
Nanoparticles-Delivered Circular RNA Strategy as a Novel Antitumor Approach
 , , , and
, , , and
Abstract
:1. Introduction
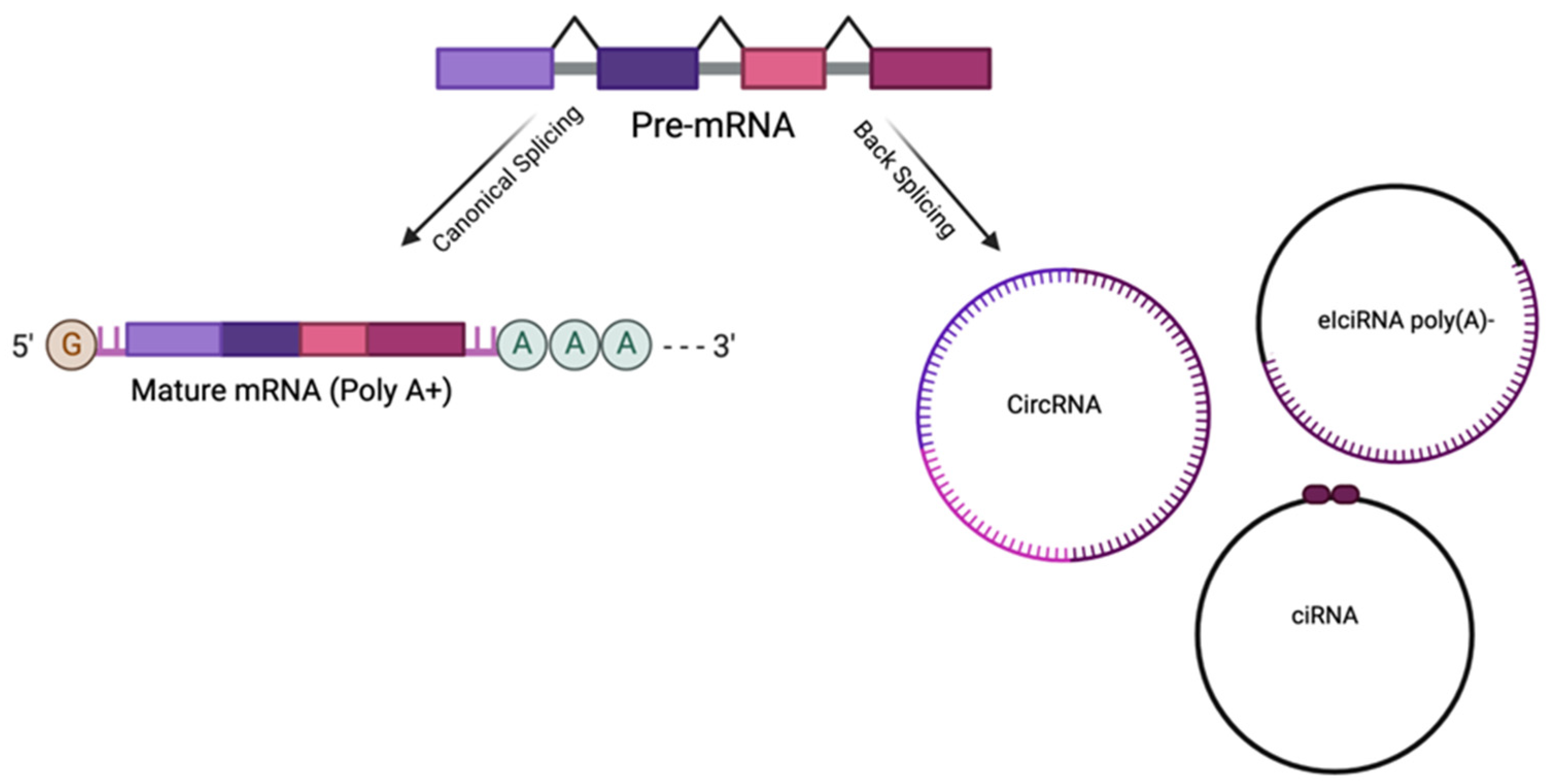
2. Circular RNAs
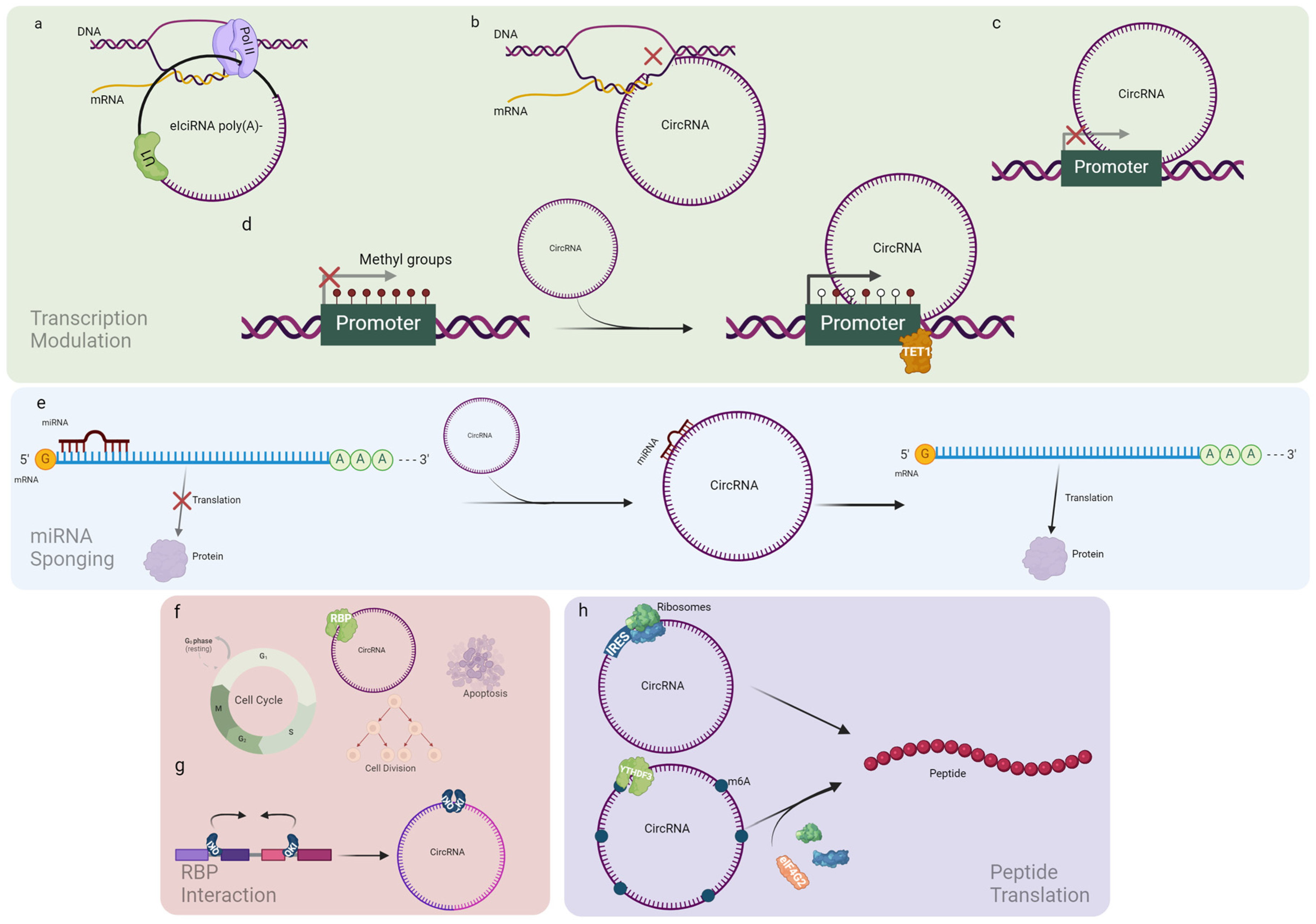
2.1. CircRNA Biological Role
2.1.1. Transcription Modulation
2.1.2. miRNA Sponging
2.1.3. RBP Interaction
2.1.4. Peptide Translation
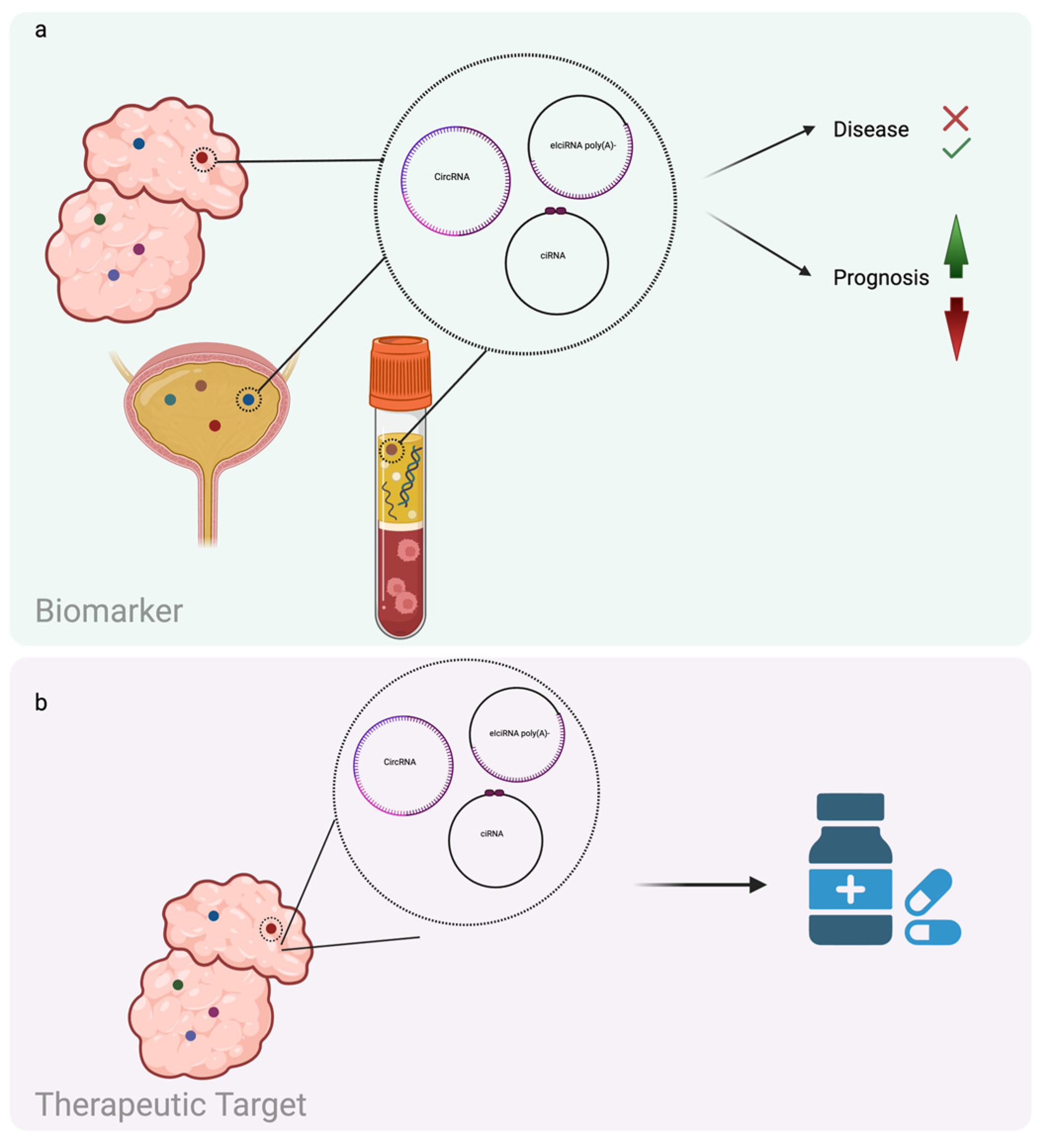
2.2. CircRNAs and Cancer
3. Nanomedicine
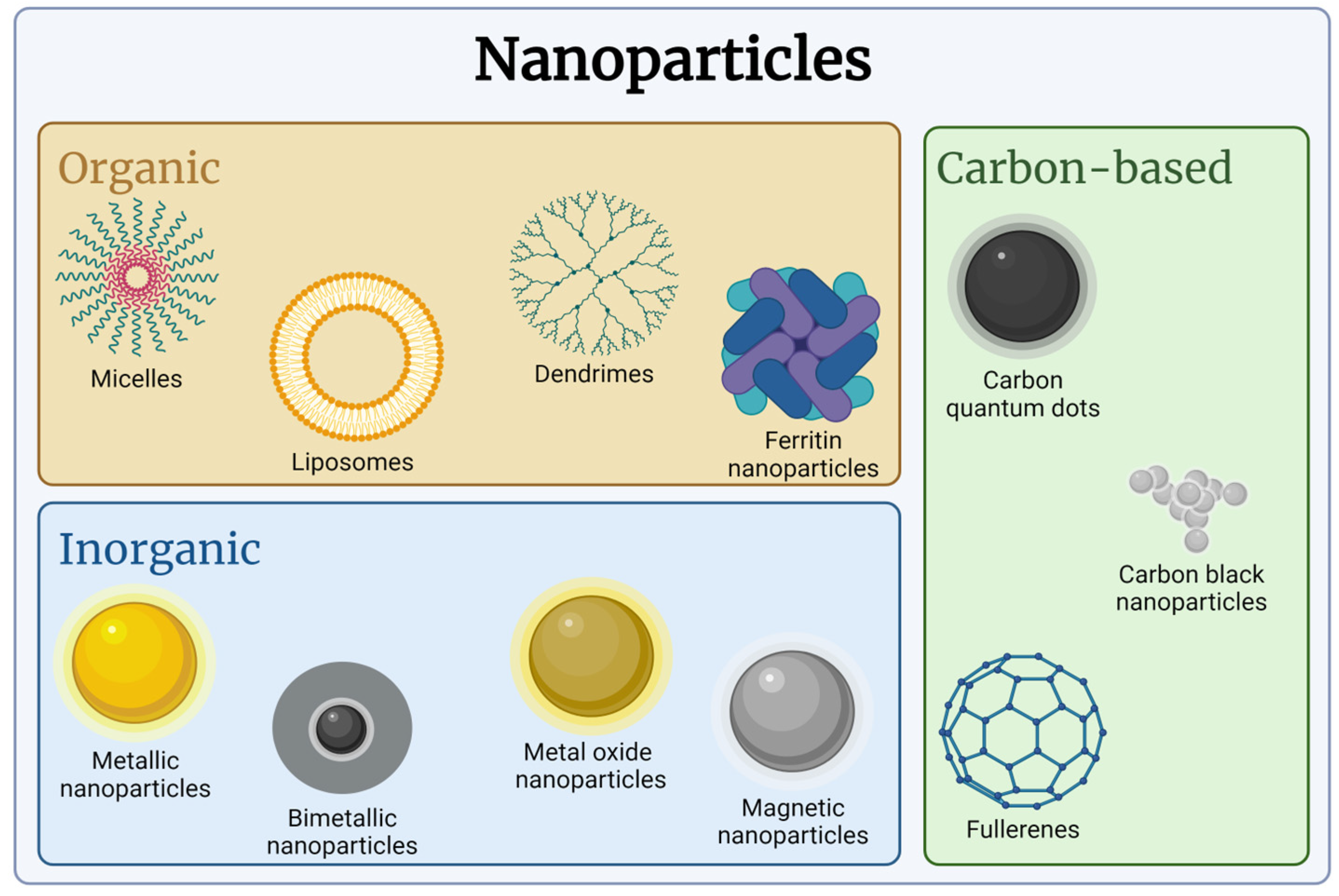
3.1. Nanomaterials and NPs
3.2. NPs in the Body Fluids and Coating Agents
3.3. EVs as a Natural and Pathological Delivery System
3.4. Nanomaterial and NPs Applications
4. NP Delivery of circRNA Targeting Agents

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Cancer | NPs | CircRNA | Molecular Mechanism | Antitumor Activity | Ref. |
|---|---|---|---|---|---|
| Breast | Gold NPs/siRNA or ASO | circDNMT1 | p53 transcription inhibition | Autophagy suppression | [44] |
| Melanoma | Gold NPs/circRNA | circFoxo3 | Interaction with MDM2/p53 | Apoptosis induction | [131] |
| Melanoma | Gold NPs/ASO | circCcnb1 | Interaction with Ccnb1/Cdk1 | Cell invasion suppression | [131] |
| Hepatocellular carcinoma | Superparamagnetic iron oxide NPs/siRNA | circ_0058051 | P38 downregulation | Migration/proliferation inhibition | [133] |
| Hepatocellular carcinoma | PAEs NPs/siRNA | circMDK | PI3K/AKT/mTOR pathway modulation | Proliferation/invasion/migration inhibition | [134] |
| Breast | Gold NPs/expression vector | circEHMT1 | MMP2 pathway modulation | Migration/invasion inhibition | [134] |
| Breast | Micropoly-Transfecter NPs/expression vector | circ-1073 | Cleaved Caspase-3/9 overexpression | Apoptosis induction | [135] |
| Renal cell carcinoma | CS-EGCG NPs/expression vector | circSPIRE1 | E-cadherin glycosylation | Proliferation/angiogenesis/metastasis suppression | [137] |
| Gastric | Organic cholesterol NPs/siRNA | circNRIP1 | miR-149-5p sponging | Chemotherapy sensitivity restoration | [138] |
| Breast | PLGA-NPs/siRNA | cSERPINE2 | MALT1-NF-κB-IL-6 axis | Proliferation/migration inhibition | [139] |
| Hepatocellular carcinoma | PLGA-NPs/siRNA | circROBO1 | miR-130a-5p/CCNT2 axis modulation | Proliferation/invasion inhibition | [140] |
| Lung adenocarcinoma | Polyethyleneimine-based NPs/circRNA | miR-21-5p ciRs | miR-21-5p decoy | Proliferation/invasion inhibition | [140] |
| Colorectal | Exosomes/siRNA | ciRS-122 | PKM2 downregulation | Chemotherapy sensitivity restoration | [142] |
5. Conclusions
Author Contributions
Funding
Conflicts of Interest
References
- Zeng, L.; Yuan, S.; Zhou, P.; Gong, J.; Kong, X.; Wu, M. Circular RNA Pvt1 oncogene (CircPVT1) promotes the progression of papillary thyroid carcinoma by activating the Wnt/beta-catenin signaling pathway and modulating the ratio of microRNA-195 (miR-195) to vascular endothelial growth factor A (VEGFA) expression. Bioengineered 2021, 12, 11795–11810. [Google Scholar] [CrossRef] [PubMed]
- Chaudhary, R.; Muys, B.R.; Grammatikakis, I.; De, S.; Abdelmohsen, K.; Li, X.L.; Zhu, Y.; Daulatabad, S.V.; Tsitsipatis, D.; Meltzer, P.S.; et al. A Circular RNA from the MDM2 Locus Controls Cell Cycle Progression by Suppressing p53 Levels. Mol. Cell. Biol. 2020, 40, 965–975. [Google Scholar] [CrossRef] [PubMed]
- Xue, C.; Li, G.; Lu, J.; Li, L. Crosstalk between circRNAs and the PI3K/AKT signaling pathway in cancer progression. Signal Transduct. Target. Ther. 2021, 6, 400. [Google Scholar] [CrossRef] [PubMed]
- Kong, Y.; Luo, Y.; Zheng, S.; Yang, J.; Zhang, D.; Zhao, Y.; Zheng, H.; An, M.; Lin, Y.; Ai, L.; et al. Mutant KRAS Mediates circARFGEF2 Biogenesis to Promote Lymphatic Metastasis of Pancreatic Ductal Adenocarcinoma. Cancer Res. 2023, 83, 3077–3094. [Google Scholar] [CrossRef]
- Zhou, C.; Li, W.; Liang, Z.; Wu, X.; Cheng, S.; Peng, J.; Zeng, K.; Li, W.; Lan, P.; Yang, X.; et al. Mutant KRAS-activated circATXN7 fosters tumor immunoescape by sensitizing tumor-specific T cells to activation-induced cell death. Nat. Commun. 2024, 15, 499. [Google Scholar] [CrossRef] [PubMed]
- Kristensen, L.S.; Jakobsen, T.; Hager, H.; Kjems, J. The emerging roles of circRNAs in cancer and oncology. Nat. Rev. Clin. Oncol. 2022, 19, 188–206. [Google Scholar] [CrossRef]
- Qiu, S.; Li, B.; Xia, Y.; Xuan, Z.; Li, Z.; Xie, L.; Gu, C.; Lv, J.; Lu, C.; Jiang, T.; et al. CircTHBS1 drives gastric cancer progression by increasing INHBA mRNA expression and stability in a ceRNA- and RBP-dependent manner. Cell. Death. Dis. 2022, 13, 266. [Google Scholar] [CrossRef] [PubMed]
- Chen, J.; Wu, Y.; Luo, X.; Jin, D.; Zhou, W.; Ju, Z.; Wang, D.; Meng, Q.; Wang, H.; Fu, X.; et al. Circular RNA circRHOBTB3 represses metastasis by regulating the HuR-mediated mRNA stability of PTBP1 in colorectal cancer. Theranostics 2021, 11, 7507–7526. [Google Scholar] [CrossRef]
- Wang, X.; Chen, M.; Fang, L. hsa_circ_0068631 promotes breast cancer progression through c-Myc by binding to EIF4A3. Mol. Ther. Nucleic Acids. 2021, 26, 122–134. [Google Scholar] [CrossRef]
- Kim, W.R.; Park, E.G.; Lee, D.H.; Lee, Y.J.; Bae, W.H.; Kim, H.S. The Tumorigenic Role of Circular RNA-MicroRNA Axis in Cancer. Int. J. Mol. Sci. 2023, 24, 3050. [Google Scholar] [CrossRef] [PubMed]
- Pisignano, G.; Michael, D.C.; Visal, T.H.; Pirlog, R.; Ladomery, M.; Calin, G.A. Going circular: History, present, and future of circRNAs in cancer. Oncogene 2023, 42, 2783–2800. [Google Scholar] [CrossRef]
- He, A.T.; Liu, J.; Li, F.; Yang, B.B. Targeting circular RNAs as a therapeutic approach: Current strategies and challenges. Signal Transduct. Target. Ther. 2021, 6, 185. [Google Scholar] [CrossRef]
- Karedath, T.; Ahmed, I.; Al Ameri, W.; Al-Dasim, F.M.; Andrews, S.S.; Samuel, S.; Al-Azwani, I.K.; Mohamoud, Y.A.; Rafii, A.; Malek, J.A. Silencing of ANKRD12 circRNA induces molecular and functional changes associated with invasive phenotypes. BMC Cancer 2019, 19, 565. [Google Scholar] [CrossRef] [PubMed]
- Hama Faraj, G.S.; Hussen, B.M.; Abdullah, S.R.; Fatih Rasul, M.; Hajiesmaeili, Y.; Baniahmad, A.; Taheri, M. Advanced approaches of the use of circRNAs as a replacement for cancer therapy. Non-Coding RNA Res. 2024, 9, 811–830. [Google Scholar] [CrossRef] [PubMed]
- Liang, D.; Tatomer, D.C.; Luo, Z.; Wu, H.; Yang, L.; Chen, L.L.; Cherry, S.; Wilusz, J.E. The Output of Protein-Coding Genes Shifts to Circular RNAs When the Pre-mRNA Processing Machinery Is Limiting. Mol. Cell 2017, 68, 940–954.e3. [Google Scholar] [CrossRef]
- Coscujuela Tarrero, L.; Ferrero, G.; Miano, V.; De Intinis, C.; Ricci, L.; Arigoni, M.; Riccardo, F.; Annaratone, L.; Castellano, I.; Calogero, R.A.; et al. Luminal breast cancer-specific circular RNAs uncovered by a novel tool for data analysis. Oncotarget 2018, 9, 14580–14596. [Google Scholar] [CrossRef] [PubMed]
- Ashwal-Fluss, R.; Meyer, M.; Pamudurti, N.R.; Ivanov, A.; Bartok, O.; Hanan, M.; Evantal, N.; Memczak, S.; Rajewsky, N.; Kadener, S. circRNA biogenesis competes with pre-mRNA splicing. Mol. Cell 2014, 56, 55–66. [Google Scholar] [CrossRef] [PubMed]
- Li, X.; Yang, L.; Chen, L.L. The Biogenesis, Functions, and Challenges of Circular RNAs. Mol. Cell 2018, 71, 428–442. [Google Scholar] [CrossRef]
- Zhang, Y.; Zhang, X.O.; Chen, T.; Xiang, J.F.; Yin, Q.F.; Xing, Y.H.; Zhu, S.; Yang, L.; Chen, L.L. Circular intronic long noncoding RNAs. Mol. Cell 2013, 51, 792–806. [Google Scholar] [CrossRef]
- Li, Z.; Huang, C.; Bao, C.; Chen, L.; Lin, M.; Wang, X.; Zhong, G.; Yu, B.; Hu, W.; Dai, L.; et al. Exon-intron circular RNAs regulate transcription in the nucleus. Nat. Struct. Mol. Biol. 2015, 22, 256–264. [Google Scholar] [CrossRef]
- Zhou, W.Y.; Cai, Z.R.; Liu, J.; Wang, D.S.; Ju, H.Q.; Xu, R.H. Circular RNA: Metabolism, functions and interactions with proteins. Mol. Cancer 2020, 19, 172. [Google Scholar] [CrossRef]
- Su, R.; Zhou, M.; Lin, J.; Shan, G.; Huang, C. A circular RNA-gawky-chromatin regulatory axis modulates stress-induced transcription. Nucleic Acids Res. 2024, 52, 3702–3721. [Google Scholar] [CrossRef] [PubMed]
- Zhao, Q.; Liu, J.; Deng, H.; Ma, R.; Liao, J.Y.; Liang, H.; Hu, J.; Li, J.; Guo, Z.; Cai, J.; et al. Targeting Mitochondria-Located circRNA SCAR Alleviates NASH via Reducing mROS Output. Cell 2020, 183, 76–93.e22. [Google Scholar] [CrossRef] [PubMed]
- Hon, K.W.; Ab-Mutalib, N.S.; Abdullah, N.M.A.; Jamal, R.; Abu, N. Extracellular Vesicle-derived circular RNAs confers chemoresistance in Colorectal cancer. Sci. Rep. 2019, 9, 16497. [Google Scholar] [CrossRef]
- Tao, W.; Luo, Z.H.; He, Y.D.; Wang, B.Y.; Xia, T.L.; Deng, W.M.; Zhang, L.X.; Tang, X.M.; Meng, Z.A.; Gao, X.; et al. Plasma extracellular vesicle circRNA signature and resistance to abiraterone in metastatic castration-resistant prostate cancer. Br. J. Cancer 2023, 128, 1320–1332. [Google Scholar] [CrossRef]
- Shi, X.; Wang, B.; Feng, X.; Xu, Y.; Lu, K.; Sun, M. circRNAs and Exosomes: A Mysterious Frontier for Human Cancer. Mol. Ther. Nucleic Acids 2020, 19, 384–392. [Google Scholar] [CrossRef]
- Chen, N.; Zhao, G.; Yan, X.; Lv, Z.; Yin, H.; Zhang, S.; Song, W.; Li, X.; Li, L.; Du, Z.; et al. A novel FLI1 exonic circular RNA promotes metastasis in breast cancer by coordinately regulating TET1 and DNMT1. Genome Biol. 2018, 19, 218. [Google Scholar] [CrossRef] [PubMed]
- Shao, T.; Pan, Y.H.; Xiong, X.D. Circular RNA: An important player with multiple facets to regulate its parental gene expression. Mol. Ther. Nucleic Acids 2021, 23, 369–376. [Google Scholar] [CrossRef] [PubMed]
- O’Brien, J.; Hayder, H.; Zayed, Y.; Peng, C. Overview of MicroRNA Biogenesis, Mechanisms of Actions, and Circulation. Front. Endocrinol. 2018, 9, 402. [Google Scholar] [CrossRef]
- Hentze, M.W.; Castello, A.; Schwarzl, T.; Preiss, T. A brave new world of RNA-binding proteins. Nat. Rev. Mol. Cell Biol. 2018, 19, 327–341. [Google Scholar] [CrossRef]
- Conn, S.J.; Pillman, K.A.; Toubia, J.; Conn, V.M.; Salmanidis, M.; Phillips, C.A.; Roslan, S.; Schreiber, A.W.; Gregory, P.A.; Goodall, G.J. The RNA binding protein quaking regulates formation of circRNAs. Cell 2015, 160, 1125–1134. [Google Scholar] [CrossRef] [PubMed]
- Du, W.W.; Yang, W.; Liu, E.; Yang, Z.; Dhaliwal, P.; Yang, B.B. Foxo3 circular RNA retards cell cycle progression via forming ternary complexes with p21 and CDK2. Nucleic Acids Res. 2016, 44, 2846–2858. [Google Scholar] [CrossRef] [PubMed]
- Holdt, L.M.; Stahringer, A.; Sass, K.; Pichler, G.; Kulak, N.A.; Wilfert, W.; Kohlmaier, A.; Herbst, A.; Northoff, B.H.; Nicolaou, A.; et al. Circular non-coding RNA ANRIL modulates ribosomal RNA maturation and atherosclerosis in humans. Nat. Commun. 2016, 7, 12429. [Google Scholar] [CrossRef] [PubMed]
- Stoll, L.; Rodriguez-Trejo, A.; Guay, C.; Brozzi, F.; Bayazit, M.B.; Gattesco, S.; Menoud, V.; Sobel, J.; Marques, A.C.; Veno, M.T.; et al. A circular RNA generated from an intron of the insulin gene controls insulin secretion. Nat. Commun. 2020, 11, 5611. [Google Scholar] [CrossRef]
- Fan, X.; Yang, Y.; Chen, C.; Wang, Z. Pervasive translation of circular RNAs driven by short IRES-like elements. Nat. Commun. 2022, 13, 3751. [Google Scholar] [CrossRef] [PubMed]
- Di Timoteo, G.; Dattilo, D.; Centron-Broco, A.; Colantoni, A.; Guarnacci, M.; Rossi, F.; Incarnato, D.; Oliviero, S.; Fatica, A.; Morlando, M.; et al. Modulation of circRNA Metabolism by m(6)A Modification. Cell Rep. 2020, 31, 107641. [Google Scholar] [CrossRef]
- Li, C.; Zhang, J.; Yang, X.; Hu, C.; Chu, T.; Zhong, R.; Shen, Y.; Hu, F.; Pan, F.; Xu, J.; et al. hsa_circ_0003222 accelerates stemness and progression of non-small cell lung cancer by sponging miR-527. Cell Death Dis. 2021, 12, 807. [Google Scholar] [CrossRef] [PubMed]
- Rengganaten, V.; Huang, C.J.; Wang, M.L.; Chien, Y.; Tsai, P.H.; Lan, Y.T.; Ong, H.T.; Chiou, S.H.; Choo, K.B. Circular RNA ZNF800 (hsa_circ_0082096) regulates cancer stem cell properties and tumor growth in colorectal cancer. BMC Cancer 2023, 23, 1088. [Google Scholar] [CrossRef] [PubMed]
- Zhang, Y.; Luo, J.; Yang, W.; Ye, W.C. CircRNAs in colorectal cancer: Potential biomarkers and therapeutic targets. Cell Death Dis. 2023, 14, 353. [Google Scholar] [CrossRef]
- Wang, X.; Zhang, J.; Luo, F.; Shen, Y. Application of Circular RNA Circ_0071662 in the Diagnosis and Prognosis of Hepatocellular Carcinoma and Its Response to Radiotherapy. Dig. Dis. 2023, 41, 431–438. [Google Scholar] [CrossRef]
- Begliarzade, S.; Sufianov, A.; Ilyasova, T.; Shumadalova, A.; Sufianov, R.; Beylerli, O.; Yan, Z. Circular RNA in cervical cancer: Fundamental mechanism and clinical potential. Noncoding RNA Res. 2024, 9, 116–124. [Google Scholar] [CrossRef] [PubMed]
- Shi, M.; Li, Z.Y.; Zhang, L.M.; Wu, X.Y.; Xiang, S.H.; Wang, Y.G.; Zhang, Y.Q. Hsa_circ_0007456 regulates the natural killer cell-mediated cytotoxicity toward hepatocellular carcinoma via the miR-6852-3p/ICAM-1 axis. Cell Death Dis. 2021, 12, 94. [Google Scholar] [CrossRef]
- Zhu, J.; Li, Q.; Wu, Z.; Xu, W.; Jiang, R. Circular RNA-mediated miRNA sponge & RNA binding protein in biological modulation of breast cancer. Non-Coding RNA Res. 2024, 9, 262–276. [Google Scholar]
- Du, W.W.; Yang, W.; Li, X.; Awan, F.M.; Yang, Z.; Fang, L.; Lyu, J.; Li, F.; Peng, C.; Krylov, S.N.; et al. A circular RNA circ-DNMT1 enhances breast cancer progression by activating autophagy. Oncogene 2018, 37, 5829–5842. [Google Scholar] [CrossRef] [PubMed]
- He, X.; Xu, T.; Hu, W.; Tan, Y.; Wang, D.; Wang, Y.; Zhao, C.; Yi, Y.; Xiong, M.; Lv, W.; et al. Circular RNAs: Their Role in the Pathogenesis and Orchestration of Breast Cancer. Front. Cell Dev. Biol. 2021, 9, 647736. [Google Scholar] [CrossRef]
- Liang, X.; Liu, X.; Song, Z.; Zhu, J.; Zhang, J. Hsa_circ_0097922 promotes tamoxifen resistance and cell malignant behaviour of breast cancer cells by regulating ACTN4 expression via miR-876-3p. Clin. Exp. Pharmacol. Physiol. 2022, 49, 1257–1269. [Google Scholar] [CrossRef]
- Wang, X.; Jian, W.; Luo, Q.; Fang, L. CircSEMA4B inhibits the progression of breast cancer by encoding a novel protein SEMA4B-211aa and regulating AKT phosphorylation. Cell Death Dis. 2022, 13, 794. [Google Scholar] [CrossRef]
- Hanniford, D.; Ulloa-Morales, A.; Karz, A.; Berzoti-Coelho, M.G.; Moubarak, R.S.; Sanchez-Sendra, B.; Kloetgen, A.; Davalos, V.; Imig, J.; Wu, P.; et al. Epigenetic Silencing of CDR1as Drives IGF2BP3-Mediated Melanoma Invasion and Metastasis. Cancer Cell 2020, 37, 55–70.e15. [Google Scholar] [CrossRef]
- Shang, Q.; Du, H.; Wu, X.; Guo, Q.; Zhang, F.; Gong, Z.; Jiao, T.; Guo, J.; Kong, Y. FMRP ligand circZNF609 destabilizes RAC1 mRNA to reduce metastasis in acral melanoma and cutaneous melanoma. J. Exp. Clin. Cancer Res. 2022, 41, 170. [Google Scholar] [CrossRef]
- Hao, T.; Yang, Y.; He, J.; Bai, J.; Zheng, Y.; Luo, Z. Knockdown of circular RNA hsa_circ_0062270 suppresses the progression of melanoma via downregulation of CDC45. Histol. Histopathol 2022, 37, 373–383. [Google Scholar]
- Li, J.; Ma, M.; Yang, X.; Zhang, M.; Luo, J.; Zhou, H.; Huang, N.; Xiao, F.; Lai, B.; Lv, W.; et al. Circular HER2 RNA positive triple negative breast cancer is sensitive to Pertuzumab. Mol. Cancer 2020, 19, 142. [Google Scholar] [CrossRef] [PubMed]
- Zhang, L.; Gao, H.; Li, X.; Yu, F.; Li, P. The important regulatory roles of circRNA-encoded proteins or peptides in cancer pathogenesis (Review). Int. J. Oncol. 2024, 64, 19. [Google Scholar] [CrossRef]
- Mecozzi, N.; Nenci, A.; Vera, O.; Bok, I.; Falzone, A.; DeNicola, G.M.; Karreth, F.A. Genetic tools for the stable overexpression of circular RNAs. RNA Biol. 2022, 19, 353–363. [Google Scholar] [CrossRef]
- Salvioni, L.; Rizzuto, M.A.; Bertolini, J.A.; Pandolfi, L.; Colombo, M.; Prosperi, D. Thirty Years of Cancer Nanomedicine: Success, Frustration, and Hope. Cancers 2019, 11, 1855. [Google Scholar] [CrossRef]
- Soares, S.; Sousa, J.; Pais, A.; Vitorino, C. Nanomedicine: Principles, Properties, and Regulatory Issues. Front. Chem. 2018, 6, 360. [Google Scholar] [CrossRef]
- Sultana, A.; Zare, M.; Thomas, V.; Kumar, T.S.S.; Ramakrishna, S. Nano-based drug delivery systems: Conventional drug delivery routes, recent developments and future prospects. Med. Drug Discov. 2022, 15, 100134. [Google Scholar] [CrossRef]
- Jain, A.; Bhattacharya, S. Recent advances in nanomedicine preparative methods and their therapeutic potential for colorectal cancer: A critical review. Front. Oncol. 2023, 13, 1211603. [Google Scholar] [CrossRef]
- Giri, P.M.; Banerjee, A.; Layek, B. A Recent Review on Cancer Nanomedicine. Cancers 2023, 15, 2256. [Google Scholar] [CrossRef]
- Fathi-Karkan, S.; Arshad, R.; Rahdar, A.; Ramezani, A.; Behzadmehr, R.; Ghotekar, S.; Pandey, S. Recent advancements in the targeted delivery of etoposide nanomedicine for cancer therapy: A comprehensive review. Eur. J. Med. Chem. 2023, 259, 115676. [Google Scholar] [CrossRef]
- Bosetti, R.; Jones, S.L. Cost-effectiveness of nanomedicine: Estimating the real size of nano-costs. Nanomedicine 2019, 14, 1367–1370. [Google Scholar] [CrossRef] [PubMed]
- Afzal, O.; Altamimi, A.S.A.; Nadeem, M.S.; Alzarea, S.I.; Almalki, W.H.; Tariq, A.; Mubeen, B.; Murtaza, B.N.; Iftikhar, S.; Riaz, N.; et al. Nanoparticles in Drug Delivery: From History to Therapeutic Applications. Nanomaterials 2022, 12, 4494. [Google Scholar] [CrossRef] [PubMed]
- Yao, Y.; Zhou, Y.; Liu, L.; Xu, Y.; Chen, Q.; Wang, Y.; Wu, S.; Deng, Y.; Zhang, J.; Shao, A. Nanoparticle-Based Drug Delivery in Cancer Therapy and Its Role in Overcoming Drug Resistance. Front. Mol. Biosci. 2020, 7, 193. [Google Scholar] [CrossRef] [PubMed]
- Joudeh, N.; Linke, D. Nanoparticle classification, physicochemical properties, characterization, and applications: A comprehensive review for biologists. J. Nanobiotechnol. 2022, 20, 262. [Google Scholar] [CrossRef] [PubMed]
- Bose, A.; Roy Burman, D.; Sikdar, B.; Patra, P. Nanomicelles: Types, properties and applications in drug delivery. IET Nanobiotechnol. 2021, 15, 19–27. [Google Scholar] [CrossRef] [PubMed]
- Fang, X.; Gao, K.; Huang, J.; Liu, K.; Chen, L.; Piao, Y.; Liu, X.; Tang, J.; Shen, Y.; Zhou, Z. Molecular level precision and high molecular weight peptide dendrimers for drug-specific delivery. J. Mater. Chem. B 2021, 9, 8594–8603. [Google Scholar] [CrossRef] [PubMed]
- Roy, M.; Roy, A.; Rustagi, S.; Pandey, N. An Overview of Nanomaterial Applications in Pharmacology. Biomed. Res. Int. 2023, 2023, 4838043. [Google Scholar] [CrossRef]
- Pasarin, D.; Ghizdareanu, A.I.; Enascuta, C.E.; Matei, C.B.; Bilbie, C.; Paraschiv-Palada, L.; Veres, P.A. Coating Materials to Increase the Stability of Liposomes. Polymers 2023, 15, 782. [Google Scholar] [CrossRef]
- Lim, S.A.; Cox, A.; Tung, M.; Chung, E.J. Clinical progress of nanomedicine-based RNA therapies. Bioact. Mater. 2022, 12, 203–213. [Google Scholar] [CrossRef] [PubMed]
- Mitchell, M.J.; Billingsley, M.M.; Haley, R.M.; Wechsler, M.E.; Peppas, N.A.; Langer, R. Engineering precision nanoparticles for drug delivery. Nat. Rev. Drug. Discov. 2021, 20, 101–124. [Google Scholar] [CrossRef] [PubMed]
- Thapa, R.K.; Kim, J.O. Nanomedicine-based commercial formulations: Current developments and future prospects. J. Pharm. Investig. 2023, 53, 19–33. [Google Scholar] [CrossRef]
- Hoshyar, N.; Gray, S.; Han, H.; Bao, G. The effect of nanoparticle size on in vivo pharmacokinetics and cellular interaction. Nanomedicine 2016, 11, 673–692. [Google Scholar] [CrossRef] [PubMed]
- Fang, R.H.; Gao, W.; Zhang, L. Targeting drugs to tumours using cell membrane-coated nanoparticles. Nat. Rev. Clin. Oncol. 2023, 20, 33–48. [Google Scholar] [CrossRef]
- Carofiglio, M.; Laurenti, M.; Vighetto, V.; Racca, L.; Barui, S.; Garino, N.; Gerbaldo, R.; Laviano, F.; Cauda, V. Iron-Doped ZnO Nanoparticles as Multifunctional Nanoplatforms for Theranostics. Nanomaterials 2021, 11, 2628. [Google Scholar] [CrossRef] [PubMed]
- Dumontel, B.; Canta, M.; Engelke, H.; Chiodoni, A.; Racca, L.; Ancona, A.; Limongi, T.; Canavese, G.; Cauda, V. Enhanced biostability and cellular uptake of zinc oxide nanocrystals shielded with a phospholipid bilayer. J. Mater. Chem. B 2017, 5, 8799–8813. [Google Scholar] [CrossRef] [PubMed]
- Amina, S.J.; Guo, B. A Review on the Synthesis and Functionalization of Gold Nanoparticles as a Drug Delivery Vehicle. Int. J. Nanomed. 2020, 15, 9823–9857. [Google Scholar] [CrossRef] [PubMed]
- Boughbina-Portoles, A.; Sanjuan-Navarro, L.; Hakobyan, L.; Gomez-Ferrer, M.; Moliner-Martinez, Y.; Sepulveda, P.; Campins-Falco, P. Reliable assessment of carbon black nanomaterial of a variety of cell culture media for in vitro toxicity assays by asymmetrical flow field-flow fractionation. Anal. Bioanal. Chem. 2023, 415, 2121–2132. [Google Scholar] [CrossRef] [PubMed]
- Hashikawa, Y.; Sadai, S.; Ikemoto, Y.; Murata, Y. Water Adsorption on pi-Surfaces of Open-Fullerenes. J. Phys. Chem. A 2024, 128, 2782–2788. [Google Scholar] [CrossRef] [PubMed]
- Ajdary, M.; Moosavi, M.A.; Rahmati, M.; Falahati, M.; Mahboubi, M.; Mandegary, A.; Jangjoo, S.; Mohammadinejad, R.; Varma, R.S. Health Concerns of Various Nanoparticles: A Review of Their in Vitro and in Vivo Toxicity. Nanomaterials 2018, 8, 634. [Google Scholar] [CrossRef]
- Guerrini, L.; Alvarez-Puebla, R.A.; Pazos-Perez, N. Surface Modifications of Nanoparticles for Stability in Biological Fluids. Materials 2018, 11, 1154. [Google Scholar] [CrossRef]
- Soenen, S.J.; Parak, W.J.; Rejman, J.; Manshian, B. (Intra)cellular stability of inorganic nanoparticles: Effects on cytotoxicity, particle functionality, and biomedical applications. Chem. Rev. 2015, 115, 2109–2135. [Google Scholar] [CrossRef]
- Simak, J.; De Paoli, S. The effects of nanomaterials on blood coagulation in hemostasis and thrombosis. Wiley Interdiscip. Rev. Nanomed. Nanobiotechnol. 2017, 9, 1148. [Google Scholar] [CrossRef] [PubMed]
- Dumontel, B.; Jimenez-Jimenez, C.; Vallet-Regi, M.; Manzano, M. Bioinspired extracellular vesicle-coated silica nanoparticles as selective delivery systems. Mater. Today Bio. 2023, 23, 100850. [Google Scholar] [CrossRef] [PubMed]
- Juan, A.; Cimas, F.J.; Bravo, I.; Pandiella, A.; Ocana, A.; Alonso-Moreno, C. Antibody Conjugation of Nanoparticles as Therapeutics for Breast Cancer Treatment. Int. J. Mol. Sci. 2020, 21, 6018. [Google Scholar] [CrossRef]
- Miao, L.; Kang, Y.; Zhang, X.F. Nanotechnology for the theranostic opportunity of breast cancer lung metastasis: Recent advancements and future challenges. Front. Bioeng. Biotechnol. 2024, 12, 1410017. [Google Scholar] [CrossRef]
- Shao, M.; Lopes, D.; Lopes, J.; Yousefiasl, S.; Macário-Soares, A.; Peixoto, D.; Ferreira-Faria, I.; Veiga, F.; Conde, J.; Huang, Y.; et al. Exosome membrane-coated nanosystems: Exploring biomedical applications in cancer diagnosis and therapy. Matter 2023, 6, 761–799. [Google Scholar] [CrossRef]
- Wang, Z.; Mo, H.; He, Z.; Chen, A.; Cheng, P. Extracellular vesicles as an emerging drug delivery system for cancer treatment: Current strategies and recent advances. Biomed. Pharmacother. 2022, 153, 113480. [Google Scholar] [CrossRef]
- Mejia-Mendez, J.L.; Vazquez-Duhalt, R.; Hernandez, L.R.; Sanchez-Arreola, E.; Bach, H. Virus-like Particles: Fundamentals and Biomedical Applications. Int. J. Mol. Sci. 2022, 23, 8579. [Google Scholar] [CrossRef] [PubMed]
- Costa-Silva, B.; Aiello, N.M.; Ocean, A.J.; Singh, S.; Zhang, H.; Thakur, B.K.; Becker, A.; Hoshino, A.; Mark, M.T.; Molina, H.; et al. Pancreatic cancer exosomes initiate pre-metastatic niche formation in the liver. Nat. Cell Biol. 2015, 17, 816–826. [Google Scholar] [CrossRef]
- Si, G.; Chen, X.; Li, Y.; Yuan, X. Exosomes promote pre-metastatic niche formation in colorectal cancer. Heliyon 2024, 10, 27572. [Google Scholar] [CrossRef]
- Bhatia, R.; Chang, J.; Munoz, J.L.; Walker, N.D. Forging New Therapeutic Targets: Efforts of Tumor Derived Exosomes to Prepare the Pre-Metastatic Niche for Cancer Cell Dissemination and Dormancy. Biomedicines 2023, 11, 1614. [Google Scholar] [CrossRef]
- Agnoletto, C.; Pignochino, Y.; Caruso, C.; Garofalo, C. Exosome-Based Liquid Biopsy Approaches in Bone and Soft Tissue Sarcomas: Review of the Literature, Prospectives, and Hopes for Clinical Application. Int. J. Mol. Sci. 2023, 24, 5159. [Google Scholar] [CrossRef] [PubMed]
- Yu, D.; Li, Y.; Wang, M.; Gu, J.; Xu, W.; Cai, H.; Fang, X.; Zhang, X. Exosomes as a new frontier of cancer liquid biopsy. Mol. Cancer 2022, 21, 56. [Google Scholar] [CrossRef]
- Romero, M.R.; Veglia, A.V.; Amé, M.V.; Bracamonte, A.G. Multimodal Spectroscopy Assays for Advanced Nano-Optics Approaches by Tuning Nano-Tool Surface Chemistry and Metal-Enhanced Fluorescence. Crystals 2024, 14, 338. [Google Scholar] [CrossRef]
- Vargas-Nadal, G.; Kober, M.; Nsamela, A.; Terenziani, F.; Sissa, C.; Pescina, S.; Sonvico, F.; Gazzali, A.M.; Wahab, H.A.; Grisanti, L.; et al. Fluorescent Multifunctional Organic Nanoparticles for Drug Delivery and Bioimaging: A Tutorial Review. Pharmaceutics 2022, 14, 2498. [Google Scholar] [CrossRef]
- Zhao, Z.; Yan, R.; Yi, X.; Li, J.; Rao, J.; Guo, Z.; Yang, Y.; Li, W.; Li, Y.Q.; Chen, C. Bacteria-Activated Theranostic Nanoprobes against Methicillin-Resistant Staphylococcus aureus Infection. ACS Nano 2017, 11, 4428–4438. [Google Scholar] [CrossRef] [PubMed]
- Thwala, L.N.; Ndlovu, S.C.; Mpofu, K.T.; Lugongolo, M.Y.; Mthunzi-Kufa, P. Nanotechnology-Based Diagnostics for Diseases Prevalent in Developing Countries: Current Advances in Point-of-Care Tests. Nanomaterials 2023, 13, 1247. [Google Scholar] [CrossRef] [PubMed]
- Chow, J.C.L. 9-Magnetic nanoparticles as contrast agents in magnetic resonance imaging and radiosensitizers in radiotherapy. In Fundamentals and Industrial Applications of Magnetic Nanoparticles; Hussain, C.M., Patankar, K.K., Eds.; Woodhead Publishing: Cambridge, UK, 2022; pp. 291–316. [Google Scholar]
- Hosseini, S.M.; Mohammadnejad, J.; Salamat, S.; Beiram Zadeh, Z.; Tanhaei, M.; Ramakrishna, S. Theranostic polymeric nanoparticles as a new approach in cancer therapy and diagnosis: A review. Mater. Today Chem. 2023, 29, 101400. [Google Scholar] [CrossRef]
- Tripathi, P.; Shukla, P.; Bieberich, E. Theranostic Applications of Nanomaterials in Alzheimer’s Disease: A Multifunctional Approach. Curr. Pharm. Des. 2022, 28, 116–132. [Google Scholar] [CrossRef]
- Jiang, L.; Ding, L.; Liu, G. Nanoparticle formulations for therapeutic delivery, pathogen imaging and theranostic applications in bacterial infections. Theranostics 2023, 13, 1545–1570. [Google Scholar] [CrossRef]
- Itani, R.; Tobaiqy, M.; Al Faraj, A. Optimizing use of theranostic nanoparticles as a life-saving strategy for treating COVID-19 patients. Theranostics 2020, 10, 5932–5942. [Google Scholar] [CrossRef]
- Racca, L.; Canta, M.; Dumontel, B.; Ancona, A.; Limongi, T.; Garino, N.; Laurenti, M.; Canavese, G.; Cauda, V. 12—Zinc Oxide Nanostructures in Biomedicine. In Smart Nanoparticles for Biomedicine; Ciofani, G., Ed.; Elsevier: Amsterdam, The Netherlands, 2018; pp. 171–187. [Google Scholar]
- Leon-Buitimea, A.; Garza-Cervantes, J.A.; Gallegos-Alvarado, D.Y.; Osorio-Concepcion, M.; Morones-Ramirez, J.R. Nanomaterial-Based Antifungal Therapies to Combat Fungal Diseases Aspergillosis, Coccidioidomycosis, Mucormycosis, and Candidiasis. Pathogens 2021, 10, 1303. [Google Scholar] [CrossRef] [PubMed]
- Ratan, Z.A.; Mashrur, F.R.; Chhoan, A.P.; Shahriar, S.M.; Haidere, M.F.; Runa, N.J.; Kim, S.; Kweon, D.-H.; Hosseinzadeh, H.; Cho, J.Y. Silver Nanoparticles as Potential Antiviral Agents. Pharmaceutics 2021, 13, 2034. [Google Scholar] [CrossRef] [PubMed]
- Wolfgruber, S.; Rieger, J.; Cardozo, O.; Punz, B.; Himly, M.; Stingl, A.; Farias, P.M.A.; Abuja, P.M.; Zatloukal, K. Antiviral Activity of Zinc Oxide Nanoparticles against SARS-CoV-2. Int. J. Mol. Sci. 2023, 24, 8425. [Google Scholar] [CrossRef]
- Vighetto, V.; Racca, L.; Canta, M.; Matos, J.C.; Dumontel, B.; Goncalves, M.C.; Cauda, V. Smart Shockwave Responsive Titania-Based Nanoparticles for Cancer Treatment. Pharmaceutics 2021, 13, 1423. [Google Scholar] [CrossRef] [PubMed]
- Carofiglio, M.; Conte, M.; Racca, L.; Cauda, V. Synergistic Phenomena between Iron-Doped ZnO Nanoparticles and Shock Waves Exploited against Pancreatic Cancer Cells. ACS Appl. Nano Mater. 2022, 5, 17212–17225. [Google Scholar] [CrossRef] [PubMed]
- Racca, L.; Limongi, T.; Vighetto, V.; Dumontel, B.; Ancona, A.; Canta, M.; Canavese, G.; Garino, N.; Cauda, V. Zinc Oxide Nanocrystals and High-Energy Shock Waves: A New Synergy for the Treatment of Cancer Cells. Front. Bioeng. Biotechnol. 2020, 8, 577. [Google Scholar] [CrossRef] [PubMed]
- Haque, S.T.; Banaszak Holl, M.M.; Chowdhury, E.H. Strategies to assemble therapeutic and imaging molecules into inorganic nanocarriers. Front. Mater. Sci. 2022, 16, 220604. [Google Scholar] [CrossRef]
- Ma, H.; Xing, F.; Zhou, Y.; Yu, P.; Luo, R.; Xu, J.; Xiang, Z.; Rommens, P.M.; Duan, X.; Ritz, U. Design and fabrication of intracellular therapeutic cargo delivery systems based on nanomaterials: Current status and future perspectives. J. Mater. Chem. B 2023, 11, 7873–7912. [Google Scholar] [CrossRef]
- Yang, C.; Merlin, D. Nanoparticle-Mediated Drug Delivery Systems For The Treatment Of IBD: Current Perspectives. Int. J. Nanomed. 2019, 14, 8875–8889. [Google Scholar] [CrossRef]
- Yeh, Y.C.; Huang, T.H.; Yang, S.C.; Chen, C.C.; Fang, J.Y. Nano-Based Drug Delivery or Targeting to Eradicate Bacteria for Infection Mitigation: A Review of Recent Advances. Front. Chem. 2020, 8, 286. [Google Scholar] [CrossRef]
- Deng, Y.; Zhang, X.; Shen, H.; He, Q.; Wu, Z.; Liao, W.; Yuan, M. Application of the Nano-Drug Delivery System in Treatment of Cardiovascular Diseases. Front. Bioeng. Biotechnol. 2019, 7, 489. [Google Scholar] [CrossRef]
- Sell, M.; Lopes, A.R.; Escudeiro, M.; Esteves, B.; Monteiro, A.R.; Trindade, T.; Cruz-Lopes, L. Application of Nanoparticles in Cancer Treatment: A Concise Review. Nanomaterials 2023, 13, 2887. [Google Scholar] [CrossRef]
- Narum, S.M.; Le, T.; Le, D.P.; Lee, J.C.; Donahue, N.D.; Yang, W.; Wilhelm, S. Chapter 4—Passive targeting in nanomedicine: Fundamental concepts, body interactions, and clinical potential. In Nanoparticles for Biomedical Applications; Chung, E.J., Leon, L., Rinaldi, C., Eds.; Elsevier: Amsterdam, The Netherlands, 2020; pp. 37–53. [Google Scholar]
- Amer Ridha, A.; Kashanian, S.; Rafipour, R.; Hemati Azandaryani, A.; Zhaleh, H.; Mahdavian, E. A promising dual-drug targeted delivery system in cancer therapy: Nanocomplexes of folate-apoferritin-conjugated cationic solid lipid nanoparticles. Pharm. Dev. Technol. 2021, 26, 673–681. [Google Scholar] [CrossRef]
- Yamamoto, S.; Iwamaru, Y.; Shimizu, Y.; Ueda, Y.; Sato, M.; Yamaguchi, K.; Nakanishi, J. Epidermal growth factor-nanoparticle conjugates change the activity from anti-apoptotic to pro-apoptotic at membrane rafts. Acta Biomater. 2019, 88, 383–391. [Google Scholar] [CrossRef] [PubMed]
- Li, X.; Peng, X.; Zoulikha, M.; Boafo, G.F.; Magar, K.T.; Ju, Y.; He, W. Multifunctional nanoparticle-mediated combining therapy for human diseases. Signal Transduct. Target. Ther. 2024, 9, 1. [Google Scholar] [CrossRef]
- Debele, T.A.; Yeh, C.F.; Su, W.P. Cancer Immunotherapy and Application of Nanoparticles in Cancers Immunotherapy as the Delivery of Immunotherapeutic Agents and as the Immunomodulators. Cancers 2020, 12, 3773. [Google Scholar] [CrossRef]
- Mendes, B.B.; Conniot, J.; Avital, A.; Yao, D.; Jiang, X.; Zhou, X.; Sharf-Pauker, N.; Xiao, Y.; Adir, O.; Liang, H.; et al. Nanodelivery of nucleic acids. Nat. Rev. Methods Prim. 2022, 2, 24. [Google Scholar] [CrossRef] [PubMed]
- Piperno, A.; Sciortino, M.T.; Giusto, E.; Montesi, M.; Panseri, S.; Scala, A. Recent Advances and Challenges in Gene Delivery Mediated by Polyester-Based Nanoparticles. Int. J. Nanomed. 2021, 16, 5981–6002. [Google Scholar] [CrossRef] [PubMed]
- Durymanov, M.; Reineke, J. Non-viral Delivery of Nucleic Acids: Insight into Mechanisms of Overcoming Intracellular Barriers. Front. Pharmacol 2018, 9, 971. [Google Scholar] [CrossRef]
- Li, L.; Li, X.; Wu, Y.; Song, L.; Yang, X.; He, T.; Wang, N.; Yang, S.; Zeng, Y.; Wu, Q.; et al. Multifunctional Nucleus-targeting Nanoparticles with Ultra-high Gene Transfection Efficiency for In Vivo Gene Therapy. Theranostics 2017, 7, 1633–1649. [Google Scholar] [CrossRef]
- Yao, J.; Fan, Y.; Li, Y.; Huang, L. Strategies on the nuclear-targeted delivery of genes. J. Drug. Target. 2013, 21, 926–939. [Google Scholar] [CrossRef] [PubMed]
- Huo, S.; Jin, S.; Ma, X.; Xue, X.; Yang, K.; Kumar, A.; Wang, P.C.; Zhang, J.; Hu, Z.; Liang, X.J. Ultrasmall gold nanoparticles as carriers for nucleus-based gene therapy due to size-dependent nuclear entry. ACS Nano 2014, 8, 5852–5862. [Google Scholar] [CrossRef] [PubMed]
- Zare, M.; Pemmada, R.; Madhavan, M.; Shailaja, A.; Ramakrishna, S.; Kandiyil, S.P.; Donahue, J.M.; Thomas, V. Encapsulation of miRNA and siRNA into Nanomaterials for Cancer Therapeutics. Pharmaceutics 2022, 14, 1620. [Google Scholar] [CrossRef]
- Choi, K.Y.; Silvestre, O.F.; Huang, X.; Hida, N.; Liu, G.; Ho, D.N.; Lee, S.; Lee, S.W.; Hong, J.I.; Chen, X. A nanoparticle formula for delivering siRNA or miRNAs to tumor cells in cell culture and in vivo. Nat. Protoc. 2014, 9, 1900–1915. [Google Scholar] [CrossRef]
- Bravo-Vazquez, L.A.; Mendez-Garcia, A.; Rodriguez, A.L.; Sahare, P.; Pathak, S.; Banerjee, A.; Duttaroy, A.K.; Paul, S. Applications of nanotechnologies for miRNA-based cancer therapeutics: Current advances and future perspectives. Front. Bioeng. Biotechnol. 2023, 11, 1208547. [Google Scholar] [CrossRef] [PubMed]
- Chaudhari, R.; Nasra, S.; Meghani, N.; Kumar, A. MiR-206 conjugated gold nanoparticle based targeted therapy in breast cancer cells. Sci. Rep. 2022, 12, 4713. [Google Scholar] [CrossRef]
- Ghafouri-Fard, S.; Shoorei, H.; Noferesti, L.; Hussen, B.M.; Moghadam, M.H.B.; Taheri, M.; Rashnoo, F. Nanoparticle-mediated delivery of microRNAs-based therapies for treatment of disorders. Pathol. Res. Pract. 2023, 248, 154667. [Google Scholar] [CrossRef]
- Du, W.W.; Fang, L.; Yang, W.; Wu, N.; Awan, F.M.; Yang, Z.; Yang, B.B. Induction of tumor apoptosis through a circular RNA enhancing Foxo3 activity. Cell Death Differ. 2017, 24, 357–370. [Google Scholar] [CrossRef]
- Fang, L.; Du, W.W.; Awan, F.M.; Dong, J.; Yang, B.B. The circular RNA circ-Ccnb1 dissociates Ccnb1/Cdk1 complex suppressing cell invasion and tumorigenesis. Cancer Lett. 2019, 459, 216–226. [Google Scholar] [CrossRef]
- You, S.; Luo, Z.; Cheng, N.; Wu, M.; Lai, Y.; Wang, F.; Zheng, X.; Wang, Y.; Liu, X.; Liu, J.; et al. Magnetically responsive nanoplatform targeting circRNA circ_0058051 inhibits hepatocellular carcinoma progression. Drug Deliv. Transl. Res. 2023, 13, 782–794. [Google Scholar] [CrossRef]
- Du, A.; Li, S.; Zhou, Y.; Disoma, C.; Liao, Y.; Zhang, Y.; Chen, Z.; Yang, Q.; Liu, P.; Liu, S.; et al. M6A-mediated upregulation of circMDK promotes tumorigenesis and acts as a nanotherapeutic target in hepatocellular carcinoma. Mol. Cancer 2022, 21, 109. [Google Scholar] [CrossRef]
- Lu, M.; Wu, Y.; Zeng, B.; Sun, J.; Li, Y.; Luo, J.; Wang, L.; Yi, Z.; Li, H.; Ren, G. CircEHMT1 inhibits metastatic potential of breast cancer cells by modulating miR-1233-3p/KLF4/MMP2 axis. Biochem. Biophys. Res. Commun. 2020, 526, 306–313. [Google Scholar] [CrossRef]
- Yi, Z.; Li, Y.; Wu, Y.; Zeng, B.; Li, H.; Ren, G.; Wang, X. Circular RNA 0001073 Attenuates Malignant Biological Behaviours in Breast Cancer Cell and Is Delivered by Nanoparticles to Inhibit Mice Tumour Growth. OncoTargets Ther. 2020, 13, 6157–6169. [Google Scholar] [CrossRef]
- Shu, G.; Lu, X.; Pan, Y.; Cen, J.; Huang, K.; Zhou, M.; Lu, J.; Dong, J.; Han, H.; Chen, W.; et al. Exosomal circSPIRE1 mediates glycosylation of E-cadherin to suppress metastasis of renal cell carcinoma. Oncogene 2023, 42, 1802–1820. [Google Scholar] [CrossRef]
- Zhang, X.; Wang, S.; Wang, H.; Cao, J.; Huang, X.; Chen, Z.; Xu, P.; Sun, G.; Xu, J.; Lv, J.; et al. Circular RNA circNRIP1 acts as a microRNA-149-5p sponge to promote gastric cancer progression via the AKT1/mTOR pathway. Mol. Cancer 2019, 18, 20. [Google Scholar] [CrossRef]
- Zhou, B.; Mo, Z.; Lai, G.; Chen, X.; Li, R.; Wu, R.; Zhu, J.; Zheng, F. Targeting tumor exosomal circular RNA cSERPINE2 suppresses breast cancer progression by modulating MALT1-NF-κB-IL-6 axis of tumor-associated macrophages. J. Exp. Clin. Cancer Res. 2023, 42, 48. [Google Scholar] [CrossRef]
- Meng, H.; Li, R.; Xie, Y.; Mo, Z.; Zhai, H.; Zhang, G.; Liang, G.; Shi, X.; Zhou, B. Nanoparticles Mediated circROBO1 Silencing to Inhibit Hepatocellular Carcinoma Progression by Modulating miR-130a-5p/CCNT2 Axis. Int. J. Nanomed. 2023, 18, 1677–1693. [Google Scholar] [CrossRef] [PubMed]
- Müller, S.; Wedler, A.; Breuer, J.; Glaß, M.; Bley, N.; Lederer, M.; Haase, J.; Misiak, C.; Fuchs, T.; Ottmann, A.; et al. Synthetic circular miR-21 RNA decoys enhance tumor suppressor expression and impair tumor growth in mice. NAR Cancer 2020, 2, zcaa014. [Google Scholar] [CrossRef] [PubMed]
- Wang, X.; Zhang, H.; Yang, H.; Bai, M.; Ning, T.; Deng, T.; Liu, R.; Fan, Q.; Zhu, K.; Li, J.; et al. Exosome-delivered circRNA promotes glycolysis to induce chemoresistance through the miR-122-PKM2 axis in colorectal cancer. Mol. Oncol. 2020, 14, 539–555. [Google Scholar] [CrossRef] [PubMed]
- Yang, Z.G.; Awan, F.M.; Du, W.W.; Zeng, Y.; Lyu, J.; Wu, D.; Gupta, S.; Yang, W.; Yang, B.B. The Circular RNA Interacts with STAT3, Increasing Its Nuclear Translocation and Wound Repair by Modulating Dnmt3a and miR-17 Function. Mol. Ther. 2017, 25, 2062–2074. [Google Scholar] [CrossRef] [PubMed]
- Liu, Y.; Zhang, T.; Zou, X.; Yuan, Z.; Li, Y.; Zang, J.; He, N.; He, L.; Xu, A.; Lu, D. Penumbra-targeted CircOGDH siRNA-loaded nanoparticles alleviate neuronal apoptosis in focal brain ischaemia. Stroke Vasc. Neurol. 2024, 9, 134–144. [Google Scholar] [CrossRef] [PubMed]

Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Racca, L.; Liuzzi, E.; Comparato, S.; Giordano, G.; Pignochino, Y. Nanoparticles-Delivered Circular RNA Strategy as a Novel Antitumor Approach. Int. J. Mol. Sci. 2024, 25, 8934. https://doi.org/10.3390/ijms25168934
Racca L, Liuzzi E, Comparato S, Giordano G, Pignochino Y. Nanoparticles-Delivered Circular RNA Strategy as a Novel Antitumor Approach. International Journal of Molecular Sciences. 2024; 25(16):8934. https://doi.org/10.3390/ijms25168934
Chicago/Turabian StyleRacca, Luisa, Elisabetta Liuzzi, Simona Comparato, Giorgia Giordano, and Ymera Pignochino. 2024. "Nanoparticles-Delivered Circular RNA Strategy as a Novel Antitumor Approach" International Journal of Molecular Sciences 25, no. 16: 8934. https://doi.org/10.3390/ijms25168934