1706. [Google Scholar] [CrossRef] [PubMed]
- Erickson, S.M.; Xi, Z.; Mayhew, G.F.; Ramirez, J.L.; Aliota, M.T.; Christensen, B.M.; Dimopoulos, G. Mosquito infection responses to developing filarial worms. PLoS Negl. Trop. Dis. 2009, 3, e529. [Google Scholar] [CrossRef]
- Cupp, M.S.; Chen, Y.; Cupp, E.W. Cellular hemolymph response of Simulium vittatum (Diptera: Simuliidae) to intrathoracic injection of Onchocerca lienalis (Filarioidea: Onchocercidae) microfilariae. J. Med. Entomol. 1997, 34, 56–63. [Google Scholar] [CrossRef]
- Bartholomay, L.C.; Waterhouse, R.M.; Mayhew, G.F.; Campbell, C.L.; Michel, K.; Zou, Z.; Ramirez, J.L.; Das, S.; Alvarez, K.; Arensburger, P.; et al. Pathogenomics of Culex quinquefasciatus and meta-analysis of infection responses to diverse pathogens. Science 2010, 330, 88–90. [Google Scholar] [CrossRef] [PubMed]
- Kambris, Z.; Cook, P.E.; Phuc, H.K.; Sinkins, S.P. Immune activation by life-shortening Wolbachia and reduced filarial competence in mosquitoes. Science 2009, 326, 134–136. [Google Scholar] [CrossRef]
- Christensen, B.M.; Li, J.; Chen, C.C.; Nappi, A.J. Melanization immune responses in mosquito vectors. Trends Parasitol. 2005, 21, 192–199. [Google Scholar] [CrossRef] [PubMed]
- Wang, X.; Fuchs, J.F.; Infanger, L.C.; Rocheleau, T.A.; Hillyer, J.F.; Chen, C.C.; Christensen, B.M. Mosquito innate immunity: Involvement β1,3-glucan recognition protein in melanotic encapsulation immune responses in Armigeres subalbatus. Mol. Biochem. Parasitol. 2005, 139, 65–73. [Google Scholar] [CrossRef]
- Armer, C.A.; Rao, S.; Berry, R.E. Insect cellular and chemical limitations to pathogen development: The Colorado potato beetle, the nematode Heterorhabditis marelatus, and its symbiotic bacteria. J. Invertebr. Pathol. 2004, 87, 114–122. [Google Scholar] [CrossRef]
- van Sambeek, J.; Wiesner, A. Successful parasitation of locusts by entomopathogenic nematodes is correlated with inhibition of insect phagocytes. J. Invertebr. Pathol. 2009, 73, 154–161. [Google Scholar] [CrossRef]
- Liu, C.T.; Hou, R.F.; Chen, C.C. Formation of basement membrane-like structure terminates the cellular encapsulation of microfilariae in the haemocoel of Anopheles quadrimaculatus. Parasitology 1998, 116, 511–518. [Google Scholar] [CrossRef]
- Taylor, D.B.; Szalanski, A.L.; Adams, B.J.; Peterson, R.D., II. Susceptibility of house fly (Diptera: Muscidae) larvae to entomopathogenic nematodes (Rhabditida: Heterorhabditidae, Steinernematidae). Environ. Entomol. 1998, 27, 1514–1519. [Google Scholar] [CrossRef]
- Zhao, X.; Ferdig, M.T.; Li, J.; Christensen, B.M. Biochemical pathway of melanotic encapsulation of Brugia malayi in the mosquito, Armigeres subalbatus. Dev. Comp. Immunol. 1995, 19, 205–215. [Google Scholar] [CrossRef] [PubMed]
- Chalk, R.; Townson, H.; Natori, S.; Desmond, H.; Ham, P.J. Purification of an insect defensin from the mosquito, Aedes aegypti. Insect Biochem. Mol. Biol. 1994, 24, 403–410. [Google Scholar] [CrossRef] [PubMed]
- Vegni Talluri, M.; Cancrini, G. An ultrastructural study on the early cellular response to Dirofilaria immitis (Nematoda) in the Malpighian tubules of Aedes aegypti (refractory strains). Parasite 1994, 1, 343–348. [Google Scholar] [CrossRef] [PubMed]
- Christensen, B.M.; Forton, K.F. Hemocyte-mediated melanization of microfilariae in Aedes aegypti. J. Parasitol. 1986, 72, 220–225. [Google Scholar] [CrossRef]
- Lu, D.; Macchietto, M.; Chang, D.; Barros, M.M.; Baldwin, J.; Mortazavi, A.; Dillman, A.R. Activated entomopathogenic nematode infective juveniles release lethal venom proteins. PLoS Pathog. 2017, 13, e1006302. [Google Scholar] [CrossRef]
- Brivio, M.F.; Mastore, M.; Nappi, A.J. A pathogenic parasite interferes with phagocytosis of insect immunocompetent cells. Dev. Comp. Immunol. 2010, 34, 991–998. [Google Scholar] [CrossRef]
- Mastore, M.; Brivio, M.F. Cuticular surface lipids are responsible for disguise properties of an entomoparasite against host cellular responses. Dev. Comp. Immunol. 2008, 32, 1050–1062. [Google Scholar] [CrossRef]
- Li, X.Y.; Cowles, R.S.; Cowles, E.A.; Gaugler, R.; Cox-Foster, D.L. Relationship between the successful infection by entomopathogenic nematodes and the host immune response. Int. J. Parasitol. 2007, 37, 365–374. [Google Scholar] [CrossRef] [PubMed]
- Brivio, M.F.; Moro, M.; Mastore, M. Down-regulation of antibacterial peptide synthesis in an insect model induced by the body-surface of an entomoparasite (Steinernema feltiae). Dev. Comp. Immunol. 2006, 30, 627–638. [Google Scholar] [CrossRef] [PubMed]
- Brivio, M.F.; Mastore, M.; Moro, M. The role of Steinernema feltiae body-surface lipids in host-parasite immunological interactions. Mol. Biochem. Parasitol. 2004, 135, 111–121. [Google Scholar] [CrossRef] [PubMed]
- Brivio, M.F.; Pagani, M.; Restelli, S. Immune suppression of Galleria mellonella (Insecta, Lepidoptera) humoral defenses induced by Steinernema feltiae (Nematoda, Rhabditida): Involvement of the parasite cuticle. Exp. Parasitol. 2002, 101, 149–156. [Google Scholar] [CrossRef] [PubMed]
- Wang, Y.; Gaugler, R. Steinernema glaseri surface coat protein suppresses the immune response of Popillia japonica (Coleoptera: Scarabaeidae) larvae. Biol. Control 1999, 14, 45–50. [Google Scholar] [CrossRef]
- Jarosz, J. Active resistance of entomophagous rhabditid Heterorhabditis bacteriophora to insect immunity. Parasitology 1998, 117, 201–208. [Google Scholar] [CrossRef] [PubMed]
- Hallem, E.A.; Rengarajan, M.; Ciche, T.A.; Sternberg, P.W. Nematodes, bacteria, and flies: A tripartite model for nematode parasitism. Curr. Biol. 2007, 17, 898–904. [Google Scholar] [CrossRef] [PubMed]
- Peña, J.M.; Carrillo, M.A.; Hallem, E.A. Variation in the susceptibility of Drosophila to different entomopathogenic nematodes. Infect. Immun. 2015, 83, 1130–1138. [Google Scholar] [CrossRef]
- Castillo, J.C.; Shokal, U.; Eleftherianos, I. A novel method for infecting Drosophila adult flies with insect pathogenic nematodes. Virulence 2012, 3, 339–347. [Google Scholar] [CrossRef]
- Castillo, J.C.; Shokal, U.; Eleftherianos, I. Immune gene transcription in Drosophila adult flies infected by entomopathogenic nematodes and their mutualistic bacteria. J. Insect Physiol. 2013, 59, 179–185. [Google Scholar] [CrossRef]
- Arefin, B.; Kucerova, L.; Dobes, P.; Markus, R.; Strnad, H.; Wang, Z.; Hyrsl, P.; Zurovec, M.; Theopold, U. Genome-wide transcriptional analysis of Drosophila larvae infected by entomopathogenic nematodes shows involvement of complement, recognition and extracellular matrix proteins. J. Innate Immun. 2014, 6, 192–204. [Google Scholar] [CrossRef] [PubMed]
- Dobes, P.; Wang, Z.; Markus, R.; Theopold, U.; Hyrsl, P. An improved method for nematode infection assays in Drosophila larvae. Fly 2012, 6, 75–79. [Google Scholar] [CrossRef] [PubMed]
- Hyrsl, P.; Dobes, P.; Wang, Z.; Hauling, T.; Wilhelmsson, C.; Theopold, U. Clotting factors and eicosanoids protect against nematode infections. J. Innate Immun. 2011, 3, 65–70. [Google Scholar] [CrossRef]
- Wang, Z.; Wilhelmsson, C.; Hyrsl, P.; Loof, T.G.; Dobes, P.; Klupp, M.; Loseva, O.; Mörgelin, M.; Iklé, J.; Cripps, R.M.; et al. Pathogen entrapment by transglutaminase—A conserved early innate immune mechanism. PLoS Pathog. 2010, 6, e1000763. [Google Scholar] [CrossRef]
- Vadnal, J.; Ratnappan, R.; Keaney, M.; Kenney, E.; Eleftherianos, I.; O’Halloran, D.; Hawdon, J.M. Identification of candidate infection genes from the model entomopathogenic nematode Heterorhabditis bacteriophora. BMC Genom. 2017, 18, 8. [Google Scholar] [CrossRef] [PubMed]
- Bastin, A.; Eleftherianos, I. Functional role of the TGF-β signaling in the Drosophila immune response. Curr. Res. Immunol. 2023, 4, 100071. [Google Scholar] [CrossRef]
- Ozakman, Y.; Eleftherianos, I. TGF-beta Signaling Interferes with the Drosophila Innate Immune and Metabolic Response to Parasitic Nematode Infection. Front. Physiol. 2019, 10, 716. [Google Scholar] [CrossRef]
- Eleftherianos, I.; Castillo, J.C.; Patrnogic, J. TGF-β regulates resistance to parasitic nematode infection in Drosophila melanogaster. Immunobiology 2016, 221, 1362–1368. [Google Scholar] [CrossRef]
- Patrnogic, J.; Heryanto, C.; Eleftherianos, I. Wounding-induced upregulation of the Bone Morphogenic Protein signaling pathway in Drosophila promotes survival against parasitic nematode infection. Gene 2018, 673, 112–118. [Google Scholar] [CrossRef]
- Patrnogic, J.; Heruanto, C.; Eleftherianos, I. Transcriptional up-regulation of the TGF-β intracellular signaling transducer Mad of Drosophila larvae in response to parasitic nematode infection. Innate Immun. 2018, 24, 349–356. [Google Scholar] [CrossRef]
- Patrnogic, J.; Heryanto, C.; Ozakman, Y.; Eleftherianos, I. Transcript analysis reveals the requirement of NF-κB transcription factors for the activation of TGF-β signaling in nematode infected Drosophila. Immunogenetics 2019, 71, 501–510. [Google Scholar] [CrossRef] [PubMed]
- Kenney, E.; Hawdon, J.M.; O’Halloran, D.; Eleftherianos, I. Heterorhabditis bacteriophora Excreted-Secreted Products Enable Infection by Photorhabdus luminescens Through Suppression of the Imd Pathway. Front. Immunol. 2019, 10, 2372. [Google Scholar] [CrossRef] [PubMed]
- Harsh, S.; Heryanto, C.; Eleftherianos, I. Intestinal lipid droplets as novel mediators of host-pathogen interaction in Drosophila. Biol. Open 2019, 8, bio039040. [Google Scholar]
- Clark, R.I.; Woodcock, K.J.; Geissmann, F.; Trouillet, C.; Dionne, M.S. Multiple TGF-β superfamily signals modulate the adult Drosophila immune response. Curr. Biol. 2011, 21, 1672–1677. [Google Scholar] [CrossRef]
- Jones, K.; Tafesh-Edwards, G.; Kenney, E.; Toubarro, N.; Simões, N.; Eleftherianos, I. Excreted secreted products from the parasitic nematode Steinernema carpocapsae manipulate the Drosophila melanogaster immune response. Sci. Rep. 2022, 12, 14237. [Google Scholar] [CrossRef] [PubMed]
- Ozakman, Y.; Eleftherianos, I. Nematode infection and antinematode immunity in Drosophila. Trends Parasitol. 2021, 37, 1002–1013. [Google Scholar] [CrossRef] [PubMed]
- Eleftherianos, I.; Heryanto, C. Transcriptomic Insights into the Insect Immune Response to Nematode Infection. Genes 2021, 12, 202. [Google Scholar] [CrossRef] [PubMed]
- Dziedziech, A.; Shivankar, S.; Theopold, U. Drosophila melanogaster Responses against Entomopathogenic Nematodes: Focus on Hemolymph Clots. Insects 2020, 11, 62. [Google Scholar] [CrossRef]
- Ozakman, Y.; Raval, D.; Eleftherianos, I. Activin and BMP Signaling Activity Affects Different Aspects of Host Anti-Nematode Immunity in Drosophila melanogaster. Front. Immunol. 2021, 12, 795331. [Google Scholar] [CrossRef]
- White, G.F. A Method for Obtaining Infective Nematode Larvae from Cultures. Science 1927, 66, 302–303. [Google Scholar] [CrossRef]
- Heryanto, C.; Ratnappan, R.; O’Halloran, D.M.; Hawdon, J.C.; Eleftherianos, I. Culturing and Genetically Manipulating Entomopathogenic Nematodes. J. Vis. Exp. 2022, 181, e63885. [Google Scholar]
- Kenney, E.; Yaparla, A.; Hawdon, J.M.; O’Halloran, D.M.; Grayfer, L.; Eleftherianos, I. A putative lysozyme and serine carboxypeptidase from Heterorhabditis bacteriophora show differential virulence capacities in Drosophila melanogaster. Dev. Comp. Immunol. 2021, 114, 103820. [Google Scholar] [CrossRef] [PubMed]
- Livak, K.J.; Schmittgen, T.D. Analysis of relative gene expression data using real-time quantitative PCR and the 2−ΔΔCT method. Methods 2001, 25, 402–408. [Google Scholar] [CrossRef] [PubMed]
- Schmittgen, T.D.; Livak, K.J. Analyzing real-time PCR data by the comparative CT method. Nat. Protoc. 2008, 3, 1101–1108. [Google Scholar] [CrossRef] [PubMed]


Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Mallick, S.; Kenney, E.; Eleftherianos, I. The Activin Branch Ligand Daw Regulates the Drosophila melanogaster Immune Response and Lipid Metabolism against the Heterorhabditis bacteriophora Serine Carboxypeptidase. Int. J. Mol. Sci. 2024, 25, 7970. https://doi.org/10.3390/ijms25147970
Mallick S, Kenney E, Eleftherianos I. The Activin Branch Ligand Daw Regulates the Drosophila melanogaster Immune Response and Lipid Metabolism against the Heterorhabditis bacteriophora Serine Carboxypeptidase. International Journal of Molecular Sciences. 2024; 25(14):7970. https://doi.org/10.3390/ijms25147970
Chicago/Turabian StyleMallick, Sreeradha, Eric Kenney, and Ioannis Eleftherianos. 2024. "The Activin Branch Ligand Daw Regulates the Drosophila melanogaster Immune Response and Lipid Metabolism against the Heterorhabditis bacteriophora Serine Carboxypeptidase" International Journal of Molecular Sciences 25, no. 14: 7970. https://doi.org/10.3390/ijms25147970
APA StyleMallick, S., Kenney, E., & Eleftherianos, I. (2024). The Activin Branch Ligand Daw Regulates the Drosophila melanogaster Immune Response and Lipid Metabolism against the Heterorhabditis bacteriophora Serine Carboxypeptidase. International Journal of Molecular Sciences, 25(14), 7970. https://doi.org/10.3390/ijms25147970