
QTL Mapping and Candidate Gene Analysis for Starch-Related Traits in Tartary Buckwheat (Fagopyrum tataricum (L.) Gaertn)

 , , and
, , and
Abstract
1. Introduction
2. Results
2.1. Variation Analysis for Starch-Related Traits
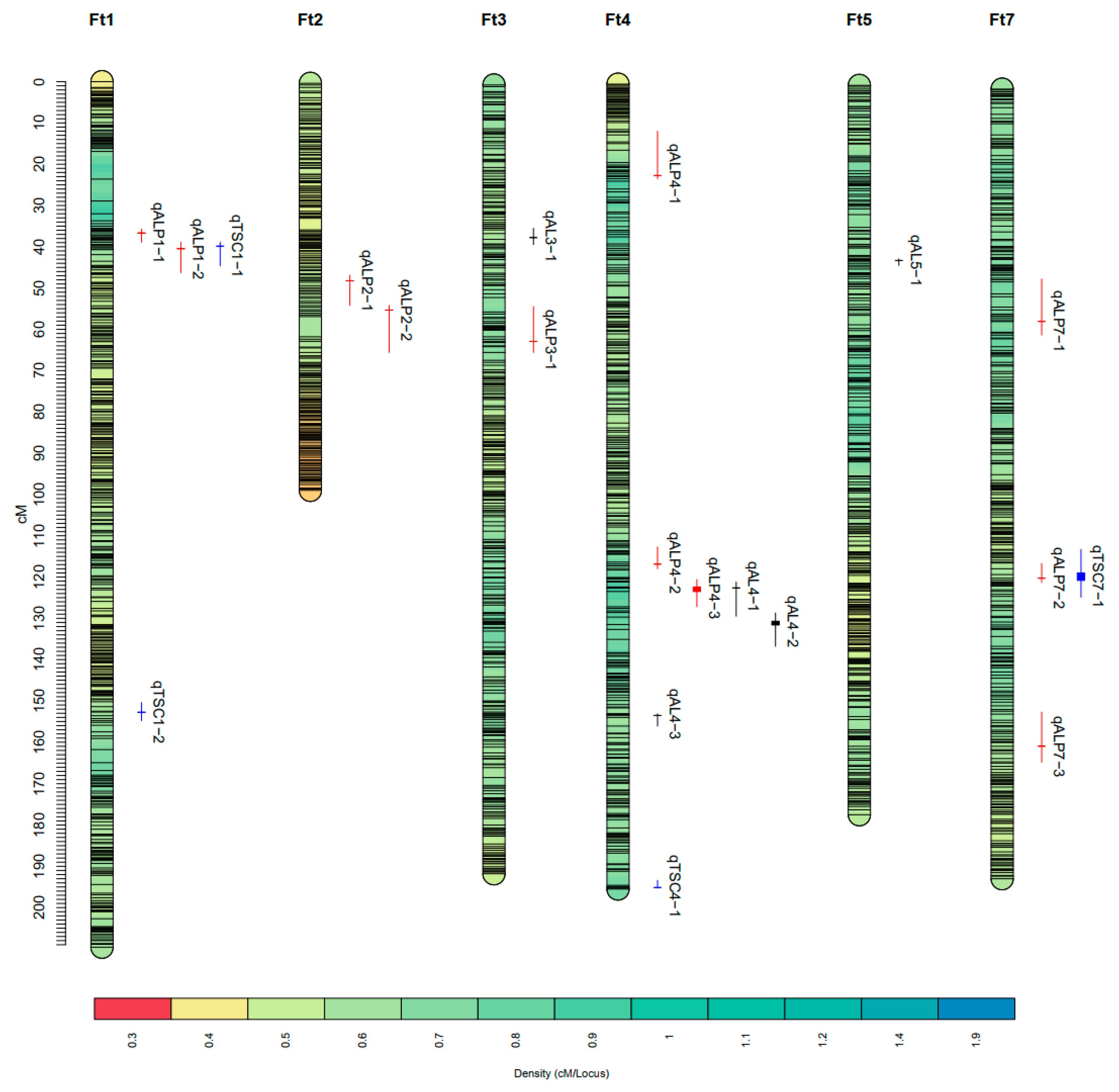
2.2. QTL Mapping of Starch-Related Traits
2.3. QTL Cluster Analysis of Starch-Related Traits
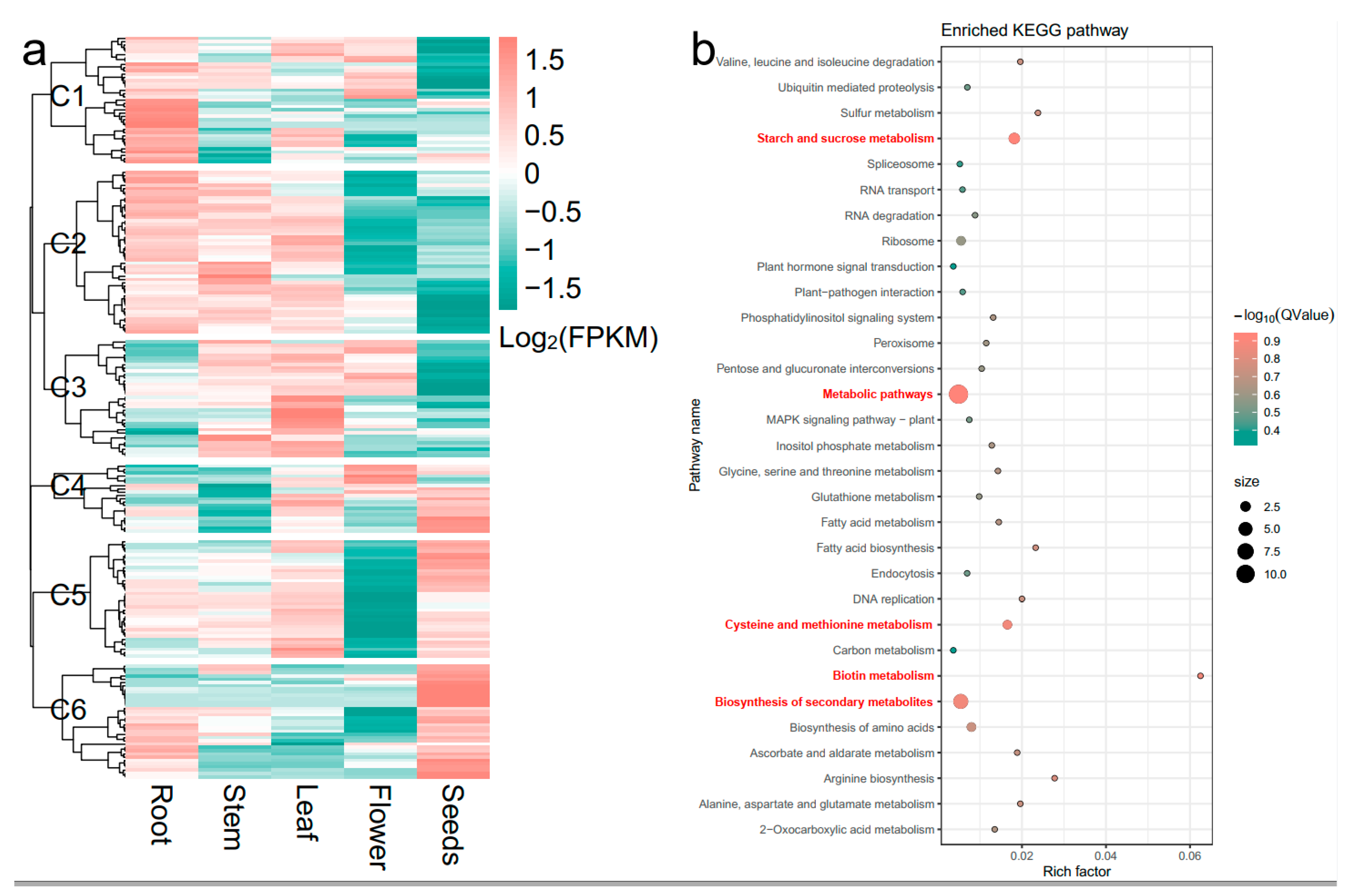
2.4. Identification of Candidate Genes within the Major QTL Cluster qClu-4-1
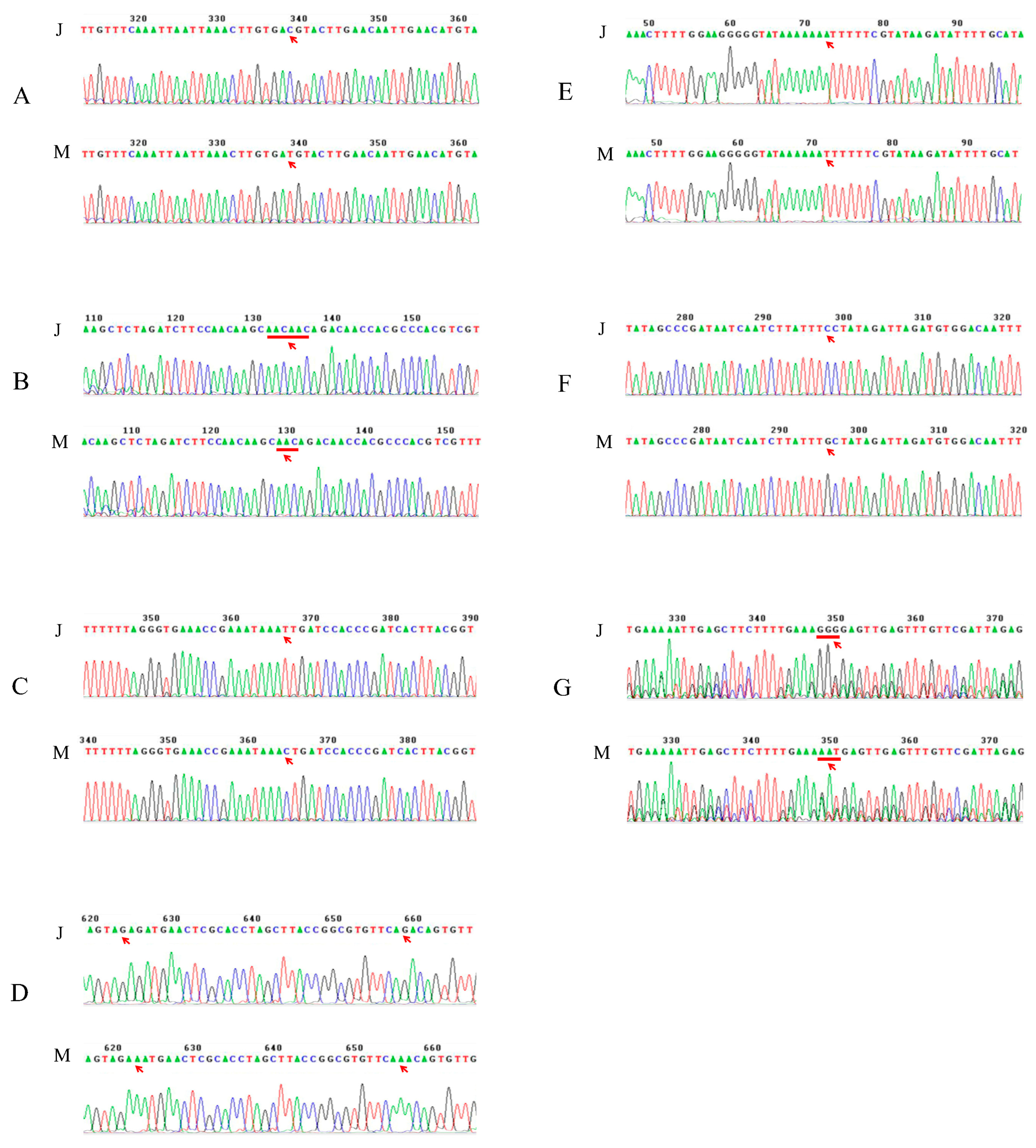
2.5. Analysis of the SNP/InDel Variations of Candidate Genes for qClu-4-1
2.6. Starch Content and Expression of Candidate Genes in Two Parents
3. Discussion
4. Materials and Methods
4.1. Materials Planting and Sampling
4.2. Measurement and Analysis of the Content of Starch-Related Traits
4.3. QTL Mapping
4.4. Candidate Gene Prediction and Analyses
4.5. Verification of the SNP/InDel Variations of Candidate Genes
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Ohnishi, O. Search for the wild ancestor of buckwheat. I. Description of new Fagopyrum (Polygonaceae) species and their distribution in China and Himalayan hills. Fagopyru 1998, 15, 18–28. [Google Scholar]
- Luthar, Z.; Golob, A.; Germ, M.; Vombergar, B.; Kreft, I. Tartary Buckwheat in Human Nutrition. Plants 2021, 10, 700. [Google Scholar] [CrossRef] [PubMed]
- Li, S.Q.; Zhang, Q.H. Advances in the development of functional foods from buckwheat. Crit. Rev. Food Sci. Nutr. 2001, 41, 451–464. [Google Scholar] [CrossRef]
- Koval, D.; Plocková, M.; Kyselka, J.; Skřivan, P.; Sluková, M.; Horáčková, Š. Buckwheat Secondary Metabolites: Potential Antifungal Agents. J. Agric. Food Chem. 2020, 68, 11631–11643. [Google Scholar] [CrossRef] [PubMed]
- Pinski, A.; Zhou, M.; Betekhtin, A. Editorial: Advances in buckwheat research. Front. Plant Sci. 2023, 14, 1190090. [Google Scholar] [CrossRef]
- Zhu, F. Chemical composition and health effects of Tartary buckwheat. Food Chem. 2016, 203, 231–245. [Google Scholar] [CrossRef]
- Zou, L.; Wu, D.; Ren, G.; Hu, Y.; Peng, L.; Zhao, J.; Garcia-Perez, P.; Carpena, M.; Prieto, M.A.; Cao, H.; et al. Bioactive compounds, health benefits, and industrial applications of Tartary buckwheat (Fagopyrum tataricum). Crit. Rev. Food Sci. Nutr. 2023, 63, 657–673. [Google Scholar] [CrossRef]
- Salimi, M.; Channab, B.E.; El Idrissi, A.; Zahouily, M.; Motamedi, E. A comprehensive review on starch: Structure, modification, and applications in slow/controlled-release fertilizers in agriculture. Carbohydr. Polym. 2023, 322, 121326. [Google Scholar] [CrossRef]
- Seung, D. Amylose in starch: Towards an understanding of biosynthesis, structure and function. New Phytol. 2020, 228, 1490–1504. [Google Scholar] [CrossRef]
- Seung, D.; Smith, A.M. Starch granule initiation and morphogenesis-progress in Arabidopsis and cereals. J. Exp. Bot. 2019, 70, 771–784. [Google Scholar] [CrossRef]
- Huang, L.; Tan, H.; Zhang, C.; Li, Q.; Liu, Q. Starch biosynthesis in cereal endosperms: An updated review over the last decade. Plant Commun. 2021, 2, 100237. [Google Scholar] [CrossRef] [PubMed]
- Kharshiing, G.; Chrungoo, N.K. Wx alleles in rice: Relationship with apparent amylose content of starch and a possible role in rice domestication. J. Genet. 2021, 100, 65. [Google Scholar] [CrossRef] [PubMed]
- Crofts, N.; Sugimoto, K.; Oitome, N.F.; Nakamura, Y.; Fujita, N. Differences in specificity and compensatory functions among three major starch synthases determine the structure of amylopectin in rice endosperm. Plant Mol. Biol. 2017, 94, 399–417. [Google Scholar] [CrossRef] [PubMed]
- Utsumi, Y.; Utsumi, C.; Tanaka, M.; Takahashi, S.; Okamoto, Y.; Ono, M.; Nakamura, Y.; Seki, M. Suppressed expression of starch branching enzyme 1 and 2 increases resistant starch and amylose content and modifies amylopectin structure in cassava. Plant Mol. Biol. 2022, 108, 413–427. [Google Scholar] [CrossRef]
- Jin, S.K.; Xu, L.N.; Leng, Y.J.; Zhang, M.Q.; Yang, Q.Q.; Wang, S.L.; Jia, S.W.; Song, T.; Wang, R.A.; Tao, T.; et al. The OsNAC24-OsNAP protein complex activates OsGBSSI and OsSBEI expression to fine-tune starch biosynthesis in rice endosperm. Plant Biotechnol. J. 2023, 21, 2224–2240. [Google Scholar] [CrossRef]
- Cao, R.; Zhao, S.; Jiao, G.; Duan, Y.; Ma, L.; Dong, N.; Lu, F.; Zhu, M.; Shao, G.; Hu, S.; et al. OPAQUE3, encoding a transmembrane bZIP transcription factor, regulates endosperm storage protein and starch biosynthesis in rice. Plant Commun. 2022, 3, 100463. [Google Scholar] [CrossRef]
- Guo, Y.; Wang, G.; Guo, X.; Chi, S.; Yu, H.; Jin, K.; Huang, H.; Wang, D.; Wu, C.; Tian, J.; et al. Genetic dissection of protein and starch during wheat grain development using QTL mapping and GWAS. Front. Plant Sci. 2023, 14, 1189887. [Google Scholar] [CrossRef]
- Guo, Y.; Mu, P.; Liu, J.; Lu, Y.; Li, Z. QTL mapping and QxE interactions of grain cooking and nutrient qualities in rice under upland and lowland environments. J. Genet. Genom. = Yi Chuan Xue Bao 2007, 34, 420–428. [Google Scholar] [CrossRef]
- Hu, S.; Wang, M.; Zhang, X.; Chen, W.; Song, X.; Fu, X.; Fang, H.; Xu, J.; Xiao, Y.; Li, Y.; et al. Genetic basis of kernel starch content decoded in a maize multi-parent population. Plant Biotechnol. J. 2021, 19, 2192–2205. [Google Scholar] [CrossRef]
- Zhu, F. Buckwheat starch: Structures, properties, and applications. Trends Food Sci. Technol. 2016, 49, 121–135. [Google Scholar] [CrossRef]
- Wang, X.; Feng, B.; Xu, Z.; Sestili, F.; Zhao, G.; Xiang, C.; Lafiandra, D.; Wang, T. Identification and characterization of granule bound starch synthase I (GBSSI) gene of tartary buckwheat (Fagopyrum tataricum Gaertn.). Gene 2014, 534, 229–235. [Google Scholar] [CrossRef] [PubMed]
- Huang, J.; Deng, J.; Shi, T.; Chen, Q.; Liang, C.; Meng, Z.; Zhu, L.; Wang, Y.; Zhao, F.; Yu, S.; et al. Global transcriptome analysis and identification of genes involved in nutrients accumulation during seed development of rice tartary buckwheat (Fagopyrum tararicum). Sci. Rep. 2017, 7, 11792. [Google Scholar] [CrossRef] [PubMed]
- Huang, J.; Tang, B.; Ren, R.; Wu, M.; Liu, F.; Lv, Y.; Shi, T.; Deng, J.; Chen, Q. Understanding the Potential Gene Regulatory Network of Starch Biosynthesis in Tartary Buckwheat by RNA-Seq. Int. J. Mol. Sci. 2022, 23, 15774. [Google Scholar] [CrossRef] [PubMed]
- Wang, L.; Liu, L.; Wu, H.; Li, C.; Zhao, H.; Wu, Q. Evolutionary and expression analysis of starch synthase genes from Tartary buckwheat revealed the potential function of FtGBSSII-4 and FtGBSSII-5 in seed amylose biosynthesis. Crop Sci. 2023, 63, 2925–2940. [Google Scholar] [CrossRef]
- Shi, T.-X.; Li, R.-Y.; Zheng, R.; Chen, Q.-F.; Li, H.-Y.; Huang, J.; Zhu, L.-W.; Liang, C.-G. Mapping QTLs for 1000-grain weight and genes controlling hull type using SNP marker in Tartary buckwheat (Fagopyrum tataricum). BMC Genom. 2021, 22, 142. [Google Scholar] [CrossRef]
- Zhang, L.; Li, X.; Ma, B.; Gao, Q.; Du, H.; Han, Y.; Li, Y.; Cao, Y.; Qi, M.; Zhu, Y.; et al. The Tartary Buckwheat Genome Provides Insights into Rutin Biosynthesis and Abiotic Stress Tolerance. Mol. Plant 2017, 10, 1224–1237. [Google Scholar] [CrossRef]
- Zhu, A.; Zhang, Y.; Zhang, Z.; Wang, B.; Xue, P.; Cao, Y.; Chen, Y.; Li, Z.; Liu, Q.; Cheng, S.; et al. Genetic Dissection of qPCG1 for a Quantitative Trait Locus for Percentage of Chalky Grain in Rice (Oryza sativa L.). Front. Plant Sci. 2018, 9, 1173. [Google Scholar] [CrossRef]
- Liu, N.; Zhang, Z.; Xue, Y.; Meng, S.; Huang, Y.; Li, W.; Huang, J.; Tang, J. Identification of Quantitative Trait Loci and Candidate Genes for Maize Starch Granule Size through Association Mapping. Sci. Rep. 2018, 8, 14236. [Google Scholar] [CrossRef] [PubMed]
- Rhazi, L.; Méléard, B.; Daaloul, O.; Grignon, G.; Branlard, G.; Aussenac, T. Genetic and Environmental Variation in Starch Content, Starch Granule Distribution and Starch Polymer Molecular Characteristics of French Bread Wheat. Foods 2021, 10, 205. [Google Scholar] [CrossRef]
- Yin, Y.; Yu, C.; Yu, L.; Zhao, J.; Sun, C.; Ma, Y.; Zhou, G. The influence of light intensity and photoperiod on duckweed biomass and starch accumulation for bioethanol production. Bioresour. Technol. 2015, 187, 84–90. [Google Scholar] [CrossRef]
- Li, R.; Chen, Z.; Zheng, R.; Chen, Q.; Deng, J.; Li, H.; Huang, J.; Liang, C.; Shi, T. QTL mapping and candidate gene analysis for yield and grain weight/size in Tartary buckwheat. BMC Plant Biol. 2023, 23, 58. [Google Scholar] [CrossRef] [PubMed]
- Chen, K.; Łyskowski, A.; Jaremko, Ł.; Jaremko, M. Genetic and Molecular Factors Determining Grain Weight in Rice. Front. Plant Sci. 2021, 12, 605799. [Google Scholar] [CrossRef] [PubMed]
- Li, G.; Wang, D.; Yang, R.; Logan, K.; Chen, H.; Zhang, S.; Skaggs, M.I.; Lloyd, A.; Burnett, W.J.; Laurie, J.D.; et al. Temporal patterns of gene expression in developing maize endosperm identified through transcriptome sequencing. Proc. Natl. Acad. Sci. USA 2014, 111, 7582–7587. [Google Scholar] [CrossRef]
- Cai, H.; Chen, Y.; Zhang, M.; Cai, R.; Cheng, B.; Ma, Q.; Zhao, Y. A novel GRAS transcription factor, ZmGRAS20, regulates starch biosynthesis in rice endosperm. Physiol. Mol. Biol. Plants Int. J. Funct. Plant Biol. 2017, 23, 143–154. [Google Scholar] [CrossRef]
- Wang, L.; Ni, D.; Yang, F.; Lin, L.; Yang, Y.; Sun, C.; Ye, X.; Cao, J.; Kong, X. Transcriptome profiling of Sorghum bicolor reveals cultivar-specific molecular signatures associated with starch and phenolic compounds biosyntheses and accumulation during sorghum grain development. Czech J. Genet. Plant Breed. 2023, 59, 235–252. [Google Scholar] [CrossRef]
- Abt, M.R.; Zeeman, S.C. Evolutionary innovations in starch metabolism. Curr. Opin. Plant Biol. 2020, 55, 109–117. [Google Scholar] [CrossRef]
- Abt, M.R.; Pfister, B.; Sharma, M.; Eicke, S.; Bürgy, L.; Neale, I.; Seung, D.; Zeeman, S.C. STARCH SYNTHASE5, a Noncanonical Starch Synthase-Like Protein, Promotes Starch Granule Initiation in Arabidopsis. Plant Cell 2020, 32, 2543–2565. [Google Scholar] [CrossRef] [PubMed]
- Tetlow, I.J.; Wait, R.; Lu, Z.; Akkasaeng, R.; Bowsher, C.G.; Esposito, S.; Kosar-Hashemi, B.; Morell, M.K.; Emes, M.J. Protein phosphorylation in amyloplasts regulates starch branching enzyme activity and protein-protein interactions. Plant Cell 2004, 16, 694–708. [Google Scholar] [CrossRef] [PubMed]
- Grimaud, F.; Rogniaux, H.; James, M.G.; Myers, A.M.; Planchot, V. Proteome and phosphoproteome analysis of starch granule-associated proteins from normal maize and mutants affected in starch biosynthesis. J. Exp. Bot. 2008, 59, 3395–3406. [Google Scholar] [CrossRef]
- Guo, S.; Chai, S.; Guo, Y.; Shi, X.; Han, F.; Qu, T.; Xing, L.; Yang, Q.; Gao, J.; Gao, X.; et al. Mapping of major QTL and candidate gene analysis for hull colour in foxtail millet (Setaria italica (L.) P. Beauv.). BMC Genom. 2023, 24, 458. [Google Scholar] [CrossRef]
- Bu, D.; Luo, H.; Huo, P.; Wang, Z.; Zhang, S.; He, Z.; Wu, Y.; Zhao, L.; Liu, J.; Guo, J.; et al. KOBAS-i: Intelligent prioritization and exploratory visualization of biological functions for gene enrichment analysis. Nucleic Acids Res. 2021, 49, W317–W325. [Google Scholar] [CrossRef] [PubMed]
- Li, L.; Chang, H.; Zhao, S.; Liu, R.; Yan, M.; Li, F.; El-Sheery, N.I.; Feng, Z.; Yu, S. Combining high-throughput deep learning phenotyping and GWAS to reveal genetic variants of fruit branch angle in upland cotton. Ind. Crops Prod. 2024, 220, 119180. [Google Scholar] [CrossRef]
- Zhou, Z.; Chen, Y.; Yan, M.; Zhao, S.; Li, F.; Yu, S.; Feng, Z.; Li, L. Genome-wide identification and mining elite allele variation of the Monoacylglycerol lipase (MAGL) gene family in upland cotton (Gossypium hirsutum L.). BMC Plant Biol. 2024, 24, 587. [Google Scholar] [CrossRef] [PubMed]



{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Traits | Environments | Parents | XJ-RILs Population | |||||
|---|---|---|---|---|---|---|---|---|
| Jinqiaomai2 | Xiaomiqiao | Mean ± SD | Range | Kurtosis | Skewness | CV (%) | ||
| Amylose (%) | 2021S | 20.90 ± 0.24 b | 22.28 ± 0.91 a | 19.26 ± 1.98 | 11.19~23.45 | 0.375 | −0.528 | 10% |
| 2021A | 17.80 ± 0.33 b | 19.44 ± 0.61 a | 19.76 ± 1.20 | 15.70~22.50 | 0.449 | 0.031 | 6% | |
| 2022S | 24.97 ± 0.64 a | 26.19 ± 0.61 b | 22.39 ± 2.15 | 17.20~28.02 | −0.238 | 0.004 | 10% | |
| Amylopectin (%) | 2021S | 47.03 ± 1.22 b | 50.95 ± 1.87 a | 49.83 ± 5.20 | 36.80~59.68 | −0.442 | −0.310 | 10% |
| 2021A | 41.65 ± 1.69 a | 43.61 ± 1.24 b | 32.09 ± 2.86 | 23.66~39.93 | 0.634 | 0.632 | 9% | |
| 2022S | 42.98 ± 1.62 a | 43.57 ± 1.08 a | 40.46 ± 4.32 | 30.26~51.61 | 0.013 | 0.208 | 11% | |
| Total Starch (%) | 2021S | 67.92 ± 1.17 b | 73.24 ± 1.85 a | 69.08 ± 4.94 | 57.73~78.72 | −0.492 | −0.178 | 7% |
| 2021A | 59.45 ± 1.50 a | 63.05 ± 1.62 b | 51.84 ± 2.54 | 45.61~61.52 | 2.419 | 0.970 | 5% | |
| 2022S | 67.96 ± 1.49 a | 69.76 ± 1.38 b | 62.85 ± 4.46 | 53.07~74.15 | −0.201 | 0.476 | 7% |
| Trait | Environment | Total Starch | Amylopectin |
|---|---|---|---|
| Amylose | 2021S | 0.065 | −0.315 ** |
| 2021A | −0.016 | −0.425 ** | |
| 2022S | 0.307 ** | −0.180 * | |
| Amylopectin | 2021S | 0.927 ** | |
| 2021A | 0.912 ** | ||
| 2022S | 0.881 ** |
| NO. | Trait | QTL | Environment | Chromosome | Position (cM) | LOD | R2 (%) | Additive Effect | Confidence Interval (cM) | Flanking Markers | Physical Interval (Mbp) |
|---|---|---|---|---|---|---|---|---|---|---|---|
| 1 | Amylose | qAL3-1 | 2021S | Ft3 | 37.51 | 2.68 | 5.2 | −0.51 | 35.2~39.2 | Block5303~Block5318 | 11.8~12.55 |
| 2 | Amylose | qAL4-1 | 2021A | Ft4 | 122.31 | 3.31 | 6.46 | 0.55 | 120.9~129.2 | Block8744~Block8836 | 39.85~42.74 |
| BLUP | Ft4 | 122.31 | 5.82 | 9.52 | 0.002 | ||||||
| 3 | Amylose | qAL4-2 | 2021S | Ft4 | 130.41 | 4.8 | 10.12 | 0.65 | 128.4~136.5 | Block8830~Block8844 | 42.10~43.34 |
| 2022S | Ft4 | 131.31 | 2.58 | 5.08 | 0.49 | ||||||
| BLUP | Ft4 | 131.31 | 4.42 | 7.34 | 0.002 | ||||||
| 4 | Amylose | qAL4-3 | 2021A | Ft4 | 153.21 | 2.7 | 6.22 | −0.34 | 152.8~155.8 | Block9021~Block9046 | 46.58~48.92 |
| 5 | Amylose | qAL5-1 | 2022S | Ft5 | 43.31 | 4.74 | 11.04 | −0.49 | 42.9~44.5 | Block9728~Block9753 | 8.39~9.06 |
| 6 | Amylopectin | qALP1-1 | BLUP | Ft1 | 36.61 | 5.31 | 8.09 | −0.17 | 35.7~38.9 | Block320~Block337 | 6.47~6.10 |
| 7 | Amylopectin | qALP1-2 | 2022S | Ft1 | 40.51 | 3.54 | 6.33 | −1.13 | 38.9~46.3 | Block334~Block428 | 6.45~9.15 |
| 8 | Amylopectin | qALP2-1 | 2021A | Ft2 | 48.21 | 4.09 | 9.04 | 2.15 | 46.9~54.2 | Block2872~Block2992 | 15.34~21.46 |
| 9 | Amylopectin | qALP2-2 | 2022S | Ft2 | 55.31 | 3.47 | 7.73 | 1.96 | 54.2~65.6 | Block3073~Block3468 | 12.57~31.04 |
| 10 | Amylopectin | qALP3-1 | 2021S | Ft3 | 62.61 | 3.37 | 7.22 | −0.79 | 54.2~65.3 | Block5579~Block5697 | 16.07~26.81 |
| 11 | Amylopectin | qALP4-1 | 2021S | Ft4 | 22.41 | 2.97 | 5.52 | −1.24 | 11.7~23.2 | Block7563~Block7612 | 3.01~5.65 |
| 12 | Amylopectin | qALP4-2 | 2021A | Ft4 | 116.61 | 3.29 | 6.76 | −1.77 | 112.4~117.7 | Block8396~Block8758 | 34.64~40.42 |
| 13 | Amylopectin | qALP4-3 | 2021S | Ft4 | 122.11 | 4.72 | 8.59 | −1.28 | 120.3~126.9 | Block8752~Block8836 | 39.85~42.65 |
| 2022S | Ft4 | 123.21 | 7.37 | 14.41 | −2.28 | ||||||
| BLUP | Ft4 | 123.21 | 6.44 | 10.16 | −0.19 | ||||||
| 14 | Amylopectin | qALP7-1 | 2021A | Ft7 | 58.11 | 2.62 | 5.53 | 0.68 | 47.8~61.4 | Block13692~Block13766 | 12.1~15.32 |
| 15 | Amylopectin | qALP7-2 | 2021S | Ft7 | 120.31 | 3.61 | 6.8 | 1.37 | 116.7~121.3 | Block14329~Block14351 | 37.07~38.89 |
| 16 | Amylopectin | qALP7-3 | BLUP | Ft7 | 161.01 | 2.74 | 4.07 | 0.12 | 152.7~164.8 | Block14552~Block14603 | 44.53~46.04 |
| 17 | Total Starch | qTSC1-1 | 2022S | Ft1 | 39.81 | 2.91 | 5.76 | −1.11 | 38.9~44.6 | Block334~Block412 | 6.45~8.72 |
| 18 | Total Starch | qTSC1-2 | BLUP | Ft1 | 152.71 | 4.33 | 7.45 | 0.04 | 150.4~154.8 | Block709~Block714 | 44.85~45.09 |
| 19 | Total Starch | qTSC4-1 | 2021S | Ft4 | 194.81 | 3.01 | 5.98 | −1.12 | 193.1~195 | Block9392~Block9381 | 55.13~56.65 |
| 20 | Total Starch | qTSC7-1 | BLUP | Ft7 | 119.01 | 2.54 | 4.41 | 0.03 | 113.3~124.9 | Block14276~Block14405 | 35.85~40.63 |
| 2021A | Ft7 | 120.81 | 3.9 | 7.99 | 1.42 |
| NO. | Cluster Name | Chromosome | Confidence Interval (cM) | Flanking Markers | Physical Interval (Mbp) | QTL Number | QTL |
|---|---|---|---|---|---|---|---|
| 1 | qClu-1-1 | Ft1 | 35.7~46.3 | Block320~Block428 | 6.45~9.15 | 3 | qALP1-1, qTSC1-1, qALP1-2 |
| 2 | qClu-4-1 | Ft4 | 120.3~136.5 | Block8744~Block8844 | 39.85~43.34 | 3 | qALP4-3*, qAL4-1#, qAL4-2* |
| 3 | qClu-7-1 | Ft7 | 113.3~124.9 | Block14276~Block14405 | 35.85~40.63 | 2 | qTSC7-1#, qALP7-2 |
| NO. | Name | Chr | Orientation | Times of Mutation | Mutation Type | Expression Cluster | Mutation Region | Rice Annotation |
|---|---|---|---|---|---|---|---|---|
| 1 | FtPinG0004897000.01 | Ft4 | − | 14 | SNP/Indel | C5 | 5’ UTR/3’ UTR | LOC_Os12g05430|ribosomal protein L24, putative, expressed |
| 2 | FtPinG0004897100.01 | Ft4 | + | 6 | SNP/Indel | C6 | 5’ UTR/3’ UTR | LOC_Os07g39820|SHR, putative, expressed |
| 3 | FtPinG0004899000.01 | Ft4 | − | 1 | SNP | C6 | 5’ UTR | LOC_Os03g31690|GCN5-related N-acetyltransferase, putative, expressed |
| 4 | FtPinG0002636200.01 | Ft4 | + | 1 | SNP | C5 | 3’ UTR | LOC_Os02g49140|glycosyltransferase, putative, expressed |
| 5 | FtPinG0002635900.01 | Ft4 | − | 1 | Indel | C5 | 3’ UTR | LOC_Os03g51990|ACT domain-containing protein, expressed |
| 6 | FtPinG0008593800.01 | Ft4 | + | 2 | SNP | C5 | exon/3’ UTR | LOC_Os07g41570|expressed protein |
| 7 | FtPinG0008592400.01 | Ft4 | + | 6 | SNP/Indel | C5 | 5’ UTR/3’ UTR | LOC_Os03g07570|aminotransferase, putative, expressed |
| 8 | FtPinG0007101700.01 | Ft4 | + | 1 | Indel | C6 | 3’ UTR | LOC_Os08g33210|expressed protein |
| 9 | FtPinG0005926000.01 | Ft4 | + | 4 | SNP/Indel | C4 | 3’ UTR | LOC_Os03g60750|ribosomal RNA large subunit methyltransferase J, putative, expressed |
| 10 | FtPinG0005925700.01 | Ft4 | − | 2 | Indel | C5 | 3’ UTR | LOC_Os01g74146|WD repeat-containing protein, putative, expressed |
| 11 | FtPinG0009329200.01 | Ft4 | + | 1 | SNP | C5 | intron | LOC_Os03g17740|transporter, putative, expressed |
| 12 | FtPinG0006225500.01 | Ft4 | − | 1 | SNP | C6 | 3’ UTR | LOC_Os03g60720|expansin precursor, putative, expressed |
| 13 | FtPinG0008469200.01 | Ft4 | + | 1 | SNP | C5 | 5’ UTR | LOC_Os03g52320|GRF-interacting factor 1, putative, expressed |
| 14 | FtPinG0008468900.01 | Ft4 | − | 1 | SNP | C5 | intron | LOC_Os03g52320|GRF-interacting factor 1, putative, expressed |
| 15 | FtPinG0007371600.01 | Ft4 | + | 1 | SNP | C5 | 3’ UTR | LOC_Os03g49510|phosphatidylinositol-4-phosphate 5-kinase, putative, expressed |
| 16 | FtPinG0007371400.01 | Ft4 | + | 13 | SNP | C6 | 5’ UTR/3’ UTR | LOC_Os09g28110|hydroxyproline-rich glycoprotein family protein, putative, expressed |
| 17 | FtPinG0006002900.01 | Ft4 | + | 1 | SNP | C4 | 5’ UTR | LOC_Os12g03260|MATE efflux family protein, putative, expressed |
| 18 | FtPinG0005124100.01 | Ft4 | − | 1 | SNP | C5 | 5’ UTR | LOC_Os03g28940|ZIM domain-containing protein, putative, expressed |
| 19 | FtPinG0005109900.01 | Ft4 | − | 1 | SNP | C5 | 3’ UTR | LOC_Os02g51070|starch synthase, putative, expressed |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Huang, J.; Liu, F.; Ren, R.; Deng, J.; Zhu, L.; Li, H.; Cai, F.; Meng, Z.; Chen, Q.; Shi, T. QTL Mapping and Candidate Gene Analysis for Starch-Related Traits in Tartary Buckwheat (Fagopyrum tataricum (L.) Gaertn). Int. J. Mol. Sci. 2024, 25, 9243. https://doi.org/10.3390/ijms25179243
Huang J, Liu F, Ren R, Deng J, Zhu L, Li H, Cai F, Meng Z, Chen Q, Shi T. QTL Mapping and Candidate Gene Analysis for Starch-Related Traits in Tartary Buckwheat (Fagopyrum tataricum (L.) Gaertn). International Journal of Molecular Sciences. 2024; 25(17):9243. https://doi.org/10.3390/ijms25179243
Chicago/Turabian StyleHuang, Juan, Fei Liu, Rongrong Ren, Jiao Deng, Liwei Zhu, Hongyou Li, Fang Cai, Ziye Meng, Qingfu Chen, and Taoxiong Shi. 2024. "QTL Mapping and Candidate Gene Analysis for Starch-Related Traits in Tartary Buckwheat (Fagopyrum tataricum (L.) Gaertn)" International Journal of Molecular Sciences 25, no. 17: 9243. https://doi.org/10.3390/ijms25179243
APA StyleHuang, J., Liu, F., Ren, R., Deng, J., Zhu, L., Li, H., Cai, F., Meng, Z., Chen, Q., & Shi, T. (2024). QTL Mapping and Candidate Gene Analysis for Starch-Related Traits in Tartary Buckwheat (Fagopyrum tataricum (L.) Gaertn). International Journal of Molecular Sciences, 25(17), 9243. https://doi.org/10.3390/ijms25179243