
A Simple Expression for the Screening of Excitonic Couplings between Chlorophylls as Inferred for Photosystem I Trimers

Abstract
1. Introduction
2. Theory
Poisson-TrEsp Method and Screening Factor
3. Results
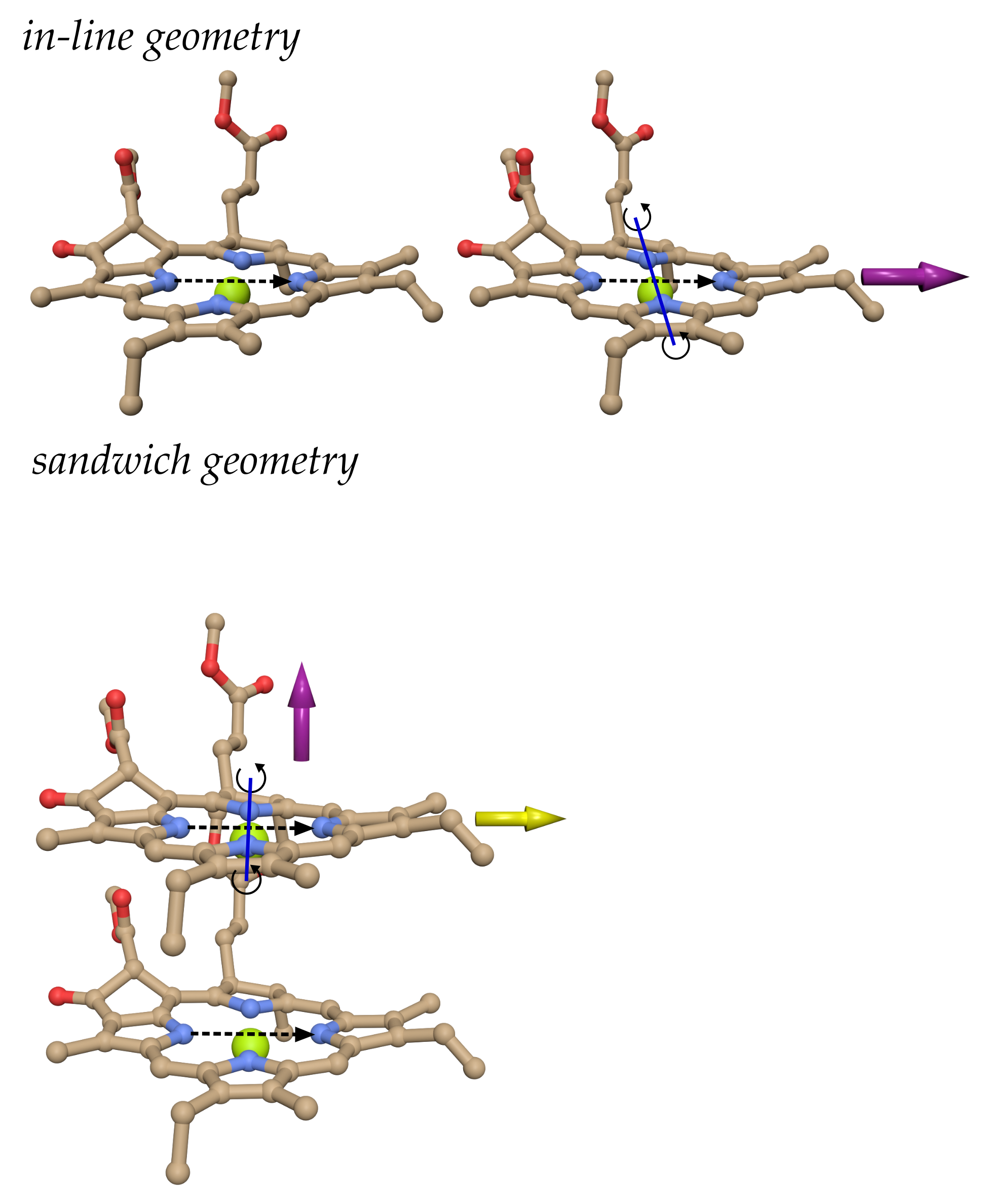
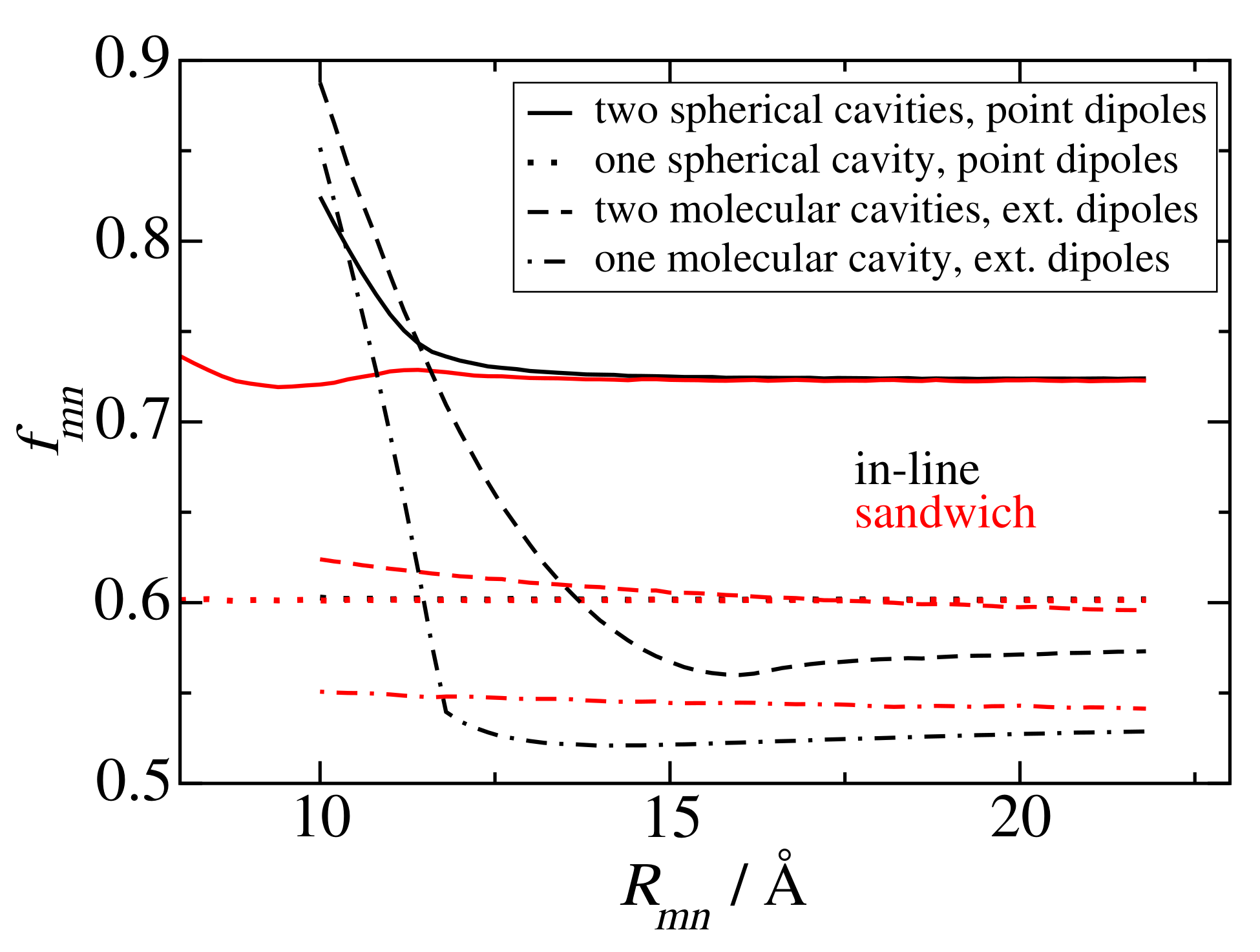
3.1. Screening Factors of PSI Trimer in Many-Cavity versus Two-Cavity Model
3.2. Dependence of Screening Factors on Mutual Orientation and Distance
4. Discussion
Model Calculations
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Data Availability Statement
Conflicts of Interest
Abbreviations
References
- Nogales, E. The development of cryo-EM into a mainstream structural biology technique. Nat. Methods 2016, 13, 24–27. [Google Scholar] [CrossRef] [PubMed]
- Cheng, Y. Single-particle cryo-EM—How did it get here and where will it go. Science 2018, 361, 876–880. [Google Scholar] [CrossRef] [PubMed]
- Croce, R.; van Amerongen, H. Light harvesting in oxygenic photosynthesis: Structural biology meets spectroscopy. Science 2020, 369, 933. [Google Scholar] [CrossRef] [PubMed]
- Pi, X.; Zhao, S.H.; Wang, W.D.; Liu, D.S.; Xu, C.Z.; Han, G.Y.; Kuang, T.Y.; Sui, S.F.; Shen, J.R. The pigment-protein network of a diatom photosystem II-light-harvesting antenna supercomplex. Science 2019, 365, 463. [Google Scholar] [CrossRef] [PubMed]
- Xie, H.; Lyratzakis, A.; Khera, R.; Kountantou, M.; Welsch, S.; Michel, H.; Tsiotis, G. Cryo-EM structure of the whole photosynthetic reaction center apparatus from the green sulfur bacterium Chlorobaculum tepidum. Proc. Natl. Acad. Sci. USA 2023, 120, e2216734120. [Google Scholar] [CrossRef] [PubMed]
- Zhang, S.; Tang, K.; Yan, Q.; Li, X.; Shen, L.; Wang, W.; He, Y.K.; Kuang, T.; Han, G.; Shen, J.R.; et al. Structural insights into a unique PSI—LHCI—LHCII—Lhcb9 supercomplex from moss Physcomitrium patens. Nat. Plants 2023, 9, 832–846. [Google Scholar] [CrossRef] [PubMed]
- Raszewski, G.; Renger, T. Light harvesting in photosystem II core complexes is limited by the transfer to the trap: Can the core complex turn into a photoprotective mode? J. Am. Chem. Soc. 2008, 130, 4431–4446. [Google Scholar] [CrossRef] [PubMed]
- Bennett, D.I.G.; Amarnath, K.; Fleming, G.R. A structure-based model of energy transfer reveals the principles of light harvesting in photosystem II supercomplexes. J. Am. Chem. Soc. 2013, 135, 9164–9173. [Google Scholar] [CrossRef] [PubMed]
- Amarnath, K.; Bennett, D.I.G.; Schneider, A.R.; Fleming, G.R. Multiscale model of light harvesting by photosystem II in plants. Proc. Natl. Acad. Sci. USA 2016, 113, 1156–1161. [Google Scholar] [CrossRef] [PubMed]
- Klinger, A.; Lindorfer, D.; Müh, F.; Renger, T. Living on the edge: Light-harvesting efficiency and photoprotection in the core of green sulfur bacteria. Phys. Chem. Chem. Phys. 2023, 25, 18698–18710. [Google Scholar] [CrossRef] [PubMed]
- Chmeliov, J.; Trinkunas, G.; van Amerongen, H.; Valkunas, L. Light harvesting in a fluctuating antenna. J. Am. Chem. Soc. 2014, 136, 8963–8972. [Google Scholar] [CrossRef] [PubMed]
- Jordan, P.; Fromme, P.; Klukas, O.; Witt, H.T.; Saenger, W.; Krauß, N. Three dimensional structure of cyanobacterial photosystem I at 2.5 Å resolution. Nature 2001, 411, 909–917. [Google Scholar] [CrossRef] [PubMed]
- Blender Online Community. Blender—A 3D Modelling and Rendering Package; Stichting Blender Foundation: Amsterdam, The Netherlands, 2024; Available online: http://www.blender.org (accessed on 1 July 2024).
- Ishizaki, A.; Fleming, G.R. Unified treatment of quantum coherent and incoherent hopping dynamics in electronic energy transfer: Reduced hierarchy equation approach. J. Chem. Phys. 2009, 130, 234111. [Google Scholar] [CrossRef] [PubMed]
- Huo, P.F.; Coker, D.F. Consistent schemes for non-adiabatic dynamics derived from partial linearized density matrix propagation. J. Chem. Phys. 2012, 137, 22A535. [Google Scholar] [CrossRef] [PubMed]
- Kreisbeck, C.; Kramer, T.; Aspuru-Guzik, A. Scalable high-performance algorithm for the simulation of exciton dynamics. Application to the light-harvesting complex II in the presence of resonant vibrational modes. J. Chem. Theory Comput. 2014, 10, 4045–4054. [Google Scholar] [CrossRef] [PubMed]
- Ma, J.; Cao, J.S. Förster resonance energy transfer, absorption and emission spectra in multichromophoric systems: I. Full cumulant expansions and system-bath entanglement. J. Chem. Phys. 2014, 142, 094106. [Google Scholar] [CrossRef] [PubMed]
- Tanimura, Y. Numerically “exact” approach to open quantum dynamics: The hierarchical equations of motion (HEOM). J. Chem. Phys. 2020, 153, 020901. [Google Scholar] [CrossRef] [PubMed]
- Reppert, M. Delocalization effects in chlorophyll fluorescence: Nonperturbative lineshape analysis of a vibronically coupled dimer. J. Phys. Chem. B 2020, 124, 10024–10033. [Google Scholar] [CrossRef] [PubMed]
- Caycedo-Soler, F.; Mattioni, A.; Lim, J.; Renger, T.; Huelga, S.F.; Plenio, M. Exact simulation of pigment-protein complexes unveils vibronic renormalization of electronic parameters in ultrafast spectroscopy. Nat. Commun. 2022, 13, 2912. [Google Scholar] [CrossRef] [PubMed]
- Kundu, S.; Makri, N. Intramolecular vibrations in excitation energy transfer: Insights from real-time path integral calculations. Annu. Rev. Phys. Chem. 2022, 73, 49–75. [Google Scholar] [CrossRef] [PubMed]
- Hsu, C. Theoretical study of photosynthetic light-harvesting processes: Application of time-dependent density functional theory. J. Chin. Chem. Soc. 2003, 50, 745–756. [Google Scholar] [CrossRef]
- Dreuw, A.; Harbach, P.H.P.; Mewes, J.M.; Wormit, M. Quantum chemical excited state calculations on pigment-protein complexes require thorough geometry re-optimization of experimental crystal structures. Theor. Chem. Acc. 2010, 125, 419–426. [Google Scholar] [CrossRef]
- Renger, T.; Müh, F. Understanding photosynthetic light-harvesting: A bottom up theoretical approach. Phys. Chem. Chem. Phys. 2013, 15, 3348–3371. [Google Scholar] [CrossRef] [PubMed]
- Renger, T.; Madjet, M.E.A.; Schmidt am Busch, M.; Adolphs, J.; Müh, F. Structure-based modeling of energy transfer in photosynthesis. Photosynth. Res. 2013, 116, 367–388. [Google Scholar] [CrossRef] [PubMed]
- König, C.; Neugebauer, J. Quantum chemical description of absorption properties and excited-state processes in photosynthtetic systems. Chem. Phys. Chem. 2012, 13, 386–425. [Google Scholar] [CrossRef] [PubMed]
- Steinmann, C.; Kongsted, J. Electronic energy transfer in polarizable heterogeneous environments: A systematic investigation of different quantum chemical approaches. J. Chem. Theory Comput. 2015, 11, 4283–4293. [Google Scholar] [CrossRef] [PubMed]
- Curutchet, C.; Mennucci, B. Quantum chemical studies of light harvesting. Chem. Rev. 2017, 117, 294–343. [Google Scholar] [CrossRef] [PubMed]
- Jang, S.J.; Mennucci, B. Delocalized excitons in natural light-harvesting complexes. Rev. Mod. Phys. 2018, 90. [Google Scholar] [CrossRef]
- Cupellini, L.; Bondanza, M.; Nottoli, M.; Mennucci, B. Successes and challenges in the atomistic modeling of light-harvesting and its photoregulation. Biochim. Biophys. Acta 2020, 1861, 148049. [Google Scholar] [CrossRef] [PubMed]
- Chang, J.C. Monopole effects on electronic excitation interactions between large molecules. I. Application to energy transfer in chlorophylls. J. Chem. Phys. 1977, 67, 3901–3909. [Google Scholar] [CrossRef]
- Krüger, B.P.; Scholes, G.D.; Fleming, G.R. Calculation of couplings and energy transfer pathways between pigments of LH2 by the ab-initio transition density cube method. J. Phys. Chem. B 2005, 4, 744–753. [Google Scholar]
- Hsu, C.; Head-Gordon, M.; Head-Gordon, T.; Fleming, G.R. Excitation energy transfer in condensed media. J. Chem. Phys. 2001, 114, 3065–3072. [Google Scholar] [CrossRef]
- Iozzi, M.F.; Mennucci, B.; Tomasi, J.; Cammi, R. Excitation energy transfer (EET) between molecules in condensed matter: A novel application of the polarizable continuum model (PCM). J. Chem. Phys. 2004, 120, 7029–7040. [Google Scholar] [CrossRef]
- Madjet, M.E.; Abdurahman, A.; Renger, T. Intermolecular Coulomb couplings from ab initio electrostatic potentials: Application to optical transitions of strongly coupled pigments in photosynthetic antennae and reaction centers. J. Phys. Chem. B 2006, 110, 17268–17281. [Google Scholar] [CrossRef]
- Curutchet, C.; Scholes, G.D.; Mennucci, B.; Cammi, R. How solvent controls electronic energy transfer and light harvesting: Toward a quantum-mechanical description of reaction field and screening effects. J. Phys. Chem. B 2007, 111, 13253–13265. [Google Scholar] [CrossRef] [PubMed]
- Adolphs, J.; Müh, F.; Madjet, M.E.; Renger, T. Calculation of pigment transition energies in the FMO protein: From simplicity to complexity and back. Photosynth. Res. 2008, 95, 197–209. [Google Scholar] [CrossRef]
- Fujimoto, K.J.; Hayashi, S. Electronic Coulombic coupling of excitation-energy transfer in xanthorhodopsin. J. Am. Chem. Soc. 2009, 131, 14152. [Google Scholar] [CrossRef]
- Curutchet, C.; Kongsted, J.; Munoz-Losa, A.; Hossein-Nejad, H.; Scholes, G.D.; Mennucci, B. Photosynthetic light-harvesting is tuned by the heterogeneous polarizable environment of the protein. J. Am. Chem. Soc. 2011, 133, 3078–3084. [Google Scholar] [CrossRef]
- Renger, T.; Müh, F. Theory of excitonic couplings in dielectric media. Photosynth. Res. 2012, 111, 47–52. [Google Scholar] [CrossRef] [PubMed]
- Blasiak, B.; Maj, M.; Cho, M.; Gora, R.W. Distributed multipolar expansion approach to calculation of excitation energy transfer couplings. J. Chem. Theory Comput. 2015, 11, 3259–3266. [Google Scholar] [CrossRef]
- Cignoni, E.; Cupellini, L.; Mennucci, B. A fast method for electronic couplings in embedded multichromophoric systems. J. Phys. Condens. Matter 2022, 34, 304004. [Google Scholar] [CrossRef] [PubMed]
- Kitoh-Nishioka, H.; Shigeta, Y.; Iro, S.; Kimura, A. Excitonic coupling on a heliobacterial symmetrical type-I reaction center: Comparison with photosystem I. J. Phys. Chem. B 2020, 124, 389–403. [Google Scholar] [CrossRef] [PubMed]
- Scholes, G.D.; Gould, I.R.; Cogdell, R.J.; Fleming, G.R. Ab Initio molecular orbital calculations of electronic couplings in the LH2 bacterial light-harvesting complex of Rps. Acidophila. J. Phys. Chem. B 1999, 103, 2543–2553. [Google Scholar] [CrossRef]
- Madjet, M.E.; Müh, F.; Renger, T. Deciphering the influence of short-range electronic couplings on optical properties of molecular dimers: Application to “special pairs” in photosynthesis. J. Phys. Chem. B 2009, 113, 12603–12614. [Google Scholar] [CrossRef] [PubMed]
- Cupellini, L.; Caprasecca, S.; Guido, C.A.; Müh, F.; Renger, T.; Mennucci, B. Coupling to charge transfer states is the key to modulate the optical bands for efficient light harvesting in purple bacteria. J. Phys. Chem. Lett. 2018, 9, 6892–6899. [Google Scholar] [CrossRef] [PubMed]
- Gemeinhardt, F.G.; Lahav, Y.; Schapiro, I.; Noy, D.; Müh, F.; Lindorfer, D.; Renger, T. Short-range effects in the special pair of photosystem II reaction centers: The nonconservative nature of circular dichroism. J. Phys. Chem. Lett. 2023, 14, 11758–11767. [Google Scholar] [CrossRef] [PubMed]
- Knox, R.S.; Spring, B.Q. Dipole strengths in the chlorophylls. Photochem. Photobiol. 2003, 77, 497–501. [Google Scholar] [CrossRef] [PubMed]
- Friedl, C.; Fedorov, D.G.; Renger, T. Towards a quantitative description of excitonic couplings in photosynthetic pigment-protein complexes: Quantum chemistry driven multiscale approaches. Phys. Chem. Chem. Phys. 2022, 24, 5014–5038. [Google Scholar] [CrossRef] [PubMed]
- Scholes, G.D.; Curutchet, C.; Mennucci, B.; Cammi, R.; Tomasi, J. How solvent controls electronic energy transfer and light harvesting. J. Phys. Chem. Lett. 2007, 111, 6978–6982. [Google Scholar] [CrossRef] [PubMed]
- Slama, V.; Müh, F.; Renger, T.; Mancal, T. Role of environmental dynamic polarizability in static excited state properties of embedded molecular systems: Application to disordered fluorographene systems. J. Phys. Chem. C 2023, 127, 381–392. [Google Scholar] [CrossRef]
- Atkins, P.W.; Friedman, R.S. Molecular Quantum Mechanics; Oxford University Press: Oxford, UK, 2005; p. 424. [Google Scholar]
- Adolphs, J.; Renger, T. How proteins trigger excitation energy transfer in the FMO complex of green sulfur bacteria. Biophys. J. 2006, 91, 2778–2797. [Google Scholar] [CrossRef] [PubMed]
- Bashford, D.; Gerwert, K. Electrostatic calculations of the pKa values of ionizable groups in bacteriorhodopsin. J. Mol. Biol. 1992, 224, 473–486. [Google Scholar] [CrossRef] [PubMed]
- Bashford, D. An object-oriented programming suite for electrostatic effects in biological molecules. In Scientific Computing in Object-Oriented Parallel Environments; Yutaka, I., Rodney, R.O., John, V.W.R., Marydell, T., Eds.; CRC Press: Boca Raton, FL, USA, 1997; pp. 233–240. [Google Scholar]
- Renger, T.; Madjet, M.E.; Müh, F.; Trostmann, I.; Schmitt, F.J.; Theiss, C.; Paulsen, H.; Eichler, H.J.; Knorr, A.; Renger, G. Thermally activated superradiance and intersystem crossing in the water-soluble chlorophyll-binding protein. J. Phys. Chem. B 2009, 113, 9948–9957. [Google Scholar] [CrossRef] [PubMed]
- Müh, F.; Zouni, A. Extinction coefficients and critical solubilisation concentrations of photosystems I and II from Thermosynechococcus elongatus. Biochim. Biophys. Acta 2005, 1708, 219–228. [Google Scholar] [CrossRef] [PubMed]
- Böttcher, C.J.F. Theory of Electric Polarization; Elsevier: Amsterdam, The Netherlands, 1973; pp. 130–134. [Google Scholar]
- Agranovich, V.M.; Galanin, M.D. Electronic Excitation Energy Transfer in Condensed Matter; North-Holland: Amsterdam, The Netherlands, 1982; p. 13. [Google Scholar]
- Juzeliunas, G.; Andrews, D.L. Quantum electrodynamics of resonant energy transfer in condensed matter. Phys. Rev. B 1994, 49, 8751–8763. [Google Scholar] [CrossRef] [PubMed]
- Renger, T.; Grundkötter, B.; Madjet, M.E.; Müh, F. Theory of solvatochromic shifts in nonpolar solvents reveals a new spectroscopic rule. Proc. Natl. Acad. Sci. USA 2008, 105, 13235–13240. [Google Scholar] [CrossRef] [PubMed]
- Renger, T.; Dankl, M.; Klinger, A.; Schlücker, T.; Langhals, H.; Müh, F. Structure-based theory of fluctuation-induced energy transfer in a molecular dyad. J. Phys. Chem. Lett. 2018, 9, 5940–5947. [Google Scholar] [CrossRef] [PubMed]








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Orientation | |
|---|---|
 | 0 |
 | 0.6 |
 | 1 |
 | 1.45 |
 | 2 |
| Group | Geometry |
|---|---|
| 1 | |
| 2 | |
| 3 |
| / | |||
|---|---|---|---|
| Group | (a) | (b) | (c) |
| 1 | 0.47 | 2.64 | 0.47 |
| 2 | 1.06 | 11.38 | 0.94 |
| 3 | 12.90 | 0.72 | 0.82 |
| all | 2.55 | 6.00 | 0.72 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Eder, M.; Renger, T. A Simple Expression for the Screening of Excitonic Couplings between Chlorophylls as Inferred for Photosystem I Trimers. Int. J. Mol. Sci. 2024, 25, 9006. https://doi.org/10.3390/ijms25169006
Eder M, Renger T. A Simple Expression for the Screening of Excitonic Couplings between Chlorophylls as Inferred for Photosystem I Trimers. International Journal of Molecular Sciences. 2024; 25(16):9006. https://doi.org/10.3390/ijms25169006
Chicago/Turabian StyleEder, Matthias, and Thomas Renger. 2024. "A Simple Expression for the Screening of Excitonic Couplings between Chlorophylls as Inferred for Photosystem I Trimers" International Journal of Molecular Sciences 25, no. 16: 9006. https://doi.org/10.3390/ijms25169006
APA StyleEder, M., & Renger, T. (2024). A Simple Expression for the Screening of Excitonic Couplings between Chlorophylls as Inferred for Photosystem I Trimers. International Journal of Molecular Sciences, 25(16), 9006. https://doi.org/10.3390/ijms25169006