
Divergent Roles of the Auxin Response Factors in Lemongrass (Cymbopogon flexuosus (Nees ex Steud.) W. Watson) during Plant Growth
 , , , ,
, , , ,
Abstract
:1. Introduction
2. Results and Discussion
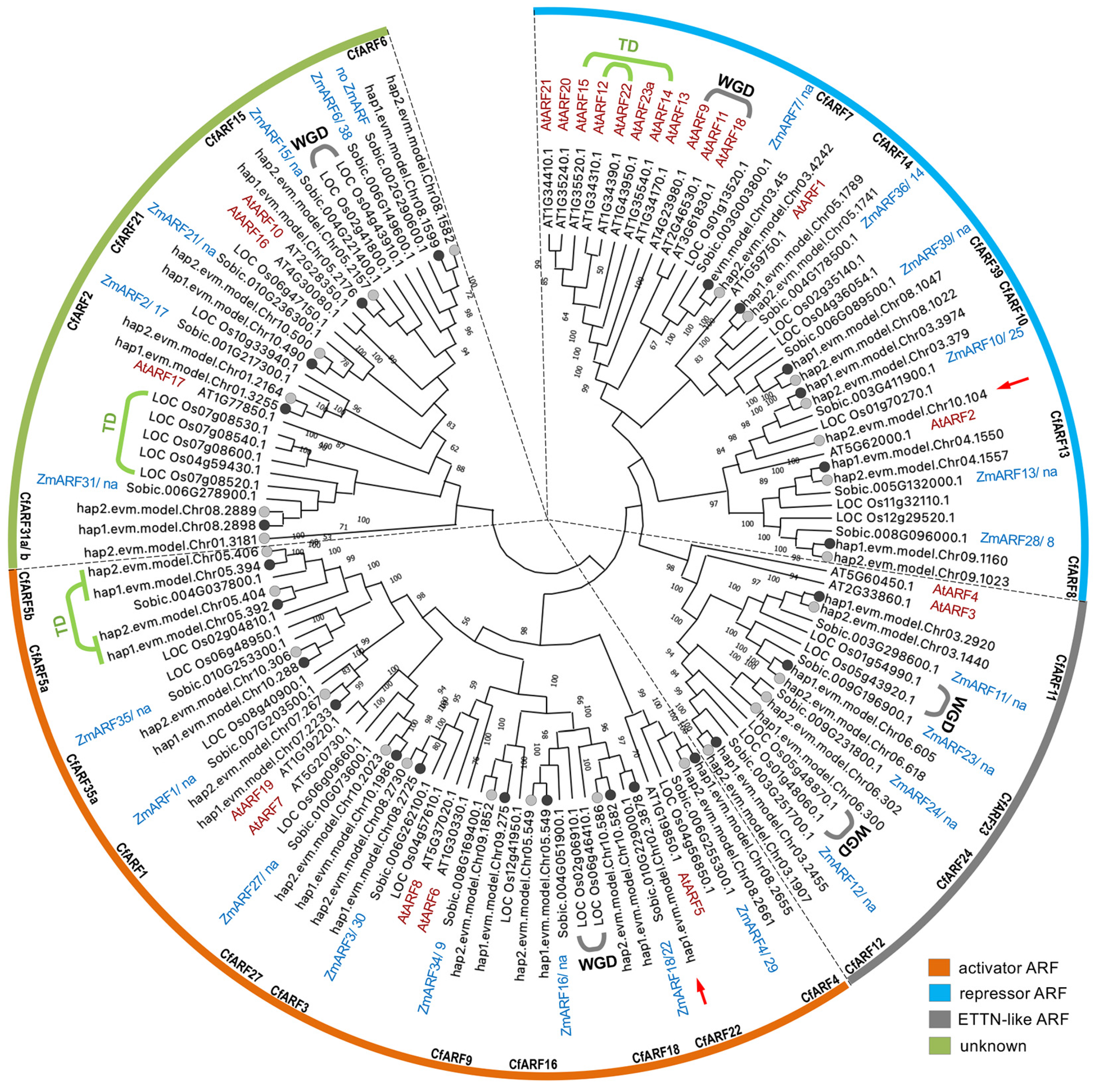
2.1. A Catalog of CfARFs in the Lemongrass Genome
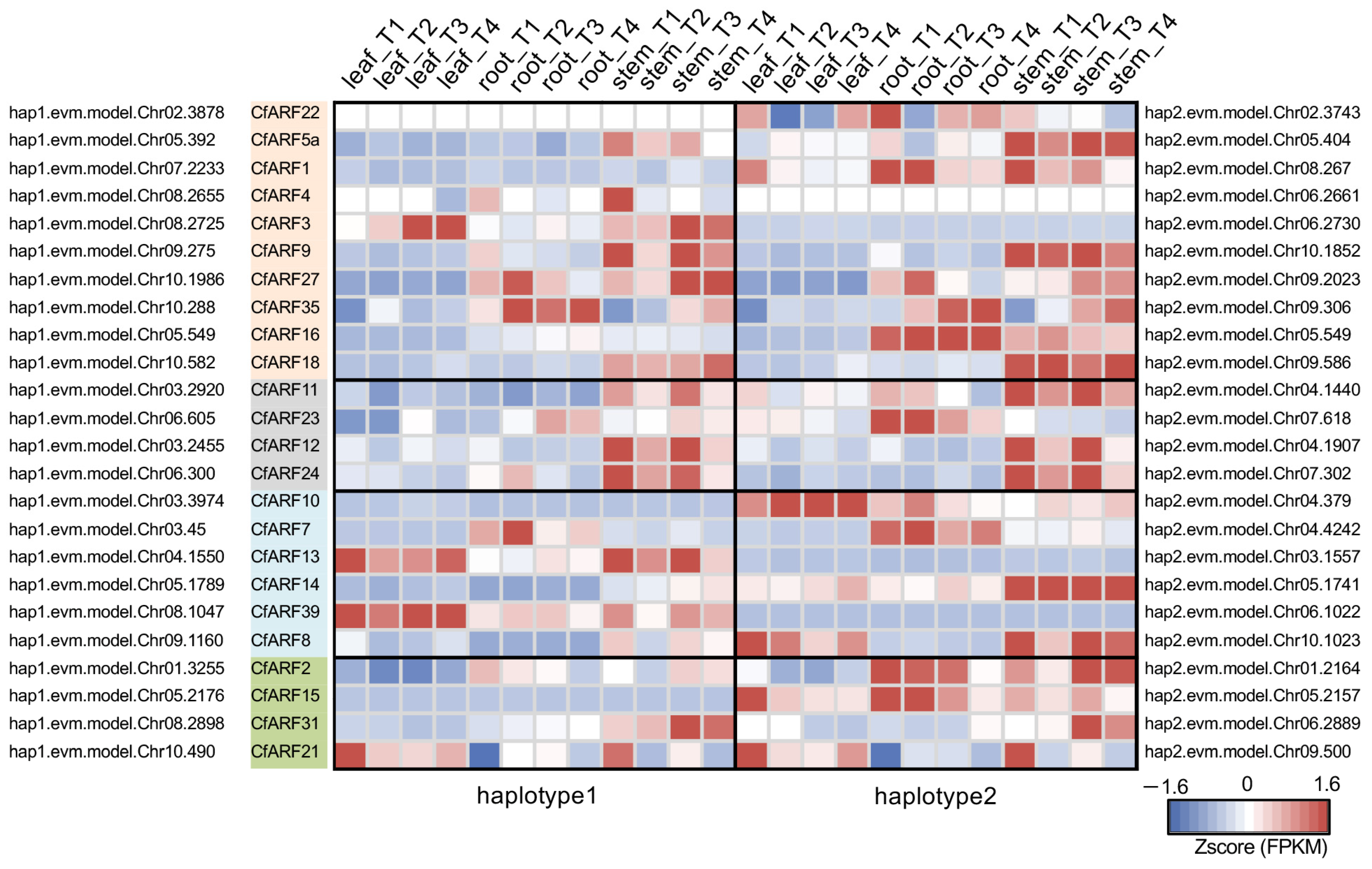
2.2. Expression Patterns of CfARFs during the Plant Growth of Lemongrass
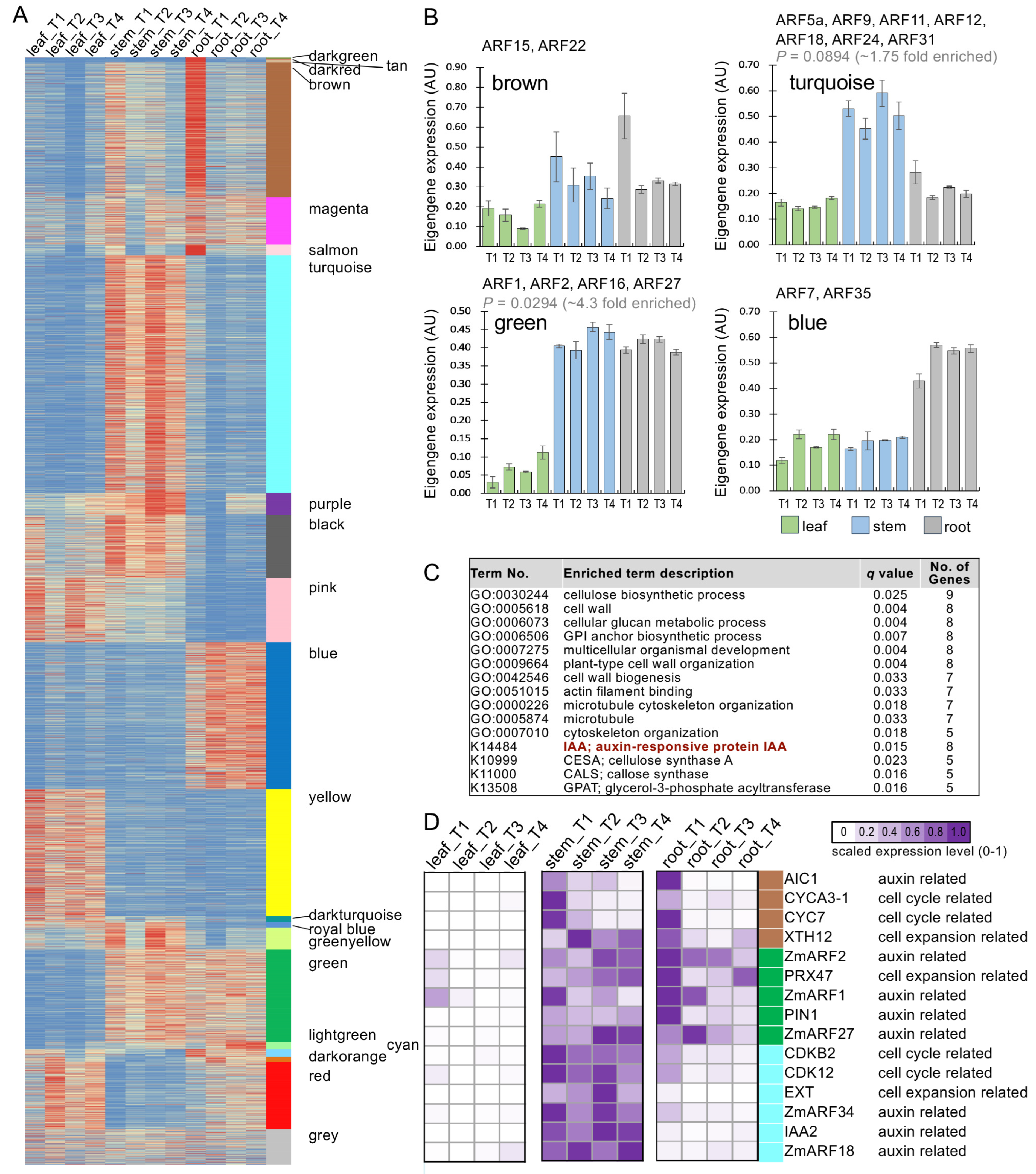
2.3. Co-Expression Analysis of CfARF Genes
2.4. Divergence of CfARFs
3. Materials and Methods
3.1. Plant Materials
3.2. Identification and Phylogenetic Analysis of CfARF Genes
3.3. Analyses of the Molecular Properties of CfARF Proteins
3.4. Sequence Analysis of CfARF Proteins
3.5. Transcriptome of the Lemongrass and Coexpression Network Analysis
3.6. qPCR-Based Expression Profiling of CfARFs
3.7. Statistically Analysis
4. Conclusions
Supplementary Materials
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Briggs, W.R. Mediation of phototropic responses of corn coleoptiles by lateral transport of auxin. Plant Physiol. 1963, 38, 237–247. [Google Scholar] [CrossRef]
- Cowling, C.L.; Dash, L.; Kelley, D.R. Roles of auxin pathways in maize biology. J. Exp. Bot. 2023, 74, 6989–6999. [Google Scholar] [CrossRef]
- Finet, C.; Berne-Dedieu, A.; Scutt, C.P.; Marletaz, F. Evolution of the ARF gene family in land plants: Old domains, new tricks. Mol. Biol. Evol. 2012, 30, 45–56. [Google Scholar] [CrossRef]
- Ramos, B.R.; Nemhauser, J.L. Expansion and innovation in auxin signaling: Where do we grow from here? Development 2021, 148, dev187120. [Google Scholar] [CrossRef]
- Matthes, M.S.; Best, N.B.; Robil, J.M.; Malcomber, S.; Gallavotti, A.; McSteen, P. Auxin EvoDevo: Conservation and diversification of genes regulating auxin biosynthesis, transport, and signaling. Mol. Plant 2019, 12, 298–320. [Google Scholar] [CrossRef]
- Weijers, D.; Wagner, D. Transcriptional responses to the auxin hormone. Annu. Rev. Plant Biol. 2016, 67, 539–574. [Google Scholar] [CrossRef]
- Roosjen, M.; Paque, S.; Weijers, D. Auxin response factors: Output control in auxin biology. J. Exp. Bot. 2017, 69, 179–188. [Google Scholar] [CrossRef]
- Mutte, S.K.; Kato, H.; Rothfels, C.; Melkonian, M.; Wong, G.K.S.; Weijers, D. Origin and evolution of the nuclear auxin response system. eLife 2018, 7, e33399. [Google Scholar] [CrossRef]
- Galli, M.; Khakhar, A.; Lu, Z.; Chen, Z.; Sen, S.; Joshi, T.; Nemhauser, J.L.; Schmitz, R.J.; Gallavotti, A. The DNA binding landscape of the maize AUXIN RESPONSE FACTOR family. Nat. Commun. 2018, 9, 4526. [Google Scholar] [CrossRef]
- Ulmasov, T.; Hagen, G.; Guilfoyle, T.J. Dimerization and DNA binding of auxin response factors. Plant J. 1999, 19, 309–319. [Google Scholar] [CrossRef]
- Kato, H.; Ishizaki, K.; Kouno, M.; Shirakawa, M.; Bowman, J.L.; Nishihama, R.; Kohchi, T. Auxin-mediated transcriptional system with a minimal set of components is critical for morphogenesis through the life cycle in Marchantia polymorpha. PLoS Genet. 2015, 11, e1005084. [Google Scholar] [CrossRef]
- Boer, D.R.; Freire-Rios, A.; Van den Berg, W.A.; Saakim, T.; Manfield, I.W.; Kepinski, S.; Lopez-Vidrieo, I.; Franco-Zorrilla, J.M.; de Vries, S.C.; Solano, R. Structural basis for DNA binding specificity by the auxin-dependent ARF transcription factors. Cell 2014, 156, 577–589. [Google Scholar] [CrossRef]
- Rienstra, J.; Hernandez-Garcia, J.; Weijers, D. To bind or not to bind: How AUXIN RESPONSE FACTORS select their target genes. J. Exp. Bot. 2023, 74, 6922–6932. [Google Scholar] [CrossRef]
- Kato, H.; Mutte, S.K.; Suzuki, H.; Crespo, I.; Das, S.; Radoeva, T.; Fontana, M.; Yoshitake, Y.; Hainiwa, E.; van den Berg, W.; et al. Design principles of a minimal auxin response system. Nat. Plants 2020, 6, 473–482. [Google Scholar] [CrossRef]
- Okushima, Y.; Overvoorde, P.J.; Arima, K.; Alonso, J.M.; Chan, A.; Chang, C.; Ecker, J.R.; Hughes, B.; Lui, A.; Nguyen, D.; et al. Functional genomic analysis of the AUXIN RESPONSE FACTOR gene family members in Arabidopsis thaliana: Unique and overlapping functions of ARF7 and ARF19. Plant Cell 2005, 17, 444–463. [Google Scholar] [CrossRef]
- Kelley, D.R.; Arreola, A.; Gallagher, T.L.; Gasser, C.S. ETTIN (ARF3) physically interacts with KANADI proteins to form a functional complex essential for integument development and polarity determination in Arabidopsis. Development 2012, 139, 1105–1109. [Google Scholar] [CrossRef]
- Powers, S.K.; Holehouse, A.S.; Korasick, D.A.; Schreiber, K.H.; Clark, N.M.; Jing, H.; Emenecker, R.; Han, S.; Tycksen, E.; Hwang, I.; et al. Nucleo-cytoplasmic partitioning of ARF proteins controls auxin responses in Arabidopsis thaliana. Mol. Cell 2019, 76, 177–190. [Google Scholar] [CrossRef] [PubMed]
- Nagpal, P.; Ellis, C.M.; Weber, H.; Ploense, S.E.; Barkawi, L.S.; Guilfoyle, T.J.; Hagen, G.; Alonso, J.M.; Cohen, J.D.; Farmer, E.E.; et al. Auxin response factors ARF6 and ARF8 promote jasmonic acid production and flower maturation. Development 2005, 132, 4107–4118. [Google Scholar] [CrossRef]
- Li, Y.; Han, S.; Qi, Y. Advances in structure and function of auxin response factor in plants. J. Integr. Plant Biol. 2022, 65, 617–632. [Google Scholar] [CrossRef]
- Galli, M.; Liu, Q.; Moss, B.L.; Malcomber, S.; Li, W.; Gaines, C.; Federici, S.; Roshkovan, J.; Meeley, R.; Nemhauser, J.L.; et al. Auxin signaling modules regulate maize inflorescence architecture. Proc. Natl. Acad. Sci. USA 2015, 112, 13372–13377. [Google Scholar] [CrossRef]
- Wang, Y.; Xu, J.; Yu, J.; Zhu, D.; Zhao, Q. Maize GSK3-like kinase ZmSK2 is involved in embryonic development. Plant Sci. 2022, 318, 111221. [Google Scholar] [CrossRef]
- Wang, H.; Huang, Y.; Li, Y.; Cui, Y.; Xiang, X.; Zhu, Y.; Wang, Q.; Wang, X.; Ma, G.; Xiao, Q.; et al. An ARF gene mutation creates flint kernel architecture in dent maize. Nat. Commun. 2024, 15, 2565. [Google Scholar] [CrossRef]
- Liu, Y.; Jiang, H.; Chen, W.; Qian, Y.; Ma, Q.; Cheng, B.; Zhu, S. Genome-wide analysis of the auxin response factor (ARF) gene family in maize (Zea mays). Plant Growth Regul. 2011, 63, 225–234. [Google Scholar] [CrossRef]
- Xing, H.; Pudake, R.N.; Guo, G.; Xing, G.; Hu, Z.; Zhang, Y.; Sun, Q.; Ni, Z. Genome-wide identification and expression profiling of auxin response factor (ARF) gene family in maize. BMC Genom. 2011, 12, 178. [Google Scholar] [CrossRef]
- Wang, Y.; Deng, D.; Shi, Y.; Miao, N.; Bian, Y.; Yin, Z. Diversification, phylogeny and evolution of auxin response factor (ARF) family: Insights gained from analyzing maize ARF genes. Mol. Biol. Rep. 2012, 39, 2401–2415. [Google Scholar] [CrossRef]
- Wang, S.K.; Bai, Y.H.; Shen, C.J.; Wu, Y.R.; Zhang, S.N.; Jiang, D.A.; Guilfoyle, T.J.; Chen, M.; Qi, Y.H. Auxin-related gene families in abiotic stress response in Sorghum bicolor. Funct. Integr. Genom. 2010, 10, 533–546. [Google Scholar] [CrossRef]
- Chen, D.; Wang, W.; Wu, Y.; Xie, H.; Zhao, L.; Zeng, Q.; Zhan, Y. Expression and distribution of the Auxin Response Factors in Sorghum bicolor during development and temperature stress. Int. J. Mol. Sci. 2019, 20, 4816. [Google Scholar] [CrossRef]
- Gupta, B.K. Chromosome numbers in some species of Cymbopogon. Proc. Indian Acad. Sci. 1965, 62, 155–157. [Google Scholar] [CrossRef]
- Ganjewala, D.; Luthra, R. Essential oil biosynthesis and regulation in the genus Cymbopogon. Nat. Prod. Commun. 2010, 5, 163–172. [Google Scholar] [CrossRef] [PubMed]
- Meena, S.; Kumar, S.R.; Rao, D.K.V.; Dwivedi, V.; Shilpashree, H.B.; Rastogi, S.; Shasany, A.K.; Nagegowda, D.A. De novo sequencing and analysis of lemongrass transcriptome provide first insights into the essential oil biosynthesis of aromatic grasses. Front. Plant Sci. 2016, 7, 1129. [Google Scholar] [CrossRef]
- Welker, C.A.D.; McKain, M.R.; Estep, M.C.; Pasquet, R.S.; Chipabika, G.; Pallangyo, B.; Kellogg, E.A. Phylogenomics enables biogeographic analysis and a new subtribal classification of Andropogoneae (Poaceae-Panicoideae). J. Syst. Evol. 2020, 58, 1003–1030. [Google Scholar] [CrossRef]
- Chakravartty, N.; Randowski, L.; Pirro, S. The complete genome sequence of Cymbopogon citratus (Poaceae, Poales), lemongrass. Biodivers. Genomes 2023, 4, 28–29. [Google Scholar] [CrossRef]
- Li, Y. A haplotype-resolved genome assembly of Cymbopogon flexuosus with the gene annotation and coding sequences. Figshare 2024, 1, 1–7. [Google Scholar] [CrossRef]
- Paterson, A.H.; Bowers, J.E.; Chapman, B.A. Ancient polyploidization predating divergence of the cereals, and its consequences for comparative genomics. Proc. Natl. Acad. Sci. USA 2004, 101, 9903–9908. [Google Scholar] [CrossRef] [PubMed]
- Wendel, J.F.; Jackson, S.A.; Meyers, B.C.; Wing, R.A. Evolution of plant genome architecture. Genome Biol. 2016, 17, 37. [Google Scholar] [CrossRef] [PubMed]
- Zhang, Y.; Ngu, D.W.; Carvalho, D.; Liang, Z.; Qiu, Y.; Roston, R.L.; Schnable, J.C. Differentially regulated orthologs in sorghum and the subgenomes of maize. Plant Cell 2017, 29, 1938–1951. [Google Scholar] [CrossRef] [PubMed]
- Paterson, A.H.; Bowers, J.E.; Bruggmann, R.; Dubchak, I.; Grimwood, J.; Gundlach, H.; Haberer, G.; Hellsten, U.; Mitros, T.; Poliakov, A.; et al. The Sorghum bicolor genome and the diversification of grasses. Nature 2009, 457, 551–556. [Google Scholar] [CrossRef]
- Bowers, J.E.; Chapman, B.A.; Rong, J.; Paterson, A.H. Unravelling angiosperm genome evolution by phylogenetic analysis of chromosomal duplication events. Nature 2003, 422, 433–438. [Google Scholar] [CrossRef]
- Panchy, N.; Lehti-Shiu, M.; Shiu, S.H. Evolution of gene duplication in plants. Plant Physiol. 2016, 171, 2294–2316. [Google Scholar] [CrossRef]
- Guilfoyle, T.J. The PB1 Domain in Auxin Response Factor and Aux/IAA Proteins: A versatile protein interaction module in the auxin response. Plant Cell 2015, 27, 33–43. [Google Scholar] [CrossRef]
- Ulmasov, T.; Hagen, G.; Guilfoyle, T.J. ARF1, a transcription factor that binds to auxin response elements. Science 1997, 276, 1865–1868. [Google Scholar] [CrossRef] [PubMed]
- Ulmasov, T.; Murfett, J.; Hagen, G.; Guilfoyle, T.J. Aux/IAA proteins repress expression of reporter genes containing natural and highly active synthetic auxin response elements. Plant Cell 1997, 9, 1963–1971. [Google Scholar] [CrossRef] [PubMed]
- Ulmasov, T.; Hagen, G.; Guilfoyle, T.J. Activation and repression of transcription by auxin-response factors. Proc. Natl. Acad. Sci. USA 1999, 96, 58445849. [Google Scholar] [CrossRef] [PubMed]
- Wang, F.; Yu, Z.; Zhang, M.; Wang, M.; Lu, X.; Liu, X.; Li, Y.; Zhang, X.; Tan, B.; Li, C.; et al. ZmTE1 promotes plant height by regulating intercalary meristem formation and internode cell elongation in maize. Plant Biotechnol. J. 2022, 20, 526–537. [Google Scholar] [CrossRef]
- Nemhauser, J.L.; Feldman, L.J.; Zambryski, P.C. Auxin and ETTIN in Arabidopsis gynoecium morphogenesis. Development 2000, 127, 3877–3888. [Google Scholar] [CrossRef] [PubMed]
- Simonini, S.; Deb, J.; Moubayidin, L.; Stephenson, P.; Valluru, M.; Freire-Rios, A.; Sorefan, K.; Weijers, D.; Friml, J.; Østergaard, L. A noncanonical auxin-sensing mechanism is required for organ morphogenesis in Arabidopsis. Genes Dev. 2016, 30, 2286–2296. [Google Scholar] [CrossRef] [PubMed]
- Simonini, S.; Deb, J.; Moubayidin, L.; Stephenson, P. Auxin-induced modulation of ETTIN activity orchestrates gene expression in Arabidopsis. Plant Cell 2017, 29, 1864–1882. [Google Scholar] [CrossRef]
- Bruggmann, R.; Bharti, A.K.; Gundlach, H.; Lai, J.; Young, S.; Pontaroli, A.C. Uneven chromosome contraction and expansion in the maize genome. Genome Res. 2006, 16, 1241–1251. [Google Scholar] [CrossRef] [PubMed]
- Schnable, J.C.; Springer, N.M.; Freeling, M. Differentiation of the maize subgenomes by genome dominance and both ancient and ongoing gene loss. Proc. Natl. Acad. Sci. USA 2011, 108, 4069–4074. [Google Scholar] [CrossRef]
- Sang, J.; Zou, D.; Wang, Z.; Wang, F.; Zhang, Y.; Xia, L.; Li, Z.; Ma, L.; Li, M.; Xu, B.; et al. IC4R-2.0: Rice Genome Reannotation Using Massive RNA-seq Data. Genom. Proteom. Bioinform. 2020, 18, 161–172. [Google Scholar] [CrossRef]
- Huang, F.; Jiang, Y.; Chen, T.; Li, H.; Fu, M.; Wang, Y.; Xu, Y.; Li, Y.; Zhou, Z.; Jia, L.; et al. New data and new features of the FunRiceGenes (Functionally Characterized Rice Genes) database: 2021 update. Rice 2022, 15, 23. [Google Scholar] [CrossRef] [PubMed]
- Wang, D.; Pei, K.; Fu, Y.; Sun, Z.; Li, S.; Liu, H.; Tang, K.; Han, B.; Tao, Y. Genome-wide analysis of the auxin response factors (ARF) gene family in rice (Oryza sativa). Gene 2007, 394, 13–24. [Google Scholar] [CrossRef]
- Sudhir, K.; Glen, S.; Michael, L.; Christina, K.; Koichiro, T. MEGA X: Molecular evolutionary genetics analysis across computing platforms. Mol. Biol. Evol. 2018, 35, 1547–1549. [Google Scholar] [CrossRef]
- Hall, B.G. Building phylogenetic trees from molecular data with MEGA. Mol. Biol. Evol. 2013, 30, 1229–1235. [Google Scholar] [CrossRef] [PubMed]
- Li, Y.; Wang, W.; Feng, Y.; Tu, M.; Wittich, P.E.; Bate, N.J.; Messing, J. Transcriptome and metabolome reveal distinct carbon allocation patterns during internode sugar accumulation in different sorghum genotypes. Plant Biotechnol. J. 2019, 17, 472–487. [Google Scholar] [CrossRef] [PubMed]
- Zhi, J.; Zeng, J.; Wang, Y.; Zhao, H.; Wang, G.; Guo, J.; Wang, Y.; Chen, M.; Yang, G.; He, G.; et al. A multi-omic resource of wheat seed tissues for nutrient deposition and improvement for human health. Sci. Data 2023, 10, 269. [Google Scholar] [CrossRef] [PubMed]
- Tu, M.; Zeng, J.; Zhang, J.; Fan, G.; Song, G. Unleashing the power within short-read RNA-seq for plant research: Beyond differential expression analysis and toward regulomics. Front. Plant Sci. 2022, 13, 1038109. [Google Scholar] [CrossRef] [PubMed]
- Wang, Y.; Tang, H.; Debarry, J.D.; Tan, X.; Li, J.; Wang, X.; Lee, T.H.; Jin, H.; Marler, B.; Guo, H.; et al. MCScanX: A toolkit for detection and evolutionary analysis of gene synteny and collinearity. Nucleic Acids Res. 2012, 40, e49. [Google Scholar] [CrossRef] [PubMed]
- Dobin, A.; Davis, C.A.; Schlesinger, F.; Drenkow, J.; Zaleski, C.; Jha, S.; Batut, P.; Chaisson, M.; Gingeras, T.R. STAR: Ultrafast universal RNA-seq aligner. Bioinformatics 2013, 29, 15–21. [Google Scholar] [CrossRef] [PubMed]
- Han, X.; Zhang, Y.; Zhang, Q.; Ma, N.; Liu, X.; Tao, W.; Lou, Z.; Zhong, C.; Deng, X.W.; Li, D.; et al. Two haplotype-resolved, gap-free genome assemblies for Actinidia latifolia and Actinidia chinensis shed light on the regulatory mechanisms of vitamin C and sucrose metabolism in kiwifruit. Mol. Plant 2023, 16, 452–470. [Google Scholar] [CrossRef]
- Li, H.; Handsaker, B.; Wysoker, A.; Fennell, T.; Ruan, J.; Homer, N.; Marth, G.; Abecasis, G.; Durbin, R.; 1000 Genome Project Data Processing Subgroup. The sequence alignment/map format and SAMtools. Bioinformatics 2009, 25, 2078–2079. [Google Scholar] [CrossRef] [PubMed]
- Hu, W.; Ji, C.; Shi, H.; Liang, Z.; Ding, Z.; Ye, J.; Ou, W.; Zhou, G.; Tie, W.; Yan, Y.; et al. Allele-defined genome reveals biallelic differentiation during cassava evolution. Mol. Plant 2021, 14, 851–854. [Google Scholar] [CrossRef] [PubMed]
- Langfelder, P.; Horvath, S. WGCNA: An R package for weighted correlation network analysis. BMC Bioinform. 2008, 9, 559. [Google Scholar] [CrossRef] [PubMed]
- Langfelder, P.; Horvath, S. Fast R functions for robust correlations and hierarchical clustering. J. Stat. Softw. 2012, 46, i11. [Google Scholar] [CrossRef]
- Wu, T.; Hu, E.; Xu, S.; Chen, M.; Guo, P.; Dai, Z.; Feng, T.; Zhou, L.; Tang, W.; Zhan, L.; et al. clusterProfiler 4.0: A universal enrichment tool for interpreting omics data. Innovation 2021, 2, 100141. [Google Scholar] [CrossRef]




{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| CfARFs | Gene ID | Chr. | Hap. | Phylogenetic Clade | CDS Length (bp) | Prot. Length (aa) | MW (Da) | pI | DD1 | B3 | DD2 | Q | LFG | III | IV | NCBI Accession |
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| 27 | hap1.Chr10.1986 | 10 | 1 | activator | 3168 | 1055 | 263,067.26 | 4.81 | √ | √ | √ | √ | √ | √ | √ | PP763312 |
| 27 | hap2.Chr10.2023 | 10 | 2 | activator | 3168 | 1055 | 262,997.19 | 4.81 | √ | √ | √ | √ | √ | √ | √ | PP763338 |
| 1 | hap1.Chr07.2233 | 7 | 1 | activator | 3258 | 1085 | 270,104.51 | 4.81 | √ | √ | √ | √ | √ | √ | √ | PP763313 |
| 1 | hap2.Chr07.267 | 7 | 2 | activator | 3258 | 1085 | 270,104.51 | 4.81 | √ | √ | √ | √ | √ | √ | √ | PP763339 |
| 9 | hap1.Chr09.275 | 9 | 1 | activator | 2691 | 896 | 223,388.16 | 4.85 | √ | √ | √ | √ | ACG | √ | √ | PP763314 |
| 9 | hap2.Chr09.1852 | 9 | 2 | activator | 2691 | 896 | 223,342.08 | 4.85 | √ | √ | √ | √ | ACG | √ | √ | PP763341 |
| 18 | hap1.Chr10.582 | 10 | 1 | activator | 2754 | 917 | 229,989.87 | 4.83 | √ | √ | √ | √ | √ | √ | √ | PP763316 |
| 18 | hap2.Chr10.586 | 10 | 2 | activator | 2754 | 917 | 230,160.25 | 4.83 | √ | √ | √ | √ | √ | √ | √ | PP763342 |
| 35 | hap1.Chr10.288 | 10 | 1 | activator | 3495 | 1164 | 290,444.40 | 4.78 | √ | √ | √ | √ | √ | √ | √ | PP763317 |
| 35 | hap2.Chr10.306 | 10 | 2 | activator | 3468 | 1155 | 288,415.02 | 4.78 | √ | √ | √ | √ | √ | √ | √ | PP763343 |
| 3 | hap1.Chr08.2725 | 8 | 1 | activator | 2454 | 817 | 204,118.89 | 4.85 | √ | √ | √ | √ | √ | √ | √ | PP763321 |
| 3 | hap2.Chr08.2730 | 8 | 2 | activator | 2175 | 724 | 180,852.58 | 4.89 | P | √ | √ | √ | √ | √ | √ | PP763347 |
| 4 | hap1.Chr08.2655 | 8 | 1 | activator | 2838 | 945 | 234,788.26 | 4.84 | √ | √ | √ | X | X | √ | √ | PP763323 |
| 4 | hap2.Chr08.2661 | 8 | 2 | activator | 2838 | 945 | 234,788.26 | 4.84 | √ | √ | √ | X | X | √ | √ | PP763349 |
| 5a | hap1.Chr05.392 | 5 | 1 | activator | 2403 | 800 | 198,766.86 | 4.88 | √ | √ | √ | √ | √ | √ | X | PP763329 |
| 5a | hap2.Chr05.404 | 5 | 2 | activator | 1242 | 413 | 102,288.83 | 5.02 | √ | √ | √ | X | X | P | X | PP763353 |
| 5b | hap1.Chr05.394 | 5 | 1 | activator | 3447 | 1148 | 285,393.94 | 4.79 | √ | √ | √ | √ | √ | √ | √ | PP763318 |
| 5b | hap2.Chr05.406 | 5 | 2 | activator | 3444 | 1147 | 285,142.60 | 4.79 | √ | √ | √ | √ | √ | √ | √ | PP763344 |
| 16 | hap1.Chr05.549 | 5 | 1 | activator | 2739 | 912 | 228,594.64 | 4.84 | √ | √ | √ | √ | √ | √ | √ | PP763319 |
| 16 | hap2.Chr05.549 | 5 | 2 | activator | 2739 | 912 | 228,566.59 | 4.84 | √ | √ | √ | √ | √ | √ | √ | PP763345 |
| 22 | hap1.Chr02.3878 | 2 | 1 | activator | 426 | 141 | 35,663.89 | 5.24 | X | X | P | X | X | X | X | PP763336 |
| 7 | hap2.Chr03.4242 | 3 | 1 | repressor | 2079 | 692 | 169,965.69 | 4.91 | √ | √ | √ | X | √ | √ | √ | PP763359 |
| 7 | hap1.Chr03.45 | 3 | 2 | repressor | 2079 | 692 | 169,963.65 | 4.91 | √ | √ | √ | X | √ | √ | √ | PP763332 |
| 8 | hap1.Chr09.1160 | 9 | 1 | repressor | 2511 | 836 | 206,575.17 | 4.87 | √ | √ | √ | X | √ | √ | √ | PP763324 |
| 8 | hap2.Chr09.1023 | 9 | 2 | repressor | 2517 | 838 | 207,017.58 | 4.87 | √ | √ | √ | X | √ | √ | √ | PP763350 |
| 10a | hap1.Chr03.3974 | 3 | 1 | repressor | 2430 | 809 | 200,683.10 | 4.86 | √ | √ | √ | X | √ | √ | √ | PP763322 |
| 10a | hap2.Chr03.379 | 3 | 2 | repressor | 2430 | 809 | 200,729.19 | 4.86 | √ | √ | √ | X | √ | √ | √ | PP763348 |
| 10b | hap2.Chr10.104 | 10 | 2 | repressor | 2559 | 852 | 212,894.83 | 4.85 | √ | √ | √ | X | √ | √ | √ | PP763358 |
| 13 | hap1.Chr04.1550 | 4 | 1 | repressor | 2565 | 854 | 209,962.57 | 4.86 | √ | √ | √ | X | √ | √ | √ | PP763326 |
| 13 | hap2.Chr04.1557 | 4 | 2 | repressor | 2565 | 854 | 210,020.65 | 4.86 | √ | √ | √ | X | √ | √ | √ | PP763352 |
| 14 | hap1.Chr05.1789 | 5 | 1 | repressor | 1977 | 658 | 163,664.80 | 4.92 | √ | √ | √ | X | √ | √ | √ | PP763325 |
| 14 | hap2.Chr05.1741 | 5 | 2 | repressor | 1977 | 658 | 163,708.85 | 4.92 | √ | √ | √ | X | √ | √ | √ | PP763351 |
| 39 | hap1.Chr08.1047 | 3 | 1 | repressor | 1986 | 661 | 164,430.61 | 4.92 | √ | √ | √ | X | √ | √ | √ | PP763328 |
| 39 | hap2.Chr08.1022 | 3 | 2 | repressor | 1986 | 661 | 164,462.67 | 4.92 | √ | √ | √ | X | √ | √ | √ | PP763355 |
| 12 | hap1.Chr03.2455 | 3 | 1 | ETTN-like | 2142 | 713 | 174,846.07 | 4.90 | √ | √ | √ | X | X | na | na | PP763331 |
| 12 | hap2.Chr03.1907 | 3 | 2 | ETTN-like | 2163 | 720 | 176,745.27 | 4.90 | √ | √ | √ | X | X | na | na | PP763357 |
| 24 | hap1.Chr06.300 | 6 | 1 | ETTN-like | 2127 | 708 | 175,645.46 | 4.88 | √ | √ | √ | X | X | na | na | PP763330 |
| 24 | hap2.Chr06.302 | 6 | 2 | ETTN-like | 2223 | 740 | 183,107.48 | 4.88 | √ | √ | √ | X | X | na | na | PP763356 |
| 11 | hap1.Chr03.2920 | 3 | 1 | ETTN-like | 2058 | 685 | 168,347.28 | 4.93 | √ | √ | √ | X | X | na | na | PP763334 |
| 11 | hap2.Chr03.1440 | 3 | 2 | ETTN-like | 2049 | 682 | 167,593.36 | 4.93 | √ | √ | √ | X | X | na | na | PP763361 |
| 23 | hap2.Chr06.618 | 6 | 1 | ETTN-like | 2052 | 683 | 167,136.26 | 4.92 | √ | √ | √ | X | X | na | na | PP763360 |
| 23 | hap1.Chr06.605 | 6 | 2 | ETTN-like | 2052 | 683 | 167,182.35 | 4.92 | √ | √ | √ | X | X | na | na | PP763333 |
| 2 | hap1.Chr01.3255 | 1 | 1 | unknown | 2073 | 690 | 169,779.02 | 4.88 | √ | √ | √ | X | √ | √ | √ | PP763311 |
| 2 | hap2.Chr01.2164 | 1 | 2 | unknown | 2073 | 690 | 169,524.45 | 4.88 | √ | √ | √ | X | √ | √ | √ | PP763337 |
| 31a | hap1.Chr08.2898 | 8 | 1 | unknown | 1572 | 523 | 133,420.31 | 4.86 | √ | √ | √ | X | √ | X | X | PP763335 |
| 31a | hap2.Chr08.2889 | 8 | 2 | unknown | 1566 | 521 | 132,723.33 | 4.87 | √ | √ | √ | X | √ | X | X | PP763362 |
| 31b | hap2.Chr01.3181 | 1 | 2 | unknown | 558 | 185 | 45,378.47 | 5.20 | √ | √ | P | X | √ | X | X | PP763363 |
| 6 | hap1.Chr08.1599 | 8 | 1 | unknown | 1830 | 609 | 152,317.09 | 4.84 | √ | √ | √ | X | √ | P | X | PP763327 |
| 6 | hap2.Chr08.1582 | 8 | 2 | unknown | 1764 | 587 | 146,798.63 | 4.85 | √ | √ | √ | X | √ | P | X | PP763354 |
| 15 | hap1.Chr05.2176 | 5 | 1 | unknown | 2121 | 706 | 175,252.12 | 4.81 | √ | √ | √ | X | √ | √ | √ | PP763320 |
| 15 | hap2.Chr05.2157 | 5 | 2 | unknown | 2121 | 706 | 175,238.09 | 4.81 | √ | √ | √ | X | √ | √ | √ | PP763346 |
| 21 | hap1.Chr10.490 | 10 | 1 | unknown | 2118 | 705 | 173,757.45 | 4.84 | √ | √ | √ | X | √ | √ | √ | PP763315 |
| 21 | hap2.Chr10.500 | 10 | 2 | unknown | 2154 | 717 | 176,664.72 | 4.83 | √ | √ | √ | X | √ | √ | √ | PP763340 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Wang, G.; Zeng, J.; Du, C.; Tang, Q.; Hua, Y.; Chen, M.; Yang, G.; Tu, M.; He, G.; Li, Y.; et al. Divergent Roles of the Auxin Response Factors in Lemongrass (Cymbopogon flexuosus (Nees ex Steud.) W. Watson) during Plant Growth. Int. J. Mol. Sci. 2024, 25, 8154. https://doi.org/10.3390/ijms25158154
Wang G, Zeng J, Du C, Tang Q, Hua Y, Chen M, Yang G, Tu M, He G, Li Y, et al. Divergent Roles of the Auxin Response Factors in Lemongrass (Cymbopogon flexuosus (Nees ex Steud.) W. Watson) during Plant Growth. International Journal of Molecular Sciences. 2024; 25(15):8154. https://doi.org/10.3390/ijms25158154
Chicago/Turabian StyleWang, Guoli, Jian Zeng, Canghao Du, Qi Tang, Yuqing Hua, Mingjie Chen, Guangxiao Yang, Min Tu, Guangyuan He, Yin Li, and et al. 2024. "Divergent Roles of the Auxin Response Factors in Lemongrass (Cymbopogon flexuosus (Nees ex Steud.) W. Watson) during Plant Growth" International Journal of Molecular Sciences 25, no. 15: 8154. https://doi.org/10.3390/ijms25158154