
NK Cell Degranulation Triggered by Rituximab Identifies Potential Markers of Subpopulations with Enhanced Cytotoxicity toward Malignant B Cells



Abstract
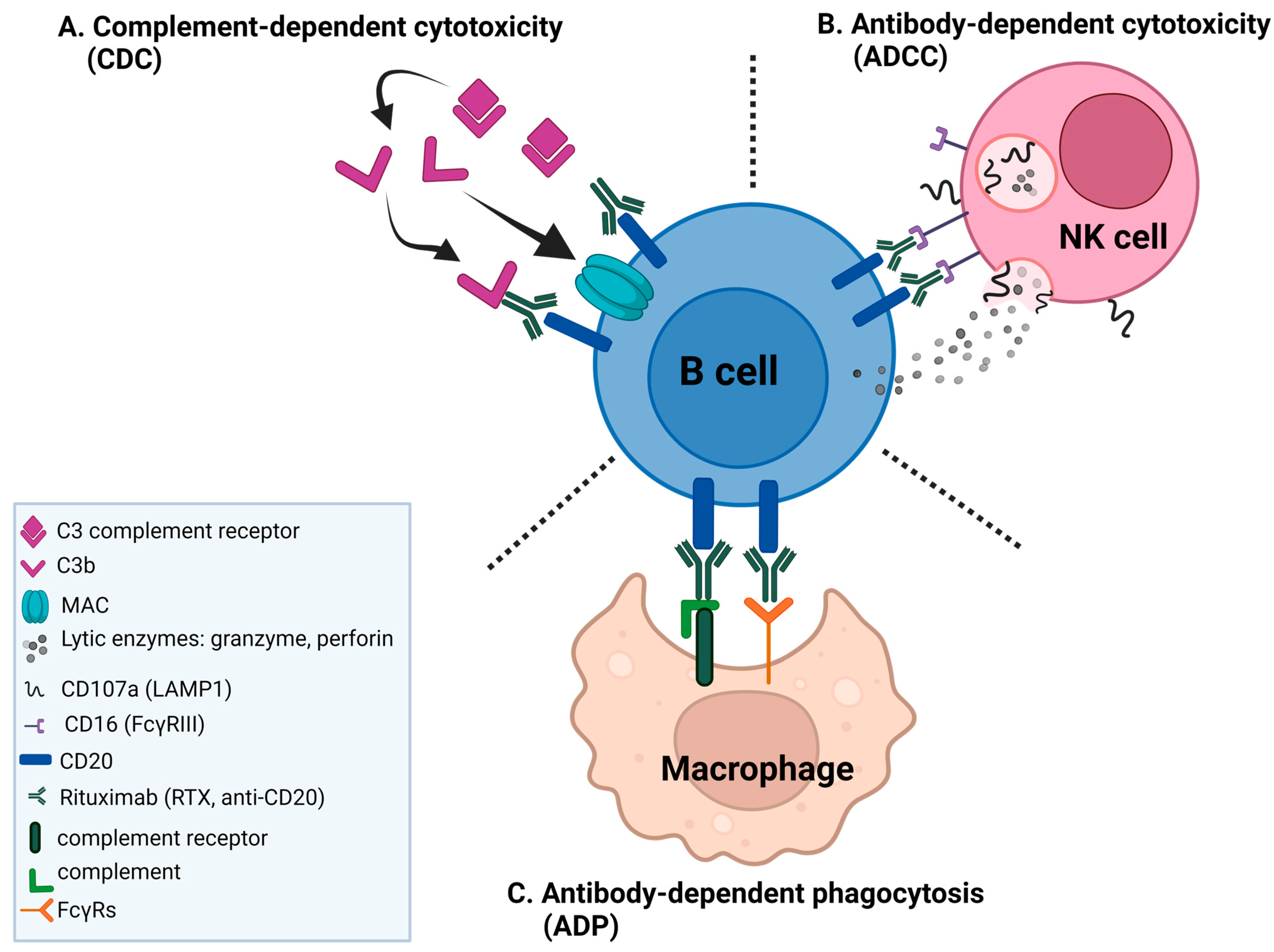
:1. Introduction
2. Results
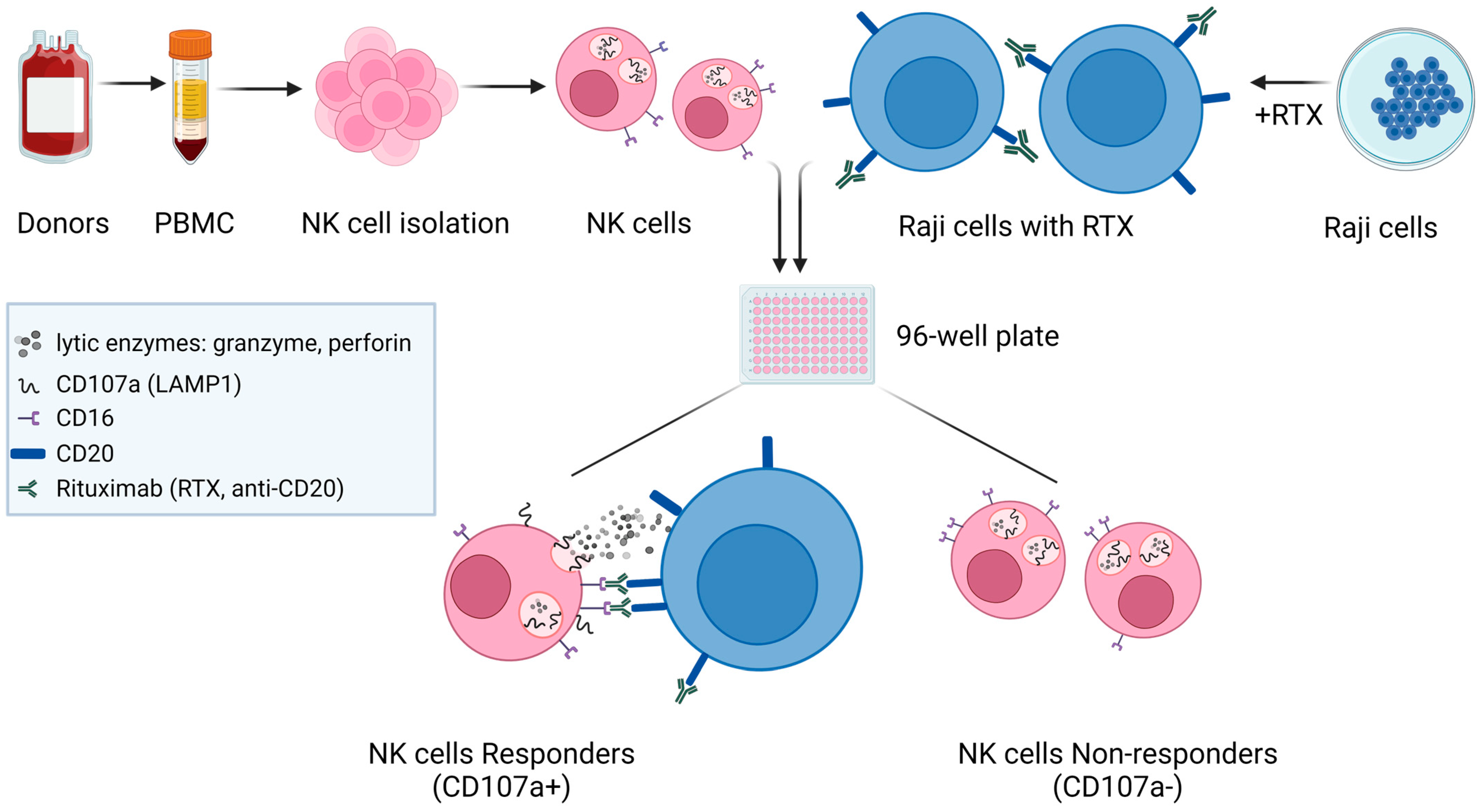
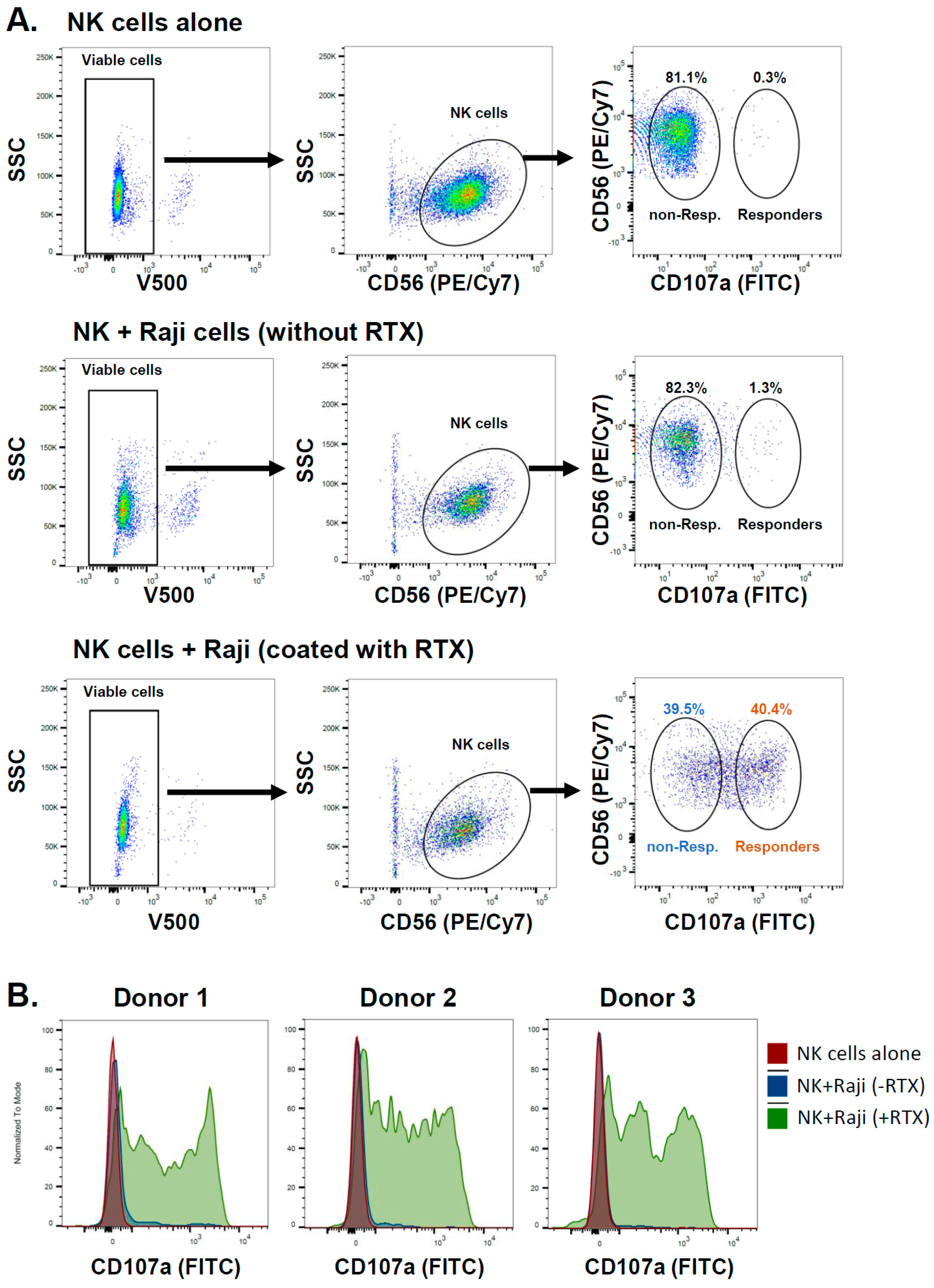
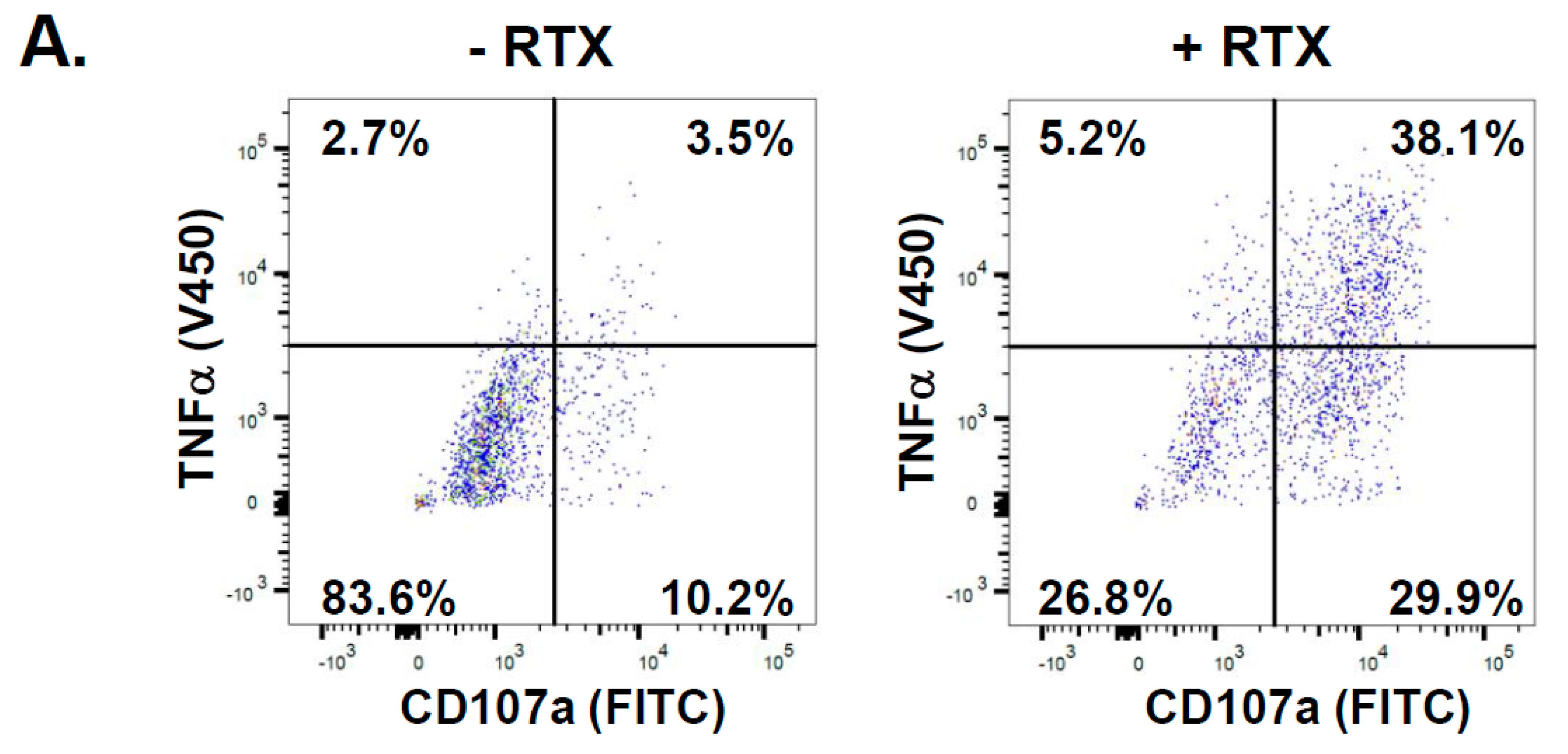
2.1. NK Cells Exposed to Rituximab-Coated Lymphoma Cells Degranulate Non-Uniformly
2.2. Analysis of CD16-Dependent Transcriptional Changes in Primary NK Cells
2.3. KIT and TNFSF4 Genes Are Down-Regulated upon CD16 Stimulation
3. Discussion
4. Materials and Methods
4.1. NK Cell Isolation
4.2. Burkitt Lymphoma Cell Line Culture
4.3. NK Cell Degranulation Analysis and Sorting
4.4. Intracellular TNFα Analysis
4.5. RNA-Seq and Bioinformatics Analysis
4.6. cDNA Synthesis and Quantitative Real-Time PCR
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Imai, K.; Matsuyama, S.; Miyake, S.; Suga, K.; Nakachi, K. Natural cytotoxic activity of peripheral-blood lymphocytes and cancer incidence: An 11-year follow-up study of a general population. Lancet 2000, 356, 1795–1799. [Google Scholar] [CrossRef] [PubMed]
- Veuillen, C.; Aurran-Schleinitz, T.; Castellano, R.; Rey, J.; Mallet, F.; Orlanducci, F.; Pouyet, L.; Just-Landi, S.; Coso, D.; Ivanov, V.; et al. Primary B-CLL resistance to NK cell cytotoxicity can be overcome in vitro and in vivo by priming NK cells and monoclonal antibody therapy. J. Clin. Immunol. 2012, 32, 632–646. [Google Scholar] [CrossRef]
- Pierpont, T.M.; Limper, C.B.; Richards, K.L. Past, Present, and Future of Rituximab-The World’s First Oncology Monoclonal Antibody Therapy. Front. Oncol. 2018, 8, 163. [Google Scholar] [CrossRef]
- Prica, A.; Baldassarre, F.; Hicks, L.K.; Imrie, K.; Kouroukis, T.; Cheung, M.; Members of the Hematology Disease Site Group of the Cancer Care Ontario Program in Evidence-Based Care. Rituximab in Lymphoma and Chronic Lymphocytic Leukaemia: A Practice Guideline. Clin. Oncol. 2017, 29, e13–e28. [Google Scholar] [CrossRef]
- Salles, G.; Barrett, M.; Foa, R.; Maurer, J.; O’Brien, S.; Valente, N.; Wenger, M.; Maloney, D.G. Rituximab in B-Cell Hematologic Malignancies: A Review of 20 Years of Clinical Experience. Adv. Ther. 2017, 34, 2232–2273. [Google Scholar] [CrossRef] [PubMed]
- Golay, J.; Introna, M. Mechanism of action of therapeutic monoclonal antibodies: Promises and pitfalls of in vitro and in vivo assays. Arch. Biochem. Biophys. 2012, 526, 146–153. [Google Scholar] [CrossRef] [PubMed]
- Grandjean, C.L.; Garcia, Z.; Lemaitre, F.; Breart, B.; Bousso, P. Imaging the mechanisms of anti-CD20 therapy in vivo uncovers spatiotemporal bottlenecks in antibody-dependent phagocytosis. Sci. Adv. 2021, 7, eabd6167. [Google Scholar] [CrossRef] [PubMed]
- Abulayha, A.; Bredan, A.; El Enshasy, H.; Daniels, I. Rituximab: Modes of action, remaining dispute and future perspective. Future Oncol. 2014, 10, 2481–2492. [Google Scholar] [CrossRef]
- Enqvist, M.; Jacobs, B.; Junlen, H.R.; Schaffer, M.; Melen, C.M.; Friberg, D.; Wahlin, B.E.; Malmberg, K.J. Systemic and Intra-Nodal Activation of NK Cells After Rituximab Monotherapy for Follicular Lymphoma. Front. Immunol. 2019, 10, 2085. [Google Scholar] [CrossRef]
- Nimmerjahn, F.; Gordan, S.; Lux, A. FcgammaR dependent mechanisms of cytotoxic, agonistic, and neutralizing antibody activities. Trends Immunol. 2015, 36, 325–336. [Google Scholar] [CrossRef]
- Capuano, C.; Pighi, C.; Battella, S.; De Federicis, D.; Galandrini, R.; Palmieri, G. Harnessing CD16-Mediated NK Cell Functions to Enhance Therapeutic Efficacy of Tumor-Targeting mAbs. Cancers 2021, 13, 2500. [Google Scholar] [CrossRef]
- Nagler-Anderson, C.; Bober, L.A.; Robinson, M.E.; Siskind, G.W.; Thorbecke, G.J. Suppression of type II collagen-induced arthritis by intragastric administration of soluble type II collagen. Proc. Natl. Acad. Sci. USA 1986, 83, 7443–7446. [Google Scholar] [CrossRef]
- Bryceson, Y.T.; March, M.E.; Ljunggren, H.G.; Long, E.O. Activation, coactivation, and costimulation of resting human natural killer cells. Immunol. Rev. 2006, 214, 73–91. [Google Scholar] [CrossRef] [PubMed]
- Leibson, P.J. Signal transduction during natural killer cell activation: Inside the mind of a killer. Immunity 1997, 6, 655–661. [Google Scholar] [CrossRef] [PubMed]
- Zompi, S.; Hamerman, J.A.; Ogasawara, K.; Schweighoffer, E.; Tybulewicz, V.L.; Di Santo, J.P.; Lanier, L.L.; Colucci, F. NKG2D triggers cytotoxicity in mouse NK cells lacking DAP12 or Syk family kinases. Nat. Immunol. 2003, 4, 565–572. [Google Scholar] [CrossRef] [PubMed]
- Bryceson, Y.T.; March, M.E.; Ljunggren, H.G.; Long, E.O. Synergy among receptors on resting NK cells for the activation of natural cytotoxicity and cytokine secretion. Blood 2006, 107, 159–166. [Google Scholar] [CrossRef] [PubMed]
- Lanier, L.L. Up on the tightrope: Natural killer cell activation and inhibition. Nat. Immunol. 2008, 9, 495–502. [Google Scholar] [CrossRef] [PubMed]
- Prevodnik, V.K.; Lavrencak, J.; Horvat, M.; Novakovic, B.J. The predictive significance of CD20 expression in B-cell lymphomas. Diagn. Pathol. 2011, 6, 33. [Google Scholar] [CrossRef] [PubMed]
- Hiraga, J.; Tomita, A.; Sugimoto, T.; Shimada, K.; Ito, M.; Nakamura, S.; Kiyoi, H.; Kinoshita, T.; Naoe, T. Down-regulation of CD20 expression in B-cell lymphoma cells after treatment with rituximab-containing combination chemotherapies: Its prevalence and clinical significance. Blood 2009, 113, 4885–4893. [Google Scholar] [CrossRef] [PubMed]
- Mishima, Y.; Terui, Y.; Takeuchi, K.; Matsumoto-Mishima, Y.; Matsusaka, S.; Utsubo-Kuniyoshi, R.; Hatake, K. The identification of irreversible rituximab-resistant lymphoma caused by CD20 gene mutations. Blood Cancer J. 2011, 1, e15. [Google Scholar] [CrossRef]
- Terui, Y.; Mishima, Y.; Sugimura, N.; Kojima, K.; Sakurai, T.; Mishima, Y.; Kuniyoshi, R.; Taniyama, A.; Yokoyama, M.; Sakajiri, S.; et al. Identification of CD20 C-terminal deletion mutations associated with loss of CD20 expression in non-Hodgkin’s lymphoma. Clin. Cancer Res. 2009, 15, 2523–2530. [Google Scholar] [CrossRef] [PubMed]
- Wu, J.; Mishra, H.K.; Walcheck, B. Role of ADAM17 as a regulatory checkpoint of CD16A in NK cells and as a potential target for cancer immunotherapy. J. Leukoc. Biol. 2019, 105, 1297–1303. [Google Scholar] [CrossRef] [PubMed]
- Romee, R.; Foley, B.; Lenvik, T.; Wang, Y.; Zhang, B.; Ankarlo, D.; Luo, X.; Cooley, S.; Verneris, M.; Walcheck, B.; et al. NK cell CD16 surface expression and function is regulated by a disintegrin and metalloprotease-17 (ADAM17). Blood 2013, 121, 3599–3608. [Google Scholar] [CrossRef] [PubMed]
- Cox, M.C.; Battella, S.; La Scaleia, R.; Pelliccia, S.; Di Napoli, A.; Porzia, A.; Cecere, F.; Alma, E.; Zingoni, A.; Mainiero, F.; et al. Tumor-associated and immunochemotherapy-dependent long-term alterations of the peripheral blood NK cell compartment in DLBCL patients. Oncoimmunology 2015, 4, e990773. [Google Scholar] [CrossRef] [PubMed]
- Stolz, C.; Schuler, M. Molecular mechanisms of resistance to Rituximab and pharmacologic strategies for its circumvention. Leuk. Lymphoma 2009, 50, 873–885. [Google Scholar] [CrossRef]
- Andre, P.; Denis, C.; Soulas, C.; Bourbon-Caillet, C.; Lopez, J.; Arnoux, T.; Blery, M.; Bonnafous, C.; Gauthier, L.; Morel, A.; et al. Anti-NKG2A mAb Is a Checkpoint Inhibitor that Promotes Anti-tumor Immunity by Unleashing Both T and NK Cells. Cell 2018, 175, 1731–1743.e13. [Google Scholar] [CrossRef]
- Krzewski, K.; Gil-Krzewska, A.; Nguyen, V.; Peruzzi, G.; Coligan, J.E. LAMP1/CD107a is required for efficient perforin delivery to lytic granules and NK-cell cytotoxicity. Blood 2013, 121, 4672–4683. [Google Scholar] [CrossRef]
- Wang, R.; Jaw, J.J.; Stutzman, N.C.; Zou, Z.; Sun, P.D. Natural killer cell-produced IFN-gamma and TNF-alpha induce target cell cytolysis through up-regulation of ICAM-1. J. Leukoc. Biol. 2012, 91, 299–309. [Google Scholar] [CrossRef]
- Khan, A.U.H.; Ali, A.K.; Marr, B.; Jo, D.; Ahmadvand, S.; Fong-McMaster, C.; Almutairi, S.M.; Wang, L.; Sad, S.; Harper, M.E.; et al. The TNFalpha/TNFR2 axis mediates natural killer cell proliferation by promoting aerobic glycolysis. Cell Mol. Immunol. 2023, 20, 1140–1155. [Google Scholar] [CrossRef]
- Bi, J.; Wang, X. Molecular Regulation of NK Cell Maturation. Front. Immunol. 2020, 11, 1945. [Google Scholar] [CrossRef]
- Wang, D.; Malarkannan, S. Transcriptional Regulation of Natural Killer Cell Development and Functions. Cancers 2020, 12, 1591. [Google Scholar] [CrossRef] [PubMed]
- Colucci, F.; Di Santo, J.P. The receptor tyrosine kinase c-kit provides a critical signal for survival, expansion, and maturation of mouse natural killer cells. Blood 2000, 95, 984–991. [Google Scholar] [CrossRef] [PubMed]
- Bosken, B.; Hepner-Schefczyk, M.; Vonderhagen, S.; Dudda, M.; Flohe, S.B. An Inverse Relationship Between c-Kit/CD117 and mTOR Confers NK Cell Dysregulation Late After Severe Injury. Front. Immunol. 2020, 11, 1200. [Google Scholar] [CrossRef] [PubMed]
- Lee, Y.; Jung, J.; Cho, K.J.; Lee, S.K.; Park, J.W.; Oh, I.H.; Kim, G.J. Increased SCF/c-kit by hypoxia promotes autophagy of human placental chorionic plate-derived mesenchymal stem cells via regulating the phosphorylation of mTOR. J. Cell Biochem. 2013, 114, 79–88. [Google Scholar] [CrossRef] [PubMed]
- Kweon, S.; Phan, M.T.; Chun, S.; Yu, H.; Kim, J.; Kim, S.; Lee, J.; Ali, A.K.; Lee, S.H.; Kim, S.K.; et al. Expansion of Human NK Cells Using K562 Cells Expressing OX40 Ligand and Short Exposure to IL-21. Front. Immunol. 2019, 10, 879. [Google Scholar] [CrossRef] [PubMed]
- Ward-Kavanagh, L.K.; Lin, W.W.; Sedy, J.R.; Ware, C.F. The TNF Receptor Superfamily in Co-stimulating and Co-inhibitory Responses. Immunity 2016, 44, 1005–1019. [Google Scholar] [CrossRef]
- Guo, M.; Sun, C.; Qian, Y.; Zhu, L.; Ta, N.; Wang, G.; Zheng, J.; Guo, F.; Liu, Y. Proliferation of Highly Cytotoxic Human Natural Killer Cells by OX40L Armed NK-92 With Secretory Neoleukin-2/15 for Cancer Immunotherapy. Front. Oncol. 2021, 11, 632540. [Google Scholar] [CrossRef]
- Eissner, G.; Kolch, W.; Scheurich, P. Ligands working as receptors: Reverse signaling by members of the TNF superfamily enhance the plasticity of the immune system. Cytokine Growth Factor Rev. 2004, 15, 353–366. [Google Scholar] [CrossRef]
- Nuebling, T.; Schumacher, C.E.; Hofmann, M.; Hagelstein, I.; Schmiedel, B.J.; Maurer, S.; Federmann, B.; Rothfelder, K.; Roerden, M.; Dorfel, D.; et al. The Immune Checkpoint Modulator OX40 and Its Ligand OX40L in NK-Cell Immunosurveillance and Acute Myeloid Leukemia. Cancer Immunol. Res. 2018, 6, 209–221. [Google Scholar] [CrossRef]
- Hwang, S.; Lim, J.; Kang, H.; Jeong, J.Y.; Joung, J.G.; Heo, J.; Jung, D.; Cho, K.; An, H.J. Predictive biomarkers for the responsiveness of recurrent glioblastomas to activated killer cell immunotherapy. Cell Biosci. 2023, 13, 17. [Google Scholar] [CrossRef] [PubMed]
- Roszik, J.; Markovits, E.; Dobosz, P.; Layani, A.; Slabodnik-Kaner, K.; Baruch, E.N.; Ben-Betzalel, G.; Grimm, E.; Berger, R.; Sidi, Y.; et al. TNFSF4 (OX40L) expression and survival in locally advanced and metastatic melanoma. Cancer Immunol. Immunother. 2019, 68, 1493–1500. [Google Scholar] [CrossRef]
- Wang, Q.W.; Lin, W.W.; Zhu, Y.J. Comprehensive analysis of a TNF family based-signature in diffuse gliomas with regard to prognosis and immune significance. Cell Commun. Signal. 2022, 20, 6. [Google Scholar] [CrossRef] [PubMed]
- Turaj, A.H.; Cox, K.L.; Penfold, C.A.; French, R.R.; Mockridge, C.I.; Willoughby, J.E.; Tutt, A.L.; Griffiths, J.; Johnson, P.W.M.; Glennie, M.J.; et al. Augmentation of CD134 (OX40)-dependent NK anti-tumour activity is dependent on antibody cross-linking. Sci. Rep. 2018, 8, 2278. [Google Scholar] [CrossRef]
- Abdel-Magid, A.F. The Potential of c-KIT Kinase inhibitors in Cancer Treatment. ACS Med. Chem. Lett. 2021, 12, 1191–1192. [Google Scholar] [CrossRef] [PubMed]
- Godesi, S.; Lee, J.; Nada, H.; Quan, G.; Elkamhawy, A.; Choi, Y.; Lee, K. Small Molecule c-KIT Inhibitors for the Treatment of Gastrointestinal Stromal Tumors: A Review on Synthesis, Design Strategies, and Structure-Activity Relationship (SAR). Int. J. Mol. Sci. 2023, 24, 9450. [Google Scholar] [CrossRef] [PubMed]
- Le, A.M.; Torres, T. OX40-OX40L Inhibition for the Treatment of Atopic Dermatitis-Focus on Rocatinlimab and Amlitelimab. Pharmaceutics 2022, 14, 2753. [Google Scholar] [CrossRef]




{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Non-Responders (% Parent) | Responders (% Parent) | |
|---|---|---|
| Donor 1 | 39.5 | 40.4 |
| Donor 2 | 38.2 | 45.1 |
| Donor 3 | 38.3 | 41.2 |
| Donor 4 | 50.5 | 24.7 |
| Donor 5 | 48.3 | 34.4 |
| Donor 6 | 23.7 | 49.2 |
| Target | Forward Primer | Reverse Primer |
|---|---|---|
| KIT | ACCAACACCGGCAAATACA | AAGCTTGGCAGGATCTCTAAC |
| TNFSF4 | TTCCAACTGAAGAAGGTCAG | GAAGTCATCCAGGGAGGTAT |
| GAPDH | AGCCACATCGCTCAGACAC | GCCCAATACGACCAAATCC |
| 18S rRNA | GCAATTATTCCCCATGAACG | GGGACTTAATCAACGCAAGC |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Wlodarczyk, M.; Torun, A.; Zerrouqi, A.; Pyrzynska, B. NK Cell Degranulation Triggered by Rituximab Identifies Potential Markers of Subpopulations with Enhanced Cytotoxicity toward Malignant B Cells. Int. J. Mol. Sci. 2024, 25, 8980. https://doi.org/10.3390/ijms25168980
Wlodarczyk M, Torun A, Zerrouqi A, Pyrzynska B. NK Cell Degranulation Triggered by Rituximab Identifies Potential Markers of Subpopulations with Enhanced Cytotoxicity toward Malignant B Cells. International Journal of Molecular Sciences. 2024; 25(16):8980. https://doi.org/10.3390/ijms25168980
Chicago/Turabian StyleWlodarczyk, Marta, Anna Torun, Abdessamad Zerrouqi, and Beata Pyrzynska. 2024. "NK Cell Degranulation Triggered by Rituximab Identifies Potential Markers of Subpopulations with Enhanced Cytotoxicity toward Malignant B Cells" International Journal of Molecular Sciences 25, no. 16: 8980. https://doi.org/10.3390/ijms25168980