
The Association between IL-1β and IL-18 Levels, Gut Barrier Disruption, and Monocyte Activation during Chronic Simian Immunodeficiency Virus Infection and Long-Term Suppressive Antiretroviral Therapy
 and
and {kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Results
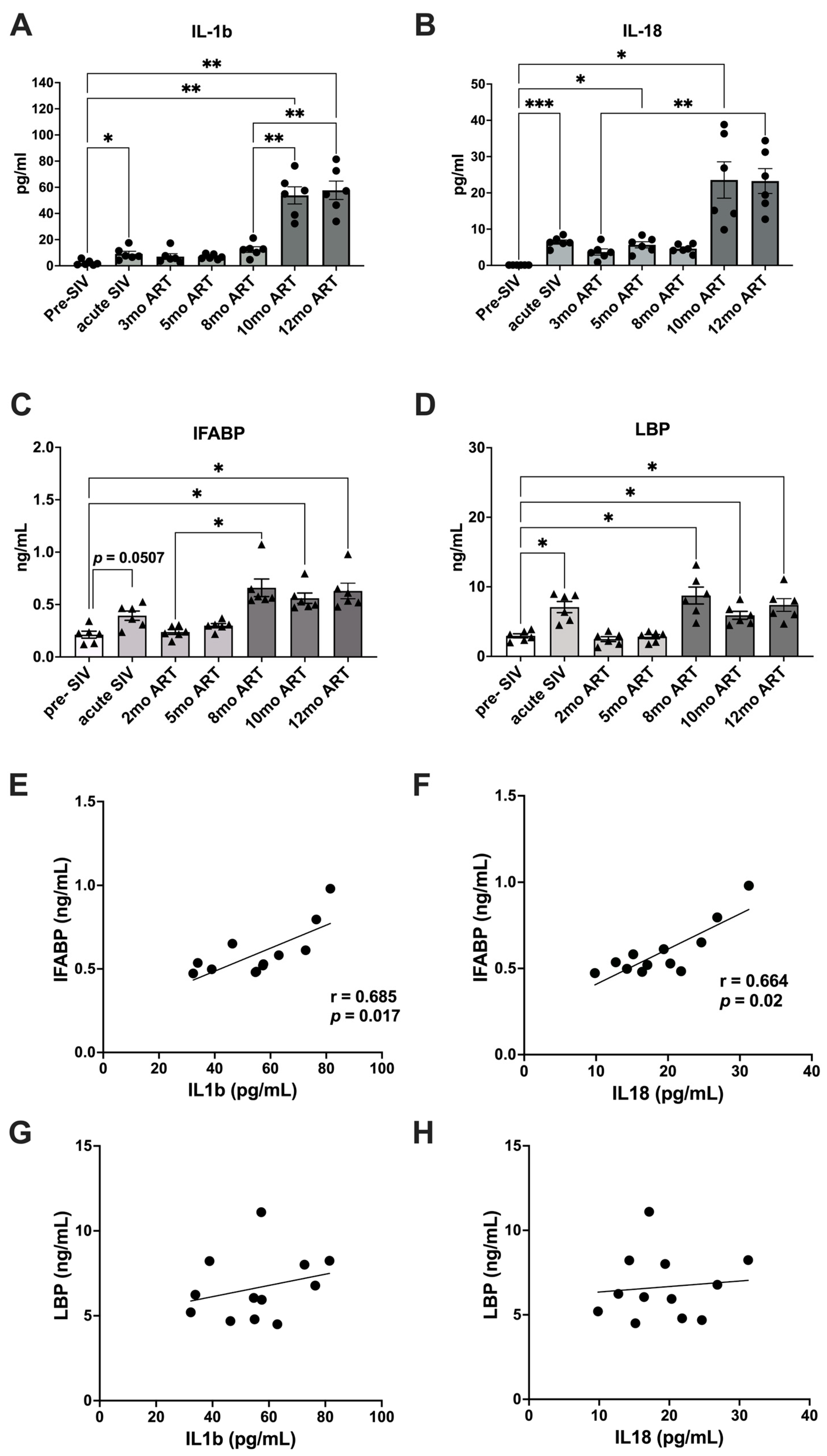
2.1. Increased Gut Permeability during Chronic SIV with ART Aligns with Elevated Levels of IL-1β and IL-18
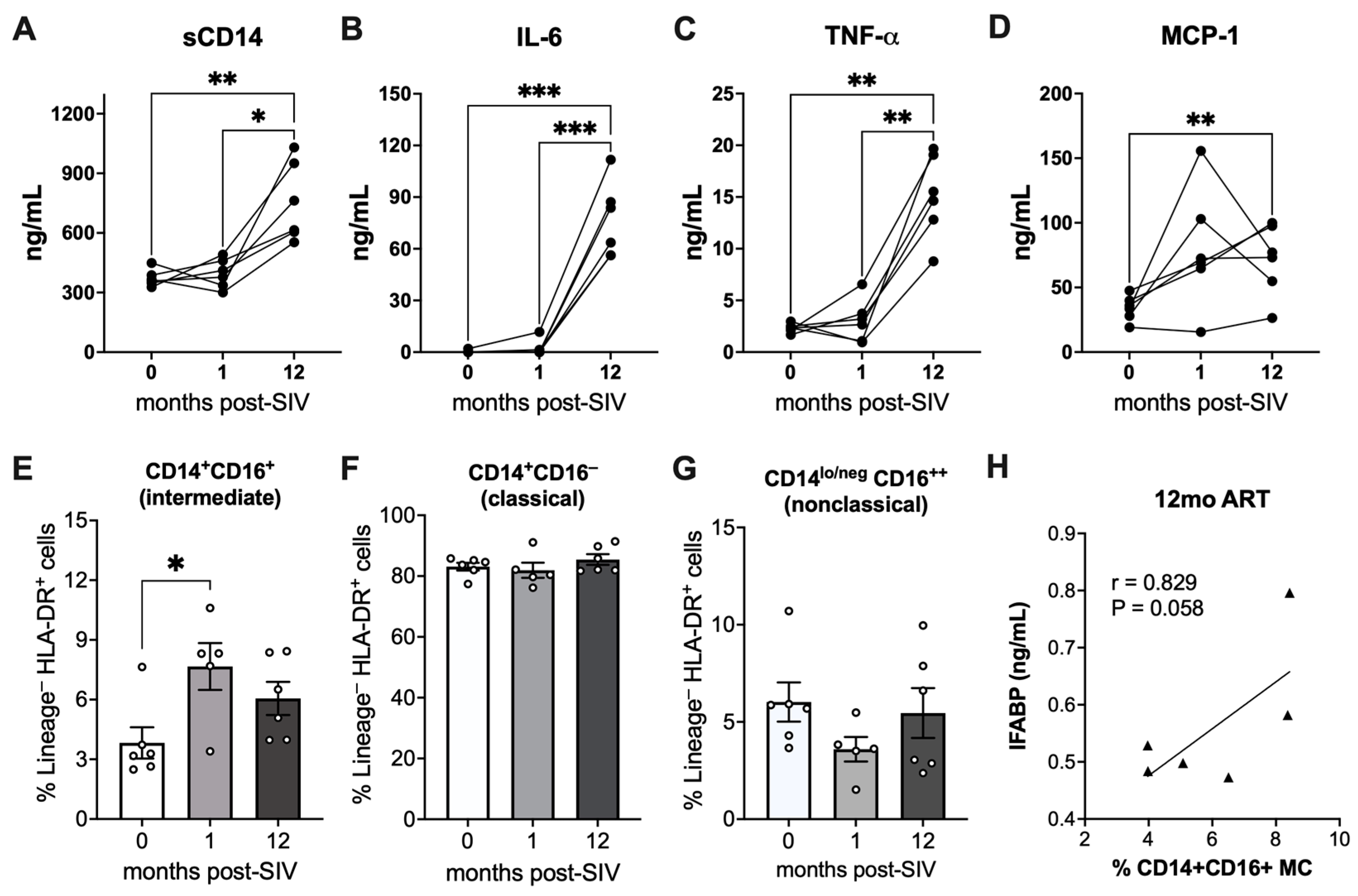
2.2. Elevated Markers of Monocyte Activation and Correlation of Intermediate Monocytes with IEBD during Chronic SIV Infection with Long-Term ART
2.3. Blood Triglycerides and Glycolytic Capacity in PBMCs during Chronic SIV Infection with Long-Term ART
3. Discussion
4. Materials and Methods
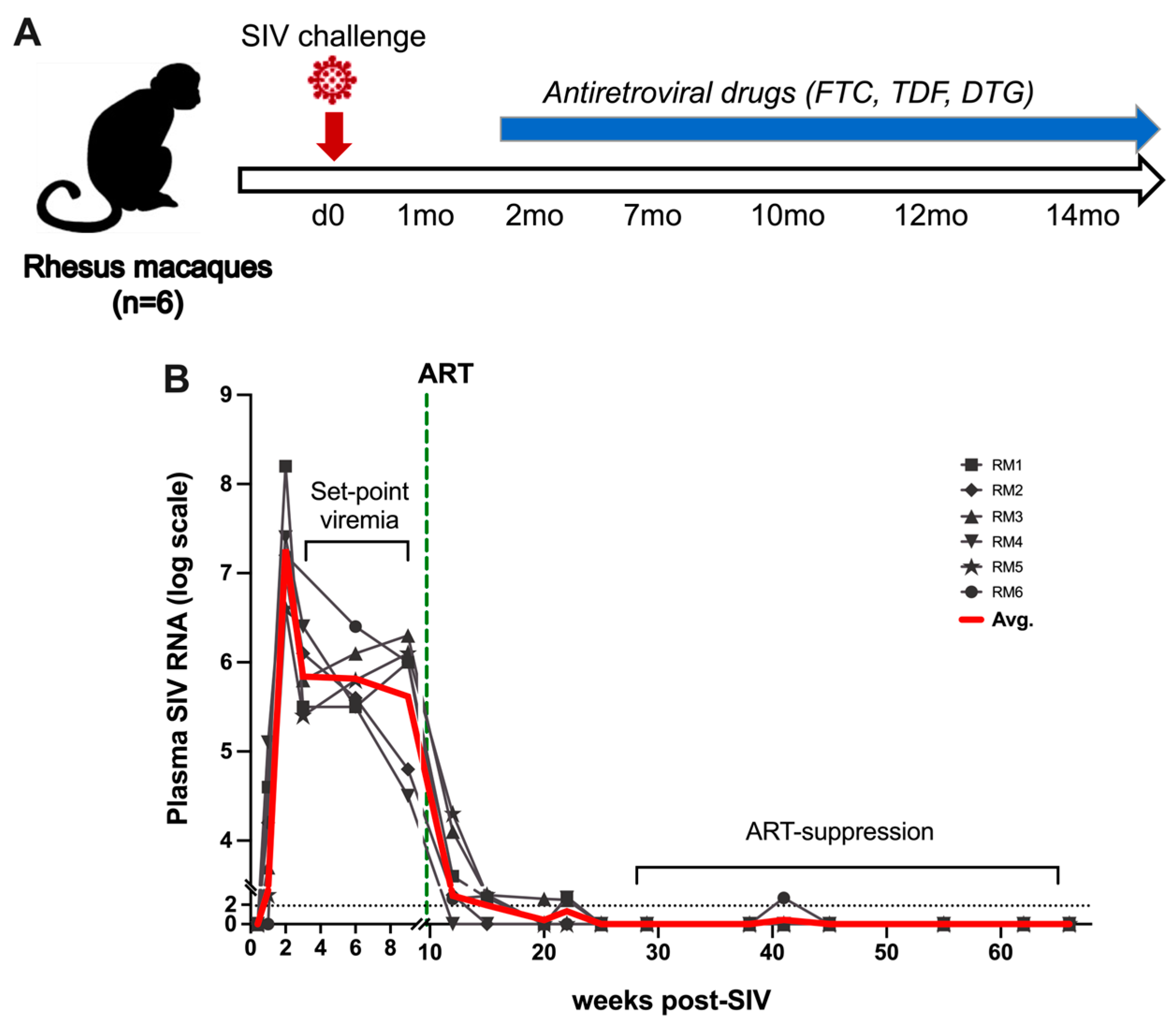
4.1. Animals, Viral Inoculation, and ART
4.2. Isolation of Peripheral Blood Mononuclear Cells (PBMCs) and Flow Cytometry
4.3. Plasma Markers of Inflammation, Microbial Translocation, and Intestinal Damage
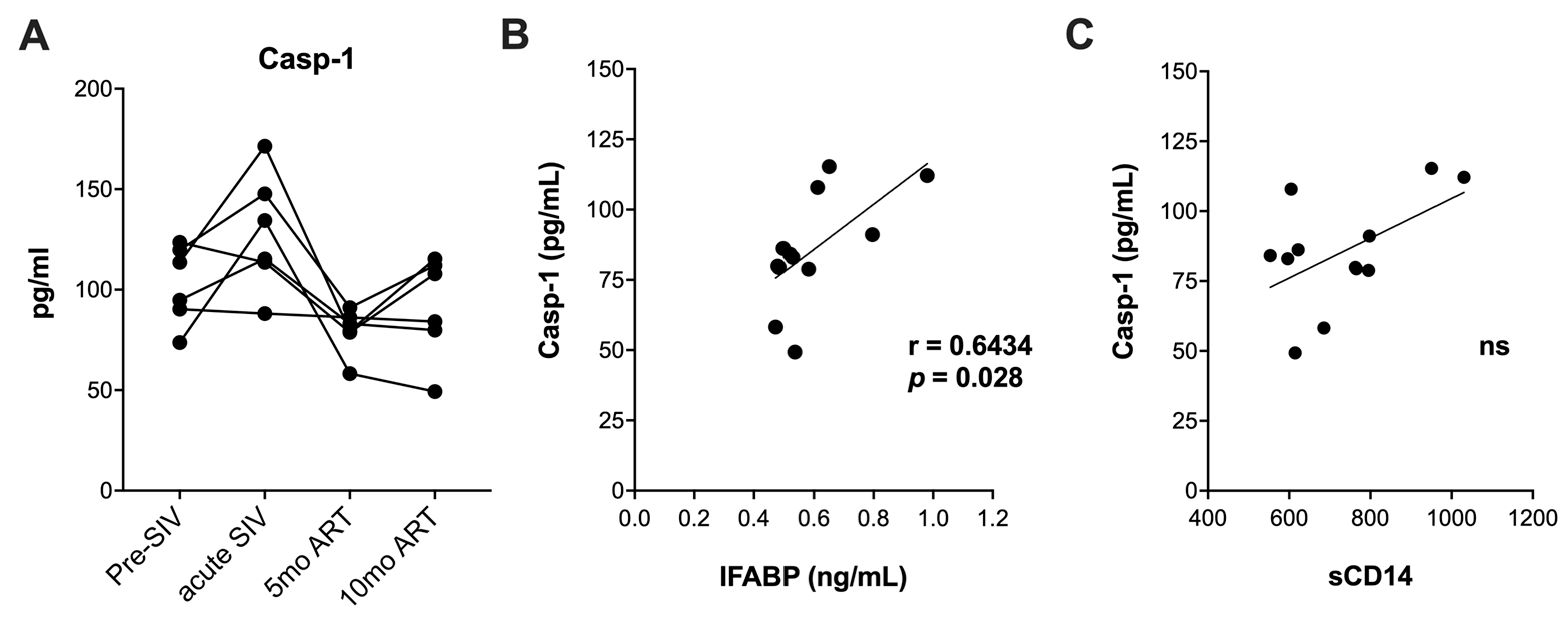
4.4. Measurement of Caspase-1 in Serum
4.5. Measurement of Glycolytic Rate and Extracellular Acidification Rates (ECARs)
4.6. Statistical Analyses
Author Contributions
Funding
Institutional Review Board Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Katz, I.T.; Maughan-Brown, B. Improved life expectancy of people living with HIV: Who is left behind? Lancet HIV 2017, 4, e324–e326. [Google Scholar] [CrossRef] [PubMed]
- Liu, F.; Dai, S.; Gordon, J.; Qin, X. Complement and HIV-I infection/HIV-associated neurocognitive disorders. J. NeuroVirology 2014, 20, 184–198. [Google Scholar] [CrossRef] [PubMed]
- Kearns, A.; Gordon, J.; Burdo, T.H.; Qin, X. HIV-1-Associated Atherosclerosis: Unraveling the Missing Link. J. Am. Coll. Cardiol. 2017, 69, 3084–3098. [Google Scholar] [CrossRef] [PubMed]
- Kearns, A.; Burdo, T.H.; Qin, X. Editorial Commentary: Clinical management of cardiovascular disease in HIV-infected patients. Trends Cardiovasc. Med. 2017, 27, 564–566. [Google Scholar] [CrossRef] [PubMed]
- Caetano, D.G.; Ribeiro-Alves, M.; Hottz, E.D.; Vilela, L.M.; Cardoso, S.W.; Hoagland, B.; Grinsztejn, B.; Veloso, V.G.; Morgado, M.G.; Bozza, P.T.; et al. Increased biomarkers of cardiovascular risk in HIV-1 viremic controllers and low persistent inflammation in elite controllers and art-suppressed individuals. Sci. Rep. 2022, 12, 6569. [Google Scholar] [CrossRef] [PubMed]
- Libby, P.; Ridker, P.M.; Hansson, G.K. Progress and challenges in translating the biology of atherosclerosis. Nature 2011, 473, 317–325. [Google Scholar] [CrossRef]
- Kuller, L.H.; Tracy, R.; Belloso, W.; De Wit, S.; Drummond, F.; Lane, H.C.; Ledergerber, B.; Lundgren, J.; Neuhaus, J.; Nixon, D.; et al. Inflammatory and coagulation biomarkers and mortality in patients with HIV infection. PLoS Med. 2008, 5, e203. [Google Scholar] [CrossRef] [PubMed]
- Borges, A.H.; O’Connor, J.L.; Phillips, A.N.; Neaton, J.D.; Grund, B.; Neuhaus, J.; Vjecha, M.J.; Calmy, A.; Koelsch, K.K.; Lundgren, J.D.; et al. Interleukin 6 Is a Stronger Predictor of Clinical Events Than High-Sensitivity C-Reactive Protein or D-Dimer During HIV Infection. J. Infect. Dis. 2016, 214, 408–416. [Google Scholar] [CrossRef] [PubMed]
- Sim, J.H.; Mukerji, S.S.; Russo, S.C.; Lo, J. Gastrointestinal Dysfunction and HIV Comorbidities. Curr. HIV/AIDS Rep. 2021, 18, 57–62. [Google Scholar] [CrossRef]
- Walker, E.M.; Slisarenko, N.; Gerrets, G.L.; Grasperge, B.F.; Mattison, J.A.; Kissinger, P.J.; Welsh, D.A.; Veazey, R.S.; Jazwinski, S.M.; Rout, N. Dysregulation of IL-17/IL-22 Effector Functions in Blood and Gut Mucosal Gamma Delta T Cells Correlates With Increase in Circulating Leaky Gut and Inflammatory Markers During cART-Treated Chronic SIV Infection in Macaques. Front. Immunol. 2021, 12, 647398. [Google Scholar] [CrossRef]
- Moss, J.W.; Ramji, D.P. Cytokines: Roles in atherosclerosis disease progression and potential therapeutic targets. Future Med. Chem. 2016, 8, 1317–1330. [Google Scholar] [CrossRef] [PubMed]
- Cai, R.; Liu, L.; Luo, B.; Wang, J.; Shen, J.; Shen, Y.; Zhang, R.; Chen, J.; Lu, H. Caspase-1 Activity in CD4 T Cells Is Downregulated Following Antiretroviral Therapy for HIV-1 Infection. AIDS Res. Hum. Retroviruses 2017, 33, 164–171. [Google Scholar] [CrossRef]
- Cai, Y.; Arikkath, J.; Yang, L.; Guo, M.L.; Periyasamy, P.; Buch, S. Interplay of endoplasmic reticulum stress and autophagy in neurodegenerative disorders. Autophagy 2016, 12, 225–244. [Google Scholar] [CrossRef]
- Hernandez, J.C.; Latz, E.; Urcuqui-Inchima, S. HIV-1 induces the first signal to activate the NLRP3 inflammasome in monocyte-derived macrophages. Intervirology 2014, 57, 36–42. [Google Scholar] [CrossRef]
- Guo, H.; Gao, J.; Taxman, D.J.; Ting, J.P.; Su, L. HIV-1 infection induces interleukin-1beta production via TLR8 protein-dependent and NLRP3 inflammasome mechanisms in human monocytes. J. Biol. Chem. 2014, 289, 21716–21726. [Google Scholar] [CrossRef]
- Chivero, E.T.; Guo, M.L.; Periyasamy, P.; Liao, K.; Callen, S.E.; Buch, S. HIV-1 Tat Primes and Activates Microglial NLRP3 Inflammasome-Mediated Neuroinflammation. J. Neurosci. 2017, 37, 3599–3609. [Google Scholar] [CrossRef]
- Song, J.; Jiao, Y.; Zhang, T.; Zhang, Y.; Huang, X.; Li, H.; Wu, H. Longitudinal changes in plasma Caspase-1 and Caspase-3 during the first 2 years of HIV-1 infection in CD4Low and CD4High patient groups. PLoS ONE 2015, 10, e0121011. [Google Scholar] [CrossRef] [PubMed]
- Lu, W.; Demers, A.J.; Ma, F.; Kang, G.; Yuan, Z.; Wan, Y.; Li, Y.; Xu, J.; Lewis, M.; Li, Q. Next-Generation mRNA Sequencing Reveals Pyroptosis-Induced CD4+ T Cell Death in Early Simian Immunodeficiency Virus-Infected Lymphoid Tissues. J. Virol. 2015, 90, 1080–1087. [Google Scholar] [CrossRef] [PubMed]
- Munoz-Arias, I.; Doitsh, G.; Yang, Z.; Sowinski, S.; Ruelas, D.; Greene, W.C. Blood-Derived CD4 T Cells Naturally Resist Pyroptosis during Abortive HIV-1 Infection. Cell Host Microbe 2015, 18, 463–470. [Google Scholar] [CrossRef]
- Galloway, N.L.; Doitsh, G.; Monroe, K.M.; Yang, Z.; Munoz-Arias, I.; Levy, D.N.; Greene, W.C. Cell-to-Cell Transmission of HIV-1 Is Required to Trigger Pyroptotic Death of Lymphoid-Tissue-Derived CD4 T Cells. Cell Rep. 2015, 12, 1555–1563. [Google Scholar] [CrossRef]
- Doitsh, G.; Galloway, N.L.; Geng, X.; Yang, Z.; Monroe, K.M.; Zepeda, O.; Hunt, P.W.; Hatano, H.; Sowinski, S.; Munoz-Arias, I.; et al. Cell death by pyroptosis drives CD4 T-cell depletion in HIV-1 infection. Nature 2014, 505, 509–514. [Google Scholar] [CrossRef] [PubMed]
- Kearns, A.C.; Liu, F.; Dai, S.; Robinson, J.A.; Kiernan, E.; Tesfaye Cheru, L.; Peng, X.; Gordon, J.; Morgello, S.; Abuova, A.; et al. Caspase-1 Activation Is Related With HIV-Associated Atherosclerosis in an HIV Transgenic Mouse Model and HIV Patient Cohort. Arterioscler. Thromb. Vasc. Biol. 2019, 39, 1762–1775. [Google Scholar] [CrossRef] [PubMed]
- Kearns, A.C.; Robinson, J.A.; Shekarabi, M.; Liu, F.; Qin, X.; Burdo, T.H. Caspase-1-associated immune activation in an accelerated SIV-infected rhesus macaque model. J. Neurovirol. 2018, 24, 420–431. [Google Scholar] [CrossRef] [PubMed]
- Buendia, J.; Sears, S.; Mgbere, O. Prevalence and risk factors of high cholesterol and triglycerides among people with HIV in Texas. AIDS Res. Ther. 2022, 19, 43. [Google Scholar] [CrossRef]
- Calza, L.; Manfredi, R.; Chiodo, F. Dyslipidaemia associated with antiretroviral therapy in HIV-infected patients. J. Antimicrob. Chemother. 2004, 53, 10–14. [Google Scholar] [CrossRef] [PubMed]
- Pedersen, K.K.; Pedersen, M.; Troseid, M.; Gaardbo, J.C.; Lund, T.T.; Thomsen, C.; Gerstoft, J.; Kvale, D.; Nielsen, S.D. Microbial translocation in HIV infection is associated with dyslipidemia, insulin resistance, and risk of myocardial infarction. J. Acquir. Immune. Defic. Syndr. 2013, 64, 425–433. [Google Scholar] [CrossRef]
- Vestad, B.; Nyman, T.A.; Hove-Skovsgaard, M.; Stensland, M.; Hoel, H.; Troseid, A.S.; Aspelin, T.; Aass, H.C.D.; Puhka, M.; Hov, J.R.; et al. Plasma extracellular vesicles in people living with HIV and type 2 diabetes are related to microbial translocation and cardiovascular risk. Sci. Rep. 2021, 11, 21936. [Google Scholar] [CrossRef]
- Palmer, C.S.; Cherry, C.L.; Sada-Ovalle, I.; Singh, A.; Crowe, S.M. Glucose Metabolism in T Cells and Monocytes: New Perspectives in HIV Pathogenesis. EBioMedicine 2016, 6, 31–41. [Google Scholar] [CrossRef]
- Henrick, B.M.; Yao, X.D.; Rosenthal, K.L.; INFANT Study Team. HIV-1 Structural Proteins Serve as PAMPs for TLR2 Heterodimers Significantly Increasing Infection and Innate Immune Activation. Front. Immunol. 2015, 6, 426. [Google Scholar] [CrossRef]
- Meas, H.Z.; Haug, M.; Beckwith, M.S.; Louet, C.; Ryan, L.; Hu, Z.; Landskron, J.; Nordbo, S.A.; Tasken, K.; Yin, H.; et al. Sensing of HIV-1 by TLR8 activates human T cells and reverses latency. Nat. Commun. 2020, 11, 147. [Google Scholar] [CrossRef]
- Nabatanzi, R.; Ssekamatte, P.; Castelnuovo, B.; Kambugu, A.; Nakanjako, D. Increased Levels of Caspase-1 and IL-1beta Among Adults With Persistent Immune Activation After 12 Years of Suppressive Antiretroviral Therapy in the Infectious Diseases Institute HIV Treatment Cohort. Open Forum. Infect. Dis. 2023, 10, ofad539. [Google Scholar] [CrossRef] [PubMed]
- Torre, D.; Pugliese, A. Interleukin-18: A proinflammatory cytokine in HIV-1 infection. Curr. HIV Res. 2006, 4, 423–430. [Google Scholar] [CrossRef] [PubMed]
- Feria, M.G.; Taborda, N.A.; Hernandez, J.C.; Rugeles, M.T. HIV replication is associated to inflammasomes activation, IL-1beta, IL-18 and caspase-1 expression in GALT and peripheral blood. PLoS ONE 2018, 13, e0192845. [Google Scholar] [CrossRef] [PubMed]
- Liang, H.; Xie, Z.; Shen, T. Monocyte activation and cardiovascular disease in HIV infection. Cell Mol. Immunol. 2017, 14, 960–962. [Google Scholar] [CrossRef] [PubMed]
- Miles, E.A.; Rees, D.; Banerjee, T.; Cazzola, R.; Lewis, S.; Wood, R.; Oates, R.; Tallant, A.; Cestaro, B.; Yaqoob, P.; et al. Age-related increases in circulating inflammatory markers in men are independent of BMI, blood pressure and blood lipid concentrations. Atherosclerosis 2008, 196, 298–305. [Google Scholar] [CrossRef]
- Funderburg, N.T.; Mayne, E.; Sieg, S.F.; Asaad, R.; Jiang, W.; Kalinowska, M.; Luciano, A.A.; Stevens, W.; Rodriguez, B.; Brenchley, J.M.; et al. Increased tissue factor expression on circulating monocytes in chronic HIV infection: Relationship to in vivo coagulation and immune activation. Blood 2010, 115, 161–167. [Google Scholar] [CrossRef] [PubMed]
- Ryan, L.A.; Zheng, J.; Brester, M.; Bohac, D.; Hahn, F.; Anderson, J.; Ratanasuwan, W.; Gendelman, H.E.; Swindells, S. Plasma levels of soluble CD14 and tumor necrosis factor-alpha type II receptor correlate with cognitive dysfunction during human immunodeficiency virus type 1 infection. J. Infect. Dis. 2001, 184, 699–706. [Google Scholar] [CrossRef] [PubMed]
- Rogacev, K.S.; Cremers, B.; Zawada, A.M.; Seiler, S.; Binder, N.; Ege, P.; Grosse-Dunker, G.; Heisel, I.; Hornof, F.; Jeken, J.; et al. CD14++CD16+ monocytes independently predict cardiovascular events: A cohort study of 951 patients referred for elective coronary angiography. J. Am. Coll. Cardiol. 2012, 60, 1512–1520. [Google Scholar] [CrossRef] [PubMed]
- Teer, E.; Joseph, D.E.; Driescher, N.; Nell, T.A.; Dominick, L.; Midgley, N.; Deshpande, G.; Page, M.J.; Pretorius, E.; Woudberg, N.J.; et al. HIV and cardiovascular diseases risk: Exploring the interplay between T-cell activation, coagulation, monocyte subsets, and lipid subclass alterations. Am. J. Physiol. Heart Circ. Physiol. 2019, 316, H1146–H1157. [Google Scholar] [CrossRef]
- Veenhuis, R.T.; Williams, D.W.; Shirk, E.N.; Abreu, C.M.; Ferreira, E.A.; Coughlin, J.M.; Brown, T.T.; Maki, P.M.; Anastos, K.; Berman, J.W.; et al. Higher circulating intermediate monocytes are associated with cognitive function in women with HIV. JCI Insight 2021, 6, e146215. [Google Scholar] [CrossRef]
- Cheru, L.T.; Park, E.A.; Saylor, C.F.; Burdo, T.H.; Fitch, K.V.; Looby, S.; Weiner, J.; Robinson, J.A.; Hubbard, J.; Torriani, M.; et al. I-FABP Is Higher in People With Chronic HIV Than Elite Controllers, Related to Sugar and Fatty Acid Intake and Inversely Related to Body Fat in People With HIV. Open Forum. Infect. Dis. 2018, 5, ofy288. [Google Scholar] [CrossRef] [PubMed]
- Kausalya, B.; Saravanan, S.; Pallikkuth, S.; Pahwa, R.; Saini, S.R.; Iqbal, S.; Solomon, S.; Murugavel, K.G.; Poongulali, S.; Kumarasamy, N.; et al. Immune correlates of cardiovascular co-morbidity in HIV infected participants from South India. BMC Immunol. 2022, 23, 24. [Google Scholar] [CrossRef] [PubMed]
- Sukumaran, L.; Kunisaki, K.M.; Bakewell, N.; Winston, A.; Mallon, P.W.G.; Doyle, N.; Anderson, J.; Boffito, M.; Haddow, L.; Post, F.A.; et al. Association between inflammatory biomarker profiles and cardiovascular risk in individuals with and without HIV. AIDS 2023, 37, 595–603. [Google Scholar] [CrossRef] [PubMed]
- Ding, A.; Wallis, Z.; White, K.; Efe, C.; Kim, W.K.; Ardeshir, A.; Williams, K. Galectin-3, galectin-9, and IL-18 are associated with monocyte activation and turnover more so than SIV-associated cardiac pathology or encephalitis. AIDS Res. Hum. Retroviruses 2024. [Google Scholar] [CrossRef] [PubMed]
- Grinspoon, S.; Carr, A. Cardiovascular risk and body-fat abnormalities in HIV-infected adults. N. Engl. J. Med. 2005, 352, 48–62. [Google Scholar] [CrossRef] [PubMed]
- Palmer, C.S.; Anzinger, J.J.; Zhou, J.; Gouillou, M.; Landay, A.; Jaworowski, A.; McCune, J.M.; Crowe, S.M. Glucose transporter 1-expressing proinflammatory monocytes are elevated in combination antiretroviral therapy-treated and untreated HIV+ subjects. J. Immunol. 2014, 193, 5595–5603. [Google Scholar] [CrossRef] [PubMed]
- Valle-Casuso, J.C.; Angin, M.; Volant, S.; Passaes, C.; Monceaux, V.; Mikhailova, A.; Bourdic, K.; Avettand-Fenoel, V.; Boufassa, F.; Sitbon, M.; et al. Cellular Metabolism Is a Major Determinant of HIV-1 Reservoir Seeding in CD4+ T Cells and Offers an Opportunity to Tackle Infection. Cell Metab. 2019, 29, 611–626.e615. [Google Scholar] [CrossRef] [PubMed]
- Thet, D.; Siritientong, T. Antiretroviral Therapy-Associated Metabolic Complications: Review of the Recent Studies. HIV AIDS 2020, 12, 507–524. [Google Scholar] [CrossRef]
- Del Prete, G.Q.; Smedley, J.; Macallister, R.; Jones, G.S.; Li, B.; Hattersley, J.; Zheng, J.; Piatak, M., Jr.; Keele, B.F.; Hesselgesser, J.; et al. Short Communication: Comparative Evaluation of Coformulated Injectable Combination Antiretroviral Therapy Regimens in Simian Immunodeficiency Virus-Infected Rhesus Macaques. AIDS Res. Hum. Retroviruses 2016, 32, 163–168. [Google Scholar] [CrossRef]
- Sugimoto, C.; Hasegawa, A.; Saito, Y.; Fukuyo, Y.; Chiu, K.B.; Cai, Y.; Breed, M.W.; Mori, K.; Roy, C.J.; Lackner, A.A.; et al. Differentiation Kinetics of Blood Monocytes and Dendritic Cells in Macaques: Insights to Understanding Human Myeloid Cell Development. J. Immunol. 2015, 195, 1774–1781. [Google Scholar] [CrossRef]

Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Thirugnanam, S.; Wang, C.; Zheng, C.; Grasperge, B.F.; Datta, P.K.; Rappaport, J.; Qin, X.; Rout, N. The Association between IL-1β and IL-18 Levels, Gut Barrier Disruption, and Monocyte Activation during Chronic Simian Immunodeficiency Virus Infection and Long-Term Suppressive Antiretroviral Therapy. Int. J. Mol. Sci. 2024, 25, 8702. https://doi.org/10.3390/ijms25168702
Thirugnanam S, Wang C, Zheng C, Grasperge BF, Datta PK, Rappaport J, Qin X, Rout N. The Association between IL-1β and IL-18 Levels, Gut Barrier Disruption, and Monocyte Activation during Chronic Simian Immunodeficiency Virus Infection and Long-Term Suppressive Antiretroviral Therapy. International Journal of Molecular Sciences. 2024; 25(16):8702. https://doi.org/10.3390/ijms25168702
Chicago/Turabian StyleThirugnanam, Siva, Chenxiao Wang, Chen Zheng, Brooke F. Grasperge, Prasun K. Datta, Jay Rappaport, Xuebin Qin, and Namita Rout. 2024. "The Association between IL-1β and IL-18 Levels, Gut Barrier Disruption, and Monocyte Activation during Chronic Simian Immunodeficiency Virus Infection and Long-Term Suppressive Antiretroviral Therapy" International Journal of Molecular Sciences 25, no. 16: 8702. https://doi.org/10.3390/ijms25168702