
Myosin Isoform-Dependent Effect of Omecamtiv Mecarbil on the Regulation of Force Generation in Human Cardiac Muscle
 , , , and
, , , and
Abstract
:1. Introduction
2. Results
2.1. OM Induces a Rapid and Fully Reversible Modulation of Force Generation in Human Atrial and Ventricular Myofibrils
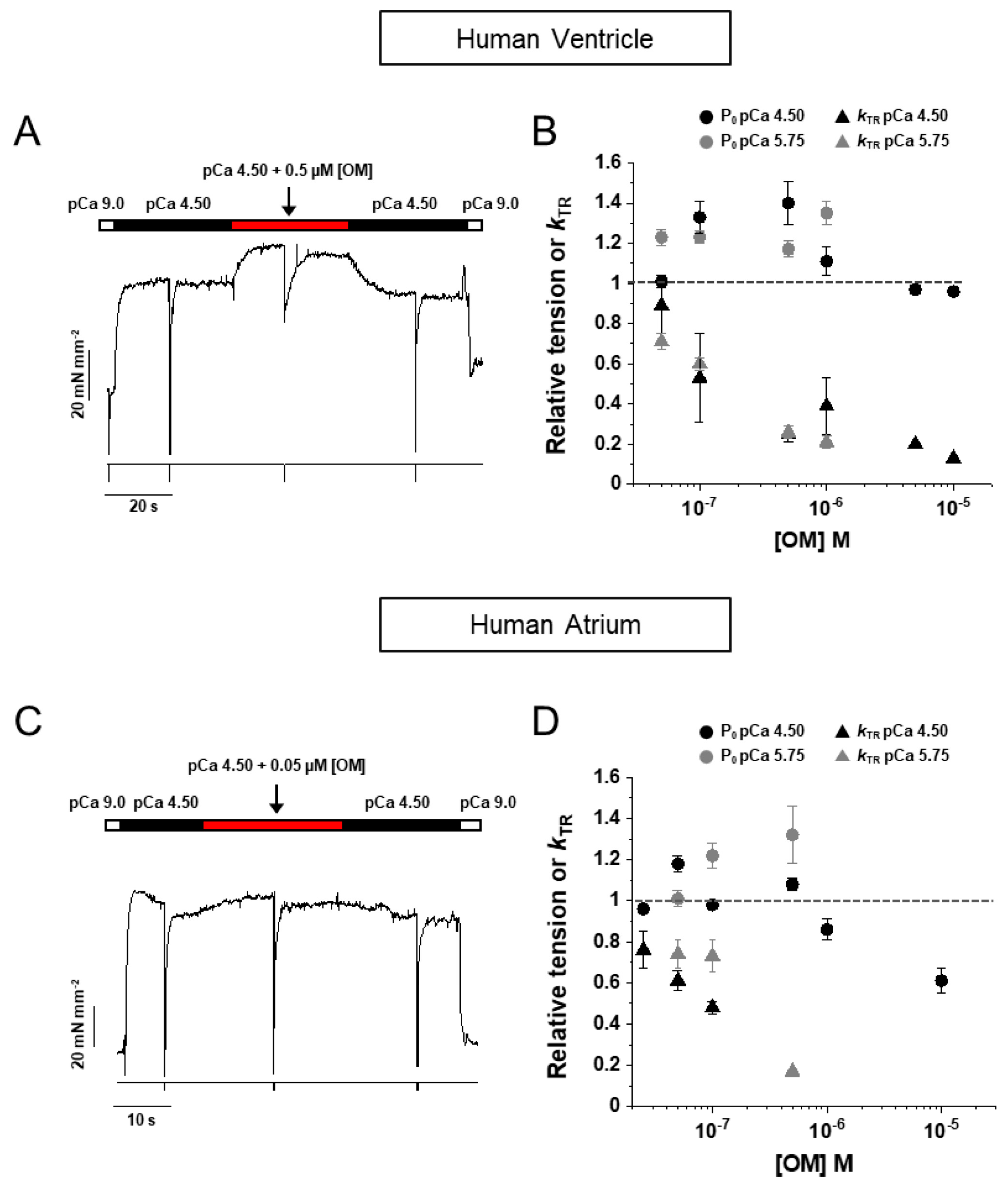
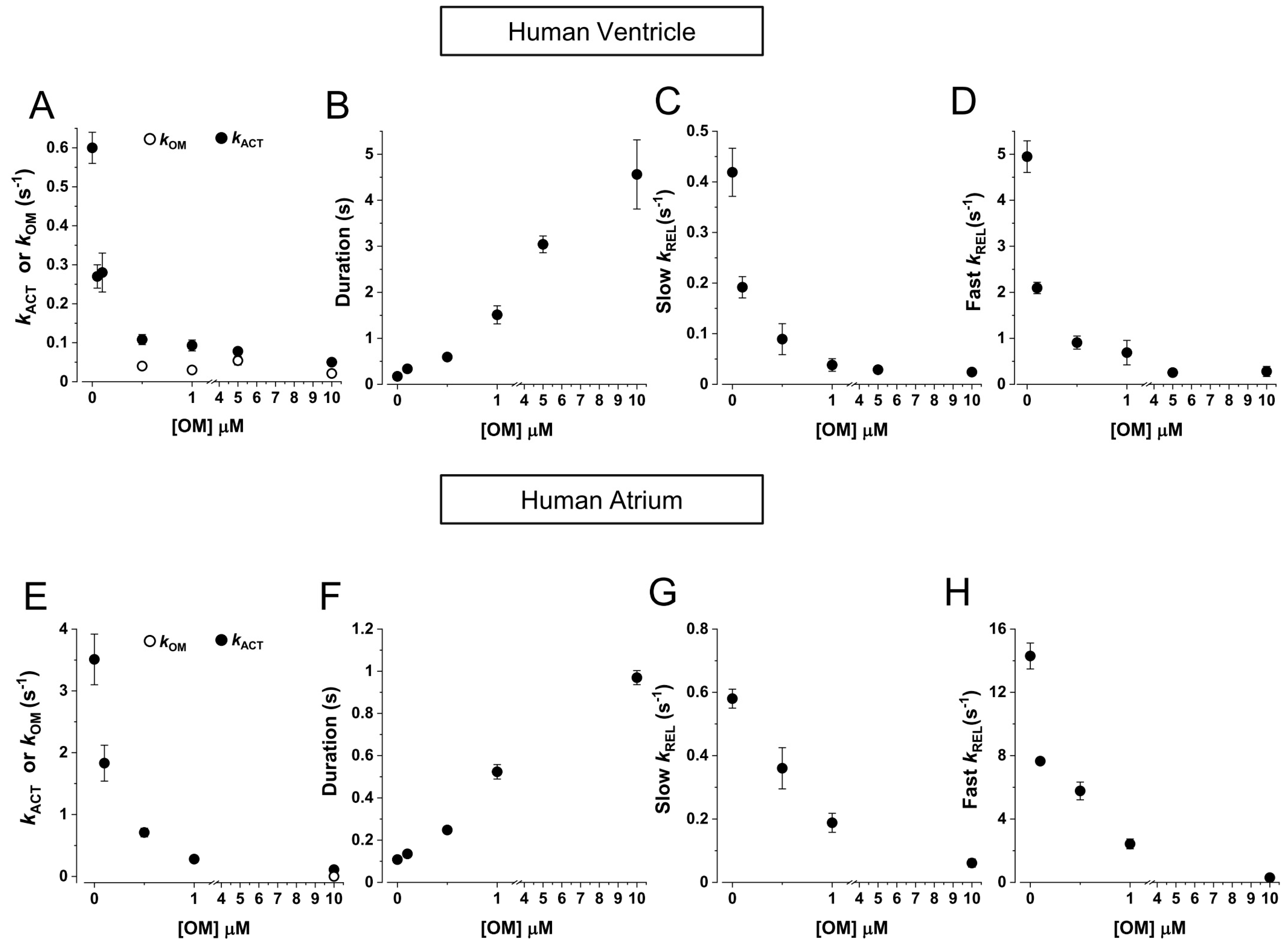
2.2. HV Myofibrils: OM Increases Tension and Decreases the Kinetics of Force Generation and Relaxation Simultaneously Perturbing the Regulation State of Ventricular β-Slow MyHC-7 Sarcomeres
2.3. HA Myofibrils: OM Increases Submaximal Tension and Decreases the Kinetics of Force Generation and Relaxation with Little Effect on the Regulatory State of Atrial α-Fast MyHC-6 Sarcomeres
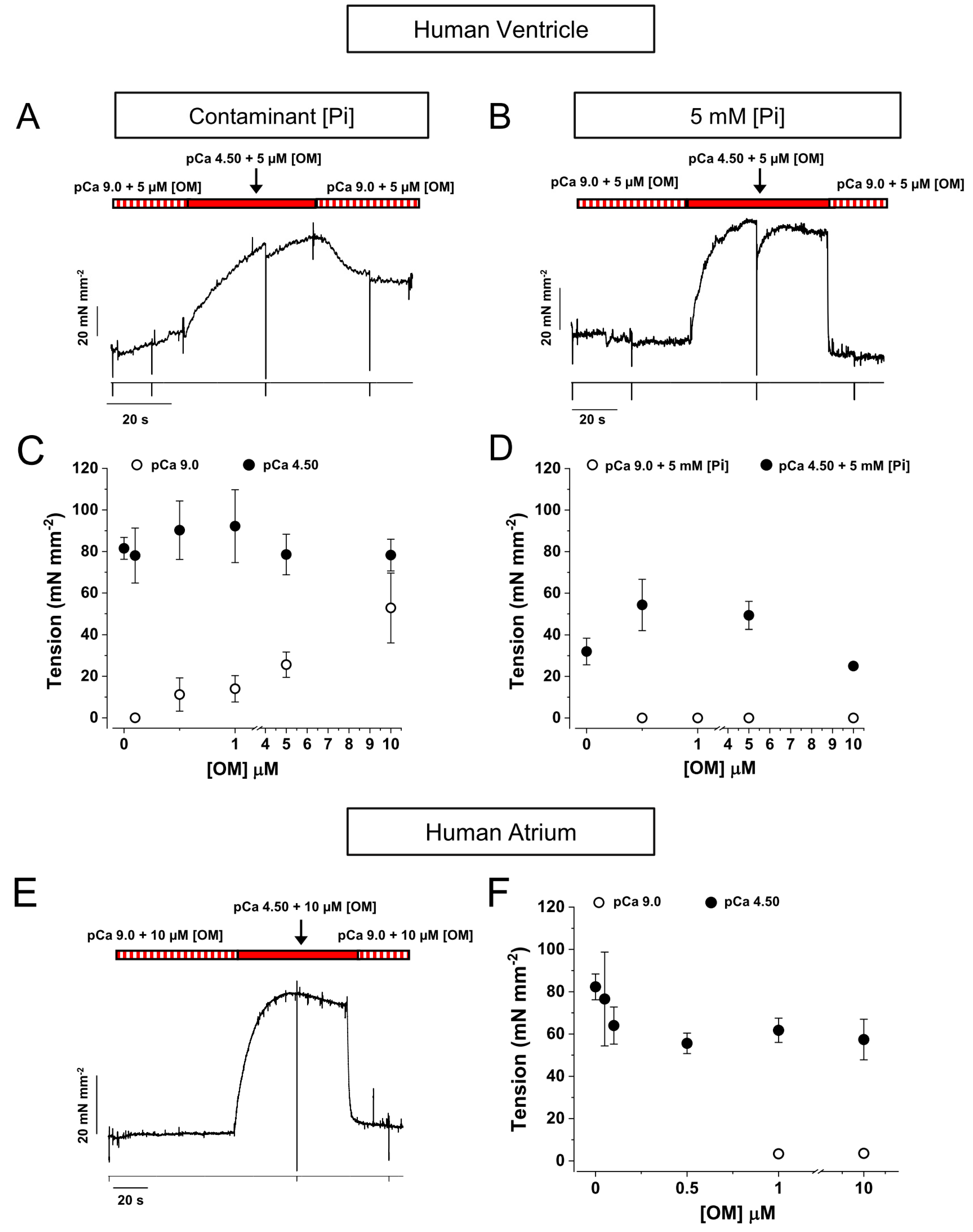
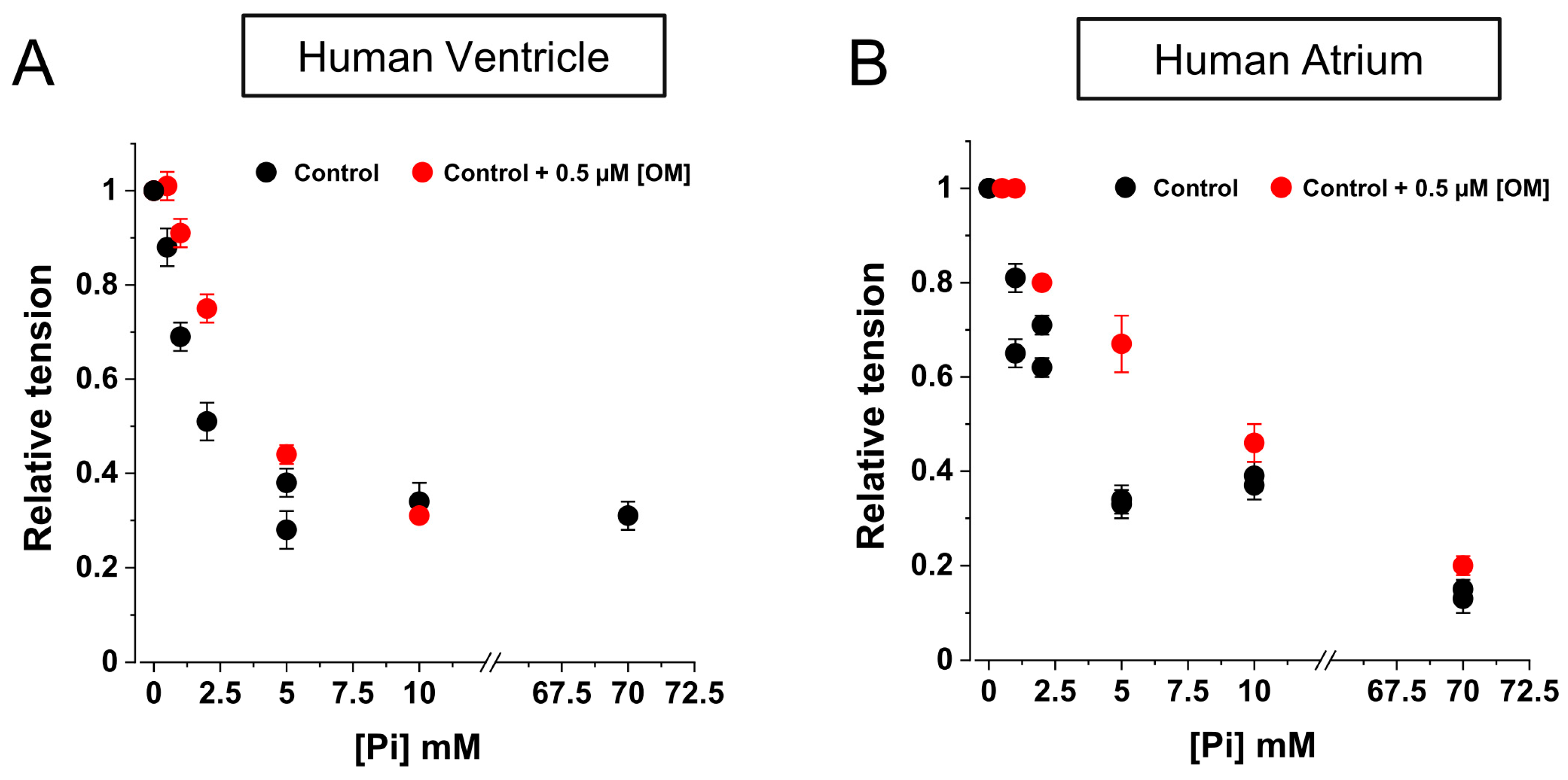
2.4. OM Reduces the Inhibition of Force by Pi in the Low [Pi] Range
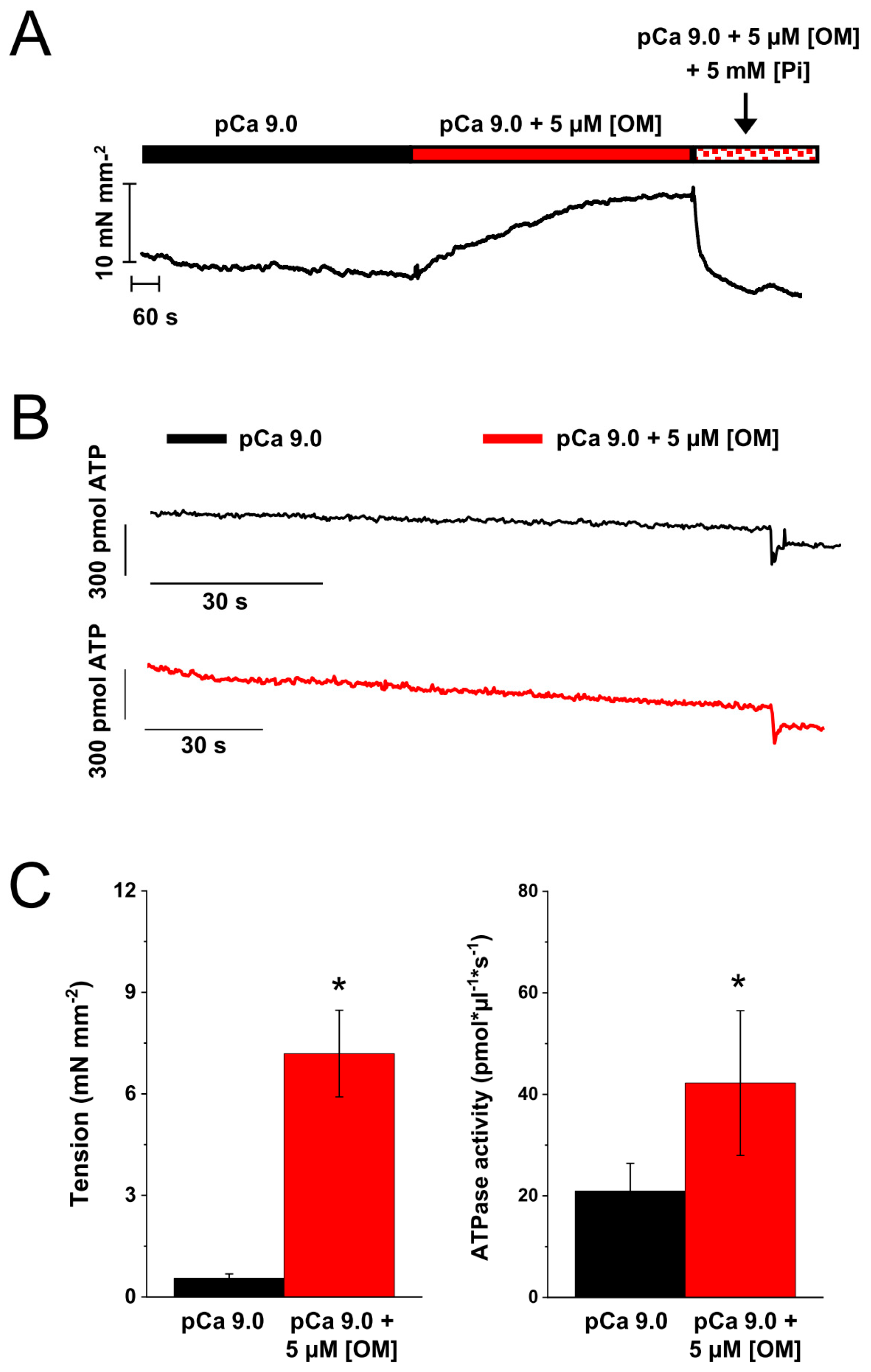
2.5. Effect of OM on the Resting Tension and ATPase of Skinned Strips of Human Ventricular Muscle
3. Discussion
4. Methods
4.1. Preparation of Myofibrils from Human Cardiac Sample
4.2. Electrophoretic Assays
4.3. Myofibril Experiments
4.4. Cardiac Strip Experiments
4.5. Solutions for Mechanical Experiments
4.6. Data Acquisition and Analysis
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Malik, F.I.; Hartman, J.J.; Elias, K.A.; Morgan, B.P.; Rodriguez, H.; Brejc, K.; Anderson, R.L.; Sueoka, S.H.; Lee, K.H.; Finer, J.T.; et al. Cardiac Myosin Activation: A Potential Therapeutic Approach for Systolic Heart Failure. Science 2011, 331, 1439–1443. [Google Scholar] [CrossRef] [PubMed]
- Lehman, S.J.; Crocini, C.; Leinwand, L.A. Targeting the sarcomere in inherited cardiomyopathies. Nat. Rev. Cardiol. 2022, 19, 353–363. [Google Scholar] [CrossRef] [PubMed]
- Day, S.M.; Tardiff, J.C.; Ostap, E.M. Myosin modulators: Emerging approaches for the treatment of cardiomyopathies and heart failure. J. Clin. Investig. 2022, 132, e148557. [Google Scholar] [CrossRef]
- Teerlink, J.R.; Felker, G.M.; McMurray, J.J.; Ponikowski, P.; Metra, M.; Filippatos, G.S.; Ezekowitz, J.A.; Dickstein, K.; Cleland, J.G.; Kim, J.B.; et al. Acute Treatment with Omecamtiv Mecarbil to Increase Contractility in Acute Heart Failure. J. Am. Coll. Cardiol. 2016, 67, 1444–1455. [Google Scholar] [CrossRef]
- Teerlink, J.R.; Felker, G.M.; McMurray, J.J.V.; Solomon, S.D.; Adams, K.F.; Cleland, J.G.F.; Ezekowitz, J.A.; Goudev, A.; Macdonald, P.; Metra, M.; et al. Chronic Oral Study of Myosin Activation to Increase Contractility in Heart Failure (COSMIC-HF): A phase 2, pharmacokinetic, randomised, placebo-controlled trial. Lancet 2016, 388, 2895–2903. [Google Scholar] [CrossRef]
- Teerlink, J.R.; Diaz, R.; Felker, G.M.; McMurray, J.J.; Metra, M.; Solomon, S.D.; Biering-Sørensen, T.; Böhm, M.; Bonderman, D.; Fang, J.C.; et al. Effect of Ejection Fraction on Clinical Outcomes in Patients Treated with Omecamtiv Mecarbil in GALACTIC-HF. J. Am. Coll. Cardiol. 2021, 78, 97–108. [Google Scholar] [CrossRef] [PubMed]
- Lewis, G.D.; Voors, A.A.; Cohen-Solal, A.; Metra, M.; Whellan, D.J.; Ezekowitz, J.A.; Böhm, M.; Teerlink, J.R.; Docherty, K.F.; Lopes, R.D.; et al. Effect of Omecamtiv Mecarbil on Exercise Capacity in Chronic Heart Failure with Reduced Ejection Fraction. JAMA 2022, 328, 259–269. [Google Scholar] [CrossRef]
- Morelli, C.; Ingrasciotta, G.; Jacoby, D.; Masri, A.; Olivotto, I. Sarcomere protein modulation: The new frontier in cardiovascular medicine and beyond. Eur. J. Intern. Med. 2022, 102, 1–7. [Google Scholar] [CrossRef] [PubMed]
- Zhou, S.; Liu, Y.; Huang, X.; Wu, C.; Pórszász, R. Omecamtiv Mecarbil in the treatment of heart failure: The past, the present, and the future. Front. Cardiovasc. Med. 2024, 11, 1337154. [Google Scholar] [CrossRef]
- Mozzini, C.; Pagani, M. The Heart Failure Knights. Curr. Probl. Cardiol. 2023, 48, 101834. [Google Scholar] [CrossRef]
- Karimi, E.; Gohlke, J.; van der Borgh, M.; Lindqvist, J.; Hourani, Z.; Kolb, J.; Cossette, S.; Lawlor, M.W.; Ottenheijm, C.; Granzier, H. Characterization of NEB pathogenic variants in patients reveals novel nemaline myopathy disease mechanisms and omecamtiv mecarbil force effects. Acta Neuropathol. 2024, 147, 72. [Google Scholar] [CrossRef] [PubMed]
- Planelles-Herrero, V.J.; Hartman, J.J.; Robert-Paganin, J.; Malik, F.I.; Houdusse, A. Mechanistic and structural basis for activation of cardiac myosin force production by omecamtiv mecarbil. Nat. Commun. 2017, 8, 190. [Google Scholar] [CrossRef] [PubMed]
- Sparrow, A.J.; Sievert, K.; Patel, S.; Chang, Y.-F.; Broyles, C.N.; Brook, F.A.; Watkins, H.; Geeves, M.A.; Redwood, C.S.; Robinson, P.; et al. Measurement of Myofilament-Localized Calcium Dynamics in Adult Cardiomyocytes and the Effect of Hypertrophic Cardiomyopathy Mutations. Circ. Res. 2019, 124, 1228–1239. [Google Scholar] [CrossRef] [PubMed]
- Kampourakis, T.; Zhang, X.; Sun, Y.; Irving, M. Omecamtiv mercabil and blebbistatin modulate cardiac contractility by perturbing the regulatory state of the myosin filament. J. Physiol. 2017, 596, 31–46. [Google Scholar] [CrossRef]
- Swenson, A.M.; Tang, W.; Blair, C.A.; Fetrow, C.M.; Unrath, W.C.; Previs, M.J.; Campbell, K.S.; Yengo, C.M. Omecamtiv Mecarbil Enhances the Duty Ratio of Human β-Cardiac Myosin Resulting in Increased Calcium Sensitivity and Slowed Force Development in Cardiac Muscle. J. Biol. Chem. 2017, 292, 3768–3778. [Google Scholar] [CrossRef]
- Governali, S.; Caremani, M.; Gallart, C.; Pertici, I.; Stienen, G.; Piazzesi, G.; Ottenheijm, C.; Lombardi, V.; Linari, M. Orthophosphate increases the efficiency of slow muscle-myosin isoform in the presence of omecamtiv mecarbil. Nat. Commun. 2020, 11, 3405. [Google Scholar] [CrossRef]
- Brunello, E.; Fusi, L. Regulating Striated Muscle Contraction: Through Thick and Thin. Annu. Rev. Physiol. 2024, 86, 255–275. [Google Scholar] [CrossRef]
- Johnson, C.A.; Walklate, J.; Svicevic, M.; Mijailovich, S.M.; Vera, C.; Karabina, A.; Leinwand, L.A.; Geeves, M.A. The ATPase cycle of human muscle myosin II isoforms: Adaptation of a single mechanochemical cycle for different physiological roles. J. Biol. Chem. 2019, 294, 14267–14278. [Google Scholar] [CrossRef]
- Walklate, J.; Ferrantini, C.; Johnson, C.A.; Tesi, C.; Poggesi, C.; Geeves, M.A. Alpha and beta myosin isoforms and human atrial and ventricular contraction. Cell. Mol. Life Sci. 2021, 78, 7309–7337. [Google Scholar] [CrossRef]
- McKillop, D.; Geeves, M. Regulation of the interaction between actin and myosin subfragment 1: Evidence for three states of the thin filament. Biophys. J. 1993, 65, 693–701. [Google Scholar] [CrossRef]
- Shchepkin, D.V.; Nabiev, S.R.; Nikitina, L.V.; Kochurova, A.M.; Berg, V.Y.; Bershitsky, S.Y.; Kopylova, G.V. Myosin from the ventricle is more sensitive to omecamtiv mecarbil than myosin from the atrium. Biochem. Biophys. Res. Commun. 2020, 528, 658–663. [Google Scholar] [CrossRef] [PubMed]
- Nakanishi, T.; Oyama, K.; Tanaka, H.; Kobirumaki-Shimozawa, F.; Ishii, S.; Terui, T.; Ishiwata, S.; Fukuda, N. Effects of omecamtiv mecarbil on the contractile properties of skinned porcine left atrial and ventricular muscles. Front. Physiol. 2022, 13, 947206. [Google Scholar] [CrossRef] [PubMed]
- Piroddi, N.; Belus, A.; Scellini, B.; Tesi, C.; Giunti, G.; Cerbai, E.; Mugelli, A.; Poggesi, C. Tension generation and relaxation in single myofibrils from human atrial and ventricular myocardium. Pflügers Arch. Eur. J. Physiol. 2006, 454, 63–73. [Google Scholar] [CrossRef] [PubMed]
- Scellini, B.; Piroddi, N.; Dente, M.; Vitale, G.; Pioner, J.M.; Coppini, R.; Ferrantini, C.; Poggesi, C.; Tesi, C. Mavacamten has a differential impact on force generation in myofibrils from rabbit psoas and human cardiac muscle. J. Gen. Physiol. 2021, 153, e202012789. [Google Scholar] [CrossRef]
- Tesi, C.; Colomo, F.; Nencini, S.; Piroddi, N.; Poggesi, C. The Effect of Inorganic Phosphate on Force Generation in Single Myofibrils from Rabbit Skeletal Muscle. Biophys. J. 2000, 78, 3081–3092. [Google Scholar] [CrossRef]
- Barsotti, R.J.; Ferenczi, M.A. Kinetics of ATP hydrolysis and tension production in skinned cardiac muscle of the guinea pig. J. Biol. Chem. 1988, 263, 16750–16756. [Google Scholar] [CrossRef]
- Drazner, M.H. Omecamtiv Mecarbil as a Therapy for Heart Failure with Low Ejection Fraction. JAMA 2022, 328, 249–250. [Google Scholar] [CrossRef]
- Mamidi, R.; Li, J.; Gresham, K.S.; Verma, S.; Doh, C.Y.; Li, A.; Lal, S.; dos Remedios, C.G.; Stelzer, J.E. Dose-Dependent Effects of the Myosin Activator Omecamtiv Mecarbil on Cross-Bridge Behavior and Force Generation in Failing Human Myocardium. Circ. Hear. Fail. 2017, 10, e004257. [Google Scholar] [CrossRef]
- Nagy, L.; Kovács, Á.; Bódi, B.; Pásztor, E.T.; Fülöp, G.Á.; Tóth, A.; Édes, I.; Papp, Z. The novel cardiac myosin activator omecamtiv mecarbil increases the calcium sensitivity of force production in isolated cardiomyocytes and skeletal muscle fibres of the rat. Br. J. Pharmacol. 2015, 172, 4506–4518. [Google Scholar] [CrossRef]
- Kieu, T.T.; Awinda, P.O.; Tanner, B.C. Omecamtiv Mecarbil Slows Myosin Kinetics in Skinned Rat Myocardium at Physiological Temperature. Biophys. J. 2019, 116, 2149–2160. [Google Scholar] [CrossRef]
- Gollapudi, S.K.; Reda, S.M.; Chandra, M. Omecamtiv Mecarbil Abolishes Length-Mediated Increase in Guinea Pig Cardiac Myofiber Ca2+ Sensitivity. Biophys. J. 2017, 113, 880–888. [Google Scholar] [CrossRef] [PubMed]
- Fülöp, G.Á.; Oláh, A.; Csipo, T.; Kovács, Á.; Pórszász, R.; Veress, R.; Horváth, B.; Nagy, L.; Bódi, B.; Fagyas, M.; et al. Omecamtiv mecarbil evokes diastolic dysfunction and leads to periodic electromechanical alternans. Basic Res. Cardiol. 2021, 116, 24. [Google Scholar] [CrossRef]
- Abella, L.M.R.; Höhm, C.; Hofmann, B.; Gergs, U.; Neumann, J. Effects of omecamtiv mecarbil and mavacamten in isolated human atrium. Naunyn-Schmiedeberg’s Arch. Pharmacol. 2023, 396, 499–511. [Google Scholar] [CrossRef] [PubMed]
- Woody, M.S.; Greenberg, M.J.; Barua, B.; Winkelmann, D.A.; Goldman, Y.E.; Ostap, E.M. Positive cardiac inotrope omecamtiv mecarbil activates muscle despite suppressing the myosin working stroke. Nat. Commun. 2018, 9, 3838. [Google Scholar] [CrossRef] [PubMed]
- Rohde, J.A.; Thomas, D.D.; Muretta, J.M. Heart failure drug changes the mechanoenzymology of the cardiac myosin powerstroke. Proc. Natl. Acad. Sci. USA 2017, 114, E1796–E1804. [Google Scholar] [CrossRef]
- Liu, Y.; White, H.D.; Belknap, B.; Winkelmann, D.A.; Forgacs, E. Omecamtiv Mecarbil Modulates the Kinetic and Motile Properties of Porcine β-Cardiac Myosin. Biochemistry 2015, 54, 1963–1975. [Google Scholar] [CrossRef]
- Hashem, S.; Tiberti, M.; Fornili, A. Allosteric modulation of cardiac myosin dynamics by omecamtiv mecarbil. PLOS Comput. Biol. 2017, 13, e1005826. [Google Scholar] [CrossRef]
- Winkelmann, D.A.; Forgacs, E.; Miller, M.T.; Stock, A.M. Structural basis for drug-induced allosteric changes to human β-cardiac myosin motor activity. Nat. Commun. 2015, 6, 7974. [Google Scholar] [CrossRef]
- Wang, Y.; Ajtai, K.; Burghardt, T.P. Analytical Comparison of Natural and Pharmaceutical Ventricular Myosin Activators. Biochemistry 2014, 53, 5298–5306. [Google Scholar] [CrossRef]
- Park-Holohan, S.-J.; Brunello, E.; Kampourakis, T.; Rees, M.; Irving, M.; Fusi, L. Stress-dependent activation of myosin in the heart requires thin filament activation and thick filament mechanosensing. Proc. Natl. Acad. Sci. USA 2021, 118, e2023706118. [Google Scholar] [CrossRef]
- Narolska, N.; Vanloon, R.; Boontje, N.; Zaremba, R.; Penas, S.E.; Russell, J.; Spiegelenberg, S.; Huybregts, M.; Visser, F.; Dejong, J.; et al. Myocardial contraction is 5-fold more economical in ventricular than in atrial human tissue. Cardiovasc. Res. 2005, 65, 221–229. [Google Scholar] [CrossRef]
- Wu, F.; Zhang, E.Y.; Zhang, J.; Bache, R.J.; Beard, D.A. Phosphate metabolite concentrations and ATP hydrolysis potential in normal and ischaemic hearts. J. Physiol. 2008, 586, 4193–4208. [Google Scholar] [CrossRef] [PubMed]
- Teerlink, J.R.; Clarke, C.P.; Saikali, K.G.; Lee, J.H.; Chen, M.M.; Escandon, R.D.; Elliott, L.; Bee, R.; Habibzadeh, M.R.; Goldman, J.H.; et al. Dose-dependent augmentation of cardiac systolic function with the selective cardiac myosin activator, omecamtiv mecarbil: A first-in-man study. Lancet 2011, 378, 667–675. [Google Scholar] [CrossRef]
- Reiser, P.J.; Moravec, C.S. Sex differences in myosin heavy chain isoforms of human failing and nonfailing atria. Am. J. Physiol. Circ. Physiol. 2014, 307, H265–H272. [Google Scholar] [CrossRef] [PubMed]
- Reiser, P.J.; Kline, W.O.; Broughton, K.M.; Li, J.; Sarmah, E.; Warren, C.M.; Lin, Y.-H.; Henze, M.P.; Sanchez-Freire, V.; Solaro, R.J.; et al. Electrophoretic separation and quantitation of cardiac myosin heavy chain isoforms in eight mammalian species. Am. J. Physiol. Circ. Physiol. 1998, 274, H1048–H1053. [Google Scholar] [CrossRef]
- Talmadge, R.J.; Roy, R.R. Electrophoretic separation of rat skeletal muscle myosin heavy-chain isoforms. J. Appl. Physiol. 1993, 75, 2337–2340. [Google Scholar] [CrossRef] [PubMed]
- Brenner, B. Effect of Ca2+ on cross-bridge turnover kinetics in skinned single rabbit psoas fibers: Implications for regulation of muscle contraction. Proc. Natl. Acad. Sci. USA 1988, 85, 3265–3269. [Google Scholar] [CrossRef]
- Tesi, C.; Piroddi, N.; Colomo, F.; Poggesi, C. Relaxation Kinetics Following Sudden Ca2+Reduction in Single Myofibrils from Skeletal Muscle. Biophys. J. 2002, 83, 2142–2151. [Google Scholar] [CrossRef]


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Tension Generation | Relaxation | ||||
|---|---|---|---|---|---|
| Slow Phase | Fast Phase | ||||
| P0 | kACT | kTR | Duration | slow kREL | fast kREL |
| mN mm−2 | s−1 | s−1 | ms | s−1 | s−1 |
| Human ventricle | |||||
| 82 ± 5 (55) | 0.60 ± 0.04 (23) | 0.44 ± 0.01 (54) | 173 ± 10 (23) | 0.42 ± 0.05 (22) | 4.95 ± 0.34 (23) |
| Human atrium | |||||
| 82 ± 6 (49) | 3.51 ± 0.41 (14) | 2.96 ± 0.16 (44) | 108 ± 6 (14) | 0.58 ± 0.03 (14) | 14.3 ± 0.82 (14) |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Scellini, B.; Piroddi, N.; Dente, M.; Pioner, J.M.; Ferrantini, C.; Poggesi, C.; Tesi, C. Myosin Isoform-Dependent Effect of Omecamtiv Mecarbil on the Regulation of Force Generation in Human Cardiac Muscle. Int. J. Mol. Sci. 2024, 25, 9784. https://doi.org/10.3390/ijms25189784
Scellini B, Piroddi N, Dente M, Pioner JM, Ferrantini C, Poggesi C, Tesi C. Myosin Isoform-Dependent Effect of Omecamtiv Mecarbil on the Regulation of Force Generation in Human Cardiac Muscle. International Journal of Molecular Sciences. 2024; 25(18):9784. https://doi.org/10.3390/ijms25189784
Chicago/Turabian StyleScellini, Beatrice, Nicoletta Piroddi, Marica Dente, J. Manuel Pioner, Cecilia Ferrantini, Corrado Poggesi, and Chiara Tesi. 2024. "Myosin Isoform-Dependent Effect of Omecamtiv Mecarbil on the Regulation of Force Generation in Human Cardiac Muscle" International Journal of Molecular Sciences 25, no. 18: 9784. https://doi.org/10.3390/ijms25189784