
Optimization of the Method for Isolating Bacterial DNA from the Aboveground Part of Lettuce

Abstract
1. Introduction
2. Results and Discussion
3. Materials and Methods
3.1. Lettuce Cultivation Conditions
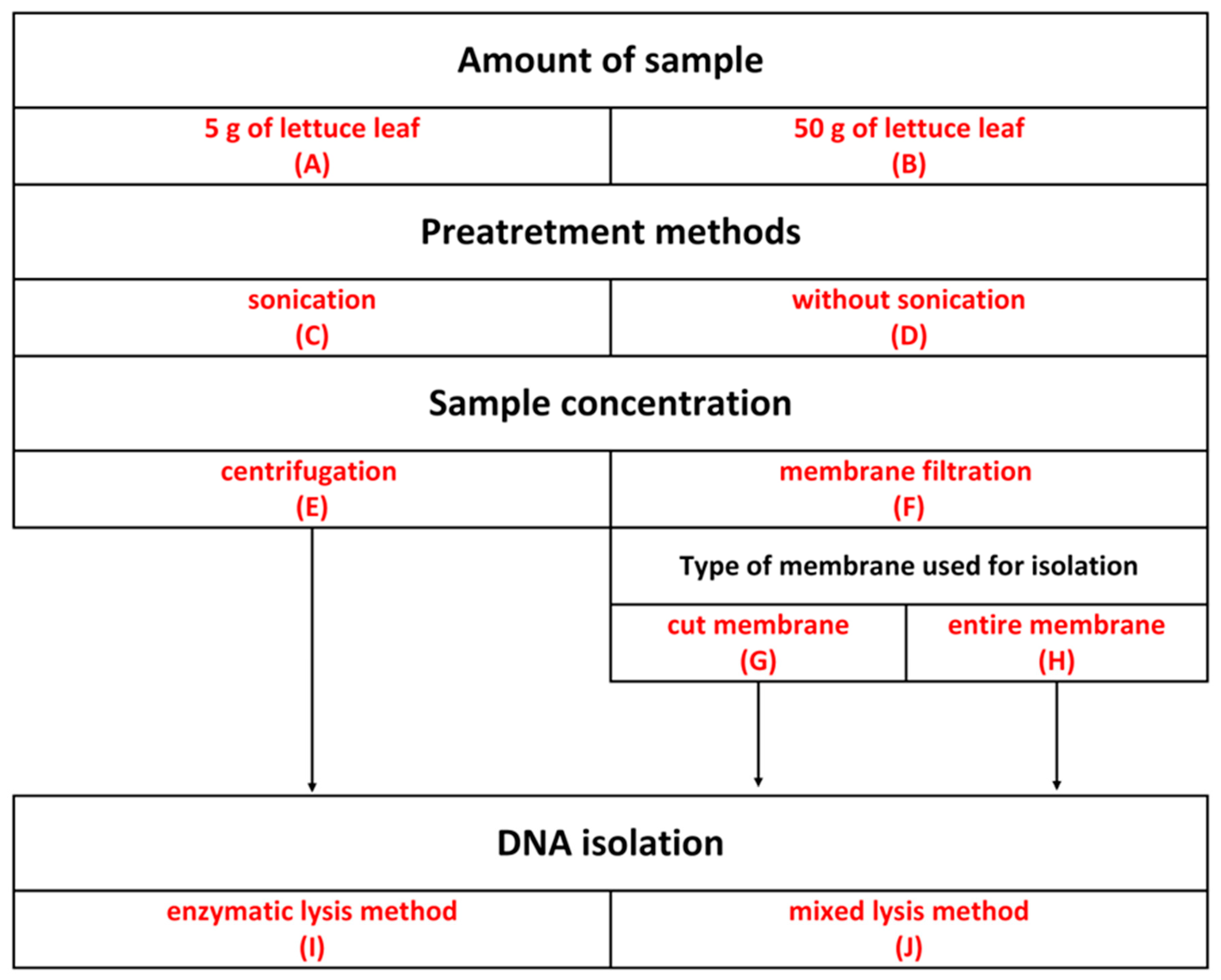
3.2. Bacterial Cell Separation from Lettuce Leaves
Sample Preparation
3.3. DNA Isolation
3.4. Experimental Variants, Quantity, and Quality of Isolated DNA
3.5. Next-Generation Sequencing
3.6. Statistical Analyses
4. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- FAO; UNEP; WHO; WOAH. Global Plan of Action on One Health. Towards a more comprehensive One Health, Approach to Global Health Threats at the Human-Animal-Environment Interface: One Health Joint Plan of Action, 2022–2026; WHO: Rome, Italy, 2022; ISBN 978-92-5-136957-9. [Google Scholar] [CrossRef]
- Ma, L.; Zhao, H.; Wu, L.B.; Cheng, Z.; Liu, C. Impact of the microbiome on human, animal, and environmental health from a One Health perspective. Sci. One Health 2023, 2, 100037. [Google Scholar] [CrossRef] [PubMed]
- Wang, F.; Fu, Y.H.; Sheng, H.J.; Topp, E.; Jiang, X.; Zhu, Y.G.; Tiedje, J.M. Antibiotic resistance in the soil ecosystem: A One Health perspective. Curr. Opin. Environ. Sci. Health. 2021, 20, 100230. [Google Scholar] [CrossRef]
- Cerqueira, F.; Matamoros, V.; Bayona, J.M.; Berendonk, T.U.; Elsinga, G.; Hornstra, L.M.; Piña, B. Antibiotic resistance gene distribution in agricultural fields and crops. A soil-to-food analysis. Environ. Res. 2019, 177, 108608. [Google Scholar] [CrossRef]
- Kläui, A.; Bütikofer, U.; Naskova, J.; Wagner, E.; Marti, E. Fresh produce as a reservoir of antimicrobial resistance genes: A case study of Switzerland. Sci. Total Environ. 2024, 907, 167671. [Google Scholar] [CrossRef] [PubMed]
- Wu, S.; Huang, J.; Wu, Q.; Zhang, J.; Zhang, F.; Yang, X.; Wu, H.; Zeng, H.; Chen, M.; Ding, Y. Staphylococcus aureus isolated from retail meat and meat products in china: Incidence, antibiotic resistance and genetic diversity. Front. Microbiol. 2018, 9, 2767. [Google Scholar] [CrossRef]
- Tóth, A.G.; Csabai, I.; Krikó, E.; Tőzsér, D.; Maróti, G.; Patai, Á.V.; Makrai, L.; Szita, G.; Solymosi, N. Antimicrobial resistance genes in raw milk for human consumption. Sci. Rep. 2020, 10, 7464. [Google Scholar] [CrossRef] [PubMed]
- Hu, L.; Cao, G.; Brown, E.W.; Allard, M.W.; Ma, L.M.; Khan, A.A.; Zhang, G. Antimicrobial resistance and related gene analysis of Salmonella from egg and chicken sources by whole-genome sequencing. Poult. Sci. 2020, 99, 7076–7083. [Google Scholar] [CrossRef] [PubMed]
- Sun, H.; Li, H.; Zhang, X.; Liu, Y.; Chen, H.; Zheng, L.; Zhai, Y.; Zheng, H. The honeybee gut resistome and its role in antibiotic resistance dissemination. Integr. Zool. 2023, 18, 1014–1026. [Google Scholar] [CrossRef]
- Qamar, M.U.; Aatika; Chughtai, M.I.; Ejaz, H.; Mazhari, B.B.Z.; Maqbool, U.; Alanazi, A.; Alruwaili, Y.; Junaid, K. Antibiotic-resistant bacteria, antimicrobial resistance genes, and antibiotic residue in food from animal sources: One Health Food Safety Concern. Microorganisms 2023, 11, 161. [Google Scholar] [CrossRef] [PubMed]
- Yuan, X.; Lv, Z.; Zhang, Z.; Han, Y.; Liu, Z.; Zhang, H. A review of antibiotics, antibiotic resistant bacteria, and resistance genes in aquaculture: Occurrence, contamination, and transmission. Toxics 2023, 11, 420. [Google Scholar] [CrossRef]
- Berg, G.; Erlacher, A.; Grube, M. The edible plant microbiome: Importance and health issues. In Principles of Plant-Microbe Interactions; Lugtenberg, B., Ed.; Springer International Publishing: Cham, Switerland, 2015; pp. 419–426. ISBN 978-3-319-08574-6. [Google Scholar]
- Soto-Giron, M.J.; Kim, J.N.; Schott, E.; Tahmin, C.; Ishoey, T.; Mincer, T.J.; DeWalt, J.; Toledo, G. The edible plant microbiome represents a diverse genetic reservoir with functional potential in the human host. Sci. Rep. 2021, 11, 24017. [Google Scholar] [CrossRef] [PubMed]
- Allard, S.M.; Walsh, C.S.; Wallis, A.E.; Ottesen, A.R.; Brown, E.W.; Micallef, S.A. Solanum lycopersicum (tomato) hosts robust phyllosphere and rhizosphere bacterial communities when grown in soil amended with various organic and synthetic fertilizers. Sci. Total Environ. 2016, 573, 555–563. [Google Scholar] [CrossRef] [PubMed]
- Wicaksono, W.A.; Cernava, T.; Wassermann, B.; Abdelfattah, A.; Soto-Giron, M.J.; Toledo, G.V.; Virtanen, S.M.; Knip, M.; Hyöty, H.; Berg, G. The edible plant microbiome: Evidence for the occurrence of fruit and vegetable bacteria in the human gut. Gut Microbes 2023, 15, 2258565. [Google Scholar] [CrossRef] [PubMed]
- Rastogi, G.; Sbodio, A.; Tech, J.J.; Suslow, T.V.; Coaker, G.L.; Leveau, J.H.J. Leaf microbiota in an agroecosystem: Spatiotemporal variation in bacterial community composition on field-grown lettuce. ISME J. 2012, 6, 1812–1822. [Google Scholar] [CrossRef] [PubMed]
- Gupta, R.; Anand, G.; Gaur, R.; Yadav, D. Plant-microbiome interactions for sustainable agriculture: A review. Physiol. Mol. Biol. Plants 2021, 27, 165–179. [Google Scholar] [CrossRef] [PubMed]
- Lucaciu, R.; Pelikan, C.; Gerner, S.M.; Zioutis, C.; Köstlbacher, S.; Marx, H.; Herbold, C.W.; Schmidt, H.; Rattei, T. A Bioinformatics guide to plant microbiome analysis. Front. Plant Sci. 2019, 10, 1313. [Google Scholar] [CrossRef] [PubMed]
- Fricker, A.M.; Podlesny, D.; Fricke, W.F. What is new and relevant for sequencing-based microbiome research? A mini-review. J. Adv. Res. 2019, 19, 105–112. [Google Scholar] [CrossRef]
- Zhang, J.; Cook, J.; Nearing, J.T.; Zhang, J.; Raudonis, R.; Glick, B.R.; Langille, M.G.I.; Cheng, Z. Harnessing the plant microbiome to promote the growth of agricultural crops. Microbiol. Res. 2021, 245, 126690. [Google Scholar] [CrossRef] [PubMed]
- Yang, S.; Lee, J.; Mahmood, M.; Min, D.; Im, J. Evaluation and optimization of lysis method for microbial DNA extraction from epiphytic phyllosphere samples. J. Microbiol. Methods 2023, 206, 106677. [Google Scholar] [CrossRef] [PubMed]
- Chen, Q.-L.; Fan, X.T.; Zhu, D.; An, X.L.; Su, J.Q.; Cui, L. Effect of biochar amendment on the alleviation of antibiotic resistance in soil and phyllosphere of Brassica chinensis L. Soil. Biol. Biochem. 2018, 119, 74–82. [Google Scholar] [CrossRef]
- Chen, X.; Wicaksono, W.A.; Berg, G.; Cernava, T. Bacterial communities in the plant phyllosphere harbour distinct responders to a broad-spectrum pesticide. Sci. Total Environ. 2021, 751, 141799. [Google Scholar] [CrossRef] [PubMed]
- Zhang, Y.J.; Hu, H.W.; Chen, Q.L.; Singh, B.K.; Yan, H.; Chen, D.; He, J.Z. Transfer of antibiotic resistance from manure-amended soils to vegetable microbiomes. Environ. Int. 2019, 130, 104912. [Google Scholar] [CrossRef] [PubMed]
- Liu, J.; Zhang, W.; Liu, Y.; Zhu, W.; Yuan, Z.; Su, X.; Ding, C. Differences in phyllosphere microbiomes among different Populus spp. in the same habitat. Front. Plant Sci. 2023, 14, 1143878. [Google Scholar] [CrossRef]
- Klumpp, J.; Fouts, D.E.; Sozhamannan, S. Next generation sequencing technologies and the changing landscape of phage genomics. Bacteriophage 2012, 2, 190–199. [Google Scholar] [CrossRef] [PubMed]
- Hamilton, A.; Harper, S.J.; Critzer, F. Optimization of a method for the concentration of genetic material in bacterial and fungal communities on fresh apple peel surfaces. Microorganisms 2020, 8, 1490. [Google Scholar] [CrossRef]
- Videnska, P.; Smerkova, K.; Zwinsova, B.; Popovici, V.; Micenkova, L.; Sedlar, K.; Budinska, E. Stool sampling and DNA isolation kits affect DNA quality and bacterial composition following 16S rRNA gene sequencing using MiSeq Illumina platform. Sci. Rep. 2019, 9, 13837. [Google Scholar] [CrossRef]
- Lear, G.; Dickie, I.; Banks, J.; Boyer, S.; Buckley, H.; Buckley, T.; Cruickshank, R.; Dopheide, A.; Handley, K.; Hermans, S. Methods for the extraction, storage, amplification and sequencing of DNA from environmental samples. N. Z. J. Ecol. 2018, 42, 10. [Google Scholar] [CrossRef]
- Baturo-Cieśniewska, A.; Loddi, G.; Prusiński, J.; Łukanowski, A. Effect of extraction method and dna quality on the reliability of molecular detection of Bradyrhizobium japonicum in soybean rhizosphere. Electron. J. Pol. Agric. Univ. Ser. Agron. 2019, 22, 5. [Google Scholar] [CrossRef]
- Mazziotti, M.; Henry, S.; Laval-Gilly, P.; Bonnefoy, A.; Falla, J. Comparison of two bacterial DNA extraction methods from non-polluted and polluted soils. Folia Microbiol. 2018, 63, 85–92. [Google Scholar] [CrossRef]
- Lee, J.; Kim, H.-S.; Jo, H.Y.; Kwon, M.J. Revisiting soil bacterial counting methods: Optimal soil storage and pretreatment methods and comparison of culture-dependent and -independent methods. PLoS ONE 2021, 16, e0246142. [Google Scholar] [CrossRef]
- Sare, A.R.; Stouvenakers, G.; Eck, M.; Lampens, A.; Goormachtig, S.; Jijakli, M.H.; Massart, S. Standardization of plant microbiome studies: Which proportion of the microbiota is really harvested? Microorganisms 2020, 8, 342. [Google Scholar] [CrossRef] [PubMed]
- Yin, Y.; Wang, Y.F.; Cui, H.L.; Zhou, R.; Li, L.; Duan, G.L.; Zhu, Y.G. Distinctive structure and assembly of phyllosphere microbial communities between wild and cultivated rice. Microbiol. Spectr. 2023, 11, e0437122. [Google Scholar] [CrossRef] [PubMed]
- Trivedi, P.; Leach, J.E.; Tringe, S.G.; Sa, T.; Singh, B.K. Plant-microbiome interactions: From community assembly to plant health. Nat. Rev. Microbiol. 2020, 18, 607–621. [Google Scholar] [CrossRef] [PubMed]
- Aliche, E.B.; Talsma, W.; Munnik, T.; Bouwmeester, H.J. Characterization of maize root microbiome in two different soils by minimizing plant DNA contamination in metabarcoding analysis. Biol. Fertil. Soils 2021, 57, 731–737. [Google Scholar] [CrossRef]
- Krupka, M.; Olkowska, E.; Klimkowicz-Pawlas, A.; Łęczyński, L.; Tankiewicz, M.; Michalczyk, D.J.; Wolska, L.; Piotrowicz-Cieślak, A.I. The Impact of soil and water pollutants released from poultry farming on the growth and development of two plant dpecies. Agriculture 2024, 14, 87. [Google Scholar] [CrossRef]
- Zhou, S.Y.D.; Zhu, D.; Giles, M.; Yang, X.R.; Daniell, T.; Neilson, R.; Zhu, Y.G. Phyllosphere of staple crops under pig manure fertilization, a reservoir of antibiotic resistance genes. Environ. Pollut. 2019, 252, 227–235. [Google Scholar] [CrossRef]
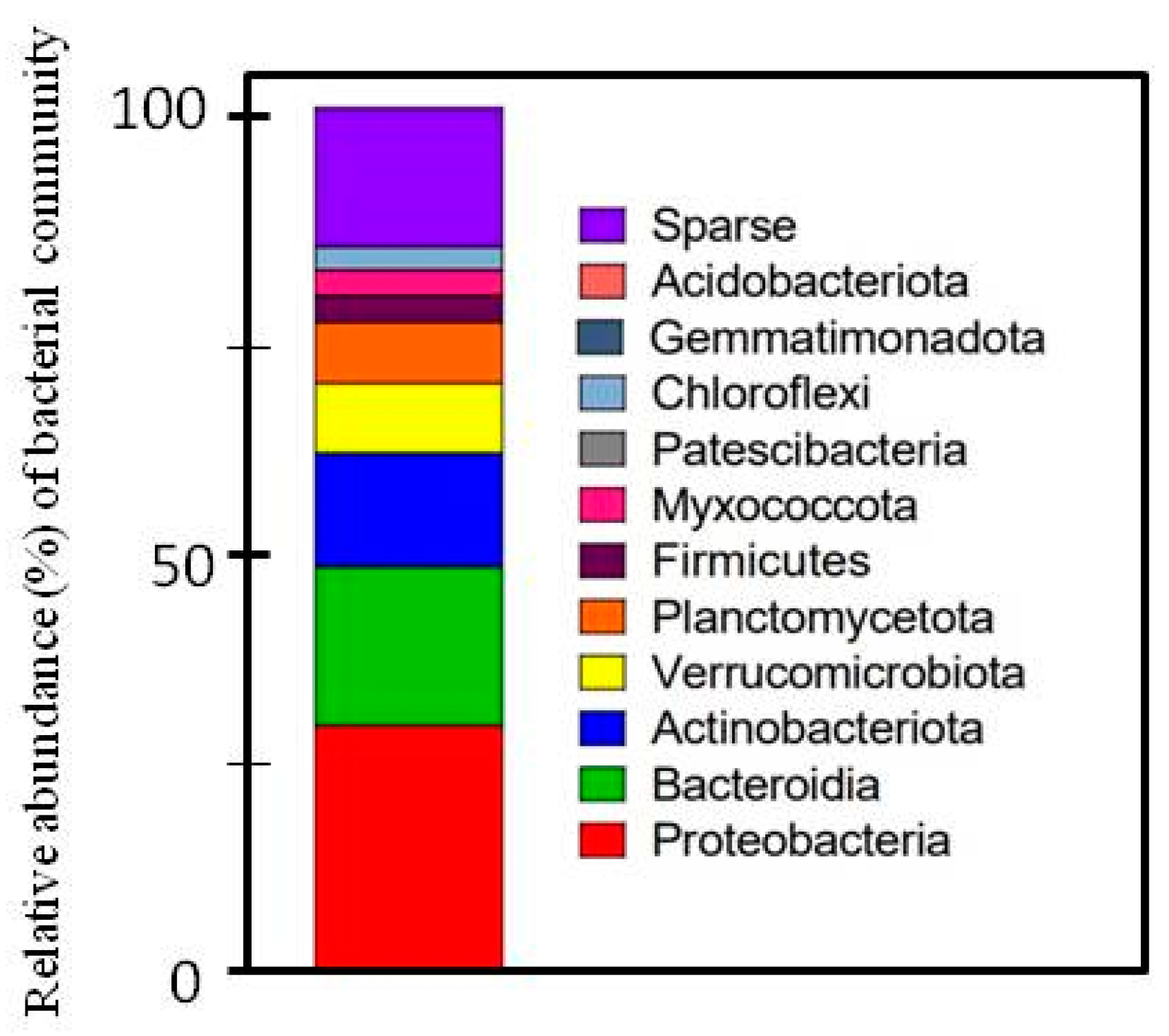

{kind=link}
{kind=link}
| Number of Experiment | DNA [ng/µL] | DNA [ng] | A260/280 | A260/230 |
|---|---|---|---|---|
| Enzymatic lysis | ||||
| 1 | 3.94 ± 0.70 a | 194 ± 39.4 a | 1.40 c | 0.77 b |
| 2 | 6.41 ± 0.95 b | 327 ± 45.0 b | 1.50c | 1.10d |
| 3 | 7.45 ± 0.95 b | 378 ± 47.0 b | 1.48 c | 0.97 c |
| 4 | 3.14 ± 0.69 a | 160 ± 35.8 a | 1.20 a | 0.75 b |
| 5 | 4.86 ± 1.02 b | 241± 53.9 b | 1.34 b | 1.00 c |
| 6 | 4.19 ± 1.03 a | 206 ± 53.7 a | 1.50 b | 1.11 d |
| 7 | 5.56 ± 1.19 b | 271 ± 59.0 b | 1.30 b | 0.70 b |
| 8 | 10.30 ± 1.38 c | 521 ± 70.5 c | 1.60 d | 1.40 e |
| 9 | 11.50 ± 1.00 c | 575 ± 54.7 c | 1.53 c | 1.13 d |
| 10 | 5.57 ± 0.70 b | 281 ± 36.4 b | 1.30 b | 0.57 a |
| 11 | 6.90 ± 0.65 b | 348 ± 32.9 b | 1.40 b | 0.73 b |
| 12 | 6.99 ± 1.16 b | 346 ± 60.5 b | 1.54 c | 0.77 b |
| Mechanical–chemical lysis | ||||
| 13 | 6.07 ± 0.81 b | 306 ± 42.0 b | 1.60 d | 1.12 d |
| 14 | 9.50 ± 1.35 c | 470 ± 69.8 c | 1.75 f | 1.79 g |
| 15 | 9.98 ± 1.20 c | 490 ± 57.4 c | 1.90 g | 1.50 f |
| 16 | 5.48 ± 0.69 b | 276± 35.8 b | 1.68 f | 1.45 f |
| 17 | 6.81 ± 0.84 b | 345 ± 42.5 b | 1.85 g | 1.78 g |
| 18 | 6.84 ± 0.75 b | 342 ±39.6 b | 1.90 g | 1.62 g |
| 19 | 5.67 ± 0.84 b | 280 ± 43.2 b | 1.78 f | 1.56 f |
| 20 | 24.29 ± 4.32 d | 1205 ± 226.6 d | 1.83 g | 1.56 f |
| 21 | 38.08 ± 4.28 f | 1929 ± 210.4 f | 1.85 g | 1.81 g |
| 22 | 5.84 ± 0.93 b | 301 ± 37.7 b | 1.76 f | 1.74 g |
| 23 | 8.41 ± 1.39 c | 420 ± 73.9 c | 1.84 g | 0.91 c |
| 24 | 9.20 ± 0.93 c | 461 ± 49.6 c | 1.90 h | 1.87 h |
| Number of Experiment | Experimental Steps |
|---|---|
| 1 | 5 g of lettuce leaf, sonication, centrifugation, enzymatic lysis |
| 2 | 5 g of lettuce leaf, sonication, membrane filtration, entire membrane, enzymatic lysis |
| 3 | 5 g of lettuce leaf, sonication, membrane filtration, cut membrane, enzymatic lysis |
| 4 | 5 g of lettuce leaf, without sonication, centrifugation, enzymatic lysis |
| 5 | 5 g of lettuce leaf, without sonication, membrane filtration, entire membrane, enzymatic lysis |
| 6 | 5 g of lettuce leaf, without sonication, membrane filtration, cut membrane, enzymatic lysis |
| 7 | 50 g of lettuce leaf, sonication, centrifugation, enzymatic lysis |
| 8 | 50 g of lettuce leaf, sonication, membrane filtration, entire membrane, enzymatic lysis |
| 9 | 50 g of lettuce leaf, sonication, membrane filtration, cut membrane, enzymatic lysis |
| 10 | 50 g of lettuce leaf, without sonication, centrifugation, enzymatic lysis |
| 11 | 50 g of lettuce leaf, without sonication, membrane filtration, entire membrane, enzymatic lysis |
| 12 | 50 g of lettuce leaf, without sonication, membrane filtration, cut membrane, enzymatic lysis |
| 13 | 5 g of lettuce leaf, sonication, centrifugation, mixed lysis |
| 14 | 5 g of lettuce leaf, sonication, membrane filtration, entire membrane, mixed lysis |
| 15 | 5 g of lettuce leaf, sonication, membrane filtration, cut membrane, mixed lysis |
| 16 | 5 g of lettuce leaf, without sonication, centrifugation, mixed lysis |
| 17 | 5 g of lettuce leaf, without sonication, membrane filtration, entire membrane, mixed lysis |
| 18 | 5 g of lettuce leaf, without sonication, membrane filtration, cut membrane, mixed lysis |
| 19 | 50 g of lettuce leaf, sonication, centrifugation, mixed lysis |
| 20 | 50 g of lettuce leaf, sonication, membrane filtration, entire membrane, mixed lysis |
| 21 | 50 g of lettuce leaf, sonication, membrane filtration, cut membrane, mixed lysis |
| 22 | 50 g of lettuce leaf, without sonication, centrifugation, mixed lysis |
| 23 | 50 g of lettuce leaf, without sonication, membrane filtration, entire membrane, mixed lysis |
| 24 | 50 g of lettuce leaf, without sonication, membrane filtration, cut membrane, mixed lysis |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Krupka, M.; Piotrowicz-Cieślak, A.I. Optimization of the Method for Isolating Bacterial DNA from the Aboveground Part of Lettuce. Int. J. Mol. Sci. 2024, 25, 8513. https://doi.org/10.3390/ijms25158513
Krupka M, Piotrowicz-Cieślak AI. Optimization of the Method for Isolating Bacterial DNA from the Aboveground Part of Lettuce. International Journal of Molecular Sciences. 2024; 25(15):8513. https://doi.org/10.3390/ijms25158513
Chicago/Turabian StyleKrupka, Magdalena, and Agnieszka I. Piotrowicz-Cieślak. 2024. "Optimization of the Method for Isolating Bacterial DNA from the Aboveground Part of Lettuce" International Journal of Molecular Sciences 25, no. 15: 8513. https://doi.org/10.3390/ijms25158513
APA StyleKrupka, M., & Piotrowicz-Cieślak, A. I. (2024). Optimization of the Method for Isolating Bacterial DNA from the Aboveground Part of Lettuce. International Journal of Molecular Sciences, 25(15), 8513. https://doi.org/10.3390/ijms25158513