
Polar-Region Soils as Novel Reservoir of Lactic Acid Bacteria from the Genus Carnobacterium
 , , and
, , and
Abstract
:1. Introduction
2. Results
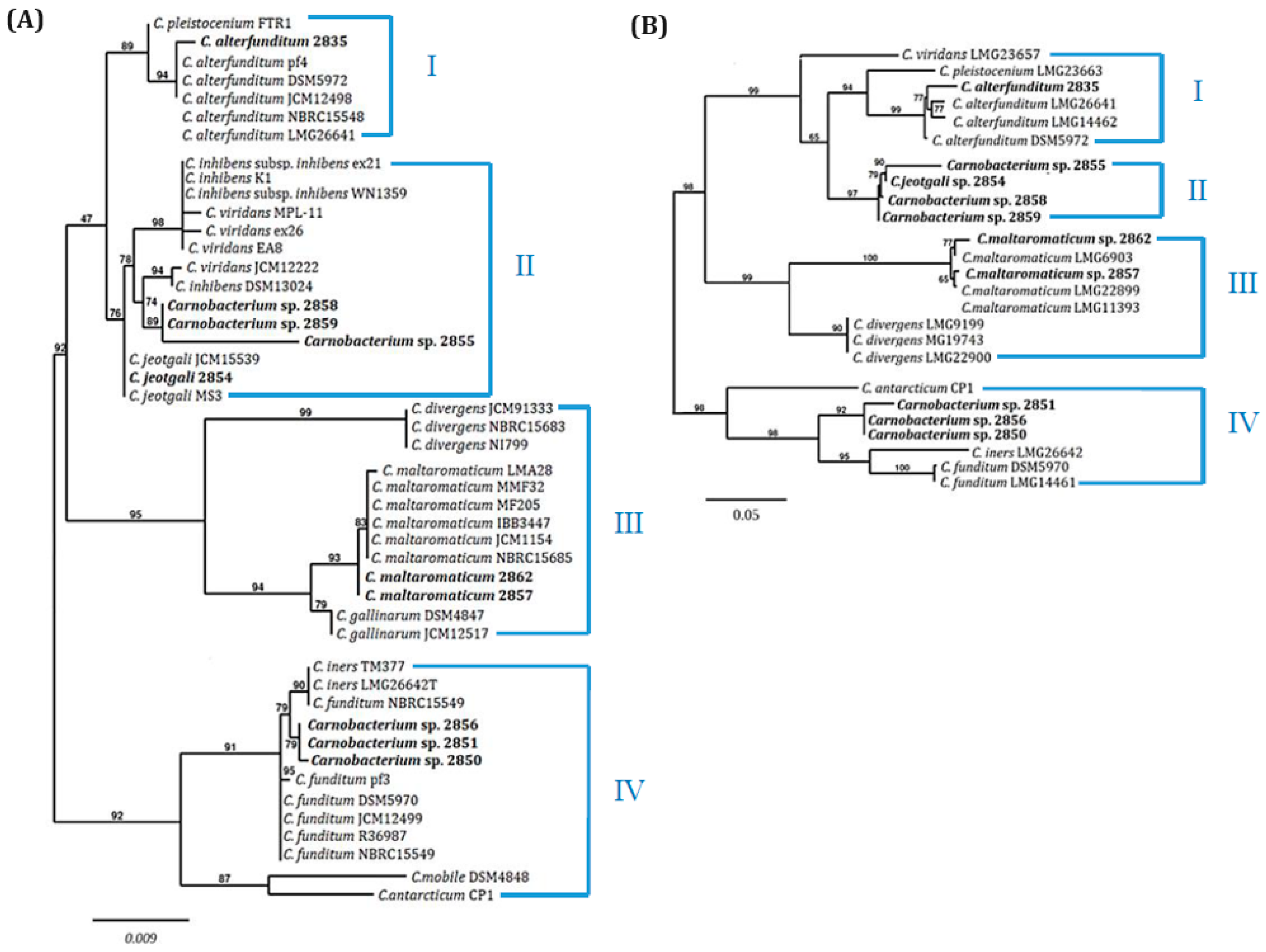
2.1. Taxonomy of Carnobacterium spp. Isolates
2.2. Physiological Characteristics
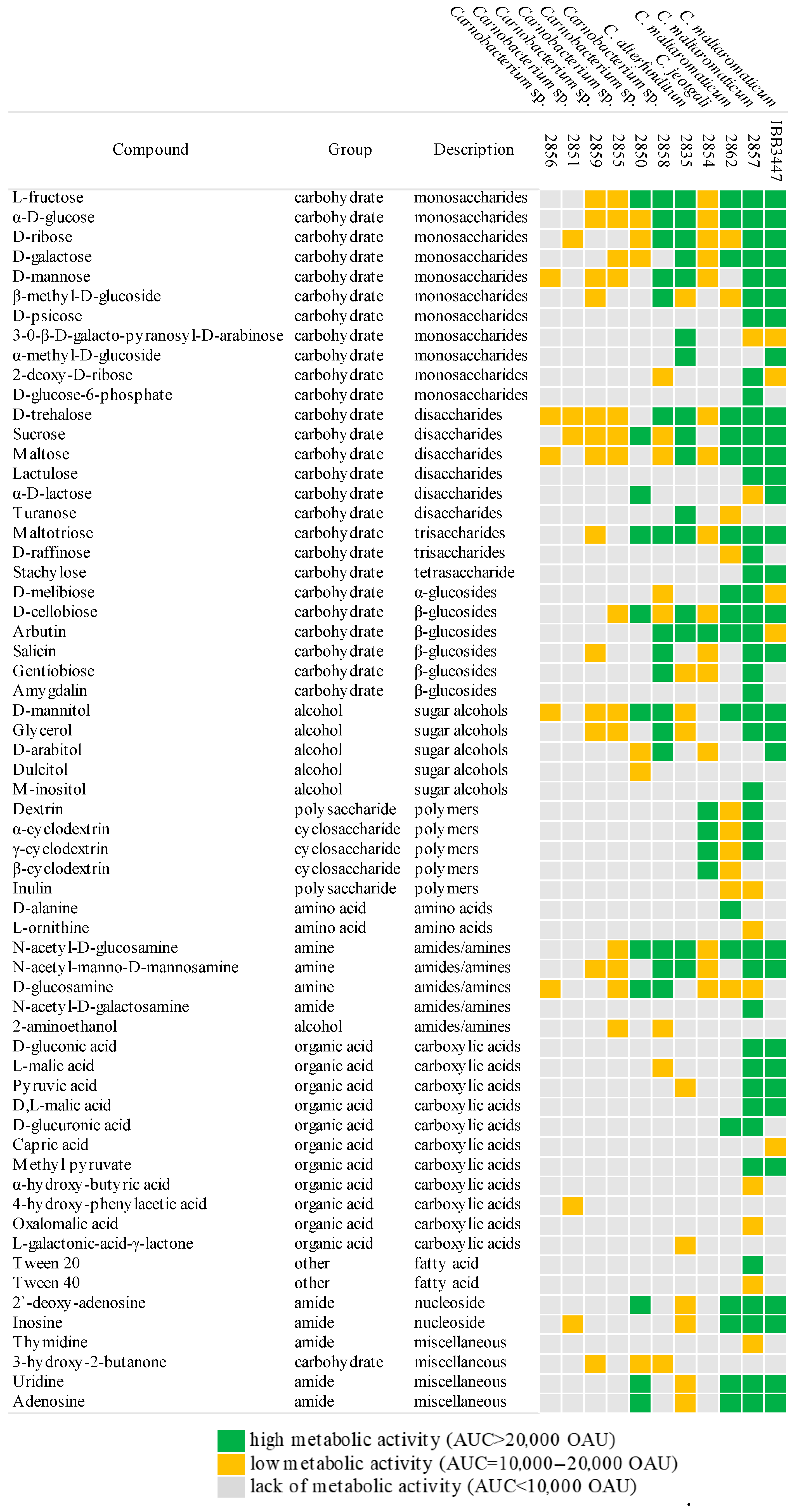
2.3. Carbon Assimilation
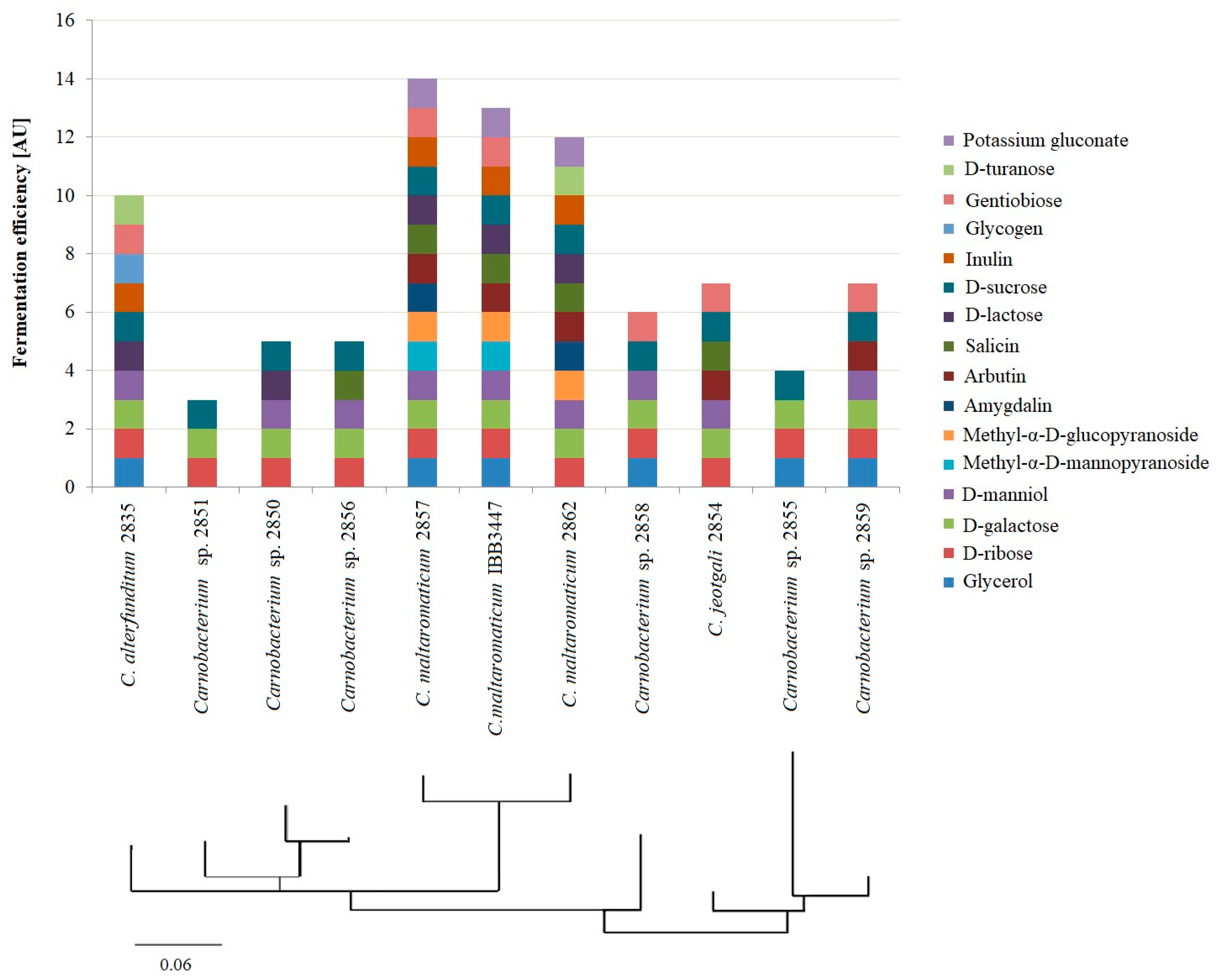
2.4. Lactic Acid Production
2.5. Antimicrobial Activity
2.6. Plasmid Characterization
3. Discussion
4. Materials and Methods
4.1. Isolation of Carnobacterium spp. Strains
4.2. Genetic Analyses
4.3. Plasmids Isolation and Sequencing
4.4. Metabolic Analyses
4.5. Antimicrobial Activity and Bacteriocin Purification
4.6. Statistical Analysis
5. Conclusions
- Identification of metabolic properties of isolated Carnobacterium strains that may suggest an ecological role involving various syntrophic interactions with other microorganisms in polar habitats, such as algae, mosses, and lichens, through the secretion and fermentation of carbohydrates and alcohols.
- Presentation of methodology and data providing guidelines for the isolation of psychrophilic LAB from environmental resources of polar regions; identification of new Carnobacterium species; and characterization of metabolic traits and antimicrobial properties that determine the adaptive potential of LAB strains to permanently cold habitats, as well as for biotechnological application.
- Description of important metabolic features of industrial importance in cold-adapted Carnobacterium strains, such as the production of fermented foods, probiotics, and antimicrobial compounds, that could be more efficiently harnessed under conditions that are less demanding than those required by thermophilic LAB strains.
- Detailed characterization of carnobacterial plasmids and identification of certain genetic elements that may be useful in targeted engineering of suitable Carnobacteriaceae strains.
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Teneva-Angelova, T.; Hristova, I.; Pavlov, A.; Beshkova, D. Lactic Acid Bacteria—From Nature Through Food to Health. In Advances in Biotechnology for Food Industry; Academic Press: Cambridge, MA, USA, 2018. [Google Scholar]
- Saha, U.B.; Saroj, S.D. Lactic Acid Bacteria: Prominent Player in the Fight against Human Pathogens. Expert Rev. Anti. Infect. Ther. 2022, 20, 1435–1453. [Google Scholar] [CrossRef] [PubMed]
- Zapaśnik, A.; Sokołowska, B.; Bryła, M. Role of Lactic Acid Bacteria in Food Preservation and Safety. Foods 2022, 11, 1283. [Google Scholar] [CrossRef] [PubMed]
- Franzmann, P.D.; Höpfl, P.; Weiss, N.; Tindall, B.J. Psychrotrophic, Lactic Acid-Producing Bacteria from Anoxic Waters in Ace Lake, Antarctica; Carnobacterium Funditum Sp. Nov. and Carnobacterium Alterfunditum Sp. Nov. Arch. Microbiol. 1991, 156, 255–262. [Google Scholar] [CrossRef]
- Pikuta, E.; Marsic, D.; Bej, A.; Tang, J.; Krader, P.; Hoover, R. Carnobacterium Pleistocenium Sp Nov., a Novel Psychrotolerant, Facultative Anaerobe Isolated from Permafrost of the Fox Tunnel in Alaska. Int. J. Syst. Evol. Microbiol. 2005, 55, 473–478. [Google Scholar] [CrossRef]
- Zhu, S.; Lin, D.; Shangling, X.; Wang, X.; Xue, Z.; Dong, B.; Shen, X.; Ma, X.; Chen, J.; Yang, J. Carnobacterium Antarcticum Sp. Nov., a Psychrotolerant, Alkaliphilic Bacterium Isolated from Sandy Soil in Antarctica. Int. J. Syst. Evol. Microbiol. 2018, 68, 1672–1677. [Google Scholar] [CrossRef] [PubMed]
- Nicholson, W.L.; Zhalnina, K.; de Oliveira, R.R.; Triplett, E.W. Proposal to Rename Carnobacterium Inhibens as Carnobacterium Inhibens Subsp. Inhibens Subsp. Nov. and Description of Carnobacterium Inhibens Subsp. Gilichinskyi Subsp. Nov., a Psychrotolerant Bacterium Isolated from Siberian Permafrost. Int. J. Syst. Evol. Microbiol. 2015, 65, 556–561. [Google Scholar] [CrossRef]
- Thomas, D.N.; Fogg, G.E.; Convey, P.; Fritsen, C.H.; Gili, J.-M.; Gradinger, R.; Laybourn-Parry, J.; Reid, K.; Walton, D.W.H. The Biology of Polar Regions; Oxford University Press: Oxford, UK, 2008; ISBN 9780199298112. [Google Scholar]
- Klekowski, R.Z.; Opaliński, K.W. Matter and Energy Flow in Spitsbergen Ornithogenic Tundra. Polar Res. 1986, 4, 187–197. [Google Scholar] [CrossRef]
- Myrcha, A.; Jadwiszczak, P.; Tambussi, C.; Noriega, J.; Gaździcki, A.; Tatur, A.; del Valle, R. Taxonomic Revision of Eocene Antarctic Penguins Based on Tarsometatarsal Morphology. Polar Res. 2002, 23, 5–46. [Google Scholar]
- Turner, J.; Overland, J. Contrasting Climate Change in the Two Polar Regions. Polar Res. 2009, 28, 146–164. [Google Scholar] [CrossRef]
- Romaniuk, K.; Ciok, A.; Decewicz, P.; Uhrynowski, W.; Budzik, K.; Nieckarz, M.; Pawlowska, J.; Zdanowski, M.K.; Bartosik, D.; Dziewit, L. Insight into Heavy Metal Resistome of Soil Psychrotolerant Bacteria Originating from King George Island (Antarctica). Polar Biol. 2018, 41, 1319–1333. [Google Scholar] [CrossRef]
- Zdanowski, M.K.; Żmuda-Baranowska, M.J.; Borsuk, P.; Świątecki, A.; Górniak, D.; Wolicka, D.; Jankowska, K.M.; Grzesiak, J. Culturable Bacteria Community Development in Postglacial Soils of Ecology Glacier, King George Island, Antarctica. Polar Biol. 2013, 36, 511–527. [Google Scholar] [CrossRef]
- Grzesiak, J.; Górniak, D.; Świątecki, A.; Aleksandrzak-Piekarczyk, T.; Szatraj, K.; Zdanowski, M.K. Microbial Community Development on the Surface of Hans and Werenskiold Glaciers (Svalbard, Arctic): A Comparison. Extremophiles 2015, 19, 885–897. [Google Scholar] [CrossRef]
- Grzesiak, J.; Zdanowski, M.K.; Górniak, D.; Świątecki, A.; Aleksandrzak-Piekarczyk, T.; Szatraj, K.; Sasin-Kurowska, J.; Nieckarz, M. Microbial Community Changes along the Ecology Glacier Ablation Zone (King George Island, Antarctica). Polar Biol. 2015, 38, 2069–2083. [Google Scholar] [CrossRef]
- Zdanowski, M.K.; Bogdanowicz, A.; Gawor, J.; Gromadka, R.; Wolicka, D.; Grzesiak, J. Enrichment of Cryoconite Hole Anaerobes: Implications for the Subglacial Microbiome. Microb. Ecol. 2017, 73, 532–538. [Google Scholar] [CrossRef] [PubMed]
- Cukier, S.; Fudala, K.; Bialik, R.J. Are Antarctic Aquatic Invertebrates Hitchhiking on Your Footwear? J. Nat. Conserv. 2023, 72, 126354. [Google Scholar] [CrossRef]
- Górniak, D.; Marszałek, H.; Kwaśniak-Kominek, M.; Rzepa, G.; Manecki, M. Soil Formation and Initial Microbiological Activity on a Foreland of an Arctic Glacier (SW Svalbard). Appl. Soil Ecol. 2017, 114, 34–44. [Google Scholar] [CrossRef]
- Duda, M.P.; Hargan, K.E.; Michelutti, N.; Smol, J.P. Freshwater Diatom Assemblages from Seabird-Inhabited Ponds in Hudson Strait, Sub-Arctic Canada. Polar Biol. 2019, 42, 1549–1560. [Google Scholar] [CrossRef]
- Teixeira, L.C.R.S.; Peixoto, R.S.; Cury, J.C.; Sul, W.J.; Pellizari, V.H.; Tiedje, J.; Rosado, A.S. Bacterial Diversity in Rhizosphere Soil from Antarctic Vascular Plants of Admiralty Bay, Maritime Antarctica. ISME J. 2010, 4, 989–1001. [Google Scholar] [CrossRef]
- Schmidt, S.K.; Johnson, B.W.; Solon, A.J.; Sommers, P.; Darcy, J.L.; Vincent, K.; Vimercati, L.; Fountain, A.G.; Porazinska, D.L. Microbial Biogeochemistry and Phosphorus Limitation in Cryoconite Holes on Glaciers across the Taylor Valley, McMurdo Dry Valleys, Antarctica. Biogeochemistry 2022, 158, 313–326. [Google Scholar] [CrossRef]
- Jin, S.; Wang, Y.; Zhao, X. Cold-Adaptive Mechanism of Psychrophilic Bacteria in Food and Its Application. Microb. Pathog. 2022, 169, 105652. [Google Scholar] [CrossRef]
- Znój, A.; Gawor, J.; Gromadka, R.; Chwedorzewska, K.J.; Grzesiak, J. Root-Associated Bacteria Community Characteristics of Antarctic Plants: Deschampsia Antarctica and Colobanthus Quitensis—A Comparison. Microb. Ecol. 2021, 84, 808–820. [Google Scholar] [CrossRef] [PubMed]
- Dziurzynski, M.; Gorecki, A.; Pawlowska, J.; Istel, L.; Decewicz, P.; Golec, P.; Styczynski, M.; Poszytek, K.; Rokowska, A.; Gorniak, D.; et al. Revealing the Diversity of Bacteria and Fungi in the Active Layer of Permafrost at Spitsbergen Island (Arctic)-Combining Classical Microbiology and Metabarcoding for Ecological and Bioprospecting Exploration. Sci. Total Environ. 2023, 856, 159072. [Google Scholar] [CrossRef] [PubMed]
- Abakumov, E.; Abramov, A.; Aislabie, J.; Alekseeva, T.; Balks, M.; Blume, H.-P.; Bockheim, J.; Bölter, M.; Dergacheva, M.; Dolgikh, A.; et al. The Soils of Antarctica; Springer: Berlin/Heidelberg, Germany, 2015; ISBN 978-3-319-05497-1. [Google Scholar]
- Białkowska, A.M.; Grzelczyk, A.; Długołęcka, A.; Cieśliński, H.; Kalinowska, H.; Kur, J.; Turkiewicz, M. Physicochemical and Biological Characterization of Soils from the Vicinity of the Arctowski Polish Antarctic Station. Biotechnol. Food Sci. 2012, 76, 13–26. [Google Scholar] [CrossRef]
- Szymański, W.; Siwek, J.; Waścińska, J.; Wojtuń, B. Texture and Geochemistry of Surface Horizons of Arctic Soils from a Non-Glaciated Catchment, SW Spitsbergen. Pol. Polar Res. 2016, 37, 361–377. [Google Scholar] [CrossRef]
- Whon, T.W.; Hyun, D.-W.; Nam, Y.-D.; Kim, M.-S.; Song, E.-J.; Jang, Y.K.; Jung, E.S.; Shin, N.-R.; Oh, S.J.; Kim, P.S.; et al. Genomic and Phenotypic Analyses of Carnobacterium Jeotgali Strain MS3T, a Lactate-Producing Candidate Biopreservative Bacterium Isolated from Salt-Fermented Shrimp. FEMS Microbiol. Lett. 2015, 362, fnv058. [Google Scholar] [CrossRef]
- Mora, D.; Scarpellini, M.; Franzetti, L.; Colombo, S.; Galli, A. Reclassification of Lactobacillus Maltaromicus (Miller et Al. 1974) DSM 20342T and DSM 20344 and Carnobacterium Piscicola (Collins et Al. 1987) DSM 20730T and DSM 20722 as Carnobacterium Maltaromaticum Comb. Nov. Int. J. Syst. Evol. Microbiol. 2003, 53, 675–678. [Google Scholar] [CrossRef] [PubMed]
- Afzal, M.I.; Ariceaga, C.C.G.; Lhomme, E.; Ali, N.K.; Payot, S.; Burgain, J.; Gaiani, C.; Borges, F.; Revol-Junelles, A.-M.; Delaunay, S.; et al. Characterization of Carnobacterium Maltaromaticum LMA 28 for Its Positive Technological Role in Soft Cheese Making. Food Microbiol. 2013, 36, 223–230. [Google Scholar] [CrossRef]
- Buda, J.; Łokas, E.; Pietryka, M.; Richter, D.; Magowski, W.; Iakovenko, N.S.; Porazinska, D.L.; Budzik, T.; Grabiec, M.; Grzesiak, J.; et al. Biotope and Biocenosis of Cryoconite Hole Ecosystems on Ecology Glacier in the Maritime Antarctic. Sci. Total Environ. 2020, 724, 138112. [Google Scholar] [CrossRef]
- Pikuta, E.V.; Hoover, R.B. The Genus Carnobacterium. In Lactic Acid Bacteria; John Wiley & Sons, Ltd.: Hoboken, NJ, USA, 2014; pp. 109–123. ISBN 9781118655252. [Google Scholar]
- Tang, T.; Martinenghi, L.D.; Hounmanou, Y.M.G.; Leisner, J.J. Distribution and Ecology of the Generalist Lactic Acid Bacterium Carnobacterium Maltaromaticum in Different Freshwater Habitats: Metabolic and Antagonistic Abilities. Environ. Microbiol. 2023, 25, 3556–3576. [Google Scholar] [CrossRef]
- Yadav, A.N.; Verma, P.; Sachan, S.G.; Kaushik, R.; Saxena, A.K. Psychrotrophic Microbiomes: Molecular Diversity and Beneficial Role in Plant Growth Promotion and Soil Health. In Microorganisms for Green Revolution: Volume 2: Microbes for Sustainable Agro-ecosystem; Panpatte, D.G., Jhala, Y.K., Shelat, H.N., Vyas, R.V., Eds.; Springer: Singapore, 2018; pp. 197–240. ISBN 978-981-10-7146-1. [Google Scholar]
- Gunjal, A.; Badodekar, N. Halophiles; IGI Global: Hershey, PA, USA, 2021; pp. 13–34. ISBN 9781799891444. [Google Scholar]
- Papadopoulou, E.; Rodriguez de Evgrafov, M.C.; Kalea, A.; Tsapekos, P.; Angelidaki, I. Adaptive Laboratory Evolution to Hypersaline Conditions of Lactic Acid Bacteria Isolated from Seaweed. N. Biotechnol. 2023, 75, 21–30. [Google Scholar] [CrossRef]
- Magalhães, C.; Stevens, M.I.; Cary, S.C.; Ball, B.A.; Storey, B.C.; Wall, D.H.; Türk, R.; Ruprecht, U. At Limits of Life: Multidisciplinary Insights Reveal Environmental Constraints on Biotic Diversity in Continental Antarctica. PLoS ONE 2012, 7, e44578. [Google Scholar] [CrossRef]
- Wohl, C.; Jones, A.E.; Sturges, W.T.; Nightingale, P.D.; Else, B.; Butterworth, B.J.; Yang, M. Sea Ice Concentration Impacts Dissolved Organic Gases in the Canadian Arctic. Biogeosciences 2022, 19, 1021–1045. [Google Scholar] [CrossRef]
- Cowan, D.A.; Tow, L.A. Endangered Antarctic Environments. Annu. Rev. Microbiol. 2004, 58, 649–690. [Google Scholar] [CrossRef] [PubMed]
- Dimitrellou, D.; Salamoura, C.; Kontogianni, A.; Katsipi, D.; Kandylis, P.; Zakynthinos, G.; Varzakas, T. Effect of Milk Type on the Microbiological, Physicochemical and Sensory Characteristics of Probiotic Fermented Milk. Microorganisms 2019, 7, 274. [Google Scholar] [CrossRef] [PubMed]
- Cabrerizo, A.; Dachs, J.; Barceló, D.; Jones, K.C. Influence of Organic Matter Content and Human Activities on the Occurrence of Organic Pollutants in Antarctic Soils, Lichens, Grass, and Mosses. Environ. Sci. Technol. 2012, 46, 1396–1405. [Google Scholar] [CrossRef]
- Underwood, G.J.C.; Aslam, S.N.; Michel, C.; Niemi, A.; Norman, L.; Meiners, K.M.; Laybourn-Parry, J.; Paterson, H.; Thomas, D.N. Broad-Scale Predictability of Carbohydrates and Exopolymers in Antarctic and Arctic Sea Ice. Proc. Natl. Acad. Sci. USA 2013, 110, 15734–15739. [Google Scholar] [CrossRef]
- Roh, H.; Kim, B.S.; Lee, M.K.; Park, C.-I.; Kim, D.-H. Genome-Wide Comparison of Carnobacterium Maltaromaticum Derived from Diseased Fish Harbouring Important Virulence-Related Genes. J. Fish Dis. 2020, 43, 1029–1037. [Google Scholar] [CrossRef]
- van Belkum, M.J.; Stiles, M.E. Characterization of the Theta-Type Plasmid PCD3.4 from Carnobacterium Divergens, and Modulation of Its Host Range by RepA Mutation. Microbiology 2006, 152, 171–178. [Google Scholar] [CrossRef]
- Heinl, S.; Wibberg, D.; Eikmeyer, F.; Szczepanowski, R.; Blom, J.; Linke, B.; Goesmann, A.; Grabherr, R.; Schwab, H.; Pühler, A.; et al. Insights into the Completely Annotated Genome of Lactobacillus Buchneri CD034, a Strain Isolated from Stable Grass Silage. J. Biotechnol. 2012, 161, 153–166. [Google Scholar] [CrossRef]
- Górecki, R.K.; Koryszewska-Bagińska, A.; Gołębiewski, M.; Żylińska, J.; Grynberg, M.; Bardowski, J.K. Adaptative Potential of the Lactococcus Lactis IL594 Strain Encoded in Its 7 Plasmids. PLoS ONE 2011, 6, e22238. [Google Scholar] [CrossRef]
- Casanueva, A.; Tuffin, M.; Cary, C.; Cowan, D.A. Molecular Adaptations to Psychrophily: The Impact of ‘Omic’ Technologies. Trends Microbiol. 2010, 18, 374–381. [Google Scholar] [CrossRef] [PubMed]
- Kosiorek, K.; Koryszewska-Bagińska, A.; Skoneczny, M.; Stasiak-Różańska, L.; Aleksandrzak-Piekarczyk, T. The Presence of Plasmids in Lactococcus Lactis IL594 Determines Changes in the Host Phenotype and Expression of Chromosomal Genes. Int. J. Mol. Sci. 2023, 24, 793. [Google Scholar] [CrossRef]
- Cowan, D.; Khan, N.; Heath, C.; Mutondo, M. Polar Microbiology, The Ecology, Biodiversity and Bioremediation Potential of Microorganisms in Extremely Cold Environments; CRC Press: Boca Raton, FL, USA, 2010; ISBN 9781420083842. [Google Scholar]
- Cole, G.B.; Bateman, T.J.; Moraes, T.F. The Surface Lipoproteins of Gram-Negative Bacteria: Protectors and Foragers in Harsh Environments. J. Biol. Chem. 2021, 296, 100147. [Google Scholar] [CrossRef] [PubMed]
- Deming, J.W.; Young, J.N. The Role of Exopolysaccharides in Microbial Adaptation to Cold Habitats. In Psychrophiles: From Biodiversity to Biotechnology; Margesin, R., Ed.; Springer International Publishing: Cham, Switzerland, 2017; pp. 259–284. ISBN 978-3-319-57057-0. [Google Scholar]
- Leisner, J.J.; Hansen, M.A.; Larsen, M.H.; Hansen, L.; Ingmer, H.; Sørensen, S.J. The Genome Sequence of the Lactic Acid Bacterium, Carnobacterium Maltaromaticum ATCC 35586 Encodes Potential Virulence Factors. Int. J. Food Microbiol. 2012, 152, 107–115. [Google Scholar] [CrossRef] [PubMed]
- Greene, N.P.; Kaplan, E.; Crow, A.; Koronakis, V. Antibiotic Resistance Mediated by the MacB ABC Transporter Family: A Structural and Functional Perspective. Front. Microbiol. 2018, 9, 950. [Google Scholar] [CrossRef]
- Bakthavatchalam, Y.D.; Ramaswamy, B.; Janakiraman, R.; Steve, R.J.; Veeraraghavan, B. Genomic Insights of Community-Acquired Methicillin-Resistant Staphylococcus aureus (MRSA) with Reduced Teicoplanin Susceptibility: A Case of Fatal Necrotizing Fasciitis. J. Glob. Antimicrob. Resist. 2018, 14, 242–245. [Google Scholar] [CrossRef]
- Scott, L.C.; Lee, N.; Aw, T.G. Antibiotic Resistance in Minimally Human-Impacted Environments. Int. J. Environ. Res. Public Health 2020, 17, 3939. [Google Scholar] [CrossRef]
- Aleksandrzak-Piekarczyk, T.; Kok, J.; Renault, P.; Bardowski, J. Alternative Lactose Catabolic Pathway in Lactococcus Lactis IL1403. Appl. Environ. Microbiol. 2005, 71, 6060–6069. [Google Scholar] [CrossRef]
- Aleksandrzak-Piekarczyk, T.; Szatraj, K.; Kosiorek, K. GlaR (YugA)-a Novel RpiR-Family Transcription Activator of the Leloir Pathway of Galactose Utilization in Lactococcus Lactis IL1403. Microbiologyopen 2019, 8, e00714. [Google Scholar] [CrossRef]
- Martin-Visscher, L.A.; van Belkum, M.J.; Garneau-Tsodikova, S.; Whittal, R.M.; Zheng, J.; McMullen, L.M.; Vederas, J.C. Isolation and Characterization of Carnocyclin a, a Novel Circular Bacteriocin Produced by Carnobacterium Maltaromaticum UAL307. Appl. Environ. Microbiol. 2008, 74, 4756–4763. [Google Scholar] [CrossRef]
- Acedo, J.Z.; Towle, K.M.; Lohans, C.T.; Miskolzie, M.; McKay, R.T.; Doerksen, T.A.; Vederas, J.C.; Martin-Visscher, L.A. Identification and Three-Dimensional Structure of Carnobacteriocin XY, a Class IIb Bacteriocin Produced by Carnobacteria. FEBS Lett. 2017, 591, 1349–1359. [Google Scholar] [CrossRef] [PubMed]
- Begrem, S.; Ivaniuk, F.; Gigout-Chevalier, F.; Kolypczuk, L.; Bonnetot, S.; Leroi, F.; Grovel, O.; Delbarre-Ladrat, C.; Passerini, D. New Insight into Antimicrobial Compounds from Food and Marine-Sourced Carnobacterium Species through Phenotype and Genome Analyses. Microorganisms 2020, 8, 1093. [Google Scholar] [CrossRef]
- Stupar, J.; Holøymoen, I.G.; Hoel, S.; Lerfall, J.; Rustad, T.; Jakobsen, A.N. Diversity and Antimicrobial Activity towards Listeria Spp. and Escherichia Coli among Lactic Acid Bacteria Isolated from Ready-to-Eat Seafood. Foods 2021, 10, 271. [Google Scholar] [CrossRef]
- Lohans, C.T.; van Belkum, M.J.; Li, J.; Vederas, J.C. Characterization of Bacterial Antimicrobial Peptides Active against Campylobacter Jejuni. Can. J. Chem. 2015, 93, 381–388. [Google Scholar] [CrossRef]
- Hwanhlem, N.; Buradaleng, S.; Wattanachant, S.; Benjakul, S.; Tani, A.; Maneerat, S. Isolation and Screening of Lactic Acid Bacteria from Thai Traditional Fermented Fish (Plasom) and Production of Plasom from Selected Strains. Food Control 2011, 22, 401–407. [Google Scholar] [CrossRef]
- Abedi, E.; Hashemi, S.M.B. Lactic Acid Production–Producing Microorganisms and Substrates Sources-State of Art. Heliyon 2020, 6, e04974. [Google Scholar] [CrossRef] [PubMed]
- Cunha, B.L.C.; Bahú, J.O.; Xavier, L.F.; Crivellin, S.; de Souza, S.D.A.; Lodi, L.; Jardini, A.L.; Filho, R.M.; Schiavon, M.I.R.B.; Concha, V.O.C.; et al. Lactide: Production Routes, Properties, and Applications. Bioengineering 2022, 9, 164. [Google Scholar] [CrossRef]
- Swetha, S.; Padmavathi, T. Study of Acid Phosphatase in Solubilization of Inorganic Phosphates by Piriformospora Indica. Pol. J. Microbiol. 2016, 65, 407–412. [Google Scholar] [CrossRef]
- Zlotnikov, K.; Zlotnikov, A.K.; Kaparullina, E.; Doronina, N. Phylogenetic Position and Phosphate Solubilization Activity of Lactic Acid Bacteria Associated with Different Plants. Mikrobiologiia 2013, 82, 376–379. [Google Scholar] [CrossRef]
- Yang, G.; Tian, J.; Chen, J. Editorial: Soil Microbes in Polar Region: Response, Adaptation and Mitigation of Climate Change. Front. Microbiol. 2022, 13, 1086822. [Google Scholar] [CrossRef]
- Hoover, R.B.; Pikuta, E.V.; Bej, A.K.; Marsic, D.; Whitman, W.B.; Tang, J.; Krader, P. Spirochaeta Americana Sp. Nov., a New Haloalkaliphilic, Obligately Anaerobic Spirochaete Isolated from Soda Mono Lake in California. Int. J. Syst. Evol. Microbiol. 2003, 53, 815–821. [Google Scholar] [CrossRef]
- Naser, S.M.; Thompson, F.L.; Hoste, B.; Gevers, D.; Dawyndt, P.; Vancanneyt, M.; Swings, J. Application of Multilocus Sequence Analysis (MLSA) for Rapid Identification of Enterococcus Species Based on RpoA and PheS Genes. Microbiology 2005, 151 Pt 7, 2141–2150. [Google Scholar] [CrossRef]
- Sayers, E.W.; Beck, J.; Bolton, E.E.; Bourexis, D.; Brister, J.R.; Canese, K.; Comeau, D.C.; Funk, K.; Kim, S.; Klimke, W.; et al. Database Resources of the National Center for Biotechnology Information. Nucleic Acids Res. 2021, 49, D10–D17. [Google Scholar] [CrossRef] [PubMed]
- Madeira, F.; Pearce, M.; Tivey, A.R.N.; Basutkar, P.; Lee, J.; Edbali, O.; Madhusoodanan, N.; Kolesnikov, A.; Lopez, R. Search and Sequence Analysis Tools Services from EMBL-EBI in 2022. Nucleic Acids Res. 2022, 50, W276–W279. [Google Scholar] [CrossRef]
- Dereeper, A.; Guignon, V.; Blanc, G.; Audic, S.; Buffet, S.; Chevenet, F.; Dufayard, J.-F.; Guindon, S.; Lefort, V.; Lescot, M.; et al. Phylogeny.Fr: Robust Phylogenetic Analysis for the Non-Specialist. Nucleic Acids Res. 2008, 36, W465–W469. [Google Scholar] [CrossRef] [PubMed]
- Charnock, C.; Hagen, R.X.; Nguyen, T.N.-T.; Vo, L.T. Diversion and Phylogenetic Relatedness of Filterable Bacteria from Norwegian Tap and Bottled Waters. J. Water Health 2019, 17, 295–307. [Google Scholar] [CrossRef] [PubMed]
- Dec, M.; Urban-Chmiel, R.; Gnat, S.; Puchalski, A.; Wernicki, A. Identification of Lactobacillus Strains of Goose Origin Using MALDI-TOF Mass Spectrometry and 16S–23S RDNA Intergenic Spacer PCR Analysis. Res. Microbiol. 2014, 165, 190–201. [Google Scholar] [CrossRef] [PubMed]
- Wick, R.R.; Judd, L.M.; Gorrie, C.L.; Holt, K.E. Unicycler: Resolving Bacterial Genome Assemblies from Short and Long Sequencing Reads. PLoS Comput. Biol. 2017, 13, e1005595. [Google Scholar] [CrossRef]
- Tanizawa, Y.; Fujisawa, T.; Nakamura, Y. DFAST: A Flexible Prokaryotic Genome Annotation Pipeline for Faster Genome Publication. Bioinformatics 2017, 34, 1037–1039. [Google Scholar] [CrossRef]
- Zimmermann, L.; Stephens, A.; Nam, S.-Z.; Rau, D.; Kübler, J.; Lozajic, M.; Gabler, F.; Söding, J.; Lupas, A.N.; Alva, V. A Completely Reimplemented MPI Bioinformatics Toolkit with a New HHpred Server at its Core. J. Mol. Biol. 2018, 430, 2237–2243. [Google Scholar] [CrossRef]
- Jecz, P.; Bartosik, A.A.; Glabski, K.; Jagura-Burdzy, G. A Single ParS Sequence from the Cluster of Four Sites Closest to OriC Is Necessary and Sufficient for Proper Chromosome Segregation in Pseudomonas Aeruginosa. PLoS ONE 2015, 10, e0120867. [Google Scholar] [CrossRef] [PubMed]
- Camesasca, L.; de Mattos, J.A.; Vila, E.; Cebreiros, F.; Lareo, C. Lactic Acid Production by Carnobacterium Sp. Isolated from a Maritime Antarctic Lake Using Eucalyptus Enzymatic Hydrolysate. Biotechnol. Rep. 2021, 31, e00643. [Google Scholar] [CrossRef]
- Zhang, H.; Cui, Y.; Zhu, S.; Feng, F.; Zheng, X. Characterization and Antimicrobial Activity of a Pharmaceutical Microemulsion. Int. J. Pharm. 2010, 395, 154–160. [Google Scholar] [CrossRef] [PubMed]
- Tymoszewska, A.; Aleksandrzak-Piekarczyk, T. The Lactococcal DgkB (YecE) and DxsA Genes for Lipid Metabolism Are Involved in the Resistance to Cell Envelope-Acting Antimicrobials. Int. J. Mol. Sci. 2021, 22, 1014. [Google Scholar] [CrossRef] [PubMed]

{kind=link}
{kind=link}
{kind=link}
| Isolate | Source of Isolation | Longitude | Latitude | Alignment Result 1 | Query Coverage [%] | Sequence Identity [%] | Taxonomic Identification | Reference for Soil Components |
|---|---|---|---|---|---|---|---|---|
| 2835 | Cryoconite, Ecology Glacier, King George Island (Antarctica) | −58.47559 | −62.17331 | C. alterfunditum JCM12498 | 100 | 100 | C. alterfunditum | [15] |
| 2850 | Ornithogenic soil, Llano Cape, King George Island (Antarctica) | −58.44761 | −62.17491 | C. funditum NBRC15549 C. funditum JCM12499 C. iners TM377 | 100 100 100 | 100 99 99 | Carnobacterium sp. | [25] |
| 2851 | Ornithogenic soil, Arctowski Station terrain, King George Island (Antarctica) | −58.45869 | −62.16317 | C. funditum NBRC15549 C. funditum JCM12499 C. funditum pf3 C. iners TM377 | 100 99 99 99 | 100 99 99 99 | Carnobacterium sp. | [23] |
| 2854 | Soil, Baranowski Glacier foreland, King George Island (Antarctica) | −58.44339 | −62.20501 | C. jeotgali MS3 C. jeotgali JM15539 | 100 100 | 100 99 | C. jeotgali | Not available |
| 2855 | −58.44084 | −62.2047 | C. jeotgali MS3 C. inhibens WN1359 C. funditum NBRC15549 | 94 93 | 98 98 | Carnobacterium sp. | ||
| 2856 | Soil, seal elephants wake, King George Island (Antarctica) | −58.46284 | −62.16255 | C. funditum NBRC15549 C. funditum JCM12499 C. iners TM377 | 100 99 99 | 100 99 98 | Carnobacterium sp. | [26] |
| 2857 | Soil, Hans Glacier foreland, Spitsbergen (Arctic) | 15.597403 | 77.015244 | C. maltaromaticum JCM1154 | 100 | 100 | C. maltaromaticum | [27] |
| 2858 | Soil, Windy Glacier foreland, King George Island (Antarctica) | −58.47487 | −62.2309 | C. jeotgali MS3 C. inhibens WN1359 C. viridans JCM12222 | 100 100 99 | 100 100 99 | Carnobacterium sp. | Not available |
| 2859 | −58.47257 | −62.23316 | C. jeotgali JCM15539 C. inhibens WN1359 C. jeotgali MS3 | 100 100 100 | 100 100 99 | Carnobacterium sp. | ||
| 2862 | Freshwater microbial mats, Jasnorzewski Garden, King George Island (Antarctica) | −58.4683 | −62.15943 | C. maltaromaticum JCM 1154 | 100 | 99 | C. maltaromaticum | Not available |
| Feature | C. alterfunditum | C. jeotgali | C. funditum | Carnobacterium sp. | C. maltaromaticum | ||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| 2835 | DSM5972 | 2854 | MS3 1 | DSM5970 | 2850 | 2851 | 2855 | 2856 | 2858 | 2859 | 2857 | 2862 | IBB3447 | LMA 28 2 | |
| Motility | − | − | − | − | − | − | − | − | − | − | − | − | − | − | − |
| Growth: | |||||||||||||||
| 50% BHI | + | +/− | + | N/A | +/− | + | + | + | + | + | + | + | + | +/− | N/A |
| 100% BHI | +/− | − | +/− | N/A | − | +/− | +/− | +/− | + | +/− | +/− | + | + | + | N/A |
| 200% BHI | +/− | − | − | N/A | − | +/− | +/− | − | +/− | − | − | + | + | + | N/A |
| LB | +/− | +/− | +/− | N/A | +/− | +/− | +/− | +/− | +/− | +/− | +/− | +/− | +/− | +/− | N/A |
| GM17 | +/− | − | +/− | N/A | − | +/− | +/− | +/− | +/− | +/− | +/− | + | + | + | N/A |
| MRS | − | − | − | N/A | − | − | − | − | − | − | − | − | − | + | N/A |
| Temp range [°C] | 4–20 (16) | 4–12 (4) | 4–20 (16) | 4–37 (30) | 4–20 (16) | 4–20 (16) | 4–20 (16) | 4–16 (16) | 4–30 (16) | 4–20 (16) | 4–20 (16) | 4–37 (16) | 4–37 (16) | 16–37 (30) | 16–37 (30) |
| NaCl tolerance [%] | 2–10 (8) | 2–6 (3) | 2–9 (7) | 0–5 (2) | 2–10 (6) | 2–6 (4) | 2–6 (4) | 2–9 (4) | 2–8 (6) | 2–6 (4) | 2–8 (7) | 2–10 (8) | 2–8 (6) | 2–8 (3) | 2–8 (3) |
| pH range | 6–9 (8) | 6–8 (7) | 6–9 (7) | 5.5–9 (8.5) | 6–9 (7) | 6–9 (8) | 6–9 (8) | 6–9 (7) | 6–9 (7) | 6–9 (7) | 6–9 (7) | 5–9 (6) | 5–9 (6) | 3–9 (6) | 3–9 (6) |
| Produced acid from: | |||||||||||||||
| Glycerol | + | +/− | − | − | − | − | − | − | − | − | − | + | +/− | + | + |
| D-ribose | + | + | +/− | − | + | − | +/− | − | + | − | − | + | + | + | + |
| D-galactose | + | +/− | +/− | − | + | +/− | + | +/− | + | +/− | +/− | + | +/− | +/− | +/− |
| D-glucose | + | + | +/− | + | + | +/− | + | +/− | + | + | +/− | + | + | + | + |
| D-fructose | + | +/− | +/− | + | + | +/− | + | +/− | + | + | +/− | + | + | + | + |
| D-mannose | + | +/− | +/− | +/− | + | +/− | + | +/− | + | + | +/− | + | + | + | + |
| D-sucrose | +/− | +/− | +/− | +/− | + | +/− | +/− | +/− | + | +/− | +/− | + | + | + | + |
| D-mannitol | − | − | +/− | + | − | − | − | +/− | − | +/− | +/− | + | +/− | + | + |
| D-melibiose | − | − | − | − | − | − | − | − | − | − | − | + | +/− | +/− | − |
| D-cellobiose | + | − | +/− | − | − | + | − | +/− | − | +/− | − | + | + | + | + |
| Arbutin | − | N/D | +/− | +/− | − | − | − | − | − | +/− | +/− | + | + | + | + |
| Salicin | + | + | +/− | N/A | − | − | − | +/− | − | +/− | +/− | + | + | + | + |
| D-lactose | + | − | − | − | − | +/− | − | +/− | − | +/− | − | + | − | + | + |
| Amygdalin | − | +/− | +/− | − | − | − | − | − | − | +/− | +/− | + | + | + | + |
| Gentiobiose | +/− | N/D | − | − | − | − | − | − | − | +/− | +/− | + | +/− | + | + |
| Inulin | +/− | − | − | − | − | − | − | − | − | − | − | +/− | − | − | − |
| Starch | − | − | − | − | − | − | − | − | − | − | − | +/− | − | − | − |
| Glycogen | +/− | − | − | − | − | − | − | − | − | − | − | − | − | − | − |
| Esculin | − | − | + | + | + | + | + | + | + | + | + | + | + | + | + |
| Produced lactic acid [g/L] | 5.5 | 3.9 | 5.9 | N/A | 3.6 | 5.5 | 5.7 | 5.3 | 5.6 | 5.8 | 5.5 | 4.5 | 4.3 | 5.3 | NA |
| L-lactic acid [%] | 98 | 99 | 98 | N/A | 98 | 98 | 96 | 98 | 96 | 98 | 97 | 100 | 100 | 100 | NA |
| D-lactic acid [%] | 2 | 1 | 2 | N/A | 2 | 2 | 4 | 2 | 4 | 2 | 3 | 0 | 0 | 0 | NA |
| Strain | C. alterfunditum | C. funditum | Carnobacterium sp. | C. jeotgali | C. maltaromaticum | ||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| 2835 | DSM5972 | DSM5970 | 2850 | 2851 | 2856 | 2855 | 2858 | 2859 | 2854 | 2857 | 2862 | IBB3447 | |
| B. cereus IBB3390 | ○ | ○ | ○ | ○ | ○ | ○ | ○ | ○ | ○ | ○ | ○ | ○ | ○ |
| B. subtilis 168 | ○ | ○ | ○ | ○ | ○ | ○ | ○ | ○ | ○ | ○ | ○ | ○ | ● |
| C. jejuni 81176 | ○ | ○ | ○ | ○ | ○ | ○ | ● | ● | ○ | ● | ○ | ○ | ○ |
| C. albicans CAI-4 | ○ | ○ | ○ | ○ | ○ | ○ | ○ | ○ | ○ | ○ | ○ | ○ | ○ |
| L. monocytogenes LMGT2604 | ○ | ○ | ○ | ○ | ○ | ○ | ○ | ○ | ○ | ○ | ● | ○ | ● |
| P. aeruginosa ATCC9027 | ○ | ○ | ○ | ○ | ○ | ○ | ○ | ○ | ○ | ○ | ○ | ○ | ○ |
| S. typhimurium TT622 | ○ | ○ | ○ | ○ | ○ | ○ | ○ | ○ | ○ | ○ | ○ | ○ | ○ |
| S. aureus ATCC638 | ○ | ○ | ○ | ○ | ○ | ○ | ● | ● | ○ | ● | ○ | ○ | ○ |
| S. caprae DSM20608 | ○ | ○ | ○ | ○ | ○ | ○ | ● | ● | ○ | ● | ○ | ○ | ○ |
| S. delphini DSM20771 | ○ | ○ | ○ | ○ | ○ | ○ | ○ | ○ | ○ | ○ | ○ | ○ | ○ |
| S. epidermidis DSM20044 | ○ | ○ | ○ | ○ | ○ | ○ | ● | ● | ● | ● | ○ | ○ | ○ |
| S. hyicus DSM20454 | ○ | ○ | ○ | ○ | ○ | ○ | ● | ○ | ○ | ● | ○ | ○ | ● |
| S. intermedius DSM20373 | ○ | ○ | ○ | ○ | ○ | ○ | ● | ○ | ○ | ● | ○ | ○ | ○ |
| S. lugdunensis DSM4804 | ○ | ○ | ○ | ○ | ○ | ○ | ● | ● | ○ | ● | ○ | ○ | ○ |
| S. pseudintermedius DSM21284 | ○ | ○ | ○ | ○ | ○ | ○ | ○ | ○ | ○ | ○ | ○ | ○ | ○ |
| S. saprophyticus DSM20229 | ○ | ○ | ○ | ○ | ○ | ○ | ○ | ○ | ○ | ○ | ○ | ○ | ○ |
| S. schleiferi DSM6628 | ○ | ○ | ○ | ○ | ○ | ○ | ○ | ○ | ○ | ○ | ○ | ○ | ○ |
| S. agalactiae IBB130 | ○ | ○ | ○ | ○ | ○ | ○ | ○ | ○ | ○ | ○ | ○ | ○ | ○ |
| S.mitis IBB3449 | ○ | ○ | ○ | ○ | ○ | ○ | ○ | ○ | ○ | ○ | ○ | ○ | ○ |
| S. pneumoniae 5005 | ○ | ○ | ○ | ○ | ○ | ○ | ○ | ○ | ○ | ○ | ○ | ○ | ○ |
| S. agalactiae IBB123 | ○ | ○ | ○ | ○ | ○ | ○ | ○ | ○ | ○ | ○ | ○ | ○ | ○ |
| S. sobrinus IBB3450 | ○ | ○ | ○ | ○ | ○ | ○ | ○ | ○ | ○ | ○ | ○ | ○ | ○ |
| L. lactis 1403 | ○ | ○ | ○ | ○ | ○ | ○ | ○ | ○ | ○ | ○ | ○ | ○ | ○ |
| Species | Strain | Plasmid | GenBank Acc. No. | Size [bp] | GC Content [%] | Replication (Rep Protein Family) | Mobility | Maintenance | Genes of Adaptation Mechanisms | ||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Transfer and Mobilization Genes | Plasmid addiction Genes | Stability Genes | Carbohydrate Metabolism Genes | Peptide Metabolism Genes | DNA Repair Genes | Cell Resistance Genes | |||||||
| C. alterfunditum | 2835 | 2835_p1 | OR786483 | 37,450 | 30.3 | RepB (HTH_11) | none | parD parE | parA | celB pdaC | ardC, oppA pepV | kdp operon macB, tcaB | |
| 2835_p2 | OR786484 | 4480 | 33.7 | RepB (Rep_3) | none | none | none | ||||||
| 2835_p3 | OR786485 | 7305 | 29.1 | RepB (Rep_3) Rep (HTH_11) | none | none | none | lacX | |||||
| Carnobacterium spp. | 2851 | 2851_p1 | OR786482 | 3360 | 28.9 | RepB (Rep_trans) | none | none | none | galM | uvrX | ||
| 2851_p2 | OR786486 | 45,450 | 28.4 | RepA (HTH) | mobP, traG ardC | parE | parA | nlpC | |||||
| 2851_p3 | OR786487 | 37,450 | 30.3 | RepB (Rep_3) | mobA | none | parA | pdaC | umuD | kdp operon | |||
| 2856 | 2856_p1 | OR786488 | 23,480 | 28.9 | RepB (Rep_3) | mobA, trwB yukC | none | parA parB | yddH | ||||
| 2859 | 2859_p1 | OR786489 | 36,820 | 31.1 | RepB (Rep_3) | mobA | none | parA | uxa, dctQ kdg operon | ||||
| C. maltaromaticum | 2862 | 2862_p1 | OR786490 | 10,598 | 33.4 | RepB (Rep_3) Rep (HTH_XRE) | none | none | none | pepA | |||
| 2862_p2 | OR786491 | 43,406 | 35.5 | Rep (HTH) | none | none | none | clp | |||||
| 2862_p3 | OR786492 | 60,935 | 33.2 | RepB (HTH_11) Rep (HTH_XRE) | none | mazF | parA parB | celB | amaP | ||||
| 2862_p4 | OR786493 | 66,284 | 33.4 | RepB (HTH_11) Rep (HTH_XRE) | mobA traC traG | parE mazF yoeB | parA | ||||||
| 2862_p5 | OR786494 | 79,510 | 32.4 | RepB (Rep_3) | none | mazE mazF yoeB | parA parB | galE ugd wecB | yjiD | ||||
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Kosiorek, K.; Grzesiak, J.; Gawor, J.; Sałańska, A.; Aleksandrzak-Piekarczyk, T. Polar-Region Soils as Novel Reservoir of Lactic Acid Bacteria from the Genus Carnobacterium. Int. J. Mol. Sci. 2024, 25, 9444. https://doi.org/10.3390/ijms25179444
Kosiorek K, Grzesiak J, Gawor J, Sałańska A, Aleksandrzak-Piekarczyk T. Polar-Region Soils as Novel Reservoir of Lactic Acid Bacteria from the Genus Carnobacterium. International Journal of Molecular Sciences. 2024; 25(17):9444. https://doi.org/10.3390/ijms25179444
Chicago/Turabian StyleKosiorek, Katarzyna, Jakub Grzesiak, Jan Gawor, Agnieszka Sałańska, and Tamara Aleksandrzak-Piekarczyk. 2024. "Polar-Region Soils as Novel Reservoir of Lactic Acid Bacteria from the Genus Carnobacterium" International Journal of Molecular Sciences 25, no. 17: 9444. https://doi.org/10.3390/ijms25179444