
The Immune Memory Response of In Vitro-Polarised Th1, Th2, and Th17 Cells in the Face of Ovalbumin-Transgenic Leishmania major in a Mouse Model

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
1. Introduction
2. Results
2.1. Cloning of sOVA and mOVA in Leishmania Expression Vector (pIR1SAT)
2.2. Expression of OVA by Transfected L. major
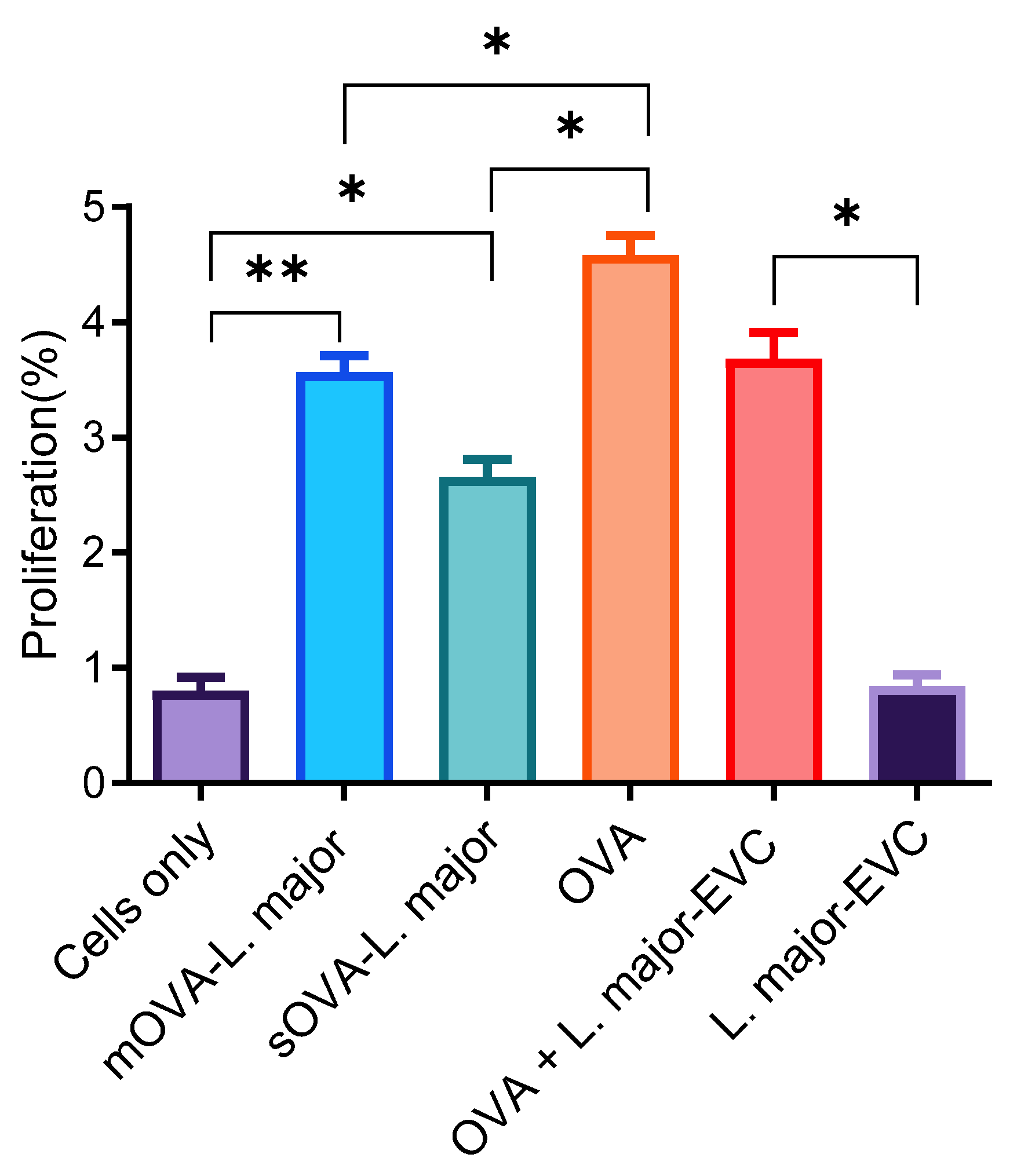
2.3. OT-II CD4+ T Cell Proliferation by OVA-Expressing L. major
2.4. Expression of Th Cytokines Following Activation with OVA-Expressing L. major
2.5. OT-II T Cells’ Response to OVA-Expressing L. major
2.6. Ex Vivo Response of OT-II T Cells to OVA-Expressing L. major
2.7. Cytokine Production of Recovered Cells Following Stimulation with OVA-Expressing L. major
2.8. Stimulation and Proliferation Rate of Recovered Antigen-Experienced Cells
2.9. Cytokine Production by Recovered Th-Polarised Cells Following Stimulation with OVA-Expressing L. major
2.10. Measuring Immune Memory Resilience during L. major Exposure
3. Discussion
4. Materials and Methods
4.1. Experimental Mice
4.2. Leishmania Major Culture
4.3. Cloning of the OVA Gene into L. major Expression Vector pIR1SAT
4.4. Genetic Manipulation of L. major
4.5. Confirming OVA Expression by L. major
4.6. Lymphocyte Preparation
4.7. In Vitro Polarisation of OT-II T Cells
4.8. Adoptive Transferral of OT-II T Cells to Congenic Mice and Parasite Injection
4.9. T Cell Proliferation Assay
4.10. Cytokine Assay
4.11. Statistical Analyses
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Pearson, R.D.; Wheeler, D.A.; Harrison, L.H.; Kay, H.D. The immunobiology of leishmaniasis. Rev. Infect. Dis. 1983, 5, 907–927. [Google Scholar] [CrossRef] [PubMed]
- Scott, P.; Novais, F. Cutaneous leishmaniasis: Immune responses in protection and pathogenesis. Nat. Rev. Immunol. 2016, 16, 581–592. [Google Scholar] [CrossRef]
- Liew, F.Y.; Hale, C.; Howard, J.G. Immunologic regulation of experimental cutaneous leishmaniasis. V. Characterization of effector and specific suppressor T cells. J. Immunol. 1982, 128, 1917–1922. [Google Scholar] [CrossRef] [PubMed]
- Carvalho, E.M.; Badaró, R.; Reed, S.G.; Jones, T.C.; Johnson, W.D., Jr. Absence of gamma interferon and interleukin 2 production during active visceral leishmaniasis. J. Clin. Investig. 1985, 76, 2066–2069. [Google Scholar] [CrossRef] [PubMed] [PubMed Central]
- Murray, H.W.; Rubin, B.Y.; Carriero, S.; Acosta, A.M. Reversible defect in antigen-induced lymphokine and gamma-interferon generation in cutaneous leishmaniasis. J. Immunol. 1984, 133, 2250–2254. [Google Scholar] [CrossRef] [PubMed]
- Sadick, M.D.; Locksley, R.M.; Tubbs, C.; Raff, H.V. Murine cutaneous leishmaniasis: Resistance correlates with the capacity to generate interferon-gamma in response to Leishmania antigens in vitro. J. Immunol. 1986, 136, 655–661. [Google Scholar] [CrossRef] [PubMed]
- Nathan, C.F.; Murray, H.W.; Wiebe, M.E.; Rubin, B.Y. Identification of interferon-gamma as the lymphokine that activates human macrophage oxidative metabolism and antimicrobial activity. J. Exp. Med. 1983, 158, 670–689. [Google Scholar] [CrossRef] [PubMed] [PubMed Central]
- Murray, H.W.; Rubin, B.Y.; Rothermel, C.D. Killing of intracellular Leishmania donovani by lymphokine-stimulated human mononuclear phagocytes. Evidence that interferon-gamma is the activating lymphokine. J. Clin. Investig. 1983, 72, 1506–1510. [Google Scholar] [CrossRef]
- Crawford, R.M.; Finbloom, D.S.; Ohara, J.; Paul, W.E.; Meltzer, M.S. B cell stimulatory factor-1 (interleukin 4) activates macrophages for increased tumoricidal activity and expression of Ia antigens. J. Immunol. 1987, 139, 135–141. [Google Scholar] [CrossRef] [PubMed]
- Blackwell, J.M.; Ezekowitz, R.A.; Roberts, M.B.; Channon, J.Y.; Sim, R.B.; Gordon, S. Macrophage complement and lectin-like receptors bind Leishmania in the absence of serum. J. Exp. Med. 1985, 162, 324–331. [Google Scholar] [CrossRef] [PubMed] [PubMed Central]
- Mosser, D.M.; Edelson, P.J. The mouse macrophage receptor for C3bi (CR3) is a major mechanism in the phagocytosis of Leishmania promastigotes. J. Immunol. 1985, 135, 2785–2789. [Google Scholar] [CrossRef] [PubMed]
- Reiner, S.L.; Locksley, R.M. The regulation of immunity to Leishmania major. Annu. Rev. Immunol. 1995, 13, 151–177. [Google Scholar] [CrossRef] [PubMed]
- Sacks, D.; Noben-Trauth, N. The immunology of susceptibility and resistance to Leishmania major in mice. Nat. Rev. Immunol. 2002, 2, 845–858. [Google Scholar] [CrossRef] [PubMed]
- Scott, P.; Artis, D.; Uzonna, J.; Zaph, C. The development of effector and memory T cells in cutaneous leishmaniasis: The implications for vaccine development. Immunol. Rev. 2004, 201, 318–338. [Google Scholar] [CrossRef] [PubMed]
- Mond, J.J.; Carman, J.; Sarma, C.; Ohara, J.; Finkelman, F.D. Interferon-gamma suppresses B cell stimulation factor (BSF-1) induction of class II MHC determinants on B cells. J. Immunol. 1986, 137, 3534–3537. [Google Scholar] [CrossRef] [PubMed]
- Reynolds, D.S.; Boom, W.H.; Abbas, A.K. Inhibition of B lymphocyte activation by interferon-gamma. J. Immunol. 1987, 139, 767–773. [Google Scholar] [CrossRef] [PubMed]
- Gajewski, T.F.; Joyce, J.; Fitch, F.W. Antiproliferative effect of IFN-gamma in immune regulation. III. Differential selection of TH1 and TH2 murine helper T lymphocyte clones using recombinant IL-2 and recombinant IFN-gamma. J. Immunol. 1989, 143, 15–22. [Google Scholar] [CrossRef] [PubMed]
- Lehn, M.; Weiser, W.Y.; Engelhorn, S.; Gillis, S.; Remold, H.G. IL-4 inhibits H2O2 production and antileishmanial capacity of human cultured monocytes mediated by IFN-gamma. J. Immunol. 1989, 143, 3020–3024. [Google Scholar] [CrossRef] [PubMed]
- Pagán, A.J.; Peters, N.C.; Debrabant, A.; Ribeiro-Gomes, F.; Pepper, M.; Karp, C.L.; Jenkins, M.K.; Sacks, D.L. Tracking antigen-specific CD4+ T cells throughout the course of chronic Leishmania major infection in resistant mice. Eur. J. Immunol. 2013, 43, 427–438. [Google Scholar] [CrossRef] [PubMed] [PubMed Central]
- Heinzel, F.P.; Sadick, M.D.; Holaday, B.J.; Coffman, R.L.; Locksley, R.M. Reciprocal expression of interferon gamma or interleukin 4 during the resolution or progression of murine leishmaniasis. Evidence for expansion of distinct helper T cell subsets. J. Exp. Med. 1989, 169, 59–72. [Google Scholar] [CrossRef] [PubMed] [PubMed Central]
- Scott, P. The role of TH1 and TH2 cells in experimental cutaneous leishmaniasis. Exp. Parasitol. 1989, 68, 369–372. [Google Scholar] [CrossRef] [PubMed]
- Romano, A.; Carneiro, M.B.H.; Doria, N.A.; Roma, E.H.; Ribeiro-Gomes, F.L.; Inbar, E.; Lee, S.H.; Mendez, J.; Paun, A.; Sacks, D.L.; et al. Divergent roles for Ly6C+CCR2+CX3CR1+ inflammatory monocytes during primary or secondary infection of the skin with the intra-phagosomal pathogen Leishmania major. PLoS Pathog. 2017, 13, e1006479. [Google Scholar] [CrossRef] [PubMed] [PubMed Central]
- Peters, N.C.; Pagán, A.J.; Lawyer, P.G.; Hand, T.W.; Henrique Roma, E.; Stamper, L.W.; Romano, A.; Sacks, D.L. Chronic parasitic infection maintains high frequencies of short-lived Ly6C+CD4+ effector T cells that are required for protection against re-infection. PLoS Pathog. 2014, 10, e1004538. [Google Scholar] [CrossRef] [PubMed] [PubMed Central]
- Volpedo, G.; Pacheco-Fernandez, T.; Bhattacharya, P.; Oljuskin, T.; Dey, R.; Gannavaram, S.; Satoskar, A.R.; Nakhasi, H.L. Determinants of Innate Immunity in Visceral Leishmaniasis and Their Implication in Vaccine Development. Front. Immunol. 2021, 12, 748325. [Google Scholar] [CrossRef] [PubMed] [PubMed Central]
- Volpedo, G.; Pacheco-Fernandez, T.; Holcomb, E.A.; Zhang, W.W.; Lypaczewski, P.; Cox, B.; Fultz, R.; Mishan, C.; Verma, C.; Huston, R.H.; et al. Centrin-deficient Leishmania mexicana confers protection against New World cutaneous leishmaniasis. NPJ Vaccines 2022, 7, 32. [Google Scholar] [CrossRef] [PubMed] [PubMed Central]
- Zutshi, S.; Kumar, S.; Chauhan, P.; Bansode, Y.; Nair, A.; Roy, S.; Sarkar, A.; Saha, B. Anti-Leishmanial Vaccines: Assumptions, Approaches, and Annulments. Vaccines 2019, 7, 156. [Google Scholar] [CrossRef] [PubMed] [PubMed Central]
- Prickett, S.; Gray, P.M.; Colpitts, S.L.; Scott, P.; Kaye, P.M.; Smith, D.F. In vivo recognition of ovalbumin expressed by transgenic Leishmania is determined by its subcellular localization. J. Immunol. 2006, 176, 4826–4833. [Google Scholar] [CrossRef] [PubMed] [PubMed Central]
- Beattie, L.; Evans, K.J.; Kaye, P.M.; Smith, D.F. Transgenic Leishmania and the immune response to infection. Parasite Immunol. 2008, 30, 255–266. [Google Scholar] [CrossRef] [PubMed] [PubMed Central]
- Kaye, P.M.; Coburn, C.; McCrossan, M.; Beverley, S.M. Antigens targeted to the Leishmania phagolysosome are processed for CD4+ T cell recognition. Eur. J. Immunol. 1993, 23, 2311–2319. [Google Scholar] [CrossRef] [PubMed]
- Olivier, M.; Gregory, D.J.; Forget, G. Subversion mechanisms by which Leishmania parasites can escape the host immune response: A signaling point of view. Clin. Microbiol. Rev. 2005, 18, 293–305. [Google Scholar] [CrossRef]
- Sutterwala, F.S.; Noel, G.J.; Salgame, P.; Mosser, D.M. Reversal of proinflammatory responses by ligating the macrophage Fcgamma receptor type I. J. Exp. Med. 1998, 188, 217–222. [Google Scholar] [CrossRef] [PubMed] [PubMed Central]
- Cunningham, A.C. Parasitic adaptive mechanisms in infection by leishmania. Exp. Mol. Pathol. 2002, 72, 132–141. [Google Scholar] [CrossRef] [PubMed]
- Kane, M.M.; Mosser, D.M. Leishmania parasites and their ploys to disrupt macrophage activation. Curr. Opin. Hematol. 2000, 7, 26–31. [Google Scholar] [CrossRef] [PubMed]
- Assreuy, J.; Cunha, F.Q.; Epperlein, M.; Noronha-Dutra, A.; O’Donnell, C.A.; Liew, F.Y.; Moncada, S. Production of nitric oxide and superoxide by activated macrophages and killing of Leishmania major. Eur. J. Immunol. 1994, 24, 672–676. [Google Scholar] [CrossRef] [PubMed]
- Peters, N.C.; Bertholet, S.; Lawyer, P.G.; Charmoy, M.; Romano, A.; Ribeiro-Gomes, F.L.; Stamper, L.W.; Sacks, D.L. Evaluation of recombinant Leishmania polyprotein plus glucopyranosyl lipid A stable emulsion vaccines against sand fly transmitted Leishmania major in C57BL/6 mice. J. Immunol. 2012, 189, 4832–4841. [Google Scholar] [CrossRef] [PubMed] [PubMed Central]
- Hohman, L.S.; Mou, Z.; Carneiro, M.B.; Ferland, G.; Kratofil, R.M.; Kubes, P.; Uzonna, J.E.; Peters, N.C. Protective CD4+ Th1 cell-mediated immunity is reliant upon execution of effector function prior to the establishment of the pathogen niche. PLoS Pathog. 2021, 17, e1009944. [Google Scholar] [CrossRef] [PubMed] [PubMed Central]
- Hohman, L.S.; Peters, N.C. CD4+ T Cell-Mediated Immunity against the Phagosomal Pathogen Leishmania: Implications for Vaccination. Trends Parasitol. 2019, 35, 423–435. [Google Scholar] [CrossRef]
- Zaph, C.; Uzonna, J.; Beverley, S.M.; Scott, P. Central memory T cells mediate long-term immunity to Leishmania major in the absence of persistent parasites. Nat. Med. Oct. 2004, 10, 1104–1110. [Google Scholar] [CrossRef]
- Glennie, N.D.; Yeramilli, V.A.; Beiting, D.P.; Volk, S.W.; Weaver, C.T.; Scott, P. Skin-resident memory CD4+ T cells enhance protection against Leishmania major infection. J. Exp. Med. Aug. 2015, 212, 1405–1414. [Google Scholar] [CrossRef]
- Glennie, N.D.; Volk, S.W.; Scott, P. Skin-resident CD4+ T cells protect against Leishmania major by recruiting and activating inflammatory monocytes. PLoS Pathog. 2017, 13, e1006349. [Google Scholar] [CrossRef]
- Glennie, N.D.; Scott, P. Memory T cells in cutaneous leishmaniasis. Cell Immunol. Nov. 2016, 309, 50–54. [Google Scholar] [CrossRef] [PubMed]
- Mahmoudzadeh-Niknam, H.; Khalili, G.; Abrishami, F.; Najafy, A.; Khaze, V. The route of Leishmania tropica infection determines disease outcome and protection against Leishmania major in BALB/c mice. Korean J. Parasitol. 2013, 51, 69–74. [Google Scholar] [CrossRef] [PubMed] [PubMed Central]
- Robinson, K.A.; Beverley, S.M. Improvements in transfection efficiency and tests of RNA interference (RNAi) approaches in the protozoan parasite Leishmania. Mol. Biochem. Parasitol. 2003, 128, 217–228. [Google Scholar] [CrossRef]
- Capul, A.A.; Barron, T.; Dobson, D.E.; Turco, S.J.; Beverley, S.M. Two functionally divergent UDP-Gal nucleotide sugar transporters participate in phosphoglycan synthesis in Leishmania major. J. Biol. Chem. 2007, 282, 14006–14017. [Google Scholar] [CrossRef] [PubMed]
- Mandal, S.; Maharjan, M.; Ganguly, S.; Chatterjee, M.; Singh, S.; Buckner, F.S.; Madhubala, R. High-throughput screening of amastigotes of Leishmania donovani clinical isolates against drugs using a colorimetric beta-lactamase assay. Indian J. Exp. Biol. 2009, 47, 475–479. [Google Scholar] [PubMed]
- Vandenhoff, M.J.B.; Moorman, A.F.M.; Lamers, W.H. Electroporation in Intracellular Buffer Increases Cell-Survival. Nucleic Acids Res. 1992, 20, 2902. [Google Scholar] [CrossRef] [PubMed]
- Bifeld, E.; Chrobak, M.; Zander, D.; Schleicher, U.; Schönian, G.; Clos, J. Geographical sequence variation in the Leishmania major virulence factor P46. Infect. Genet. Evol. 2015, 30, 195–205. [Google Scholar] [CrossRef] [PubMed]
- Tedla, M.G.; Every, A.L.; Scheerlinck, J.Y. Measuring the Manipulation of T Helper Immune Responses by Schistosoma mansoni. Int. J. Mol. Sci. 2022, 23, 1462. [Google Scholar] [CrossRef] [PubMed] [PubMed Central]
- Kondrack, R.M.; Harbertson, J.; Tan, J.T.; McBreen, M.E.; Surh, C.D.; Bradley, L.M. Interleukin 7 regulates the survival and generation of memory CD4 cells. J. Exp. Med. 2003, 198, 1797–1806. [Google Scholar] [CrossRef]
- Tedla, M.G.; Nahar, M.F.; Hagen, J.; Every, A.L.; Scheerlinck, J.-P.Y. Recognition of Schistosoma mansoni egg-expressed ovalbumin by T cell receptor transgenic mice. Exp. Parasitol. 2019, 206, 107767. [Google Scholar] [CrossRef]









Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Tedla, M.G.; Nahar, M.F.; Every, A.L.; Scheerlinck, J.-P.Y. The Immune Memory Response of In Vitro-Polarised Th1, Th2, and Th17 Cells in the Face of Ovalbumin-Transgenic Leishmania major in a Mouse Model. Int. J. Mol. Sci. 2024, 25, 8753. https://doi.org/10.3390/ijms25168753
Tedla MG, Nahar MF, Every AL, Scheerlinck J-PY. The Immune Memory Response of In Vitro-Polarised Th1, Th2, and Th17 Cells in the Face of Ovalbumin-Transgenic Leishmania major in a Mouse Model. International Journal of Molecular Sciences. 2024; 25(16):8753. https://doi.org/10.3390/ijms25168753
Chicago/Turabian StyleTedla, Mebrahtu G., Musammat F. Nahar, Alison L. Every, and Jean-Pierre Y. Scheerlinck. 2024. "The Immune Memory Response of In Vitro-Polarised Th1, Th2, and Th17 Cells in the Face of Ovalbumin-Transgenic Leishmania major in a Mouse Model" International Journal of Molecular Sciences 25, no. 16: 8753. https://doi.org/10.3390/ijms25168753
APA StyleTedla, M. G., Nahar, M. F., Every, A. L., & Scheerlinck, J.-P. Y. (2024). The Immune Memory Response of In Vitro-Polarised Th1, Th2, and Th17 Cells in the Face of Ovalbumin-Transgenic Leishmania major in a Mouse Model. International Journal of Molecular Sciences, 25(16), 8753. https://doi.org/10.3390/ijms25168753