
Decoding LncRNA in COPD: Unveiling Prognostic and Diagnostic Power and Their Driving Role in Lung Cancer Progression
 , , , , , ,
, , , , , , 
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Chronological Progression of COPD
2.1. Disease Progression in COPD
2.2. The Major Causative Factors of COPD
2.3. Diagnosis and Monitoring of COPD
3. Exploring LncRNA Profiles and Clinical Significance in COPD
3.1. Identification and Profiling of COPD-Associated LncRNAs
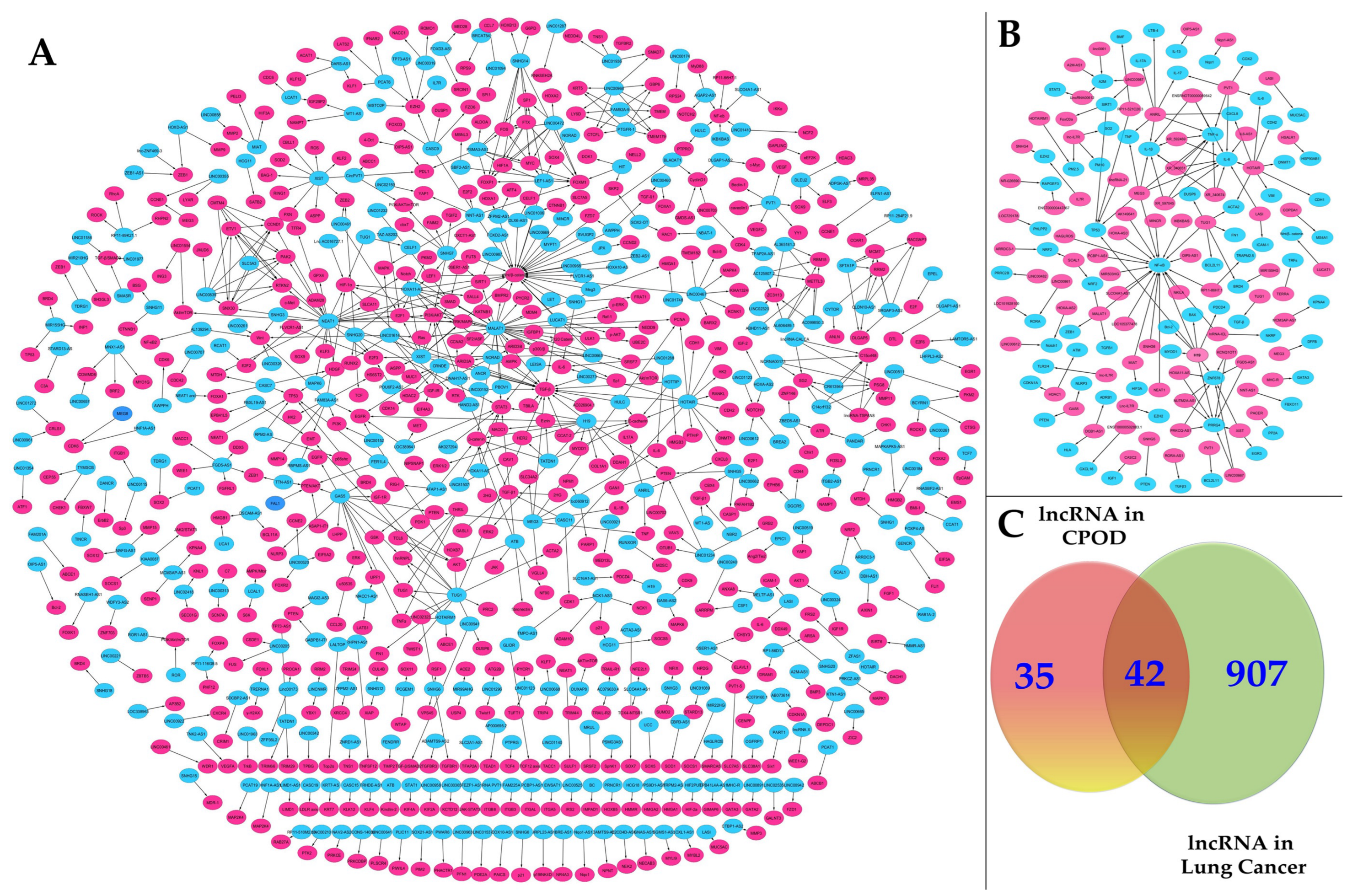
3.2. Master Regulators of Gene Expression in Lung Cancer Progression
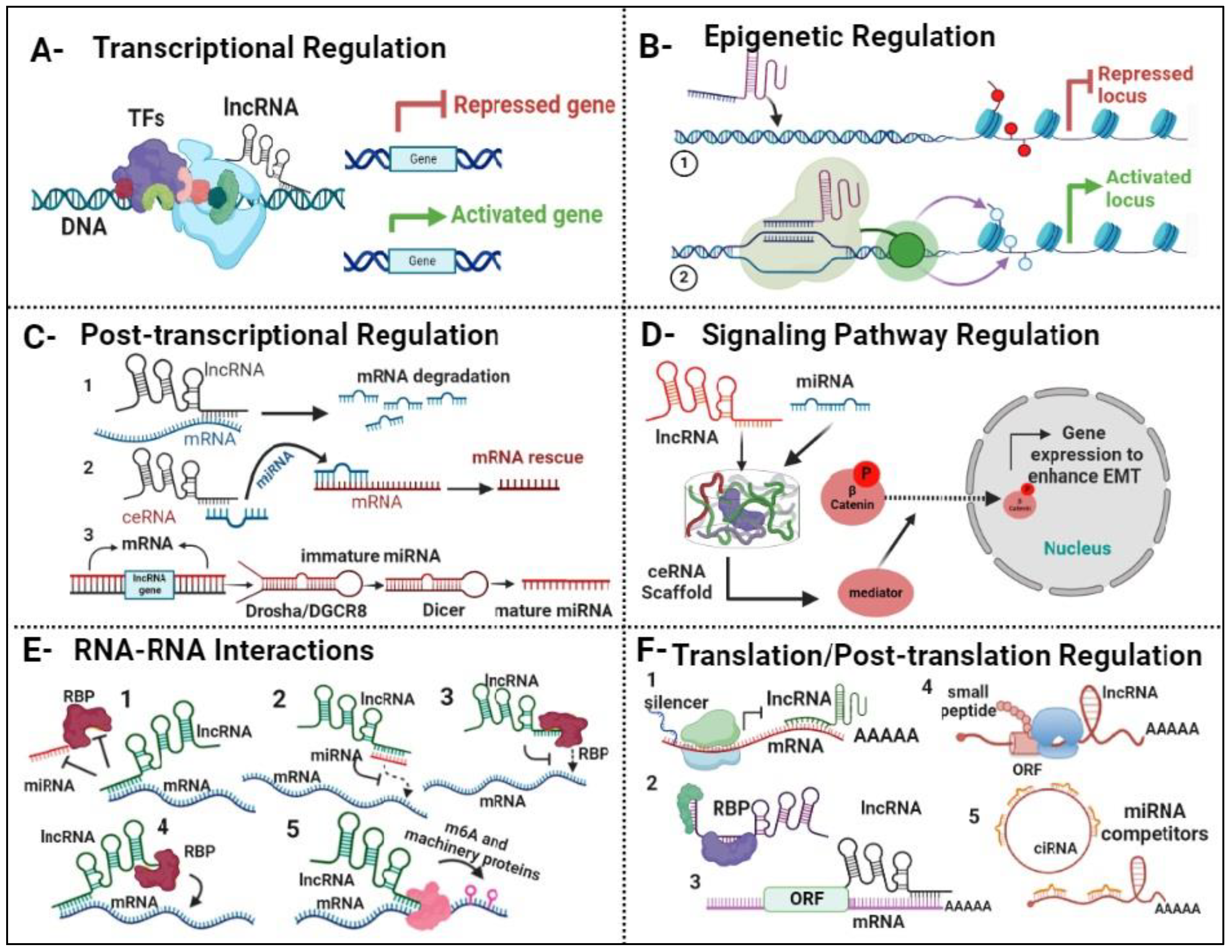
3.3. Tobacco Smoke Exposure and Expression Profiles of LncRNA in COPD

3.4. Correlation Between LncRNA Expression and COPD Severity
3.5. Prognostic and Diagnostic Value of LncRNA in Predicting COPD Outcomes
4. Regulatory Functions of LncRNAs in COPD
4.1. LncRNA-Mediated Immune Response in COPD
4.2. Complexities of LncRNA-Mediated Inflammation in COPD
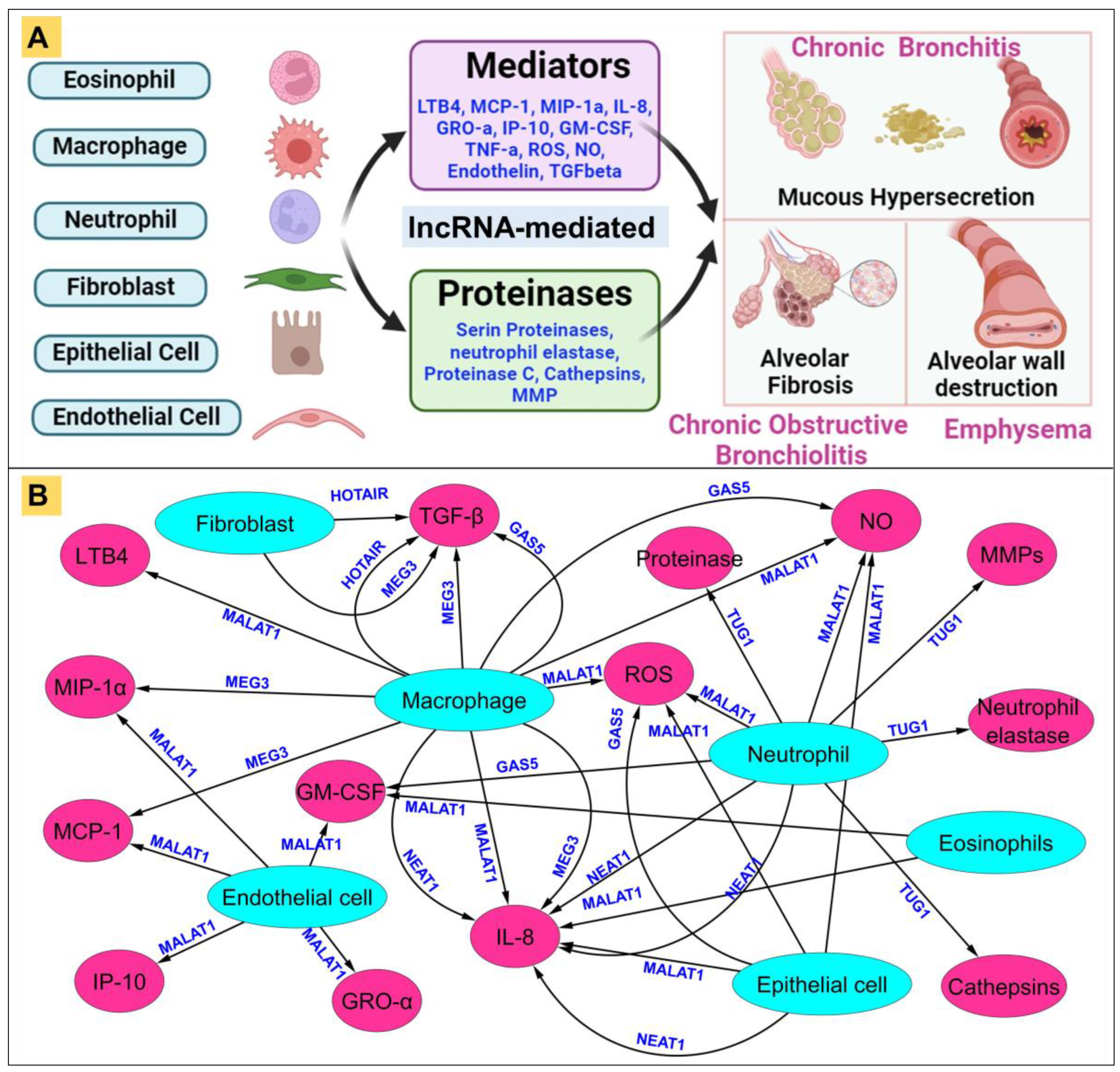
4.3. Modulation of Cellular Processes by LncRNA in COPD
5. Modulatory Mechanisms of COPD-Associated LncRNA in Lung Cancer Progression
5.1. Interaction Between COPD-Associated LncRNA and Lung Cancer Pathways
5.1.1. Shared Molecular Pathways
5.1.2. Regulatory Roles of COPD-Associated LncRNA in Lung Cancer
5.2. Dysregulated Expression of COPD-Associated LncRNAs in Lung Cancer
6. LncRNA in Body Fluids for COPD Diagnosis and Therapy
6.1. Detection of LncRNA Biomarkers in Blood Samples
6.2. Urinary LncRNA as a Diagnostic Tool for COPD
6.3. Pharmacological Actions of LncRNA Molecules as Potential Therapeutics for COPD
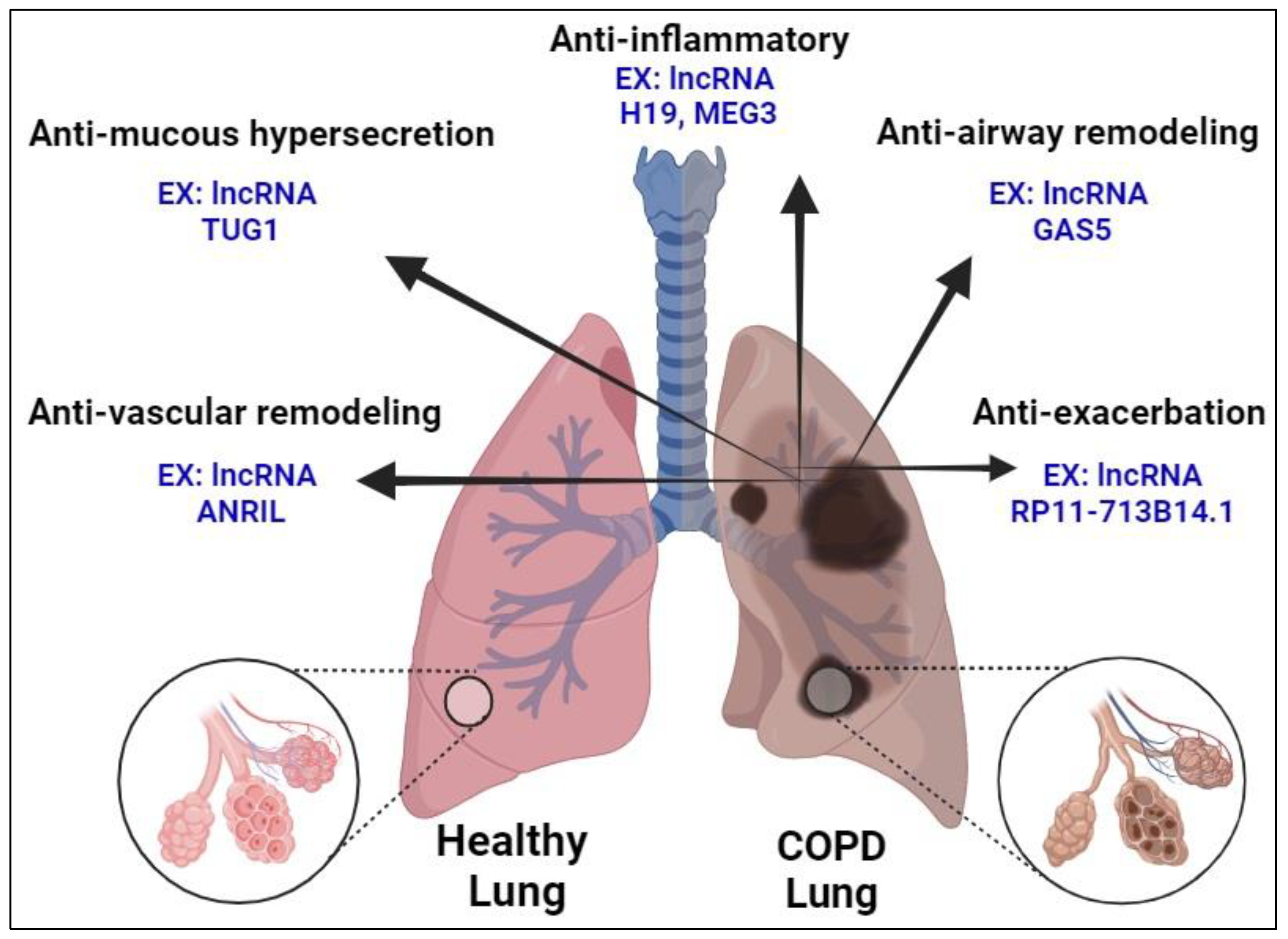
6.4. Potential Therapeutic Applications of LncRNA in COPD
7. Conclusions and Future Directions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Global Initiative for Chronic Obstructive Lung Disease (GOLD). Global Strategy for the Diagnosis, Management, and Prevention of Chronic Obstructive Pulmonary Disease. 2021. Available online: https://goldcopd.org/gold-reports/ (accessed on 18 January 2024).
- Soriano, J.B.; Kendrick, P.J.; Paulson, K.R.; Gupta, V.; Abrams, E.M.; Adedoyin, R.A.; Adhikari, T.B.; Advani, S.M.; Agrawal, A.; Moradi, M.; et al. Prevalence and Attributable Health Burden of Chronic Respiratory Diseases, 1990–2017: A Systematic Analysis for the Global Burden of Disease Study 2017. Lancet Respir. Med. 2020, 8, 585–596. [Google Scholar] [CrossRef] [PubMed]
- Gurer, D.C.; Erdogan, İ.; Ahmadov, U.; Basol, M.; Sweef, O.; Cakan-Akdogan, G.; Akgül, B. Transcriptomics Profiling Identifies Cisplatin-Inducible Death Receptor 5 Antisense Long Non-coding RNA as a Modulator of Proliferation and Metastasis in HeLa Cells. Front. Cell Dev. Biol. 2021, 9, 688855. [Google Scholar] [CrossRef] [PubMed] [PubMed Central]
- Quinn, J.J.; Chang, H.Y. Unique Features of Long Non-Coding RNA Biogenesis and Function. Nat. Rev. Genet. 2016, 17, 47–62. [Google Scholar] [CrossRef] [PubMed]
- Mattick, J.S.; Amaral, P.P.; Carninci, P.; Carpenter, S.; Chang, H.Y.; Chen, L.L.; Chen, R.; Dean, C.; Dinger, M.E.; Fitzgerald, K.A.; et al. Long non-coding RNAs: Definitions, functions, challenges and recommendations. Nat. Rev. Mol. Cell Biol. 2023, 24, 430–447. [Google Scholar] [CrossRef] [PubMed] [PubMed Central]
- Zhou, T.; Ding, J.W.; Wang, X.A. Dysregulation of lncRNAs and mRNAs Expression in the Development of Chronic Obstructive Pulmonary Disease. J. Cell. Biochem. 2018, 119, 4600–4610. [Google Scholar] [CrossRef]
- de-Torres, J.P.; Wilson, D.O.; Sanchez-Salcedo, P.; Weissfeld, J.L.; Berto, J.; Campo, A.; Zulueta, J.J. Lung Cancer in Patients with Chronic Obstructive Pulmonary Disease: Development and Validation of the COPD Lung Cancer Screening Score. Am. J. Respir. Crit. Care Med. 2015, 191, 285–291. [Google Scholar] [CrossRef]
- Durham, A.L.; Adcock, I.M. The Relationship between COPD and Lung Cancer. Lung Cancer 2015, 90, 121–127. [Google Scholar] [CrossRef] [PubMed]
- Erdoğan, İ.; Sweef, O.; Akgül, B. Long Noncoding RNAs in Human Cancer and Apoptosis. Curr. Pharm. Biotechnol. 2023, 24, 872–888. [Google Scholar] [CrossRef] [PubMed]
- Walther, K.; Schulte, L.N. The Role of lncRNAs in Innate Immunity and Inflammation. RNA Biol. 2021, 18, 587–603. [Google Scholar] [CrossRef]
- Xie, J.; Wu, Y.; Tao, Q.; Liu, H.; Wang, J.; Zhang, C.; Zhou, Y.; Wei, C.; Chang, Y.; Jin, Y.; et al. The Role of lncRNA in the Pathogenesis of Chronic Obstructive Pulmonary Disease. Heliyon 2023, 9, e22460. [Google Scholar] [CrossRef]
- Agusti, A.; Edwards, L.D.; Rennard, S.I.; MacNee, W.; Tal-Singer, R.; Miller, B.E. Persistent systemic inflammation is associated with poor clinical outcomes in COPD: A novel phenotype. PLoS ONE 2012, 7, e37483. [Google Scholar] [CrossRef] [PubMed]
- Barnes, P.J. Inflammatory mechanisms in patients with chronic obstructive pulmonary disease. J. Allergy Clin. Immunol. 2016, 138, 16–27. [Google Scholar] [CrossRef] [PubMed]
- Vogelmeier, C.F.; Criner, G.J.; Martinez, F.J.; Anzueto, A.; Barnes, P.J.; Bourbeau, J.; Agusti, A. Global Strategy for the Diagnosis, Management, and Prevention of Chronic Obstructive Lung Disease 2017 Report. GOLD Executive Summary. Am. J. Respir. Crit. Care Med. 2017, 195, 557. [Google Scholar] [CrossRef] [PubMed]
- Wedzicha, J.A.; Seemungal, T.A. COPD exacerbations: Defining their cause and prevention. Lancet 2007, 370, 786–796. [Google Scholar] [CrossRef]
- Celli, B.R.; MacNee, W. ATS/ERS Task Force. Standards for the diagnosis and treatment of patients with COPD: A summary of the ATS/ERS position paper. Eur. Respir. J. 2004; 23, 932–946. [Google Scholar] [CrossRef]
- Ko, F.W.; Chan, K.P.; Hui, D.S.; Goddard, J.R.; Shaw, J.G.; Reid, D.W.; Yang, I.A. Acute exacerbation of COPD. Respirology 2016, 21, 1152–1165. [Google Scholar] [CrossRef] [PubMed]
- Pellegrino, R.; Viegi, G.; Brusasco, V.; Crapo, R.O.; Burgos, F.; Casaburi, R.E.A.; Wanger, J. Interpretative strategies for lung function tests. Eur. Respir. J. 2005, 26, 948–968. [Google Scholar] [CrossRef] [PubMed]
- Celli, B.R.; Cote, C.G.; Marin, J.M.; Casanova, C.; Montes de Oca, M.; Mendez, R.A.; Cabral, H.J. The Body-Mass Index, Airflow Obstruction, Dyspnea, and Exercise Capacity Index in Chronic Obstructive Pulmonary Disease. N. Engl. J. Med. 2004, 350, 1005–1012. [Google Scholar] [CrossRef] [PubMed]
- Sin, D.D.; Miller, B.E.; Duvoix, A.; Man, S.P.; Zhang, X.; Silverman, E.K.; Lomas, D.A. Serum PARC/CCL-18 concentrations and health outcomes in chronic obstructive pulmonary disease. Am. J. Respir. Crit. Care Med. 2011, 183, 1187–1192. [Google Scholar] [CrossRef] [PubMed]
- Sweef, O.; Yang, C.; Wang, Z. The Oncogenic and Tumor Suppressive Long Non-Coding RNA-microRNA-Messenger RNA Regulatory Axes Identified by Analyzing Multiple Platform Omics Data from Cr(VI)-Transformed Cells and Their Implications in Lung Cancer. Biomedicines 2022, 10, 2334. [Google Scholar] [CrossRef] [PubMed] [PubMed Central]
- Scoditti, E.; Massaro, M.; Garbarino, S.; Toraldo, D.M. Role of Diet in Chronic Obstructive Pulmonary Disease Prevention and Treatment. Nutrients 2019, 11, 1357. [Google Scholar] [CrossRef]
- Byanova, K.L.; Abelman, R.; North, C.M.; Christenson, S.A.; Huang, L. COPD in People with HIV: Epidemiology, Pathogenesis, Management, and Prevention Strategies. Int. J. Chron. Obstruct. Pulmon. Dis. 2023, 18, 2795–2817. [Google Scholar] [CrossRef]
- Schleich, F.; Bougard, N.; Moermans, C.; Sabbe, M.; Louis, R. Cytokine-targeted therapies for asthma and COPD. Eur. Respir. Rev. 2023, 32, 220193. [Google Scholar] [CrossRef] [PubMed]
- Kaur, M.; Chandel, J.; Malik, J.; Naura, A.S. Particulate matter in COPD pathogenesis: An overview. Inflamm. Res. 2022, 71, 797–815. [Google Scholar] [CrossRef] [PubMed]
- Sin, D.D.; Doiron, D.; Agusti, A.; Anzueto, A.; Barnes, P.J.; Celli, B.R.; Criner, G.J.; Halpin, D.; Han, M.K.; Martinez, F.J.; et al. Air pollution and COPD: GOLD 2023 committee report. Eur. Respir. J. 2023, 61, 2202469. [Google Scholar] [CrossRef] [PubMed]
- Neşe, A.; Samancıoğlu Bağlama, S. The Effect of Progressive Muscle Relaxation and Deep Breathing Exercises on Dyspnea and Fatigue Symptoms of COPD Patients: A Randomized Controlled Study. Holist. Nurs. Pract. 2022, 36, E18–E26. [Google Scholar] [CrossRef]
- Ntritsos, G.; Franek, J.; Belbasis, L.; Christou, M.A.; Markozannes, G.; Altman, P.; Fogel, R.; Sayre, T.; Ntzani, E.E.; Evangelou, E. Gender-specific estimates of COPD prevalence: A systematic review and meta-analysis. Int. J. Chron. Obstruct. Pulmon. Dis. 2018, 13, 1507–1514. [Google Scholar] [CrossRef] [PubMed]
- Segal, L.N.; Martinez, F.J. Chronic obstructive pulmonary disease subpopulations and phenotyping. J. Allergy Clin. Immunol. 2018, 141, 1961–1971. [Google Scholar] [CrossRef] [PubMed]
- Duffy, S.P.; Criner, G.J. Chronic Obstructive Pulmonary Disease: Evaluation and Management. Med. Clin. N. Am. 2019, 103, 453–461. [Google Scholar] [CrossRef]
- Bagdonas, E.; Raudoniute, J.; Bruzauskaite, I.; Aldonyte, R. Novel aspects of pathogenesis and regeneration mechanisms in COPD. Int. J. Chron. Obstruct. Pulmon. Dis. 2015, 10, 995–1013. [Google Scholar] [CrossRef]
- Wang, P.S.; Liu, Z.; Sweef, O.; Saeed, A.F.; Kluz, T.; Costa, M.; Shroyer, K.R.; Kondo, K.; Wang, Z.; Yang, C. Hexavalent chromium exposure activates the non-canonical nuclear factor kappa B pathway to promote immune checkpoint protein programmed death-ligand 1 expression and lung carcinogenesis. Cancer Lett. 2024, 589, 216827. [Google Scholar] [CrossRef] [PubMed]
- Murgia, N.; Gambelunghe, A. Occupational COPD-The most under-recognized occupational lung disease? Respirology 2022, 27, 399–410. [Google Scholar] [CrossRef]
- GBD 2019 Chronic Respiratory Diseases Collaborators. Global burden of chronic respiratory diseases and risk factors, 1990–2019: An update from the Global Burden of Disease Study 2019. EClinicalMedicine 2023, 59, 101936. [CrossRef]
- Rabe, K.F.; Watz, H. Chronic obstructive pulmonary disease. Lancet 2017, 389, 1931–1940. [Google Scholar] [CrossRef]
- Lakshmanan, S.; Jankowich, M.; Wu, W.C.; Blackshear, C.; Abbasi, S.; Choudhary, G. Gender Differences in Risk Factors Associated with Pulmonary Artery Systolic Pressure, Heart Failure, and Mortality in Blacks: Jackson Heart Study. J. Am. Heart Assoc. 2020, 9, e013034. [Google Scholar] [CrossRef] [PubMed]
- Yang, I.A.; Jenkins, C.R.; Salvi, S.S. Chronic obstructive pulmonary disease in never-smokers: Risk factors, pathogenesis, and implications for prevention and treatment. Lancet Respir. Med. 2022, 10, 497–511. [Google Scholar] [CrossRef]
- Mannino, D.M.; Buist, A.S. Global burden of COPD: Risk factors, prevalence, and future trends. Lancet 2007, 370, 765–773. [Google Scholar] [CrossRef] [PubMed]
- Salvi, S.S.; Barnes, P.J. Chronic obstructive pulmonary disease in non-smokers. Lancet 2009, 374, 733–743. [Google Scholar] [CrossRef]
- Easter, M.; Bollenbecker, S.; Barnes, J.W.; Krick, S. Targeting Aging Pathways in Chronic Obstructive Pulmonary Disease. Int. J. Mol. Sci. 2020, 21, 6924. [Google Scholar] [CrossRef] [PubMed]
- Li, J.; Sun, S.; Tang, R.; Qiu, H.; Huang, Q.; Mason, T.G.; Tian, L. Major air pollutants and risk of COPD exacerbations: A systematic review and meta-analysis. Int. J. Chron. Obstruct. Pulmon. Dis. 2016, 11, 3079–3091. [Google Scholar] [CrossRef]
- Wang, P.S.; Liu, Z.; Sweef, O.; Xie, J.; Chen, J.; Zhu, H.; Zeidler-Erdely, P.C.; Yang, C.; Wang, Z. Long noncoding RNA ABHD11-AS1 interacts with SART3 and regulates CD44 RNA alternative splicing to promote lung carcinogenesis. Environ. Int. 2024, 185, 108494. [Google Scholar] [CrossRef] [PubMed]
- Bekkat-Berkani, R.; Wilkinson, T.; Buchy, P.; Dos Santos, G.; Stefanidis, D.; Devaster, J.M. Seasonal influenza vaccination in patients with COPD: A systematic literature review. BMC Pulm. Med. 2017, 17, 79. [Google Scholar] [CrossRef] [PubMed]
- Odimba, U.; Senthilselvan, A.; Farrell, J.; Gao, Z. Current Knowledge of Asthma-COPD Overlap (ACO) Genetic Risk Factors, Characteristics, and Prognosis. COPD J. Chronic Obstr. Pulm. Dis. 2021, 18, 585–595. [Google Scholar] [CrossRef]
- Silverman, E.K. Genetics of COPD. Annu. Rev. Physiol. 2020, 82, 413–431. [Google Scholar] [CrossRef]
- Huang, X.; Mu, X.; Deng, L.; Fu, A.; Pu, E.; Tang, T.; Kong, X. The etiologic origins for chronic obstructive pulmonary disease. Int. J. Chron. Obstruct. Pulmon. Dis. 2019, 14, 1139–1158. [Google Scholar] [CrossRef]
- Kahnert, K.; Jörres, R.A.; Behr, J.; Welte, T. The Diagnosis and Treatment of COPD and Its Comorbidities. Dtsch. Arztebl. Int. 2023, 120, 434–444. [Google Scholar] [CrossRef]
- Şerifoğlu, İ.; Ulubay, G. The methods other than spirometry in the early diagnosis of COPD. Tuberk Toraks 2019, 67, 63–70. [Google Scholar] [CrossRef]
- Huprikar, N.A.; Skabelund, A.J.; Bedsole, V.G.; Sjulin, T.J.; Karandikar, A.V.; Aden, J.K.; Morris, M.J. Comparison of Forced and Slow Vital Capacity Maneuvers in Defining Airway Obstruction. Respir. Care 2019, 64, 786–792. [Google Scholar] [CrossRef]
- Yuan, W.; He, X.; Xu, Q.F.; Wang, H.Y.; Casaburi, R. Increased difference between slow and forced vital capacity is associated with reduced exercise tolerance in COPD patients. BMC Pulm. Med. 2014, 14, 16. [Google Scholar] [CrossRef]
- Christenson, S.A.; Smith, B.M.; Bafadhel, M.; Putcha, N. Chronic obstructive pulmonary disease. Lancet 2022, 399, 2227–2242. [Google Scholar] [CrossRef]
- van den Bemt, L.; Schermer, T.; Smeele, I.; Bischoff, E.; Jacobs, A.; Grol, R.; van Weel, C. Monitoring of patients with COPD: A review of current guidelines’ recommendations. Respir. Med. 2008, 102, 633–641. [Google Scholar] [CrossRef]
- Dasgupta, S.; Ghosh, N.; Bhattacharyya, P.; Roy Chowdhury, S.; Chaudhury, K. Metabolomics of asthma, COPD, and asthma-COPD overlap: An overview. Crit. Rev. Clin. Lab. Sci. 2023, 60, 153–170. [Google Scholar] [CrossRef]
- Wouters, E.F.; Posthuma, R.; Koopman, M.; Liu, W.Y.; Sillen, M.J.; Hajian, B.; Sastry, M.; Spruit, M.A.; Franssen, F.M. An Update on Pulmonary Rehabilitation Techniques for Patients with Chronic Obstructive Pulmonary Disease. Expert Rev. Respir. Med. 2020, 14, 149–161. [Google Scholar] [CrossRef] [PubMed]
- Lea, S.; Higham, A.; Beech, A.; Singh, D. How Inhaled Corticosteroids Target Inflammation in COPD. Eur. Respir. Rev. 2023, 32, 230084. [Google Scholar] [CrossRef] [PubMed] [PubMed Central]
- Lu, Z.; Van Eeckhoutte, H.P.; Liu, G.; Nair, P.M.; Jones, B.; Gillis, C.M.; Nalkurthi, B.C.; Verhamme, F.; Buyle-Huybrecht, T.; Vandenabeele, P.; et al. Necroptosis Signaling Promotes Inflammation, Airway Remodeling, and Emphysema in Chronic Obstructive Pulmonary Disease. Am. J. Respir. Crit. Care Med. 2021, 204, 667–681. [Google Scholar] [CrossRef] [PubMed]
- Thaiparambil, J.; Dong, L.; Jasso, D.; Huang, J.-A.; El-Zein, R.A. Mitotic Spindle Apparatus Abnormalities in Chronic Obstructive Pulmonary Disease Cells: A Potential Pathway to Lung Cancer. Cancer Prev. Res. 2020, 13, 923–934. [Google Scholar] [CrossRef]
- Brightling, C.; Greening, N. Airway Inflammation in COPD: Progress to Precision Medicine. Eur. Respir. J. 2019, 54, 1900651. [Google Scholar] [CrossRef] [PubMed]
- Lin, S.; Liu, C.; Sun, J.; Guan, Y. RNA-Sequencing and Bioinformatics Analysis of Exosomal Long Noncoding RNAs Revealed a Novel ceRNA Network in Stable COPD. Int. J. Chron. Obstruct. Pulmon. Dis. 2023, 18, 1995–2007. [Google Scholar] [CrossRef]
- Firoozi, Z.; Shahi, A.; Mohammadisoleimani, E.; Afzali, S.; Mansoori, B.; Bahmanyar, M.; Mohaghegh, P.; Dastsooz, H.; Pezeshki, B.; Nikfar, G.; et al. CircRNA-Associated ceRNA Networks (circCeNETs) in Chronic Obstructive Pulmonary Disease (COPD). Life Sci. 2024, 349, 122715. [Google Scholar] [CrossRef]
- Fan, P.; Zhang, Y.; Ding, S.; Du, Z.; Zhou, C.; Du, X. Integrating RNA-seq and scRNA-seq to explore the mechanism of macrophage ferroptosis associated with COPD. Front. Pharmacol. 2023, 14, 1139137. [Google Scholar] [CrossRef]
- Halpin, D.M.G.; Criner, G.J.; Papi, A.; Singh, D.; Anzueto, A.; Martinez, F.J.; Agusti, A.A.; Vogelmeier, C.F. Global Initiative for the Diagnosis, Management, and Prevention of Chronic Obstructive Lung Disease. The 2020 GOLD Science Committee Report on COVID-19 and Chronic Obstructive Pulmonary Disease. Am. J. Respir. Crit. Care Med. 2021, 203, 24–36. [Google Scholar] [CrossRef]
- Zhang, J.; Zhu, Y.; Wang, R. Long noncoding RNAs in respiratory diseases. Histol. Histopathol. 2018, 33, 747–756. [Google Scholar] [CrossRef]
- Poulet, C.; Njock, M.S.; Moermans, C.; Louis, E.; Louis, R.; Malaise, M.; Guiot, J. Exosomal Long Non-Coding RNAs in Lung Diseases. Int. J. Mol. Sci. 2020, 21, 3580. [Google Scholar] [CrossRef]
- Soni, D.K.; Biswas, R. Role of Non-Coding RNAs in Post-Transcriptional Regulation of Lung Diseases. Front. Genet. 2021, 12, 767348. [Google Scholar] [CrossRef]
- Booton, R.; Lindsay, M.A. Emerging role of MicroRNAs and long noncoding RNAs in respiratory disease. Chest 2014, 146, 193–204. [Google Scholar] [CrossRef]
- Sweef, O.; Zaabout, E.; Bakheet, A.; Halawa, M.; Gad, I.; Akela, M.; Tousson, E.; Abdelghany, A.; Furuta, S. Unraveling Therapeutic Opportunities and the Diagnostic Potential of microRNAs for Human Lung Cancer. Pharmaceutics 2023, 15, 2061. [Google Scholar] [CrossRef] [PubMed] [PubMed Central]
- Fang, D.; Ou, X.; Sun, K.; Zhou, X.; Li, Y.; Shi, P.; Zhao, Z.; He, Y.; Peng, J.; Xu, J. m6A modification-mediated lncRNA TP53TG1 inhibits gastric cancer progression by regulating CIP2A stability. Cancer Sci. 2022, 113, 4135–4150. [Google Scholar] [CrossRef] [PubMed]
- Herman, A.B.; Tsitsipatis, D.; Gorospe, M. Integrated lncRNA function upon genomic and epigenomic regulation. Mol. Cell 2022, 82, 2252–2266. [Google Scholar] [CrossRef] [PubMed]
- Wu, Y.; Sarkissyan, M.; Ogah, O.; Kim, J.; Vadgama, J.V. Expression of MALAT1 Promotes Trastuzumab Resistance in HER2 Overexpressing Breast Cancers. Cancers 2020, 12, 1918. [Google Scholar] [CrossRef]
- Kim, J.; Piao, H.L.; Kim, B.J.; Yao, F.; Han, Z.; Wang, Y.; Xiao, Z.; Siverly, A.N.; Lawhon, S.E.; Ton, B.N.; et al. Long noncoding RNA MALAT1 suppresses breast cancer metastasis. Nat. Genet. 2018, 50, 1705–1715. [Google Scholar] [CrossRef]
- Gupta, R.A.; Shah, N.; Wang, K.C.; Kim, J.; Horlings, H.M.; Wong, D.J.; Tsai, M.C.; Hung, T.; Argani, P.; Rinn, J.L.; et al. Long Non-Coding RNA HOTAIR Reprograms Chromatin State to Promote Cancer Metastasis. Nature 2010, 464, 1071–1076. [Google Scholar] [CrossRef]
- Fang, S.; Shen, Y.; Chen, B.; Wu, Y.; Jia, L.; Li, Y.; Zhu, Y.; Yan, Y.; Li, M.; Chen, R.; et al. H3K27me3 Induces Multidrug Resistance in Small Cell Lung Cancer by Affecting HOXA1 DNA Methylation via Regulation of the lncRNA HOTAIR. Ann. Transl. Med. 2018, 6, 440. [Google Scholar] [CrossRef]
- Ali, M.A.; Shaker, O.G.; Khalifa, A.A.; Ezzat, E.M.; Elghobary, H.A.; Abdel Mawla, T.S.; Elkhateeb, A.F.; Elebiary, A.M.A.; Elamir, A.M. LncRNAs NEAT1, HOTAIR, and GAS5 Expression in Hypertensive and Non-Hypertensive Associated Cerebrovascular Stroke Patients, and Its Link to Clinical Characteristics and Severity Score of the Disease. Noncoding RNA Res. 2022, 8, 96–108. [Google Scholar] [CrossRef]
- Wang, Y.; Ren, X.; Yuan, Y.; Yuan, B.S. Downregulated lncRNA GAS5 and Upregulated miR-21 Lead to Epithelial-Mesenchymal Transition and Lung Metastasis of Osteosarcomas. Front. Cell Dev. Biol. 2021, 9, 707693. [Google Scholar] [CrossRef] [PubMed]
- Hao, X.; Wei, H. LncRNA H19 alleviates sepsis-induced acute lung injury by regulating the miR-107/TGFBR3 axis. BMC Pulm. Med. 2022, 22, 371. [Google Scholar] [CrossRef] [PubMed]
- Zhang, R.; Pan, T.; Xiang, Y.; Zhang, M.; Xie, H.; Liang, Z.; Chen, B.; Xu, C.; Wang, J.; Huang, X.; et al. Curcumenol triggered ferroptosis in lung cancer cells via lncRNA H19/miR-19b-3p/FTH1 axis. Bioact. Mater. 2021, 13, 23–36. [Google Scholar] [CrossRef]
- Rojas-Cruz, A.F.; Bermúdez-Santana, C.I. Computational Prediction of RNA-RNA Interactions between Small RNA Tracks from Betacoronavirus Nonstructural Protein 3 and Neurotrophin Genes during Infection of an Epithelial Lung Cancer Cell Line: Potential Role of Novel Small Regulatory RNA. Viruses 2023, 15, 1647. [Google Scholar] [CrossRef] [PubMed]
- Wang, H.; Shen, Q.; Zhang, X.; Yang, C.; Cui, S.; Sun, Y.; Wang, L.; Fan, X.; Xu, S. The Long Non-Coding RNA XIST Controls Non-Small Cell Lung Cancer Proliferation and Invasion by Modulating miR-186-5p. Cell Physiol. Biochem. 2017, 41, 2221–2229. [Google Scholar] [CrossRef] [PubMed]
- Archer, K.; Broskova, Z.; Bayoumi, A.S.; Teoh, J.P.; Davila, A.; Tang, Y.; Su, H.; Kim, I.M. Long Non-Coding RNAs as Master Regulators in Cardiovascular Diseases. Int. J. Mol. Sci. 2015, 16, 23651–23667. [Google Scholar] [CrossRef]
- Sebastian-delaCruz, M.; Gonzalez-Moro, I.; Olazagoitia-Garmendia, A.; Castellanos-Rubio, A.; Santin, I. The Role of lncRNAs in Gene Expression Regulation through mRNA Stabilization. Noncoding RNA 2021, 7, 3. [Google Scholar] [CrossRef] [PubMed] [PubMed Central]
- Lin, J.; Jiang, Z.; Liu, C.; Zhou, D.; Song, J.; Liao, Y.; Chen, J. Emerging Roles of Long Non-Coding RNAs in Renal Fibrosis. Life 2020, 10, 131. [Google Scholar] [CrossRef] [PubMed] [PubMed Central]
- Ma, T.; Tang, Y.; Wang, T.; Yang, Y.; Zhang, Y.; Wang, R.; Zhang, Y.; Li, Y.; Wu, M.; Tang, M.; et al. Chronic pulmonary bacterial infection facilitates breast cancer lung metastasis by recruiting tumor-promoting MHCIIhi neutrophils. Signal Transduct. Target. Ther. 2023, 8, 296. [Google Scholar] [CrossRef]
- Warren, G.W.; Cummings, K.M. Tobacco and lung cancer: Risks, trends, and outcomes in patients with cancer. Am. Soc. Clin. Oncol. Educ. Book 2013, 33, 359–364. [Google Scholar] [CrossRef]
- Wang, F.; Lin, H.; Su, Q.; Li, C. Cuproptosis-related lncRNA predict prognosis and immune response of lung adenocarcinoma. World J. Surg. Oncol. 2022, 20, 275. [Google Scholar] [CrossRef]
- Desind, S.Z.; Iacona, J.R.; Yu, C.Y.; Mitrofanova, A.; Lutz, C.S. PACER lncRNA regulates COX-2 expression in lung cancer cells. Oncotarget 2022, 13, 291–306. [Google Scholar] [CrossRef]
- He, Y.; Jiang, X.; Duan, L.; Xiong, Q.; Yuan, Y.; Liu, P.; Jiang, L.; Shen, Q.; Zhao, S.; Yang, C.; et al. LncRNA PKMYT1AR promotes cancer stem cell maintenance in non-small cell lung cancer via activating Wnt signaling pathway. Mol. Cancer 2021, 20, 156. [Google Scholar] [CrossRef]
- Liu, S.J.; Dang, H.X.; Lim, D.A.; Feng, F.Y.; Maher, C.A. Long noncoding RNAs in cancer metastasis. Nat. Rev. Cancer 2021, 21, 446–460. [Google Scholar] [CrossRef]
- Sun, L.; Xu, A.; Li, M.; Xia, X.; Li, P.; Han, R.; Fei, G.; Zhou, S.; Wang, R. Effect of Methylation Status of lncRNA-MALAT1 and MicroRNA-146a on Pulmonary Function and Expression Level of COX2 in Patients with Chronic Obstructive Pulmonary Disease. Front. Cell Dev. Biol. 2021, 9, 667624. [Google Scholar] [CrossRef]
- Sundar, I.K.; Yao, H.; Rahman, I. Oxidative stress and chromatin remodeling in chronic obstructive pulmonary disease and smoking-related diseases. Antioxid. Redox Signal. 2013, 18, 1956–1971. [Google Scholar] [CrossRef]
- Zhou, D.; Wang, J.; Liu, X. Development of six immune-related lncRNA signature prognostic model for smoking-positive lung adenocarcinoma. J. Clin. Lab. Anal. 2022, 36, e24467. [Google Scholar] [CrossRef]
- Zheng, R.; Gao, F.; Mao, Z.; Xiao, Y.; Yuan, L.; Huang, Z.; Lv, Q.; Qin, C.; Du, M.; Zhang, Z.; et al. LncRNA BCCE4 Genetically Enhances the PD-L1/PD-1 Interaction in Smoking-Related Bladder Cancer by Modulating miR-328-3p-USP18 Signaling. Adv. Sci. 2023, 10, e2303473. [Google Scholar] [CrossRef]
- Wang, S.; Jiang, W.; Zhang, X.; Lu, Z.; Geng, Q.; Wang, W.; Li, N.; Cai, X. LINC-PINT Alleviates Lung Cancer Progression via Sponging miR-543 and Inducing PTEN. Cancer Med. 2020, 9, 1999–2009. [Google Scholar] [CrossRef]
- Maremanda, K.P.; Sundar, I.K.; Rahman, I. Role of inner mitochondrial protein OPA1 in mitochondrial dysfunction by tobacco smoking and in the pathogenesis of COPD. Redox Biol. 2021, 45, 102055. [Google Scholar] [CrossRef]
- You, Z.; Ye, X.; Jiang, M.; Gu, N.; Liang, C. lnc-MRGPRF-6:1 Promotes ox-LDL-Induced Macrophage Ferroptosis via Suppressing GPX4. Mediators Inflamm. 2023, 2023, 1–14. [Google Scholar] [CrossRef]
- Upadhyay, P.; Wu, C.W.; Pham, A.; Zeki, A.A.; Royer, C.M.; Kodavanti, U.P.; Takeuchi, M.; Bayram, H.; Pinkerton, K.E. Animal models and mechanisms of tobacco smoke-induced chronic obstructive pulmonary disease (COPD). J. Toxicol. Environ. Health B Crit. Rev. 2023, 26, 275–305. [Google Scholar] [CrossRef]
- Wang, L.; Yu, Q.; Xiao, J.; Chen, Q.; Fang, M.; Zhao, H. Cigarette Smoke Extract-Treated Mouse Airway Epithelial Cells-Derived Exosomal LncRNA MEG3 Promotes M1 Macrophage Polarization and Pyroptosis in Chronic Obstructive Pulmonary Disease by Upregulating TREM-1 via m6A Methylation. Immune Netw. 2024, 24, e3. [Google Scholar] [CrossRef]
- Du, Y.; Ding, Y.; Shi, T.; He, W.; Feng, J.; Mei, Z.; Chen, X.; Feng, X.; Zhang, X.; Jie, Z. Long noncoding RNA GAS5 attenuates cigarette smoke-induced airway remodeling by regulating miR-217-5p/PTEN axis. Acta Biochim. Biophys. Sin. (Shanghai) 2022, 54, 931–939. [Google Scholar] [CrossRef]
- Guo, P.; Li, R.; Piao, T.H.; Wang, C.L.; Wu, X.L.; Cai, H.Y. Pathological Mechanism and Targeted Drugs of COPD. Int. J. Chron. Obstruct. Pulmon. Dis. 2022, 17, 1565–1575. [Google Scholar] [CrossRef]
- Wang, Y.; Lyu, X.; Wu, X.; Yu, L.; Hu, K. Long non-coding RNA PVT1, a Novel Biomarker for Chronic Obstructive Pulmonary Disease Progression Surveillance and Acute Exacerbation Prediction Potentially Through Interaction with microRNA-146a. J Clin Lab Anal. 2020, 34, e23346. [Google Scholar] [CrossRef]
- Lee, K.Y.; Ho, S.C.; Sun, W.L.; Feng, P.H.; Lin, C.W.; Chen, K.Y.; Chuang, H.C.; Tseng, C.H.; Chen, T.T.; Wu, S.M. Lnc-IL7R Alleviates PM2.5-mediated Cellular Senescence and Apoptosis Through EZH2 Recruitment in Chronic Obstructive Pulmonary Disease. Cell Biol Toxicol. 2022, 38, 1097–1120. [Google Scholar] [CrossRef]
- Szafranski, P.; Gambin, T.; Karolak, J.A.; Popek, E.; Stankiewicz, P. Lung-specific distant enhancer cis regulates expression of FOXF1 and lncRNA FENDRR. Hum Mutat. 2021, 42, 694–698. [Google Scholar] [CrossRef] [PubMed]
- Lewis, A.; Lee, J.Y.; Donaldson, A.V.; Natanek, S.A.; Vaidyanathan, S.; Man, W.D.; Hopkinson, N.S.; Sayer, A.A.; Patel, H.P.; Cooper, C.; et al. Increased expression of H19/miR-675 is associated with a low fat-free mass index in patients with COPD. J. Cachexia Sarcopenia Muscle 2016, 7, 330–344. [Google Scholar] [CrossRef]
- Forder, A.; Zhuang, R.; Souza, V.G.P.; Brockley, L.J.; Pewarchuk, M.E.; Telkar, N.; Stewart, G.L.; Benard, K.; Marshall, E.A.; Reis, P.P.; et al. Mechanisms Contributing to the Comorbidity of COPD and Lung Cancer. Int. J. Mol. Sci. 2023, 24, 2859. [Google Scholar] [CrossRef]
- Mo, R.; Li, J.; Chen, Y.; Ding, Y. lncRNA GAS5 promotes pyroptosis in COPD by functioning as a ceRNA to regulate the miR 223 3p/NLRP3 axis. Mol. Med. Rep. 2022, 26, 219. [Google Scholar] [CrossRef] [PubMed] [PubMed Central]
- Reiche, L.; Göttle, P.; Lane, L.; Duek, P.; Park, M.; Azim, K.; Schütte, J.; Manousi, A.; Schira-Heinen, J.; Küry, P. C21orf91 Regulates Oligodendroglial Precursor Cell Fate—A Switch in the Glial Lineage? Front. Cell. Neurosci. 2021, 15. [Google Scholar] [CrossRef] [PubMed]
- Wang, S.; Wang, X.; Xu, S.L. LINC00473 Promotes Lung Adenocarcinoma Progression by Regulating miR-1294/ROBO1 Axis. J. Biol. Regul. Homeost. Agents Epub ahead of print. 2020, 34. [Google Scholar] [PubMed]
- Xia, H.; Xue, J.; Xu, H.; Lin, M.; Shi, M.; Sun, Q.; Xiao, T.; Dai, X.; Wu, L.; Li, J.; et al. Andrographolide Antagonizes the Cigarette Smoke-Induced Epithelial-Mesenchymal Transition and Pulmonary Dysfunction Through Anti-inflammatory Inhibiting HOTAIR. Toxicology. 2019, 422, 84–94. [Google Scholar] [CrossRef] [PubMed]
- Ge, J.; Geng, S.; Jiang, H. Long noncoding RNAs antisense noncoding RNA in the INK4 locus (ANRIL) correlates with lower acute exacerbation risk, decreased inflammatory cytokines, and mild GOLD stage in patients with chronic obstructive pulmonary disease. J. Clin. Lab. Anal. 2019, 33, e22678. [Google Scholar] [CrossRef]
- Araya, J.; Tsubouchi, K.; Sato, N.; Ito, S.; Minagawa, S.; Hara, H.; Hosaka, Y.; Ichikawa, A.; Saito, N.; Kadota, T.; et al. PRKN-regulated mitophagy and cellular senescence during COPD pathogenesis. Autophagy 2019, 15, 510–526. [Google Scholar] [CrossRef]
- Dong, Z.; Yang, P.; Qiu, X.; Liang, S.; Guan, B.; Yang, H.; Li, F.; Sun, L.; Liu, H.; Zou, G.; et al. KCNQ1OT1 facilitates progression of non-small-cell lung carcinoma via modulating miRNA-27b-3p/HSP90AA1 axis. J Cell Physiol. 2019, 234, 11304–11314. [Google Scholar] [CrossRef]
- Zheng, Z.H.; Wu, D.M.; Fan, S.H.; Zhang, Z.F.; Chen, G.Q.; Lu, J. Upregulation of miR-675-5p induced by lncRNA H19 was associated with tumor progression and development by targeting tumor suppressor p53 in non-small cell lung cancer. J Cell Biochem. 2019, 120, 18724–18735. [Google Scholar] [CrossRef]
- Tao, Y.; Tang, Y.; Yang, Z.; Wu, F.; Wang, L.; Yang, L.; Lei, L.; Jing, Y.; Jiang, X.; Jin, H.; et al. Exploration of Serum Exosomal LncRNA TBILA and AGAP2-AS1 as Promising Biomarkers for Diagnosis of Non-Small Cell Lung Cancer. Int. J. Biol. Sci. 2020, 16, 471–482. [Google Scholar] [CrossRef] [PubMed]
- Liu, S.; Liu, M.; Dong, L. The clinical value of lncRNA MALAT1 and its targets miR-125b, miR-133, miR-146a, and miR-203 for predicting disease progression in chronic obstructive pulmonary disease patients. J Clin Lab Anal. 2020, 34, e23410. [Google Scholar] [CrossRef] [PubMed]
- Xu, M.; Senanayaka, D.; Zhao, R.; Chigumira, T.; Tripathi, A.; Tones, J.; Lackner, R.M.; Wondisford, A.R.; Moneysmith, L.N.; Hirschi, A.; et al. TERRA-LSD1 phase separation promotes R-loop formation for telomere maintenance in ALT cancer cells. Nat Commun. 2024, 15, 2165. [Google Scholar] [CrossRef] [PubMed]
- Wang, M.; Zhou, L.; Yu, F.; Zhang, Y.; Li, P.; Wang, K. The functional roles of exosomal long non-coding RNAs in cancer. Cell. Mol. Life Sci. 2019, 76, 2059–2076. [Google Scholar] [CrossRef] [PubMed]
- Hu, T.J.; Huang, H.B.; Shen, H.B.; Chen, W.; Yang, Z.H. Role of long non-coding RNA MALAT1 in chronic obstructive pulmonary disease. Exp. Ther. Med. 2020, 20, 2691–2697. [Google Scholar] [CrossRef] [PubMed]
- Devadoss, D.; Long, C.; Langley, R.J.; Manevski, M.; Nair, M.; Campos, M.A.; Borchert, G.; Rahman, I.; Chand, H.S. Long Noncoding Transcriptome in Chronic Obstructive Pulmonary Disease. Am. J. Respir. Cell Mol. Biol. 2019, 61, 678–688. [Google Scholar] [CrossRef] [PubMed] [PubMed Central]
- Kumar, P.; Sen, C.; Peters, K.; Frizzell, R.A.; Biswas, R. Comparative analyses of long non-coding RNA profiles in vivo in cystic fibrosis lung airway and parenchyma tissues. Respir. Res. 2019, 20, 284. [Google Scholar] [CrossRef]
- Dai, J.; Zhou, N.; Wu, R.; Du, J.; Miao, S.; Gong, K.; Yang, L.; Chen, W.; Li, X.; Li, C.; et al. LncRNA MALAT1 Regulating Lung Carcinoma Progression via the miR-491-5p/UBE2C Axis. Pathol. Oncol. Res. 2021, 27, 610159. [Google Scholar] [CrossRef] [PubMed]
- Liu, G.; Philp, A.M.; Corte, T.; Travis, M.A.; Schilter, H.; Hansbro, N.G.; Burns, C.J.; Eapen, M.S.; Sohal, S.S.; Burgess, J.K.; et al. Therapeutic targets in lung tissue remodelling and fibrosis. Pharmacol. Ther. 2021, 225, 107839. [Google Scholar] [CrossRef]
- Di Stefano, A.; Sangiorgi, C.; Gnemmi, I.; Casolari, P.; Brun, P.; Ricciardolo, F.L.M.; Contoli, M.; Papi, A.; Maniscalco, P.; Ruggeri, P.; et al. TGF-β Signaling Pathways in Different Compartments of the Lower Airways of Patients with Stable COPD. Chest 2018, 153, 851–862. [Google Scholar] [CrossRef]
- Mahmood, M.Q.; Reid, D.; Ward, C.; Muller, H.K.; Knight, D.A.; Sohal, S.S.; Walters, E.H. Transforming growth factor (TGF) β1 and Smad signalling pathways: A likely key to EMT-associated COPD pathogenesis. Respirology 2016, 22, 133–140. [Google Scholar] [CrossRef] [PubMed]
- Jarroux, J.; Morillon, A.; Pinskaya, M. History, Discovery, and Classification of lncRNAs. Adv. Exp. Med. Biol. 2017, 1008, 1–46. [Google Scholar] [CrossRef]
- Zhu, J.; Fu, H.; Wu, Y.; Zheng, X. Function of lncRNAs and approaches to lncRNA-protein interactions. Sci. China Life Sci. 2013, 56, 876–885. [Google Scholar] [CrossRef] [PubMed]
- Bridges, M.C.; Daulagala, A.C.; Kourtidis, A. LNCcation: LncRNA localization and function. J. Cell Biol. 2021, 220, e202009045. [Google Scholar] [CrossRef]
- Zhang, H.; Sun, D.; Li, D.; Zheng, Z.; Xu, J.; Liang, X.; Zhang, C.; Wang, S.; Wang, J.; Lu, W. Long non-coding RNA expression patterns in lung tissues of chronic cigarette smoke induced COPD mouse model. Sci. Rep. 2018, 8, 7609. [Google Scholar] [CrossRef] [PubMed]
- Negewo, N.A.; Gibson, P.G.; McDonald, V.M. COPD and its comorbidities: Impact, measurement and mechanisms. Respirology 2015, 20, 1160–1171. [Google Scholar] [CrossRef]
- Uwagboe, I.; Adcock, I.M.; Lo Bello, F.; Caramori, G.; Mumby, S. New drugs under development for COPD. Minerva Med. 2022, 113, 471–496. [Google Scholar] [CrossRef]
- Raherison, C.; Girodet, P.O. Epidemiology of COPD. Eur. Respir. Rev. 2009, 18, 213–221. [Google Scholar] [CrossRef]
- Adams, T.S.; Schupp, J.C.; Poli, S.; Ayaub, E.A.; Neumark, N.; Ahangari, F.; Chu, S.G.; Raby, B.A.; DeIuliis, G.; Januszyk, M.; et al. Single-Cell RNA-Seq Reveals Ectopic and Aberrant Lung-Resident Cell Populations in Idiopathic Pulmonary Fibrosis. Sci. Adv. 2020, 6, eaba1983. [Google Scholar] [CrossRef] [PubMed]
- Lin, Q.; Zhang, C.F.; Guo, J.L.; Su, J.L.; Guo, Z.K.; Li, H.Y. Involvement of NEAT1/PINK1-Mediated Mitophagy in Chronic Obstructive Pulmonary Disease Induced by Cigarette Smoke or PM2.5. Ann. Transl. Med. 2022, 10, 277. [Google Scholar] [CrossRef]
- Cao, Y.; Zhang, H.; Tang, J.; Wang, R. Long non-coding RNA FAM230B is a novel prognostic and diagnostic biomarker for lung adenocarcinoma. Bioengineered 2022, 13, 7919–7925. [Google Scholar] [CrossRef] [PubMed]
- Zhang, C.; Huang, Q.; He, F. Correlation of small nucleolar RNA host gene 16 with acute respiratory distress syndrome occurrence and prognosis in sepsis patients. J. Clin. Lab. Anal. 2022, 36, e24516. [Google Scholar] [CrossRef] [PubMed]
- Liu, W.; Liang, W.; Zhang, C.; Liu, H.; Li, H.; Zhou, L.; Zhou, L. LncR-GAS5 decrease in adenine phosphoribosyltransferase expresssion via binding TAF1 to increase kidney damage created by CIH. Heliyon 2024, 10, e33084. [Google Scholar] [CrossRef] [PubMed]
- Hussain, S.; Afzal, O.; Gupta, G.; Goyal, A.; Almalki, W.H.; Kazmi, I.; Alzarea, S.I.; Altamimi, A.S.A.; Kukreti, N.; Chakraborty, A.; et al. Unraveling Neat1’s Complex Role In Lung Cancer Biology: A Comprehensive Review. EXCLI J. 2024, 23, 34–52. [Google Scholar] [CrossRef] [PubMed]
- Jen, J.; Tang, Y.A.; Lu, Y.H.; Lin, C.C.; Lai, W.W.; Wang, Y.C. Oct4 transcriptionally regulates the expression of long non-coding RNAs NEAT1 and MALAT1 to promote lung cancer progression. Mol. Cancer 2017, 16, 104. [Google Scholar] [CrossRef] [PubMed]
- Song, Y.; Kelava, L.; Zhang, L.; Kiss, I. Microarray data analysis to identify miRNA biomarkers and construct the lncRNA-miRNA-mRNA network in lung adenocarcinoma. Medicine 2022, 101, e30393. [Google Scholar] [CrossRef] [PubMed]
- Tokgun, O.; Tokgun, P.E.; Inci, K.; Akca, H. lncRNAs as Potential Targets in Small Cell Lung Cancer: MYC-dependent Regulation. Anticancer Agents Med. Chem. 2020, 20, 2074–2081. [Google Scholar] [CrossRef] [PubMed]
- Chen, X.F.; Deng, J.M. Emerging Role of Long Noncoding RNAs in Asthma. Comb. Chem. High Throughput Screen. 2023, 26, 247–255. [Google Scholar] [CrossRef] [PubMed]
- Yuan, D.; Liu, Y.; Li, M.; Zhou, H.; Cao, L.; Zhang, X.; Li, Y. Senescence associated long non-coding RNA 1 regulates cigarette smoke-induced senescence of type II alveolar epithelial cells through sirtuin-1 signaling. J. Int. Med. Res. 2021, 49, 300060520986049. [Google Scholar] [CrossRef]
- Ezegbunam, W.; Foronjy, R. Posttranscriptional control of airway inflammation. Wiley Interdiscip. Rev. RNA 2018, 9, e1455. [Google Scholar] [CrossRef]
- Wang, W.; Min, L.; Qiu, X.; Wu, X.; Liu, C.; Ma, J.; Zhang, D.; Zhu, L. Biological Function of Long Non-coding RNA (lncRNA) Xist. Front. Cell Dev. Biol. 2021, 9, 645647. [Google Scholar] [CrossRef]
- Huang, M.; Wang, H.; Hu, X.; Cao, X. lncRNA MALAT1 binds chromatin remodeling subunit BRG1 to epigenetically promote inflammation-related hepatocellular carcinoma progression. Oncoimmunology 2018, 8, e1518628. [Google Scholar] [CrossRef]
- Dai, J.; Qu, T.; Yin, D.; Cui, Y.; Zhang, C.; Zhang, E.; Guo, R. LncRNA LINC00969 promotes acquired gefitinib resistance by epigenetically suppressing of NLRP3 at transcriptional and posttranscriptional levels to inhibit pyroptosis in lung cancer. Cell Death Dis. 2023, 14, 312. [Google Scholar] [CrossRef]
- Xing, C.; Sun, S.G.; Yue, Z.Q.; Bai, F. Role of lncRNA LUCAT1 in cancer. Biomed. Pharmacother. 2021, 134, 111158. [Google Scholar] [CrossRef] [PubMed]
- Yin, H.; Chen, L.; Piao, S.; Wang, Y.; Li, Z.; Lin, Y.; Tang, X.; Zhang, H.; Zhang, H.; Wang, X. M6A RNA methylation-mediated RMRP stability renders proliferation and progression of non-small cell lung cancer through regulating TGFBR1/SMAD2/SMAD3 pathway. Cell Death Differ. 2023, 30, 605–617. [Google Scholar] [CrossRef]
- Zhang, H.; Wang, S.Q.; Zhu, J.B.; Wang, L.N.; Lin, H.; Li, L.F.; Cheng, Y.D.; Duan, C.J.; Zhang, C.F. LncRNA CALML3-AS1 modulated by m6A modification induces BTNL9 methylation to drive non-small-cell lung cancer progression. Cancer Gene Ther. 2023, 30, 1649–1662. [Google Scholar] [CrossRef] [PubMed]
- Barreiro, E.; Sznajder, J.I. Epigenetic regulation of muscle phenotype and adaptation: A potential role in COPD muscle dysfunction. J. Appl. Physiol. (1985) 2013, 114, 1263–1272. [Google Scholar] [CrossRef] [PubMed]
- Wu, S.; Huang, K.; Chang, C.; Chu, X.; Zhang, K.; Li, B.; Yang, T. Serum Proteomic Profiling in Patients with Chronic Obstructive Pulmonary Disease. Int. J. Chron. Obstruct. Pulmon. Dis. 2023, 18, 1623–1635. [Google Scholar] [CrossRef] [PubMed]
- Salvato, I.; Ricciardi, L.; Nucera, F.; Nigro, A.; Dal Col, J.; Monaco, F.; Caramori, G.; Stellato, C. RNA-Binding Proteins as a Molecular Link between COPD and Lung Cancer. COPD J. Chronic Obstr. Pulm. Dis. 2023, 20, 18–30. [Google Scholar] [CrossRef]
- Yang, M.; Kohler, M.; Heyder, T.; Forsslund, H.; Garberg, H.K.; Karimi, R.; Grunewald, J.; Berven, F.S.; Nyrén, S.; Magnus Sköld, C.; et al. Proteomic Profiling of Lung Immune Cells Reveals Dysregulation of Phagocytotic Pathways in Female-Dominated Molecular COPD Phenotype. Respir. Res. 2018, 19, 39. [Google Scholar] [CrossRef] [PubMed] [PubMed Central]
- Kim, R.Y.; Oliver, B. Innate Immune Reprogramming in Chronic Obstructive Pulmonary Disease: New Mechanisms for Old Questions. Am. J. Respir. Cell Mol. Biol. 2023, 68, 470–471. [Google Scholar] [CrossRef]
- Mayama, T.; Marr, A.K.; Kino, T. Differential Expression of Glucocorticoid Receptor Noncoding RNA Repressor Gas5 in Autoimmune and Inflammatory Diseases. Horm. Metab. Res. 2016, 48, 550–557. [Google Scholar] [CrossRef]
- Li, F.; Sun, J.; Huang, S.; Su, G.; Pi, G. LncRNA GAS5 Overexpression Reverses LPS-Induced Inflammatory Injury and Apoptosis Through Up-Regulating KLF2 Expression in ATDC5 Chondrocytes. Cell. Physiol. Biochem. 2018, 45, 1241–1251. [Google Scholar] [CrossRef] [PubMed]
- Chen, L.; Huang, S.; Huang, J.; Chen, Q.; Zhuang, Q. Role and Mechanism of Exosome-Derived Long Noncoding RNA HOTAIR in Lung Cancer. ACS Omega 2021, 6, 17217–17227. [Google Scholar] [CrossRef]
- Tang, Q.; Zhou, Q.; Li, J.; Yang, X.; Wang, R.; Wang, X.; Xu, M.; Han, L.; Wu, W.; Wang, S. Solamargine enhanced gefitinib antitumor effect via regulating MALAT1/miR-141-3p/Sp1/IGFBP1 signaling pathway in non-small cell lung cancer. Carcinogenesis 2023, 44, 497–510. [Google Scholar] [CrossRef]
- Yang, T.; Li, H.; Chen, T.; Ren, H.; Shi, P.; Chen, M. LncRNA MALAT1 Depressed Chemo-Sensitivity of NSCLC Cells through Directly Functioning on miR-197-3p/p120 Catenin Axis. Mol Cells 2019, 42, 270–283. [Google Scholar] [CrossRef] [PubMed]
- Park, E.G.; Pyo, S.J.; Cui, Y.; Yoon, S.H.; Nam, J.W. Tumor immune microenvironment lncRNAs. Brief Bioinform. 2022, 23, bbab504. [Google Scholar] [CrossRef]
- Lv, X.; Zhang, X.Y.; Zhang, Q.; Nie, Y.J.; Luo, G.H.; Fan, X.; Yang, S.; Zhao, Q.H.; Li, J.Q. lncRNA NEAT1 aggravates sepsis-induced lung injury by regulating the miR-27a/PTEN axis. Lab Investig. 2021, 101, 1371–1381. [Google Scholar] [CrossRef] [PubMed]
- Luo, Z.; Yi, Z.J.; Ou, Z.L.; Han, T.; Wan, T.; Tang, Y.C.; Wang, Z.C.; Huang, F.Z. RELA/NEAT1/miR-302a-3p/RELA feedback loop modulates pancreatic ductal adenocarcinoma cell proliferation and migration. J. Cell Physiol. 2019, 234, 3583–3597. [Google Scholar] [CrossRef] [PubMed]
- Ma, J.; Miao, H.; Zhang, H.; Ren, J.; Qu, S.; Da, J.; Xu, F.; Zhao, H. LncRNA GAS5 modulates the progression of non-small cell lung cancer through repressing miR-221-3p and up-regulating IRF2. Diagn Pathol. 2021, 16, 46. [Google Scholar] [CrossRef]
- Zhu, L.; Zhou, D.; Guo, T.; Chen, W.; Ding, Y.; Li, W.; Huang, Y.; Huang, J.; Pan, X. LncRNA GAS5 inhibits Invasion and Migration of Lung Cancer through influencing EMT process. J. Cancer 2021, 12, 3291–3298. [Google Scholar] [CrossRef] [PubMed]
- Dai, Z.; Liu, X.; Zeng, H.; Chen, Y. Long noncoding RNA HOTAIR facilitates pulmonary vascular endothelial cell apoptosis via DNMT1 mediated hypermethylation of Bcl-2 promoter in COPD. Respir. Res. 2022, 23, 356. [Google Scholar] [CrossRef] [PubMed] [PubMed Central]
- Liu, Y.; Luo, F.; Xu, Y.; Wang, B.; Zhao, Y.; Xu, W.; Shi, L.; Lu, X.; Liu, Q. Corrigendum to “Epithelial-mesenchymal transition and cancer stem cells, mediated by a long non-coding RNA, HOTAIR, are involved in cell malignant transformation induced by cigarette smoke extract” [Toxicology and Applied Pharmacology, 282(2015), page 9–19/Article Number: TAAP-D-14-00789]. Toxicol. Appl. Pharmacol. 2023, 467, 116508. [Google Scholar] [CrossRef] [PubMed]
- Liu, J.; Ali, M.K.; Mao, Y. Emerging role of long non-coding RNA MALAT1 related signaling pathways in the pathogenesis of lung disease. Front. Cell Dev. Biol. 2023, 11, 1149499. [Google Scholar] [CrossRef] [PubMed] [PubMed Central]
- Zhang, J.; Wang, M.; Wang, J.; Wang, W. JMJD2C-mediated long non-coding RNA MALAT1/microRNA-503-5p/SEPT2 axis worsens non-small cell lung cancer. Cell Death Dis. 2022, 13, 65. [Google Scholar] [CrossRef]
- Tang, W.; Shen, Z.; Guo, J.; Sun, S. Screening of long non-coding RNA and TUG1 inhibits proliferation with TGF-β induction in patients with COPD. Int. J. Chron. Obstruct. Pulmon. Dis. 2016, 11, 2951–2964. [Google Scholar] [CrossRef] [PubMed] [PubMed Central]
- Gu, W.; Yuan, Y.; Wang, L.; Yang, H.; Li, S.; Tang, Z.; Li, Q. Long non-coding RNA TUG1 promotes airway remodelling by suppressing the miR-145-5p/DUSP6 axis in cigarette smoke-induced COPD. J. Cell Mol. Med. 2019, 23, 7200–7209. [Google Scholar] [CrossRef] [PubMed]
- Zeng, Z.; Lan, Y.; Chen, Y.; Zuo, F.; Gong, Y.; Luo, G.; Peng, Y.; Yuan, Z. LncRNA GAS5 suppresses inflammatory responses by inhibiting HMGB1 release via miR-155-5p/SIRT1 axis in sepsis. Eur. J. Pharmacol. 2023, 942, 175520. [Google Scholar] [CrossRef] [PubMed]
- Li, B.; Zhang, J.; Dong, H.; Feng, X.; Yu, L.; Zhu, J.; Zhang, J. Systematic analysis of various RNA transcripts and construction of biological regulatory networks at the post-transcriptional level for chronic obstructive pulmonary disease. J. Transl. Med. 2023, 21, 790. [Google Scholar] [CrossRef] [PubMed] [PubMed Central]
- Li, J.; Liu, S. LncRNA GAS5 suppresses inflammatory responses and apoptosis of alveolar epithelial cells by targeting miR-429/DUSP1. Exp. Mol. Pathol. 2020, 113, 104357. [Google Scholar] [CrossRef]
- Ming, X.; Duan, W.; Yi, W. Long non-coding RNA NEAT1 predicts elevated chronic obstructive pulmonary disease (COPD) susceptibility and acute exacerbation risk, and correlates with higher disease severity, inflammation, and lower miR-193a in COPD patients. Int. J. Clin. Exp. Pathol. 2019, 12, 2837–2848. Available online: https://www.ncbi.nlm.nih.gov/pmc/articles/PMC6949709/ (accessed on 17 April 2024). [PubMed] [PubMed Central]
- Wang, Y.; Xu, J.; Meng, Y.; Adcock, I.M.; Yao, X. Role of inflammatory cells in airway remodeling in COPD. Int. J. Chron. Obstruct. Pulmon. Dis. 2018, 13, 3341–3348. [Google Scholar] [CrossRef] [PubMed] [PubMed Central]
- Tu, Y.H.; Guo, Y.; Ji, S.; Shen, J.L.; Fei, G.H. The Influenza A Virus H3N2 Triggers the Hypersusceptibility of Airway Inflammatory Response via Activating the lncRNA TUG1/miR-145-5p/NF-κB Pathway in COPD. Front. Pharmacol. 2021, 12, 604590. [Google Scholar] [CrossRef] [PubMed] [PubMed Central]
- Wang, J.; Xia, B.; Ma, R.; Ye, Q. Comprehensive Analysis of a Competing Endogenous RNA Co-Expression Network in Chronic Obstructive Pulmonary Disease. Int. J. Chron. Obstruct. Pulmon. Dis. 2023, 18, 2417–2429. [Google Scholar] [CrossRef] [PubMed]
- Zhang, J.; Piao, C.D.; Ding, J.; Li, Z.W. LncRNA MALAT1 facilitates lung metastasis of osteosarcomas through miR-202 sponging. Sci Rep. 2020, 10, 12757. [Google Scholar] [CrossRef] [PubMed]
- Feng, L.; Guo, J.; Ai, F. Circulating long noncoding RNA ANRIL downregulation correlates with increased risk, higher disease severity and elevated pro-inflammatory cytokines in patients with acute ischemic stroke. J Clin Lab Anal. 2019, 33, e22629. [Google Scholar] [CrossRef]
- Wei, L.; Li, J.; Han, Z.; Chen, Z.; Zhang, Q. Silencing of lncRNA MALAT1 Prevents Inflammatory Injury after Lung Transplant Ischemia-Reperfusion by Downregulation of IL-8 via p300. Mol. Ther. Nucleic Acids 2019, 18, 285–297. [Google Scholar] [CrossRef] [PubMed] [PubMed Central]
- Bi, H.; Wang, G.; Li, Z.; Zhou, L.; Zhang, M.; Ye, J.; Wang, Z. Long Noncoding RNA (lncRNA) Maternally Expressed Gene 3 (MEG3) Participates in Chronic Obstructive Pulmonary Disease through Regulating Human Pulmonary Microvascular Endothelial Cell Apoptosis. Med. Sci. Monit. 2020, 26, e920793, Erratum in: Med Sci Monit. 2020, 26, e927410. [Google Scholar] [CrossRef] [PubMed] [PubMed Central]
- Fu, Y.S.; Kang, N.; Yu, Y.; Mi, Y.; Guo, J.; Wu, J.; Weng, C.F. Polyphenols, flavonoids and inflammasomes: The role of cigarette smoke in COPD. Eur. Respir. Rev. 2022, 31, 220028. [Google Scholar] [CrossRef] [PubMed] [PubMed Central]
- Zhang, X.; Liu, J.; Sun, Y.; Zhou, Q.; Ding, X.; Chen, X. Chinese herbal compound Huangqin Qingrechubi capsule reduces lipid metabolism disorder and inflammatory response in gouty arthritis via the LncRNA H19/APN/PI3K/AKT cascade. Pharm. Biol. 2023, 61, 541–555. [Google Scholar] [CrossRef]
- Rao, W.; Wang, S.; Duleba, M.; Niroula, S.; Goller, K.; Xie, J.; Mahalingam, R.; Neupane, R.; Liew, A.A.; Vincent, M.; et al. Regenerative Metaplastic Clones in COPD Lung Drive Inflammation and Fibrosis. Cell 2020, 181, 848–864e18. [Google Scholar] [CrossRef] [PubMed] [PubMed Central]
- Li, H.; Xie, S.; Li, H.; Zhang, R.; Zhang, H. LncRNA MALAT1 mediates proliferation of LPS treated-articular chondrocytes by targeting the miR-146a-PI3K/Akt/mTOR axis. Life Sci. 2019, 254, 116801. [Google Scholar] [CrossRef]
- Wang, Q.; Xie, Y.; He, Q.; Geng, Y.; Xu, J. LncRNA-Cox2 regulates macrophage polarization and inflammatory response through the CREB-C/EBPβ signaling pathway in septic mice. Int. Immunopharmacol. 2021, 101, 108347. [Google Scholar] [CrossRef]
- Xue, Q.; Wang, Y.; Zheng, Q.; Chen, L.; Jin, Y.; Shen, X.; Li, Y. Construction of a prognostic immune-related lncRNA model and identification of the immune microenvironment in middle- or advanced-stage lung squamous carcinoma patients. Heliyon 2022, 8, e09521. [Google Scholar] [CrossRef] [PubMed]
- Barta, I.; Paska, C.; Antus, B. Sputum Cytokine Profiling in COPD: Comparison Between Stable Disease and Exacerbation. Int. J. Chron. Obstruct. Pulmon. Dis. 2022, 17, 1897–1908. [Google Scholar] [CrossRef]
- Dinescu, S.; Ignat, S.; Lazar, A.D.; Constantin, C.; Neagu, M.; Costache, M. Epitranscriptomic Signatures in lncRNAs and Their Possible Roles in Cancer. Genes 2019, 10, 52. [Google Scholar] [CrossRef] [PubMed] [PubMed Central]
- Königshoff, M.; Kneidinger, N.; Eickelberg, O. TGF-beta signaling in COPD: Deciphering genetic and cellular susceptibilities for future therapeutic regimen. Swiss Med. Wkly. 2009, 139, 554–563. [Google Scholar] [CrossRef] [PubMed]
- Zhang, L.; Gu, H.; Gu, Y.; Zeng, X. Association between TNF-α -308 G/A polymorphism and COPD susceptibility: A meta-analysis update. Int. J. Chron. Obstruct. Pulmon. Dis. 2016, 11, 1367–1379. [Google Scholar] [CrossRef] [PubMed]
- Huang, W.; Luo, T.; Lan, M.; Zhou, W.; Zhang, M.; Wu, L.; Lu, Z.; Fan, L. Identification and Characterization of a ceRNA Regulatory Network Involving LINC00482 and PRRC2B in Peripheral Blood Mononuclear Cells: Implications for COPD Pathogenesis and Diagnosis. Int J Chron Obstruct Pulmon Dis. 2024, 19, 419–430. [Google Scholar] [CrossRef]
- Huang, X.; Zhao, M. High expression of long non-coding RNA MALAT1 correlates with raised acute respiratory distress syndrome risk, disease severity, and increased mortality in sepstic patients. Int. J. Clin. Exp. Pathol. 2019, 12, 1877–1887. Available online: https://www.ncbi.nlm.nih.gov/pmc/articles/PMC6947113/ (accessed on 18 April 2024). [PubMed] [PubMed Central]
- Dong, R.; Xie, L.; Zhao, K.; Zhang, Q.; Zhou, M.; He, P. Cigarette smoke-induced lung inflammation in COPD mediated via LTB4/BLT1/SOCS1 pathway. Int. J. Chron. Obstruct. Pulmon. Dis. 2015, 11, 31–41. [Google Scholar] [CrossRef]
- Li, R.; Fang, L.; Pu, Q.; Bu, H.; Zhu, P.; Chen, Z.; Yu, M.; Li, X.; Weiland, T.; Bansal, A.; et al. MEG3-4 is a miRNA decoy that regulates IL-1β abundance to initiate and then limit inflammation to prevent sepsis during lung infection. Sci. Signal. 2018, 11, eaao2387. [Google Scholar] [CrossRef] [PubMed] [PubMed Central]
- Christopoulou, M.E.; Papakonstantinou, E.; Stolz, D. Matrix Metalloproteinases in Chronic Obstructive Pulmonary Disease. Int. J. Mol. Sci. 2023, 24, 3786. [Google Scholar] [CrossRef] [PubMed]
- Liu, Z.; Li, J.; Chen, T.; Zhao, X.; Chen, Q.; Xiao, L.; Peng, Z.; Zhang, H. Association between dietary antioxidant levels and chronic obstructive pulmonary disease: A mediation analysis of inflammatory factors. Front. Immunol. 2024, 14, 1310399. [Google Scholar] [CrossRef] [PubMed] [PubMed Central]
- Chen, Q.W.; Cai, Q.Q.; Yang, Y.; Dong, S.; Liu, Y.Y.; Chen, Z.Y.; Kang, C.L.; Qi, B.; Dong, Y.W.; Wu, W.; et al. LncRNA BC promotes lung adenocarcinoma progression by modulating IMPAD1 alternative splicing. Clin. Transl. Med. 2023, 13, e1129. [Google Scholar] [CrossRef] [PubMed]
- Lee, H.W.; Jose, C.C.; Cuddapah, S. Epithelial-mesenchymal transition: Insights into nickel-induced lung diseases. Semin. Cancer Biol. 2021, 76, 99–109. [Google Scholar] [CrossRef]
- He, H.; Ji, X.; Cao, L.; Wang, Z.; Wang, X.; Li, X.M.; Miao, M. Medicine Targeting Epithelial-Mesenchymal Transition to Treat Airway Remodeling and Pulmonary Fibrosis Progression. Can. Respir. J. 2023, 2023, 3291957. [Google Scholar] [CrossRef]
- Qian, G.; Liao, Q.; Li, G.; Yin, F. miR-378 associated with proliferation, migration and apoptosis properties in A549 cells and targeted NPNT in COPD. PeerJ 2022, 10, e14062. [Google Scholar] [CrossRef]
- Demedts, I.K.; Demoor, T.; Bracke, K.R.; Joos, G.F.; Brusselle, G.G. Role of apoptosis in the pathogenesis of COPD and pulmonary emphysema. Respir. Res. 2006, 7, 53. [Google Scholar] [CrossRef]
- Lai, H.C.; Lin, T.L.; Chen, T.W.; Kuo, Y.L.; Chang, C.J.; Wu, T.R.; Shu, C.C.; Tsai, Y.H.; Swift, S.; Lu, C.C. Gut microbiota modulates COPD pathogenesis: Role of anti-inflammatory Parabacteroides goldsteinii lipopolysaccharide. Gut 2022, 71, 309–321. [Google Scholar] [CrossRef]
- Wang, S.M.; Liu, G.Q.; Xian, H.B.; Si, J.L.; Qi, S.X.; Yu, Y.P. LncRNA NEAT1 alleviates sepsis-induced myocardial injury by regulating the TLR2/NF-κB signaling pathway. Eur. Rev. Med. Pharmacol. Sci. 2019, 23, 4898–4907. [Google Scholar] [CrossRef]
- Wiegman, C.H.; Li, F.; Ryffel, B.; Togbe, D.; Chung, K.F. Oxidative Stress in Ozone-Induced Chronic Lung Inflammation and Emphysema: A Facet of Chronic Obstructive Pulmonary Disease. Front. Immunol. 2020, 11, 1957. [Google Scholar] [CrossRef]
- Liu, K.; Hua, S.; Song, L. PM2.5 Exposure and Asthma Development: The Key Role of Oxidative Stress. Oxid. Med. Cell. Longev. 2022, 2022, 3618806. [Google Scholar] [CrossRef] [PubMed]
- Salazar, L.M.; Herrera, A.M. Fibrotic response of tissue remodeling in COPD. Lung 2011, 189, 101–109. [Google Scholar] [CrossRef]
- Yang, L.; Wu, D.; Chen, J.; Chen, J.; Qiu, F.; Li, Y.; Liu, L.; Cao, Y.; Yang, B.; Zhou, Y.; et al. A functional CNVR_3425.1 damping lincRNA FENDRR increases lifetime risk of lung cancer and COPD in Chinese. Carcinogenesis 2018, 39, 347–359. [Google Scholar] [CrossRef]
- Kume, H.; Yamada, R.; Sato, Y.; Togawa, R. Airway Smooth Muscle Regulated by Oxidative Stress in COPD. Antioxidants 2023, 12, 142. [Google Scholar] [CrossRef] [PubMed]
- Groot, M.; Zhang, D.; Jin, Y. Long Non-Coding RNA Review and Implications in Lung Diseases. JSM Bioinform. Genom. Proteom. 2018, 3, 1033. [Google Scholar]
- Xiao, J.H.; Hao, Q.Y.; Wang, K.; Paul, J.; Wang, Y.X. Emerging Role of MicroRNAs and Long Noncoding RNAs in Healthy and Diseased Lung. Adv. Exp. Med. Biol. 2017, 967, 343–359. [Google Scholar] [CrossRef]
- de Groot, P.; Munden, R.F. Lung cancer epidemiology, risk factors, and prevention. Radiol. Clin. N. Am. 2012, 50, 863–876. [Google Scholar] [CrossRef]
- Ginn, L.; Shi, L.; Montagna, M.; Garofalo, M. LncRNAs in Non-Small-Cell Lung Cancer. Noncoding RNA 2020, 6, 25. [Google Scholar] [CrossRef]
- Wang, C.; Zhou, J.; Wang, J.; Li, S.; Fukunaga, A.; Yodoi, J.; Tian, H. Progress in the mechanism and targeted drug therapy for COPD. Signal Transduct. Target. Ther. 2020, 5, 248. [Google Scholar] [CrossRef]
- Zhao, Y.; Feng, C.; Li, Y.; Ma, Y.; Cai, R. LncRNA H19 promotes lung cancer proliferation and metastasis by inhibiting miR-200a function. Mol. Cell. Biochem. 2019, 460, 1–8. [Google Scholar] [CrossRef]
- Roh, J.; Kim, B.; Im, M.; Jang, W.; Chae, Y.; Kang, J.; Youn, B.; Kim, W. MALAT1-regulated gene expression profiling in lung cancer cell lines. BMC Cancer 2023, 23, 818. [Google Scholar] [CrossRef]
- Wang, D.; Zhang, S.; Zhao, M.; Chen, F. LncRNA MALAT1 accelerates non-small cell lung cancer progression via regulating miR-185-5p/MDM4 axis. Cancer Med. 2020, 9, 9138–9149. [Google Scholar] [CrossRef]
- Wang, J.; Mo, J.; Xie, Y.; Wang, C. Ultrasound microbubbles-mediated miR-216b affects MALAT1-miRNA axis in non-small cell lung cancer cells. Tissue Cell 2022, 74, 101703. [Google Scholar] [CrossRef]
- Attaway, A.H.; Bellar, A.; Welch, N.; Sekar, J.; Kumar, A.; Mishra, S.; Hatipoğlu, U.; McDonald, M.L.; Regan, E.A.; Smith, J.D.; et al. Gene polymorphisms associated with heterogeneity and senescence characteristics of sarcopenia in chronic obstructive pulmonary disease. J. Cachexia Sarcopenia Muscle 2023, 14, 1083–1095. [Google Scholar] [CrossRef]
- Du, Z.; Zhang, F.; Liu, L.; Shen, H.; Liu, T.; Jin, J.; Yu, N.; Wan, Z.; Wang, H.; Hu, X.; et al. LncRNA ANRIL promotes HR repair through regulating PARP1 expression by sponging miR-7-5p in lung cancer. BMC Cancer 2023, 23, 130. [Google Scholar] [CrossRef]
- Li, K.; Niu, H.; Wang, Y.; Li, R.; Zhao, Y.; Liu, C.; Cao, H.; Chen, H.; Xie, R.; Zhuang, L. LncRNA TUG1 contributes to the tumorigenesis of lung adenocarcinoma by regulating miR-138-5p-HIF1A axis. Int. J. Immunopathol. Pharmacol. 2021, 35, 20587384211048265. [Google Scholar] [CrossRef]
- Qi, F.; Lv, Z.D.; Huang, W.D.; Wei, S.C.; Liu, X.M.; Song, W.D. LncRNA TUG1 promotes pulmonary fibrosis progression via up-regulating CDC27 and activating PI3K/Akt/mTOR pathway. Epigenetics 2023, 18, 2195305. [Google Scholar] [CrossRef]
- Entezari, M.; Ghanbarirad, M.; Taheriazam, A.; Sadrkhanloo, M.; Zabolian, A.; Goharrizi, M.A.S.B.; Hushmandi, K.; Aref, A.R.; Ashrafizadeh, M.; Zarrabi, A.; et al. Long non-coding RNAs and exosomal lncRNAs: Potential functions in lung cancer progression, drug resistance and tumor microenvironment remodeling. Biomed. Pharmacother. 2022, 150, 112963. [Google Scholar] [CrossRef]
- Gupta, S.; Hashimoto, R.F. Dynamical Analysis of a Boolean Network Model of the Oncogene Role of lncRNA ANRIL and lncRNA UFC1 in Non-Small Cell Lung Cancer. Biomolecules 2022, 12, 420. [Google Scholar] [CrossRef]
- Li, Z.; Yu, X.; Shen, J. ANRIL: A pivotal tumor suppressor long non-coding RNA in human cancers. Tumour Biol. 2016, 37, 5657–5661. [Google Scholar] [CrossRef]
- Kang, Y.H.; Kim, D.; Jin, E.J. Down-regulation of Phospholipase D Stimulates Death of Lung Cancer Cells Involving Up-regulation of the Long ncRNA ANRIL. Anticancer. Res. 2015, 35, 2795–2803. Available online: https://pubmed.ncbi.nlm.nih.gov/25964559/ (accessed on 1 January 2024). [PubMed]
- Zhang, H.; Li, X.; Jia, M.; Ji, J.; Wu, Z.; Chen, X.; Yu, D.; Zheng, Y.; Zhao, Y. Roles of H19/miR-29a-3p/COL1A1 axis in COE-induced lung cancer. Environ. Pollut. 2022, 313, 120194. [Google Scholar] [CrossRef]
- Hu, X.T.; Xing, W.; Zhao, R.S.; Tan, Y.; Wu, X.F.; Ao, L.Q.; Li, Z.; Yao, M.W.; Yuan, M.; Guo, W.; et al. HDAC2 inhibits EMT-mediated cancer metastasis by downregulating the long noncoding RNA H19 in colorectal cancer. J. Exp. Clin. Cancer Res. 2020, 39, 270. [Google Scholar] [CrossRef]
- Lai, X.; Zhong, J.; Zhang, A.; Zhang, B.; Zhu, T.; Liao, R. Focus on long non-coding RNA MALAT1: Insights into acute and chronic lung diseases. Front Genet. 2022, 13, 1003964. [Google Scholar] [CrossRef]
- Bhat, A.A.; Afzal, O.; Afzal, M.; Gupta, G.; Thapa, R.; Ali, H.; Hassan Almalki, W.; Kazmi, I.; Alzarea, S.I.; Saleem, S.; et al. MALAT1: A key regulator in lung cancer pathogenesis and therapeutic targeting. Pathol. Res. Pract. 2024, 253, 154991. [Google Scholar] [CrossRef]
- Qiu, N.; Xu, X.; He, Y. LncRNA TUG1 alleviates sepsis-induced acute lung injury by targeting miR-34b-5p/GAB1. BMC Pulm. Med. 2020, 20, 49. [Google Scholar] [CrossRef]
- Li, Z.; Shen, J.; Chan, M.T.; Wu, W.K. TUG1: A Pivotal Oncogenic Long Non-Coding RNA of Human Cancers. Cell Prolif. 2016, 49, 471–475. [Google Scholar] [CrossRef]
- Loewen, G.; Jayawickramarajah, J.; Zhuo, Y.; Shan, B. Functions of lncRNA HOTAIR in lung cancer. J. Hematol. Oncol. 2014, 7, 90. [Google Scholar] [CrossRef]
- Ren, M.M.; Xu, S.; Wei, Y.B.; Yang, J.J.; Yang, Y.N.; Sun, S.S.; Li, Y.J.; Wang, P.Y.; Xie, S.Y. Roles of HOTAIR in lung cancer susceptibility and prognosis. Mol. Genet. Genomic Med. 2020, 8, e1299. [Google Scholar] [CrossRef] [PubMed]
- Tang, L.; Wang, S.; Wang, Y.; Li, K.; Li, Q. LncRNA-UCA1 regulates lung adenocarcinoma progression through competitive binding to miR-383. Cell Cycle 2023, 22, 213–228. [Google Scholar] [CrossRef]
- Shi, W.; Ling, L.; Li, C.; Wu, R.; Zhang, M.; Shao, F.; Wang, Y. LncRNA UCA1 promoted cisplatin resistance in lung adenocarcinoma with HO1 targets NRF2/HO1 pathway. J. Cancer Res. Clin. Oncol. 2023, 149, 1295–1311. [Google Scholar] [CrossRef]
- Szafranski, P.; Stankiewicz, P. Long Non-Coding RNA FENDRR: Gene Structure, Expression, and Biological Relevance. Genes (Basel). 2021, 12, 177. [Google Scholar] [CrossRef]
- Herrera-Merchan, A.; Cuadros, M.; Rodriguez, M.I.; Rodriguez, S.; Torres, R.; Estecio, M.; Coira, I.F.; Loidi, C.; Saiz, M.; Carmona-Saez, P.; et al. The value of lncRNA FENDRR and FOXF1 as a prognostic factor for survival of lung adenocarcinoma. Oncotarget 2017, 11, 1172–1185. [Google Scholar] [CrossRef] [PubMed]
- Ghafouri-Fard, S.; Shirvani-Farsani, Z.; Hussen, B.M.; Taheri, M.; Samsami, M. The key roles of non-coding RNAs in the pathophysiology of hypertension. Eur. J. Pharmacol. 2022, 931, 175220. [Google Scholar] [CrossRef] [PubMed]
- Wu, J.; Ma, Y.; Chen, Y. Extracellular vesicles and COPD: Foe or friend? J. Nanobiotechnol. 2023, 21, 147. [Google Scholar] [CrossRef]
- Fanale, D.; Taverna, S.; Russo, A.; Bazan, V. Circular RNA in Exosomes. Adv. Exp. Med. Biol. 2018, 1087, 109–117. [Google Scholar] [CrossRef]
- Sarfi, M.; Abbastabar, M.; Khalili, E. Long noncoding RNAs biomarker-based cancer assessment. J. Cell Physiol. 2019, 234, 16971–16986. [Google Scholar] [CrossRef]
- Reggiardo, R.E.; Maroli, S.V.; Kim, D.H. LncRNA Biomarkers of Inflammation and Cancer. Adv. Exp. Med. Biol. 2022, 1363, 121–145. [Google Scholar] [CrossRef]
- Chi, Y.; Wang, D.; Wang, J.; Yu, W.; Yang, J. Long Non-Coding RNA in the Pathogenesis of Cancers. Cells 2019, 8, 1015. [Google Scholar] [CrossRef] [PubMed]
- Chen, B.B.; Li, Z.H.; Gao, S. Circulating miR-146a/b Correlates with Inflammatory Cytokines in COPD and Could Predict the Risk of Acute Exacerbation COPD. Medicine 2018, 97, e9820. [Google Scholar] [CrossRef] [PubMed]
- Hoult, G.; Gillespie, D.; Wilkinson, T.M.A.; Thomas, M.; Francis, N.A. Biomarkers to Guide the Use of Antibiotics for Acute Exacerbations of COPD (AECOPD): A Systematic Review and Meta-Analysis. BMC Pulm. Med. 2022, 22, 194. [Google Scholar] [CrossRef] [PubMed]
- Walcher, L.; Kistenmacher, A.K.; Suo, H.; Kitte, R.; Dluczek, S.; Strauß, A.; Blaudszun, A.R.; Yevsa, T.; Fricke, S.; Kossatz-Boehlert, U. Cancer Stem Cells-Origins and Biomarkers: Perspectives for Targeted Personalized Therapies. Front. Immunol. 2020, 11, 1280. [Google Scholar] [CrossRef] [PubMed]
- Vogelmeier, C.F.; Román-Rodríguez, M.; Singh, D.; Han, M.K.; Rodríguez-Roisin, R.; Ferguson, G.T. Goals of COPD Treatment: Focus on Symptoms and Exacerbations. Respir. Med. 2020, 166, 105938. [Google Scholar] [CrossRef] [PubMed]
- Barnes, P.J. Inflammatory Endotypes in COPD. Allergy 2019, 74, 1249–1256. [Google Scholar] [CrossRef] [PubMed]
- Ma, Z.; Liang, H.; Cui, R.; Ji, J.; Liu, H.; Liu, X.; Shen, P.; Wang, H.; Wang, X.; Song, Z.; et al. Construction of a Risk Model and Prediction of Prognosis and Immunotherapy Based on Cuproptosis-Related LncRNAs in the Urinary System Pan-Cancer. Eur. J. Med. Res. 2023, 28, 198. [Google Scholar] [CrossRef] [PubMed]
- Brisotto, G.; Guerrieri, R.; Colizzi, F.; Steffan, A.; Montico, B.; Fratta, E. Long Noncoding RNAs as Innovative Urinary Diagnostic Biomarkers. Methods Mol. Biol. 2021, 2292, 73–94. [Google Scholar] [CrossRef] [PubMed]
- Terracciano, D.; Ferro, M.; Terreri, S.; Lucarelli, G.; D’Elia, C.; Musi, G.; de Cobelli, O.; Mirone, V.; Cimmino, A. Urinary Long Noncoding RNAs in Nonmuscle-Invasive Bladder Cancer: New Architects in Cancer Prognostic Biomarkers. Transl. Res. 2017, 184, 108–117. [Google Scholar] [CrossRef] [PubMed]
- Wang, Y.; Fu, X.; Yu, B.; Ai, F. Long Non-Coding RNA THRIL Predicts Increased Acute Respiratory Distress Syndrome Risk and Positively Correlates with Disease Severity, Inflammation, and Mortality in Sepsis Patients. J. Clin. Lab. Anal. 2019, 33, e22882. [Google Scholar] [CrossRef] [PubMed]
- Lin, Q.; Xie, D.; Pan, L.; Lou, Y.; Shi, M. Urinary Exosomal Long Noncoding RNAs Serve as Biomarkers for Early Detection of Non-Small Cell Lung Cancer. Biosci. Rep. 2021, 41, BSR20210908. [Google Scholar] [CrossRef]
- Zhang, Y.; Luo, G.; Zhang, Y.; Zhang, M.; Zhou, J.; Gao, W.; Xuan, X.; Yang, X.; Yang, D.; Tian, Z.; et al. Critical Effects of Long Non-Coding RNA on Fibrosis Diseases. Exp. Mol. Med. 2018, 50, e428. [Google Scholar] [CrossRef]
- Wang, H.; Feng, Y.; Zheng, X.; Xu, X. The Diagnostic and Therapeutic Role of snoRNA and LincRNA in Bladder Cancer. Cancers 2023, 15, 1007. [Google Scholar] [CrossRef]
- Zhang, H.; Wang, S.Q.; Wang, L.; Lin, H.; Zhu, J.B.; Chen, R.; Li, L.F.; Cheng, Y.D.; Duan, C.J.; Zhang, C.F. m6A Methyltransferase METTL3-Induced LncRNA SNHG17 Promotes Lung Adenocarcinoma Gefitinib Resistance by Epigenetically Repressing LATS2 Expression. Cell Death Dis. 2022, 13, 657. [Google Scholar] [CrossRef]
- Chen, H.; Wang, L.; Liu, J.; Wan, Z.; Zhou, L.; Liao, H.; Wan, R. LncRNA ITGB2-AS1 Promotes Cisplatin Resistance of Non-Small Cell Lung Cancer by Inhibiting Ferroptosis via Activating the FOSL2/NAMPT Axis. Cancer Biol. Ther. 2023, 24, 2223377. [Google Scholar] [CrossRef]
- Ma, H.; Hu, T.; Tao, W.; Tong, J.; Han, Z.; Herndler-Brandstetter, D.; Wei, Z.; Liu, R.; Zhou, T.; Liu, Q.; et al. A LncRNA from an Inflammatory Bowel Disease Risk Locus Maintains Intestinal Host-Commensal Homeostasis. Cell Res. 2023, 33, 372–388. [Google Scholar] [CrossRef]
- Lai, X.N.; Li, J.; Tang, L.B.; Chen, W.T.; Zhang, L.; Xiong, L.X. MiRNAs and LncRNAs: Dual Roles in TGF-β Signaling-Regulated Metastasis in Lung Cancer. Int. J. Mol. Sci. 2020, 21, 1193. [Google Scholar] [CrossRef] [PubMed]
- Du, Z.; Wang, B.; Tan, F.; Wu, Y.; Chen, J.; Zhao, F.; Liu, M.; Zhou, G.; Yuan, C. The Regulatory Role of LncRNA HCG18 in Various Cancers. J. Mol. Med. (Berl.) 2023, 101, 351–360. [Google Scholar] [CrossRef]
- Hansel, N.N.; McCormack, M.C.; Kim, V. The Effects of Air Pollution and Temperature on COPD. COPD J. Chronic Obstr. Pulm. Dis. 2016, 13, 372–379. [Google Scholar] [CrossRef]
- Hu, Q.; Egranov, S.D.; Lin, C.; Yang, L. Long Noncoding RNA Loss in Immune Suppression in Cancer. Pharmacol. Ther. 2020, 213, 107591. [Google Scholar] [CrossRef] [PubMed]
- Chen, C.; Ding, P.; Yan, W.; Wang, Z.; Lan, Y.; Yan, X.; Li, T.; Han, J. Pharmacological Roles of LncRNAs in Diabetic Retinopathy with a Focus on Oxidative Stress and Inflammation. Biochem. Pharmacol. 2023, 214, 115643. [Google Scholar] [CrossRef]
- Zeng, L.; Zhou, M.; Wang, X.; Long, X.; Ye, M.; Yuan, Y.; Tan, W. Aberrant LncRNA Expression in Patients with Proliferative Diabetic Retinopathy: Preliminary Results from a Single-Center Observational Study. BMC Ophthalmol. 2023, 23, 94. [Google Scholar] [CrossRef] [PubMed]
- Mohammad, H.M.F.; Abdelghany, A.A.; Al Ageeli, E.; Kattan, S.W.; Hassan, R.; Toraih, E.A.; Fawzy, M.S.; Mokhtar, N. Long Non-Coding RNAs Gene Variants as Molecular Markers for Diabetic Retinopathy Risk and Response to Anti-VEGF Therapy. Pharmgenomics Pers. Med. 2021, 14, 997–1014. [Google Scholar] [CrossRef] [PubMed]
- Foronjy, R. Commentary on: The Potency of LncRNA MALAT1/miR-155 in Altering Asthmatic Th1/Th2 Balance by Modulation of CTLA4. Biosci. Rep. 2020, 40, BSR20190768. [Google Scholar] [CrossRef]
- Mei, D.; Tan, W.S.D.; Tay, Y.; Mukhopadhyay, A.; Wong, W.S.F. Therapeutic RNA Strategies for Chronic Obstructive Pulmonary Disease. Trends Pharmacol. Sci. 2020, 41, 475–486. [Google Scholar] [CrossRef]
- Sato, T.; Baskoro, H.; Rennard, S.I.; Seyama, K.; Takahashi, K. MicroRNAs as Therapeutic Targets in Lung Disease: Prospects and Challenges. Chronic Obstr. Pulm. Dis. 2015, 3, 382–388. [Google Scholar] [CrossRef]
- Castaldi, P.J.; Guo, F.; Qiao, D.; Du, F.; Naing, Z.Z.C.; Li, Y.; Pham, B.; Mikkelsen, T.S.; Cho, M.H.; Silverman, E.K.; et al. Identification of Functional Variants in the FAM13A Chronic Obstructive Pulmonary Disease Genome-Wide Association Study Locus by Massively Parallel Reporter Assays. Am. J. Respir. Crit. Care Med. 2019, 199, 52–61. [Google Scholar] [CrossRef] [PubMed]
- Vij, N. Nano-based theranostics for chronic obstructive lung diseases: Challenges and therapeutic potential. Expert Opin. Drug Deliv. 2011, 8, 1105–1109. [Google Scholar] [CrossRef]
- Pleasants, R.A.; Hess, D.R. Aerosol Delivery Devices for Obstructive Lung Diseases. Respir. Care 2018, 63, 708–733. [Google Scholar] [CrossRef]
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Sweef, O.; Mahfouz, R.; Taşcıoğlu, T.; Albowaidey, A.; Abdelmonem, M.; Asfar, M.; Zaabout, E.; Corcino, Y.L.; Thomas, V.; Choi, E.-S.; et al. Decoding LncRNA in COPD: Unveiling Prognostic and Diagnostic Power and Their Driving Role in Lung Cancer Progression. Int. J. Mol. Sci. 2024, 25, 9001. https://doi.org/10.3390/ijms25169001
Sweef O, Mahfouz R, Taşcıoğlu T, Albowaidey A, Abdelmonem M, Asfar M, Zaabout E, Corcino YL, Thomas V, Choi E-S, et al. Decoding LncRNA in COPD: Unveiling Prognostic and Diagnostic Power and Their Driving Role in Lung Cancer Progression. International Journal of Molecular Sciences. 2024; 25(16):9001. https://doi.org/10.3390/ijms25169001
Chicago/Turabian StyleSweef, Osama, Reda Mahfouz, Tülin Taşcıoğlu, Ali Albowaidey, Mohamed Abdelmonem, Malek Asfar, Elsayed Zaabout, Yalitza Lopez Corcino, Venetia Thomas, Eun-Seok Choi, and et al. 2024. "Decoding LncRNA in COPD: Unveiling Prognostic and Diagnostic Power and Their Driving Role in Lung Cancer Progression" International Journal of Molecular Sciences 25, no. 16: 9001. https://doi.org/10.3390/ijms25169001