
Investigating Genetic Overlap between Alzheimer’s Disease, Lipids, and Coronary Artery Disease: A Large-Scale Genome-Wide Cross Trait Analysis


Abstract
1. Introduction
2. Results
2.1. Global Genetic Correlation of AD with Lipids and CAD Traits
2.2. Results of Gene-Level Genetic Overlap Analysis
2.3. Genome-Wide Significant (Sentinel) Genes Shared by AD, Lipids, and CAD Traits
2.4. Shared Genes Reaching Genome-Wide Significance for AD, Lipids, and CAD Traits
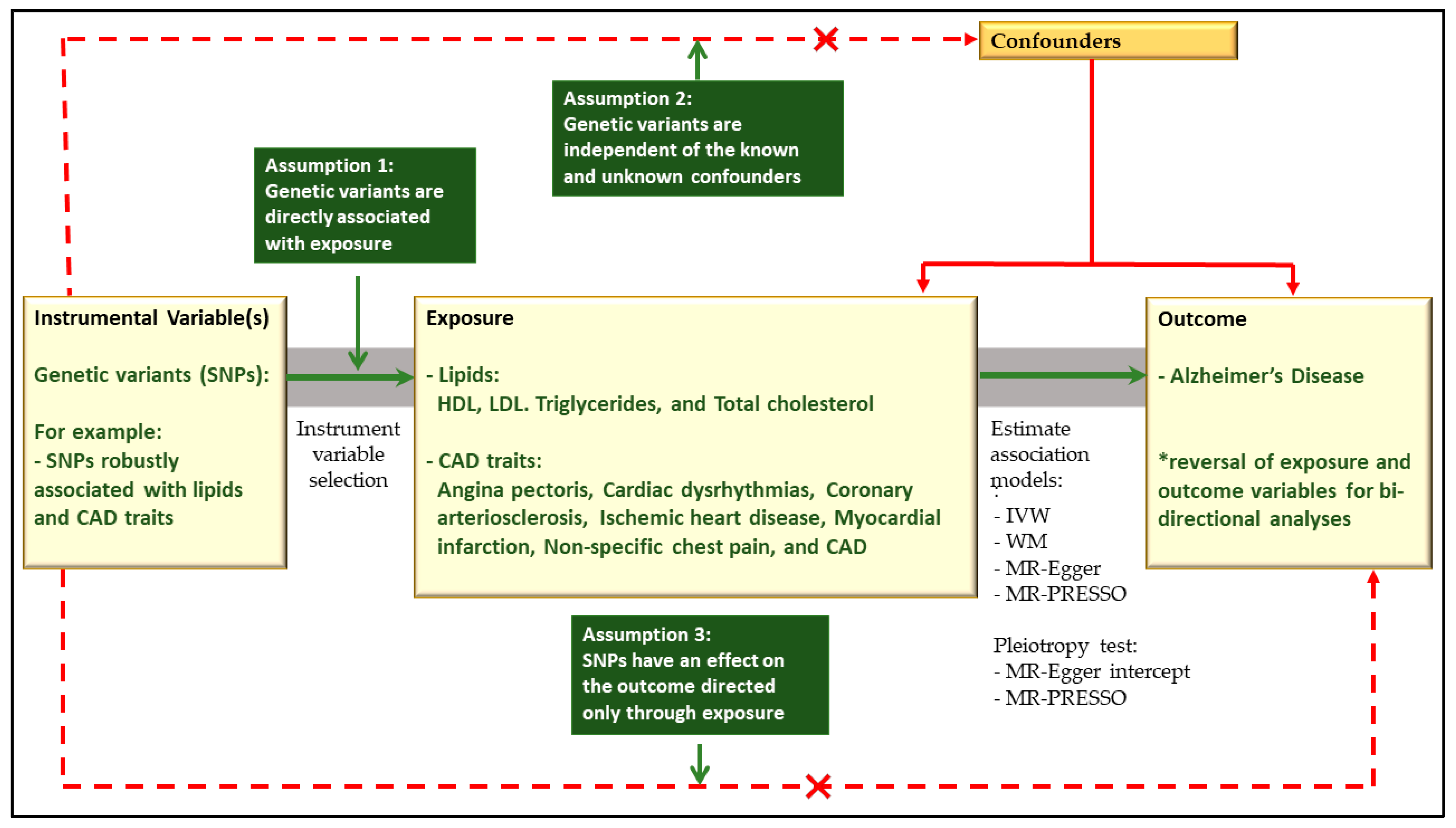
2.5. Results of Causal Relationship Assessment
2.5.1. No Causal Relationship of Lipids with Alzheimer’s Disease
2.5.2. No Causal Relationship of CAD Traits with Alzheimer’s Disease
2.6. Local Genetic Correlation of Alzheimer’s Disease with Lipids and CAD Traits
2.7. Comparing LDSC and LAVA Results
3. Discussion
4. Materials and Methods
4.1. Data Sources
4.2. Statistical Analyses
4.3. Assessing Global Genetic Correlation
4.4. Gene-Level Genetic Overlap Assessment
4.5. Identifying Genes Shared by AD, Lipids, and CAD Traits
4.6. Causal Relationship Assessment
4.7. Local Genetic Correlation Analysis
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Alzheimer’s Disease International; Guerchet, M.; Prince, M.; Prina, M. Numbers of People with Dementia around the World; 2020; Available online: https://www.alzint.org/resource/numbers-of-people-with-dementia-worldwide/ (accessed on 10 August 2024).
- Prince, M.J.; Wimo, A.; Guerchet, M.M.; Ali, G.C.; Wu, Y.-T.; Prina, M. World Alzheimer Report 2015—The Global Impact of Dementia: An Analysis of Prevalence, Incidence, Cost and Trends. Ph.D. Thesis, Alzheimer’s Disease International, London, UK, 2015. [Google Scholar]
- Alzheimer’s Disease International. Dementia Statistics. Available online: https://www.alzint.org/about/dementia-facts-figures/dementia-statistics/ (accessed on 23 August 2023).
- Australian Institute of Health and Welfare. Deaths in Australia. Available online: https://www.aihw.gov.au/reports/life-expectancy-deaths/deaths-in-australia/contents/leading-causes-of-death (accessed on 24 August 2023).
- Glenner, G.G.; Wong, C.W. Alzheimer’s disease: Initial report of the purification and characterization of a novel cerebrovascular amyloid protein. Biochem. Biophys. Res. Commun. 1984, 120, 885–890. [Google Scholar] [CrossRef] [PubMed]
- Goedert, M.; Spillantini, M.G.; Jakes, R.; Rutherford, D.; Crowther, R. Multiple isoforms of human microtubule-associated protein tau: Sequences and localization in neurofibrillary tangles of Alzheimer’s disease. Neuron 1989, 3, 519–526. [Google Scholar] [CrossRef] [PubMed]
- Fuller, S.C. Alzheimer’s disease (senium praecox): The report of a case and review of published cases. J. Nerv. Ment. Dis. 1912, 39, 440–455. [Google Scholar] [CrossRef]
- Hippius, H.; Neundörfer, G. The discovery of Alzheimer’s disease. Dialogues Clin. Neurosci. 2003, 6, 101–108. [Google Scholar] [CrossRef] [PubMed]
- Fowler, C.; Rainey-Smith, S.R.; Bird, S.; Bomke, J.; Bourgeat, P.; Brown, B.M.; Burnham, S.C.; Bush, A.I.; Chadunow, C.; Collins, S.; et al. Fifteen years of the australian imaging, biomarkers and lifestyle (AIBL) study: Progress and observations from 2,359 older adults spanning the spectrum from cognitive normality to Alzheimer’s disease. J. Alzheimer’s Dis. Rep. 2021, 5, 443–468. [Google Scholar] [CrossRef] [PubMed]
- Livingston, G.; Huntley, J.; Sommerlad, A.; Ames, D.; Ballard, C.; Banerjee, S.; Brayne, C.; Burns, A.; Cohen-Mansfield, J.; Cooper, C.; et al. Dementia prevention, intervention, and care: 2020 report of the Lancet Commission. Lancet 2020, 396, 413–446. [Google Scholar] [CrossRef] [PubMed]
- Asher, S.; Priefer, R. Alzheimer’s disease failed clinical trials. Life Sci. 2022, 306, 120861. [Google Scholar] [CrossRef] [PubMed]
- Consortium, G.L.G. Discovery and refinement of loci associated with lipid levels. Nat. Genet. 2013, 45, 1274–1283. [Google Scholar] [CrossRef]
- Schunkert, H.; König, I.R.; Kathiresan, S.; Reilly, M.P.; Assimes, T.L.; Holm, H.; Preuss, M.; Stewart, A.F.; Barbalic, M.; Gieger, C. Large-scale association analysis identifies 13 new susceptibility loci for coronary artery disease. Nat. Genet. 2011, 43, 333–338. [Google Scholar] [CrossRef]
- Cadby, G.; Giles, C.; Melton, P.E.; Huynh, K.; Mellett, N.A.; Duong, T.; Nguyen, A.; Cinel, M.; Smith, A.; Olshansky, G. Comprehensive genetic analysis of the human lipidome identifies loci associated with lipid homeostasis with links to coronary artery disease. Nat. Commun. 2022, 13, 3124. [Google Scholar] [CrossRef]
- Lord, J.; Jermy, B.; Green, R.; Wong, A.; Xu, J.; Legido-Quigley, C.; Dobson, R.; Richards, M.; Proitsi, P. Mendelian randomization identifies blood metabolites previously linked to midlife cognition as causal candidates in Alzheimer’s disease. Proc. Natl. Acad. Sci. USA 2021, 118, e2009808118. [Google Scholar] [CrossRef] [PubMed]
- van der Linden, R.J.; Reus, L.M.; De Witte, W.; Tijms, B.M.; Olde Rikkert, M.; Visser, P.J.; Poelmans, G. Genetic overlap between Alzheimer’s disease and blood lipid levels. Neurobiol. Aging 2021, 108, 189–195. [Google Scholar] [CrossRef] [PubMed]
- Grace, C.; Clarke, R.; Goel, A.; Farrall, M.; Watkins, H.; Hopewell, J.C. Lack of genetic support for shared aetiology of coronary artery disease and late-onset Alzheimer’s disease. Sci. Rep. 2018, 8, 7102. [Google Scholar] [CrossRef]
- Van Meer, G.; Voelker, D.R.; Feigenson, G.W. Membrane lipids: Where they are and how they behave. Nat. Rev. Mol. Cell Biol. 2008, 9, 112–124. [Google Scholar] [CrossRef] [PubMed]
- Adewuyi, E.O.; O’Brien, E.K.; Nyholt, D.R.; Porter, T.; Laws, S.M. A large-scale genome-wide cross-trait analysis reveals shared genetic architecture between Alzheimer’s disease and gastrointestinal tract disorders. Commun. Biol. 2022, 5, 691. [Google Scholar] [CrossRef]
- Kao, Y.-C.; Ho, P.-C.; Tu, Y.-K.; Jou, I.-M.; Tsai, K.-J. Lipids and Alzheimer’s disease. Int. J. Mol. Sci. 2020, 21, 1505. [Google Scholar] [CrossRef]
- Liu, Y.; Thalamuthu, A.; Mather, K.A.; Crawford, J.; Ulanova, M.; Wong, M.W.K.; Pickford, R.; Sachdev, P.S.; Braidy, N. Plasma lipidome is dysregulated in Alzheimer’s disease and is associated with disease risk genes. Transl. Psychiatry 2021, 11, 344. [Google Scholar] [CrossRef]
- Fox, K.M.; Ferrari, R. Heart rate: A forgotten link in coronary artery disease? Nat. Rev. Cardiol. 2011, 8, 369–379. [Google Scholar] [CrossRef]
- Lathe, R.; Sapronova, A.; Kotelevtsev, Y. Atherosclerosis and Alzheimer-diseases with a common cause? Inflammation, oxysterols, vasculature. BMC Geriatr. 2014, 14, 36. [Google Scholar] [CrossRef]
- Qiu, C.; Fratiglioni, L. A major role for cardiovascular burden in age-related cognitive decline. Nat. Rev. Cardiol. 2015, 12, 267–277. [Google Scholar] [CrossRef]
- Xie, W.; Zheng, F.; Yan, L.; Zhong, B. Cognitive decline before and after incident coronary events. J. Am. Coll. Cardiol. 2019, 73, 3041–3050. [Google Scholar] [CrossRef] [PubMed]
- Liang, J.; Li, C.; Gao, D.; Ma, Q.; Wang, Y.; Pan, Y.; Zhang, W.; Xie, W.; Zheng, F. Association Between Onset Age of Coronary Heart Disease and Incident Dementia: A Prospective Cohort Study. J. Am. Heart Assoc. 2023, 12, e031407. [Google Scholar] [CrossRef] [PubMed]
- Wolters, F.J.; Segufa, R.A.; Darweesh, S.K.; Bos, D.; Ikram, M.A.; Sabayan, B.; Hofman, A.; Sedaghat, S. Coronary heart disease, heart failure, and the risk of dementia: A systematic review and meta-analysis. Alzheimer’s Dement. 2018, 14, 1493–1504. [Google Scholar] [CrossRef] [PubMed]
- Jansen, I.E.; Savage, J.E.; Watanabe, K.; Bryois, J.; Williams, D.M.; Steinberg, S.; Sealock, J.; Karlsson, I.K.; Hägg, S.; Athanasiu, L.; et al. Genome-wide meta-analysis identifies new loci and functional pathways influencing Alzheimer’s disease risk. Nat. Genet. 2019, 51, 404–413. [Google Scholar] [CrossRef] [PubMed]
- Lambert, J.-C.; Ibrahim-Verbaas, C.A.; Harold, D.; Naj, A.C.; Sims, R.; Bellenguez, C.; Jun, G.; DeStefano, A.L.; Bis, J.C.; Beecham, G.W.; et al. Meta-analysis of 74,046 individuals identifies 11 new susceptibility loci for Alzheimer’s disease. Nat. Genet. 2013, 45, 1452–1458. [Google Scholar] [CrossRef]
- Raulin, A.-C.; Doss, S.V.; Trottier, Z.A.; Ikezu, T.C.; Bu, G.; Liu, C.-C. ApoE in Alzheimer’s disease: Pathophysiology and therapeutic strategies. Mol. Neurodegener. 2022, 17, 72. [Google Scholar] [CrossRef]
- Serrano-Pozo, A.; Das, S.; Hyman, B.T. APOE and Alzheimer’s disease: Advances in genetics, pathophysiology, and therapeutic approaches. Lancet Neurol. 2021, 20, 68–80. [Google Scholar] [CrossRef]
- Mortensen, M.B.; Nordestgaard, B.G. Elevated LDL cholesterol and increased risk of myocardial infarction and atherosclerotic cardiovascular disease in individuals aged 70–100 years: A contemporary primary prevention cohort. Lancet 2020, 396, 1644–1652. [Google Scholar] [CrossRef] [PubMed]
- Ferguson, E.L.; Zimmerman, S.C.; Jiang, C.; Choi, M.; Swinnerton, K.; Choudhary, V.; Meyers, T.J.; Hoffmann, T.J.; Gilsanz, P.; Oni-Orisan, A. Low-and high-density lipoprotein cholesterol and dementia risk over 17 Years of follow-up among members of a large Health care plan. Neurology 2023, 101, e2172–e2184. [Google Scholar] [CrossRef]
- Lee, T.; Lee, H.; The Alzheimer’s Disease Neuroimaging, I. Identification of Disease-Related Genes That Are Common between Alzheimer’s and Cardiovascular Disease Using Blood Genome-Wide Transcriptome Analysis. Biomedicines 2021, 9, 1525. [Google Scholar] [CrossRef]
- Bellou, E.; Escott-Price, V. Are Alzheimer’s and coronary artery diseases genetically related to longevity? Front. Psychiatry 2023, 13, 1102347. [Google Scholar] [CrossRef] [PubMed]
- Gauthier, S.; Webster, C.; Servaes, S.; Morais, J.A.; Rosa-Neto, P. World Alzheimer Report 2022: Life after Diagnosis: Navigating Treatment, Care and Support; Alzheimer’s Disease International: London, UK, 2022. [Google Scholar]
- Luo, J.; Thomassen, J.Q.; Bellenguez, C.; Grenier-Boley, B.; De Rojas, I.; Castillo, A.; Parveen, K.; Küçükali, F.; Nicolas, A.; Peters, O. Genetic Associations Between Modifiable Risk Factors and Alzheimer Disease. JAMA Netw. Open 2023, 6, e2313734. [Google Scholar] [PubMed]
- Peters, R.; Xu, Y.; Antikainen, R.; Beckett, N.; Gussekloo, J.; Jagger, C.; Jukema, J.W.; Keinanen-Kiukaanniemi, S.; Rydén, L.; Skoog, I.; et al. Evaluation of High Cholesterol and Risk of Dementia and Cognitive Decline in Older Adults Using Individual Patient Meta-Analysis. Dement. Geriatr. Cogn. Disord. 2021, 50, 318–325. [Google Scholar] [CrossRef] [PubMed]
- Bulik-Sullivan, B.K.; Loh, P.-R.; Finucane, H.K.; Ripke, S.; Yang, J.; Schizophrenia Working Group of the Psychiatric Genomics Consortium; Patterson, N.; Daly, M.J.; Price, A.L.; Neale, B.M. LD Score regression distinguishes confounding from polygenicity in genome-wide association studies. Nat. Genet. 2015, 47, 291–295. [Google Scholar] [CrossRef] [PubMed]
- Davies, N.M.; Holmes, M.V.; Smith, G.D. Reading Mendelian randomisation studies: A guide, glossary, and checklist for clinicians. BMJ 2018, 362, k601. [Google Scholar] [CrossRef] [PubMed]
- Watanabe, K.; Taskesen, E.; Van Bochoven, A.; Posthuma, D. Functional mapping and annotation of genetic associations with FUMA. Nat. Commun. 2017, 8, 1826. [Google Scholar] [CrossRef] [PubMed]
- Tanha, H.M.; Sathyanarayanan, A.; The International Headache Genetics Consortium; Nyholt, D.R. Genetic overlap and causality between blood metabolites and migraine. Am. J. Hum. Genet. 2021, 108, 2086–2098. [Google Scholar] [CrossRef] [PubMed]
- Adewuyi, E.O.; O’Brien, E.K.; Porter, T.; Laws, S.M. Relationship of Cognition and Alzheimer’s Disease with Gastrointestinal Tract Disorders: A Large-Scale Genetic Overlap and Mendelian Randomisation Analysis. Int. J. Mol. Sci. 2022, 23, 16199. [Google Scholar] [CrossRef]
- Adewuyi, E.O.; Mehta, D.; International Endogene Consortium (IEC); 23andMe Research Team; Nyholt, D.R. Genetic overlap analysis of endometriosis and asthma identifies shared loci implicating sex hormones and thyroid signalling pathways. Hum. Reprod. 2022, 37, 366–383. [Google Scholar] [CrossRef]
- Adewuyi, E.O.; Mehta, D.; Sapkota, Y.; International Endogene Consortium; 23andMe Research Team; Auta, A.; Yoshihara, K.; Nyegaard, M.; Griffiths, L.R.; Montgomery, G.W.; et al. Genetic analysis of endometriosis and depression identifies shared loci and implicates causal links with gastric mucosa abnormality. Hum. Genet. 2021, 140, 529–552. [Google Scholar] [CrossRef]
- Adewuyi, E.O.; Sapkota, Y.; International Endogene Consortium (IEC); 23andMe Research Team; International Headache Genetics Consortium (IHGC); Auta, A.; Yoshihara, K.; Nyegaard, M.; Griffiths, L.R.; Montgomery, G.W.; et al. Shared molecular genetic mechanisms underlie endometriosis and migraine comorbidity. Genes 2020, 11, 268. [Google Scholar] [CrossRef] [PubMed]
- Yang, Y.; Zhao, H.; Boomsma, D.I.; Ligthart, L.; Belin, A.C.; Smith, G.D.; Esko, T.; Freilinger, T.M.; Hansen, T.F.; Ikram, M.A.; et al. Molecular genetic overlap between migraine and major depressive disorder. Eur. J. Hum. Genet. 2018, 26, 1202–1216. [Google Scholar] [CrossRef] [PubMed]
- Zhao, H.; Eising, E.; De Vries, B.; Vijfhuizen, L.S.; Consortium, I.H.G.; Anttila, V.; Winsvold, B.S.; Kurth, T.; Stefansson, H.; Kallela, M.; et al. Gene-based pleiotropy across migraine with aura and migraine without aura patient groups. Cephalalgia 2016, 36, 648–657. [Google Scholar] [CrossRef] [PubMed]
- Adewuyi, E.O.; Porter, T.; O’Brien, E.K.; Olaniru, O.; Verdile, G.; Laws, S.M. Genome-wide cross-disease analyses highlight causality and shared biological pathways of type 2 diabetes with gastrointestinal disorders. Commun. Biol. 2024, 7, 643. [Google Scholar] [CrossRef] [PubMed]
- Werme, J.; van der Sluis, S.; Posthuma, D.; de Leeuw, C.A. An integrated framework for local genetic correlation analysis. Nat. Genet. 2022, 54, 274–282. [Google Scholar] [CrossRef]
- Shin, S.-Y.; Fauman, E.B.; Petersen, A.-K.; Krumsiek, J.; Santos, R.; Huang, J.; Arnold, M.; Erte, I.; Forgetta, V.; Yang, T.-P. An atlas of genetic influences on human blood metabolites. Nat. Genet. 2014, 46, 543–550. [Google Scholar] [CrossRef] [PubMed]
- Draisma, H.H.; Pool, R.; Kobl, M.; Jansen, R.; Petersen, A.-K.; Vaarhorst, A.A.; Yet, I.; Haller, T.; Demirkan, A.; Esko, T. Genome-wide association study identifies novel genetic variants contributing to variation in blood metabolite levels. Nat. Commun. 2015, 6, 7208. [Google Scholar] [CrossRef]
- Zhou, W.; Zhao, Z.; Nielsen, J.B.; Fritsche, L.G.; LeFaive, J.; Gagliano Taliun, S.A.; Bi, W.; Gabrielsen, M.E.; Daly, M.J.; Neale, B.M.; et al. Scalable generalized linear mixed model for region-based association tests in large biobanks and cohorts. Nat. Genet. 2020, 52, 634–639. [Google Scholar] [CrossRef]
- Weber, C.J.; Carrillo, M.C.; Jagust, W.; Jack, C.R., Jr.; Shaw, L.M.; Trojanowski, J.Q.; Saykin, A.J.; Beckett, L.A.; Sur, C.; Rao, N.P.; et al. The Worldwide Alzheimer’s Disease Neuroimaging Initiative: ADNI-3 updates and global perspectives. Alzheimer’s Dement. Transl. Res. Clin. Interv. 2021, 7, e12226. [Google Scholar] [CrossRef]
- Allara, E.; Morani, G.; Carter, P.; Gkatzionis, A.; Zuber, V.; Foley, C.N.; Rees, J.M.B.; Mason, A.M.; Bell, S.; Gill, D.; et al. Genetic Determinants of Lipids and Cardiovascular Disease Outcomes. Circ. Genom. Precis. Med. 2019, 12, e002711. [Google Scholar] [CrossRef]
- Arnett, D.K.; Blumenthal, R.S.; Albert, M.A.; Buroker, A.B.; Goldberger, Z.D.; Hahn, E.J.; Himmelfarb, C.D.; Khera, A.; Lloyd-Jones, D.; McEvoy, J.W. 2019 ACC/AHA guideline on the primary prevention of cardiovascular disease: A report of the American College of Cardiology/American Heart Association Task Force on Clinical Practice Guidelines. Circulation 2019, 140, e596–e646. [Google Scholar] [CrossRef] [PubMed]
- Razavi, A.C.; Mehta, A.; Jain, V.; Patel, P.; Liu, C.; Patel, N.; Eisenberg, S.; Vaccarino, V.; Isiadinso, I.; Sperling, L.S.; et al. High-Density Lipoprotein Cholesterol in Atherosclerotic Cardiovascular Disease Risk Assessment: Exploring and Explaining the “U”-Shaped Curve. Curr. Cardiol. Rep. 2023, 25, 1725–1733. [Google Scholar] [CrossRef] [PubMed]
- Borodzicz-Jażdżyk, S.; Jażdżyk, P.; Łysik, W.; Cudnoch-Jȩdrzejewska, A.; Czarzasta, K. Sphingolipid metabolism and signaling in cardiovascular diseases. Front. Cardiovasc. Med. 2022, 9, 915961. [Google Scholar] [CrossRef]
- Manzo, O.L.; Nour, J.; Sasset, L.; Marino, A.; Rubinelli, L.; Palikhe, S.; Smimmo, M.; Hu, Y.; Bucci, M.R.; Borczuk, A.; et al. Rewiring Endothelial Sphingolipid Metabolism to Favor S1P Over Ceramide Protects From Coronary Atherosclerosis. Circ. Res. 2024, 134, 990–1005. [Google Scholar] [CrossRef]
- Kovilakath, A.; Jamil, M.; Cowart, L.A. Sphingolipids in the heart: From cradle to grave. Front. Endocrinol. 2020, 11, 652. [Google Scholar] [CrossRef]
- Zhu, Z.; Lin, Y.; Li, X.; Driver, J.A.; Liang, L. Shared genetic architecture between metabolic traits and Alzheimer’s disease: A large-scale genome-wide cross-trait analysis. Hum. Genet. 2019, 138, 271–285. [Google Scholar] [CrossRef]
- Shi, H.; Mancuso, N.; Spendlove, S.; Pasaniuc, B. Local genetic correlation gives insights into the shared genetic architecture of complex traits. Am. J. Hum. Genet. 2017, 101, 737–751. [Google Scholar] [CrossRef]
- Gerring, Z.F.; Thorp, J.G.; Gamazon, E.R.; Derks, E.M. A Local Genetic Correlation Analysis Provides Biological Insights Into the Shared Genetic Architecture of Psychiatric and Substance Use Phenotypes. Biol. Psychiatry 2022, 92, 583–591. [Google Scholar] [CrossRef] [PubMed]
- Yin, F. Lipid metabolism and Alzheimer’s disease: Clinical evidence, mechanistic link and therapeutic promise. FEBS J. 2023, 290, 1420–1453. [Google Scholar] [CrossRef]
- Escott-Price, V.; Hardy, J. Genome-wide association studies for Alzheimer’s disease: Bigger is not always better. Brain Commun. 2022, 4, fcac125. [Google Scholar] [CrossRef]
- Akosile, W.; Adewuyi, E. Genetic correlation and causality assessment between post-traumatic stress disorder and coronary artery disease-related traits. Gene 2022, 842, 146802. [Google Scholar] [CrossRef] [PubMed]
- Yuan, S.; Chen, J.; Ruan, X.; Sun, Y.; Zhang, K.; Wang, X.; Li, X.; Gill, D.; Burgess, S.; Giovannucci, E.; et al. Smoking, alcohol consumption, and 24 gastrointestinal diseases: Mendelian randomization analysis. eLife 2023, 12, e84051. [Google Scholar] [CrossRef]
- Bowden, J.; Davey Smith, G.; Burgess, S. Mendelian randomization with invalid instruments: Effect estimation and bias detection through Egger regression. Int. J. Epidemiol. 2015, 44, 512–525. [Google Scholar] [CrossRef]
- Bowden, J.; Davey Smith, G.; Haycock, P.C.; Burgess, S. Consistent estimation in Mendelian randomization with some invalid instruments using a weighted median estimator. Genet. Epidemiol. 2016, 40, 304–314. [Google Scholar] [CrossRef] [PubMed]
- Verbanck, M.; Chen, C.-Y.; Neale, B.; Do, R. Detection of widespread horizontal pleiotropy in causal relationships inferred from Mendelian randomization between complex traits and diseases. Nat. Genet. 2018, 50, 693–698. [Google Scholar] [CrossRef] [PubMed]
- Maier, R.; Visscher, P.; Robinson, M.R.; Wray, N. Embracing polygenicity: A review of methods and tools for psychiatric genetics research. Psychol. Med. 2018, 48, 1055–1067. [Google Scholar] [CrossRef]
- Bulik-Sullivan, B.; Finucane, H.K.; Anttila, V.; Gusev, A.; Day, F.R.; Loh, P.-R.; Duncan, L.; Perry, J.R.B.; Patterson, N.; Robinson, E.B.; et al. An atlas of genetic correlations across human diseases and traits. Nat. Genet. 2015, 47, 1236–1241. [Google Scholar] [CrossRef]

 ; LDL:
; LDL:  ; Triglycerides:
; Triglycerides:  ; Total cholesterol:
; Total cholesterol:  ; Angina pectoris:
; Angina pectoris:  ; Cardiac dysrhythmias:
; Cardiac dysrhythmias:  ; Coronary arteriosclerosis:
; Coronary arteriosclerosis:  ; Ischemic heart disease:
; Ischemic heart disease:  ; Myocardial infarction:
; Myocardial infarction:  . Representative colour for chromosome—Chromosome 6: ORANGE; Chromosome 8: GREEN; Chromosome 17: BLUE; Chromosome 19: RED. Representative pattern fill for locus position—962:
. Representative colour for chromosome—Chromosome 6: ORANGE; Chromosome 8: GREEN; Chromosome 17: BLUE; Chromosome 19: RED. Representative pattern fill for locus position—962:  ; 963:
; 963:  ; 964:
; 964:  ; 965:
; 965:  ; 966:
; 966:  ; 1351:
; 1351:  ; 2209:
; 2209:  ; 2351:
; 2351:  .
; LDL: ; Triglycerides: ; Total cholesterol: ; Angina pectoris: ; Cardiac dysrhythmias: ; Coronary arteriosclerosis: ; Ischemic heart disease: ; Myocardial infarction: . Representative colour for chromosome—Chromosome 6: ORANGE; Chromosome 8: GREEN; Chromosome 17: BLUE; Chromosome 19: RED. Representative pattern fill for locus position—962: ; 963: ; 964: ; 965: ; 966: ; 1351: ; 2209: ; 2351: .
.
; LDL: ; Triglycerides: ; Total cholesterol: ; Angina pectoris: ; Cardiac dysrhythmias: ; Coronary arteriosclerosis: ; Ischemic heart disease: ; Myocardial infarction: . Representative colour for chromosome—Chromosome 6: ORANGE; Chromosome 8: GREEN; Chromosome 17: BLUE; Chromosome 19: RED. Representative pattern fill for locus position—962: ; 963: ; 964: ; 965: ; 966: ; 1351: ; 2209: ; 2351: .


{kind=link}
{kind=link}
{kind=link}
{kind=link}
| GWAS Summary Statistics | Cases | Controls | Sample Size | Ancestry | Phenotype Source/Definition |
|---|---|---|---|---|---|
| AD | European | ||||
| Main (Jansen et al. [28]) | 71,880 | 383,378 | 455,258 | Clinically diagnosed and UKB AD-by-proxy2 | |
| Validation (Lambert et al. [29]) * | 17,008 | 37,154 | 54,162 | Data from the EADI, GERAD, ADGC, and CHARGE study | |
| LIPID | European | ||||
| Sphingolipids: | |||||
| Palmitoyl sphingomyelin (Shin et al. [51]) | 7814 | Data from the TwinsUK and KORA study | |||
| 154 SM C16:1 sphingomyelin (Draisma et al. [52]) | 7428 | Data from dataverse | |||
| 156 SM C18:1 sphingomyelin (Draisma et al. [52]) | 7428 | Data from dataverse | |||
| Glycerophospholipids: | |||||
| Beta-glycerophosphoric acid (Shin et al. [51]) | 5912 | Data from the TwinsUK and KORA study | |||
| Lysophosphatidylinositol (Shin et al. [51]) | 7797 | Data from the TwinsUK and KORA study | |||
| Fatty Acyls: | |||||
| Palmitic acid (Shin et al. [51]) | 7800 | Data from the TwinsUK and KORA study | |||
| Stearic acid (Shin et al. [51]) | 7803 | Data from the TwinsUK and KORA study | |||
| Fatty Acyls [lipids or lipid-like molecules]: | |||||
| Caprylic acid (Shin et al. [51]) | 7802 | Data from the TwinsUK and KORA study | |||
| Organic compounds known as medium-chain fatty acids: | |||||
| Dodecanoic acid (Shin et al. [51]), (also known as lauric acid) | 7793 | Data from the TwinsUK and KORA study | |||
| Lipoproteins: | |||||
| HDL (GLGC [12]) | 188,577 | Data from the GLGC | |||
| LDL (GLGC [12]) | 188,577 | Data from the GLGC | |||
| Neutral lipids: | |||||
| TG (GLGC [12]) | 188,577 | Data from the GLGC | |||
| Steroids and steroid derivatives: | |||||
| TC (GLGC [12]) | 188,577 | Data from the GLGC | |||
| CAD trait | European | ||||
| Angina pectoris Phecode 411.3 (Lee Lab [53]) | 16,175 | 377,103 | 393,278 | Full European data subset from the Lee Lab | |
| Cardiac dysrhythmias Phecode 427 (Lee Lab [53]) | 24,681 | 380,919 | 405,600 | Full European data subset from the Lee Lab | |
| Coronary atherosclerosis Phecode 411.4 (Lee Lab [53]) | 20,023 | 377,103 | 397,126 | Full European data subset from the Lee Lab | |
| Ischemic heart disease Phecode 411 (Lee Lab [53]) | 31,355 | 377,103 | 408,458 | Full European data subset from the Lee Lab | |
| Myocardial infarction Phecode 411.2 (Lee Lab [53]) | 11,703 | 377,103 | 388,806 | Full European data subset from the Lee Lab | |
| Non-specific chest pain Phecode 418 (Lee Lab [53]) | 31,429 | 377,532 | 408,961 | Full European data subset from the Lee Lab | |
| CARDIoGRAMplusC4D (CGCC [12]) | 22,233 | 64,762 | 86,995 | Data from the CGCC |
| AD | Lipids | rg | Se | p |
| Palmitoyl sphingomyelin | −0.03 | 4.22 × 10−2 | 4.96 × 10−1 | |
| 154 SM C16:1 sphingomyelin | −0.02 | 1.07 × 10−1 | 8.78 × 10−1 | |
| 156 SM C18:1 sphingomyelin | 0.14 | 1.37 × 10−1 | 3.10 × 10−1 | |
| beta-Glycerophosphoric acid | 0.05 | 5.69 × 10−2 | 4.28 × 10−1 | |
| Lysophosphatidylinositol | 0.04 | 5.48 × 10−2 | 5.11 × 10−1 | |
| Palmitic acid | 0.00 | 5.08 × 10−2 | 9.96 × 10−1 | |
| AD | Stearic acid | −0.03 | 4.34 × 10−2 | 5.49 × 10−1 |
| Caprylic acid | 0.00 | 4.56 × 10−2 | 9.51 × 10−1 | |
| Dodecanoic acid | −0.01 | 3.94 × 10−2 | 7.20 × 10−1 | |
| HDL | −0.05 | 3.85 × 10−2 | 1.81 × 10−1 | |
| LDL | 0.13 | 7.10 × 10−2 | 6.49 × 10−2 | |
| TG | 0.09 | 3.64 × 10−2 | 1.09 × 10−2 | |
| TC | 0.13 | 7.01 × 10−2 | 5.48 × 10−2 | |
| AD | CAD traits | Rg | Se | p |
| Angina pectoris | 0.21 | 3.55 × 10−2 | 5.88 × 10−9 | |
| Cardiac dysrhythmias | 0.14 | 3.78 × 10−2 | 3.49 × 10−4 | |
| Coronary arteriosclerosis | 0.17 | 2.96 × 10−2 | 2.26 × 10−8 | |
| AD | Ischemic heart disease | 0.20 | 3.13 × 10−2 | 1.39 × 10−10 |
| Myocardial infarction | 0.17 | 3.84 × 10−2 | 1.03 × 10−5 | |
| Non-specific chest pain | 0.22 | 3.91 × 10−2 | 2.06 × 10−8 | |
| CAD | 0.15 | 4.25 × 10−2 | 3.74 × 10−4 |
| CAD Trait | Lipids Trait | rg | Se | p |
|---|---|---|---|---|
| Angina pectoris | HDL | −0.39 | 4.77 × 10−2 | 1.55 × 10−16 |
| LDL | 0.28 | 3.66 × 10−2 | 5.68 × 10−14 | |
| TG | 0.41 | 5.73 × 10−2 | 8.92 × 10−13 | |
| TC | 0.23 | 3.44 × 10−2 | 1.07 × 10−11 | |
| Cardiac dysrhythmias | HDL | −0.18 | 3.60 × 10−2 | 2.93 × 10−7 |
| TG | 0.14 | 4.00 × 10−2 | 3.70 × 10−4 | |
| Coronary arteriosclerosis | HDL | −0.36 | 4.44 × 10−2 | 8.72 × 10−16 |
| LDL | 0.3 | 3.78 × 10−2 | 4.93 × 10−15 | |
| TG | 0.37 | 4.62 × 10−2 | 2.11 × 10−15 | |
| TC | 0.25 | 3.59 × 10−2 | 2.10 × 10−12 | |
| 154 SM C16:1 sphingomyelin | −0.31 | 1.23 × 10−1 | 1.18 × 10−2 | |
| Ischemic heart disease | HDL | −0.38 | 4.66 × 10−2 | 2.65 × 10−16 |
| LDL | 0.28 | 3.55 × 10−2 | 3.08 × 10−15 | |
| TG | 0.4 | 5.07 × 10−2 | 2.99 × 10−15 | |
| TC | 0.24 | 3.28 × 10−2 | 1.73 × 10−13 | |
| Myocardial infarction | HDL | −0.37 | 5.27 × 10−2 | 1.25 × 10−12 |
| LDL | 0.29 | 3.82 × 10−2 | 6.62 × 10−14 | |
| TG | 0.41 | 5.65 × 10−2 | 2.82 × 10−13 | |
| TC | 0.26 | 3.59 × 10−2 | 3.98 × 10−13 | |
| Non-specific chest pain | HDL | −0.32 | 4.40 × 10−2 | 5.38 × 10−13 |
| LDL | 0.14 | 3.35 × 10−2 | 2.86 × 10−5 | |
| TG | 0.31 | 5.29 × 10−2 | 7.10 × 10−9 | |
| TC | 0.1 | 3.11 × 10−2 | 1.96 × 10−3 | |
| CAD | HDL | −0.37 | 4.40 × 10−2 | 6.52 × 10−17 |
| LDL | 0.39 | 4.52 × 10−2 | 1.48 × 10−17 | |
| TG | 0.42 | 4.39 × 10−2 | 1.41 × 10−21 | |
| TC | 0.35 | 4.24 × 10−2 | 2.67 × 10−16 |
| Discovery Set | Target Set | Number of Genes | Proportion of Gene Overlap | Binomial Test | |||||
|---|---|---|---|---|---|---|---|---|---|
| Lipids and CAD Traits | Total Number of Genes in the Discovery Set (Lipid or CAD Trait) | Number of Genes in the Discovery Set Pgene < 0.05 | AD | Total Number of Genes in the Target Set (AD) | Number of Genes in the Target Set at Pgene < 0.05 | Overlapping the Discovery and the Target Sets at Pgene < 0.05 | Expected (%) | Observed (%) | p Value |
| * HDL | 17,683 | 1880 | AD | 17,683 | 1768 | 294 | 10.6 | 16.6 | 9.84 × 10−15 |
| LDL | 17,669 | 1766 | AD | 17,669 | 1769 | 267 | 10.0 | 15.1 | 1.28 × 10−11 |
| Triglycerides | 17,671 | 1743 | AD | 17,671 | 1769 | 273 | 9.9 | 15.4 | 2.24 × 10−13 |
| Total cholesterol | 17,683 | 1988 | AD | 17,683 | 1767 | 320 | 11.2 | 18.1 | 2.20 × 10−16 |
| Angina pectoris | 18,960 | 2175 | AD | 18,960 | 1843 | 260 | 11.5 | 14.1 | 3.65 × 10−4 |
| Cardiac dysrhythmias | 18,960 | 1776 | AD | 18,960 | 1843 | 212 | 9.4 | 11.5 | 1.48 × 10−3 |
| Coronary arteriosclerosis | 18,960 | 2524 | AD | 18,960 | 1843 | 333 | 13.3 | 18.1 | 4.73 × 10−9 |
| Ischemic heart disease | 18,960 | 2710 | AD | 18,960 | 1843 | 315 | 14.3 | 17.1 | 4.60 × 10−4 |
| Myocardial infarction | 18,960 | 1995 | AD | 18,960 | 1843 | 244 | 10.5 | 13.2 | 1.18 × 10−4 |
| Non-specific chest pain | 18,960 | 1943 | AD | 18,960 | 1843 | 212 | 10.2 | 11.5 | 3.69 × 10−2 |
| CAD | 17,735 | 1601 | AD | 17,735 | 1781 | 201 | 9.0 | 11.3 | 6.26 × 10−4 |
| Genes | Chr | START (hg19) | STOP (hg19) | AD, Lipids, and CAD Traits |
|---|---|---|---|---|
| GWS genes (sentinel) overlapping AD and two or more CAD or lipid traits | ||||
| APOC1 | 19 | 45,417,504 | 45,422,606 | AD, CA, IHD, MI |
| APOC4 | 19 | 45,445,495 | 45,452,820 | AD, HDL, LDL, TC |
| APOC4-APOC2 | 19 | 45,445,495 | 45,452,822 | AD, HDL, TC |
| APOE | 19 | 45,409,011 | 45,412,650 | AD, AP, CA, HDL, IHD, LDL, MI, TC, TG |
| BCL3 | 19 | 45,250,962 | 45,263,301 | AD, LDL, TC |
| CBLC | 19 | 45,281,126 | 45,303,891 | AD, LDL, TC |
| CEACAM19 | 19 | 45,165,545 | 45,187,631 | AD, LDL, TC |
| IGSF23 | 19 | 45,116,940 | 45,140,081 | AD, LDL, TC |
| NKPD1 | 19 | 45,653,008 | 45,663,408 | AD, LDL, TC |
| PVR | 19 | 45,147,098 | 45,166,850 | AD, LDL, TC |
| PVRL2 | 19 | 45,349,432 | 45,392,485 | AD, CA, IHD, LDL, TG, TC |
| TOMM40 | 19 | 45,393,826 | 45,406,946 | AD, CA, HDL, IHD, LDL, MI, TC, TG |
| ZNF652 | 17 | 47,366,568 | 47,439,835 | AD, AP, CA, IHD |
| Genes reaching GWS in the FCP analysis overlapping AD and two or more CAD or lipid traits | ||||
| ACMSD | 2 | 135,596,117 | 135,659,604 | AD, LDL, TC |
| ICA1L | 2 | 203,640,690 | 203,736,708 | AD, AP, LDL, TC |
| WDR12 | 2 | 203,739,505 | 203,879,521 | AD, AP, LDL, TC |
| CARF | 2 | 203,776,937 | 203,851,786 | AD, AP, LDL, TC |
| PRRC2A | 6 | 31,588,497 | 31,605,548 | AD, CA, IHD |
| BAG6 | 6 | 31,606,805 | 31,620,482 | AD, CA, IHD, NSCP |
| C6orf10 | 6 | 32,256,303 | 32,339,684 | AD, CA, IHD, TC |
| HLA-DRA | 6 | 32,407,619 | 32,412,823 | AD, AP, CA, IHD, MI, HDL, TG |
| HLA-DQB1 | 6 | 32,627,244 | 32,636,160 | AD, AP, CA, LDL |
| TMEM106B | 7 | 12,250,867 | 12,282,993 | AD, AP, IHD |
| NDUFAF6 | 8 | 95,907,995 | 96,128,683 | AD, CA, IHD |
| TRIB1 | 8 | 126,442,563 | 126,450,647 | AD, CA, HDL |
| DOC2A | 16 | 30,016,830 | 30,034,591 | AD, CD, IHD |
| ZNF668 | 16 | 31,072,164 | 31,085,641 | AD, LDL, TC |
| PRSS8 | 16 | 31,142,756 | 31,147,083 | AD, LDL, TC |
| PLCG2 | 16 | 81,772,702 | 81,991,899 | AD, AP, CA |
| RP11-81K2.1 | 17 | 47,448,102 | 47,554,350 | AD, MI, NSCP |
| PHB | 17 | 47,481,414 | 47,492,246 | AD, CD, MI, NSCP |
| APOC2 | 19 | 45,449,243 | 45,452,822 | AD, LDL, TG |
| RSPH6A | 19 | 46,298,968 | 46,318,577 | AD, LDL, TC |
| Outcome | Exposure | nIV | IVW | Weighted Median | MR-Egger | * Ppleiotropy | # Pheterogeneity | MR-PRESSO | ||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| OR (95% CI) | p Value | OR (95% CI) | p Value | OR (95% CI) | p Value | RAW OR (95% CI) | p Value | Corrected OR (95% CI) | p Value | |||||
| HDL | 3 | 1.02 (0.94–1.12) | 5.91 × 10−1 | 1.04 (0.94–1.14) | 4.56 × 10−1 | 1.15 (0.79–1.67) | 6.01 × 10−1 | 6.49 × 10−1 | 7.24 × 10−1 | - | - | - | - | |
| LDL | 62 | 1.00 (0.99–1.02) | 4.19 × 10−1 | 1.00 (0.99–1.02) | 7.19 × 10−1 | 1.00 (0.99–1.02) | 6.19 × 10−1 | 9.93 × 10−1 | 4.15 × 10−1 | 1.00 (0.99–1.02) | 4.23 × 10−1 | NA | NA | |
| Triglycerides | 43 | 1.01 (0.99–1.02) | 2.13 × 10−1 | 1.00 (0.99–1.03) | 5.74 × 10−1 | 1.00 (0.98–1.02) | 9.33 × 10−1 | 2.31 × 10−1 | 5.84 × 10−1 | 1.01 (1.00–1.02) | 2.06 × 10−1 | NA | NA | |
| Total cholesterol | 66 | 1.00 (0.99–1.02) | 4.65 × 10−1 | 1.00 (0.98–1.02) | 7.41 × 10−1 | 1.00 (0.98–1.02) | 9.50 × 10−1 | 6.93 × 10−1 | 4.92 × 10−1 | 1.00 (0.99–1.02) | 4.66 × 10−1 | NA | NA | |
| Angina pectoris | 15 | 1.00 (0.99–1.02) | 4.70 × 10−1 | 1.01 (0.99–1.03) | 2.53 × 10−1 | 1.01 (0.98–1.04) | 5.09 × 10−1 | 7.05 × 10−1 | 1.44 × 10−1 | 1.00 (0.99–1.02) | 4.81 × 10−1 | NA | NA | |
| AD | Cardiac dysrhythmias | 24 | 1.00 (0.99–1.02) | 4.15 × 10−1 | 1.01 (0.99–1.02) | 3.95 × 10−1 | 1.02 (1.00–1.05) | 1.13 × 10−1 | 1.64 × 10−1 | 9.40 × 10−1 | 1.00 (1.00–1.01) | 2.99 × 10−1 | NA | NA |
| Coronary arteriosclerosis | 37 | 1.00 (1.00–1.01) | 4.20 × 10−1 | 1.01 (0.99–1.02) | 3.53 × 10−1 | 1.02 (1.00–1.04) | 3.06 × 10−2 | 4.16 × 10−2 | 3.88 × 10−1 | 1.00 (1.00–1.01) | 4.25 × 10−1 | NA | NA | |
| Ischemic heart disease | 33 | 1.01 (1.00–1.02) | 7.62 × 10−2 | 1.01 (0.99–1.03) | 2.15 × 10−1 | 1.01 (0.98–1.03) | 5.06 × 10−1 | 8.57 × 10−1 | 2.75 × 10−1 | 1.01 (1.00–1.02) | 8.58 × 10−2 | NA | NA | |
| Myocardial infarction | 14 | 1.00 (0.99–1.02) | 5.97 × 10−1 | 0.99 (0.98–1.01) | 4.89 × 10−1 | 1.02 (0.99–1.04) | 2.68 × 10−1 | 3.22 × 10−1 | 2.24 × 10−1 | 1.00 (0.99–1.02) | 6.06 × 10−1 | NA | NA | |
| Non-specific chest pain | 1 | - | - | - | - | - | - | - | - | - | - | - | - | |
| CAD | 13 | 1.00 (0.99–1.01) | 9.41 × 10−1 | 1.00 (0.98–1.01) | 7.21 × 10−1 | 0.99 (0.95–1.03) | 5.71 × 10−1 | 5.70 × 10−1 | 9.27 × 10−1 | 1.00 (0.99–1.01) | 9.06 × 10−1 | NA | NA | |
| HDL | 2 | 0.77 (0.30–2.00) | 5.86 × 10−1 | - | - | - | - | - | 1.26 × 10−1 | - | - | - | - | |
| LDL | 10 | 1.00 (0.79–1.26) | 9.81 × 10−1 | 1.14 (0.82–1.57) | 4.33 × 10−1 | 1.03 (0.33–3.22) | 9.56 × 10−1 | 9.51 × 10−1 | 6.00 × 10−1 | 1.00 (0.80–1.24) | 9.79 × 10−1 | NA | NA | |
| Triglycerides | 10 | 0.90 (0.69–1.18) | 4.42 × 10−1 | 0.75 (0.54–1.04) | 8.74 × 10−2 | 1.93 (0.56–6.72) | 3.31 × 10−1 | 2.55 × 10−1 | 1.19 × 10−1 | 0.90 (0.69–1.18) | 4.62 × 10−1 | NA | NA | |
| Total cholesterol | 10 | 0.94 (0.74–1.18) | 5.85 × 10−1 | 1.09 (0.80–1.47) | 5.97 × 10−1 | 0.89 (0.29–2.77) | 8.46 × 10−1 | 9.30 × 10−1 | 4.99 × 10−1 | 0.94 (0.75–1.17) | 5.85 × 10−1 | NA | NA | |
| Angina pectoris | 23 | 0.85 (0.65–1.11) | 2.25 × 10−1 | 1.17 (0.81–1.67) | 4.07 × 10−1 | 1.43 (0.93–2.20) | 1.14 × 10−1 | 8.07 × 10−3 | 2.66 × 10−1 | 0.85 (0.65–1.11) | 2.37 × 10−1 | NA | NA | |
| Cardiac dysrhythmias | AD | 27 | 0.96 (0.84–1.10) | 5.50 × 10−1 | 1.09 (0.90–1.31) | 3.85 × 10−1 | 1.11 (0.93–1.32) | 2.76 × 10−1 | 2.62 × 10−2 | 8.29 × 10−1 | 0.96 (0.86–1.08) | 4.92 × 10−1 | NA | NA |
| Coronary arteriosclerosis | 22 | 0.96 (0.75–1.22) | 7.29 × 10−1 | 1.32 (0.96–1.82) | 9.06 × 10−2 | 1.23 (0.82–1.85) | 3.36 × 10−1 | 1.59 × 10−1 | 2.96 × 10−1 | 0.96 (0.75–1.22) | 7.32 × 10−1 | NA | NA | |
| Ischemic heart disease | 2 | 1.03 (0.40–2.61) | 9.57 × 10−1 | - | - | - | - | - | 3.06 × 10−1 | - | - | - | - | |
| Myocardial infarction | 24 | 1.09 (0.82–1.44) | 5.61 × 10−1 | 1.19 (0.78–1.79) | 4.21 × 10−1 | 1.23 (0.75–2.01) | 4.27 × 10−1 | 5.67 × 10−1 | 8.77 × 10−1 | 1.09 (0.86–1.37) | 4.85 × 10−1 | NA | NA | |
| Non-specific chest pain | 25 | 0.94 (0.81–1.89) | 3.87 × 10−1 | 0.98 (0.80–1.21) | 8.61 × 10−1 | 0.92 (0.74–1.15) | 4.60 × 10−1 | 8.20 × 10−1 | 8.16 × 10−1 | 0.94 (0.82–1.06) | 3.24 × 10−1 | NA | NA | |
| CAD | 9 | 1.50 (0.73–3.10) | 2.72 × 10−1 | 2.14 (0.83–5.47) | 1.11 × 10−1 | 2.03 (0.08–53.19) | 6.82 × 10−1 | 8.57 × 10−1 | 6.12 × 10−1 | 1.50 (0.79–2.86) | 2.51 × 10−1 | NA | NA |
| Locus | Chr | Start | Stop | SNP (n) | Phenotype1 | Phenotype2 | RHO | R2 | p | Mean.RHO |
|---|---|---|---|---|---|---|---|---|---|---|
| 2351 | 19 | 45,040,933 | 45,893,307 | 375 | AD | HDL | −0.29 | 0.09 | 3.75 × 10−10 | −0.29 |
| 962 | 6 | 32,208,902 | 32,454,577 | 538 | AD | LDL | 0.64 | 0.41 | 1.69 × 10−4 | |
| 964 | 6 | 32,539,568 | 32,586,784 | 26 | AD | LDL | 0.34 | 0.11 | 1.14 × 10−3 | |
| 966 | 6 | 32,629,240 | 32,682,213 | 161 | AD | LDL | 0.76 | 0.58 | 1.72 × 10−5 | |
| 2351 | 19 | 45,040,933 | 45,893,307 | 369 | AD | LDL | 0.34 | 0.11 | 2.21 × 10−100 | 0.52 |
| 2351 | 19 | 45,040,933 | 45,893,307 | 371 | AD | Triglycerides | 0.26 | 0.07 | 1.02 × 10−4 | 0.26 |
| 964 | 6 | 32,539,568 | 32,586,784 | 26 | AD | Total cholesterol | 0.41 | 0.17 | 1.22 × 10−3 | |
| 966 | 6 | 32,629,240 | 32,682,213 | 161 | AD | Total cholesterol | 0.51 | 0.26 | 3.85 × 10−4 | |
| 1351 | 8 | 125,453,323 | 126,766,827 | 1102 | AD | Total cholesterol | 0.30 | 0.09 | 1.04 × 10−79 | |
| 2351 | 19 | 45,040,933 | 45,893,307 | 373 | AD | Total cholesterol | 0.38 | 0.14 | 3.86 × 10−9 | 0.40 |
| 965 | 6 | 32,586,785 | 32,629,239 | 651 | AD | Angina pectoris | 0.34 | 0.12 | 2.69 × 10−4 | |
| 2351 | 19 | 45,040,933 | 45,893,307 | 2620 | AD | Angina pectoris | 0.37 | 0.14 | 1.29 × 10−10 | 0.35 |
| 963 | 6 | 32,454,578 | 32,539,567 | 89 | AD | Cardiac dysrhythmias | −0.38 | 0.14 | 7.25 × 10−6 | −0.38 |
| 2351 | 19 | 45,040,933 | 45,893,307 | 2620 | AD | Coronary arteriosclerosis | 0.53 | 0.28 | 9.80 × 10−28 | 0.53 |
| 2209 | 17 | 45,883,902 | 47,516,224 | 4658 | AD | Ischemic heart disease | 0.33 | 0.11 | 1.28 × 10−3 | |
| 2351 | 19 | 45,040,933 | 45,893,307 | 2620 | AD | Ischemic heart disease | 0.44 | 0.19 | 6.70 × 10−17 | 0.38 |
| 964 | 6 | 32,539,568 | 32,586,784 | 496 | AD | Myocardial infarction | 0.41 | 0.16 | 8.95 × 10−4 | |
| 2351 | 19 | 45,040,933 | 45,893,307 | 2620 | AD | Myocardial infarction | 0.45 | 0.20 | 3.37 × 10−14 | 0.43 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Kirby, A.; Porter, T.; Adewuyi, E.O.; Laws, S.M. Investigating Genetic Overlap between Alzheimer’s Disease, Lipids, and Coronary Artery Disease: A Large-Scale Genome-Wide Cross Trait Analysis. Int. J. Mol. Sci. 2024, 25, 8814. https://doi.org/10.3390/ijms25168814
Kirby A, Porter T, Adewuyi EO, Laws SM. Investigating Genetic Overlap between Alzheimer’s Disease, Lipids, and Coronary Artery Disease: A Large-Scale Genome-Wide Cross Trait Analysis. International Journal of Molecular Sciences. 2024; 25(16):8814. https://doi.org/10.3390/ijms25168814
Chicago/Turabian StyleKirby, Artika, Tenielle Porter, Emmanuel O. Adewuyi, and Simon M. Laws. 2024. "Investigating Genetic Overlap between Alzheimer’s Disease, Lipids, and Coronary Artery Disease: A Large-Scale Genome-Wide Cross Trait Analysis" International Journal of Molecular Sciences 25, no. 16: 8814. https://doi.org/10.3390/ijms25168814
APA StyleKirby, A., Porter, T., Adewuyi, E. O., & Laws, S. M. (2024). Investigating Genetic Overlap between Alzheimer’s Disease, Lipids, and Coronary Artery Disease: A Large-Scale Genome-Wide Cross Trait Analysis. International Journal of Molecular Sciences, 25(16), 8814. https://doi.org/10.3390/ijms25168814