

The ABCG2 Transporter Affects Plasma Levels, Tissue Distribution and Milk Secretion of Lumichrome, a Natural Derivative of Riboflavin

 , , ,
, , , 
Abstract
:1. Introduction
2. Results
2.1. Murine Abcg2 and Human ABCG2 Efficiently In Vitro Transport Lumichrome
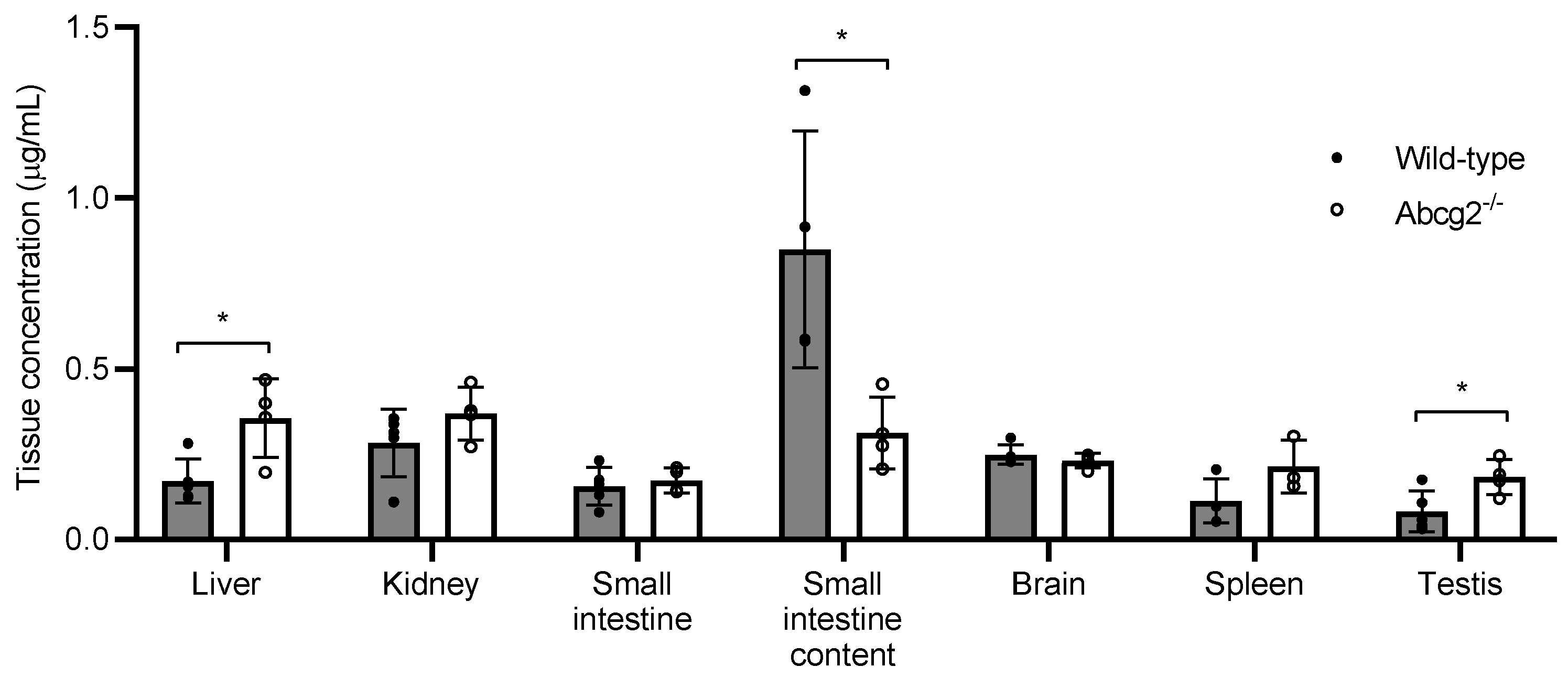
2.2. Plasma Levels and Tissue Distribution of Lumichrome in Wild-Type and Abcg2-/- Male Mice
2.3. Secretion of Lumichrome into Milk in Wild-Type and Abcg2-/- Lactating Female Mice
3. Discussion
4. Materials and Methods
4.1. Chemicals
4.2. Cell Cultures
4.3. Transcellular Transport Assays
4.4. Animals
4.4.1. Plasma and Tissue Distribution
4.4.2. Milk Secretion Experiments
4.5. High-Performance Liquid Chromatographic Analysis
4.6. Statistical Analysis
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Vlaming, M.L.; Lagas, J.S.; Schinkel, A.H. Physiological and pharmacological roles of ABCG2 (BCRP): Recent findings in Abcg2 knockout mice. Adv. Drug Deliv. Rev. 2008, 61, 14–25. [Google Scholar] [CrossRef] [PubMed]
- Horsey, A.J.; Cox, M.H.; Sarwat, S.; Kerr, I.D. The multidrug transporter ABCG2: Still more questions than answers. Biochem. Soc. Trans. 2016, 44, 824–830. [Google Scholar] [CrossRef]
- Khunweeraphong, N.; Szöllősi, D.; Stockner, T.; Kuchler, K. The ABCG2 multidrug transporter is a pump gated by a valve and an extracellular lid. Nat. Commun. 2019, 10, 1–14. [Google Scholar] [CrossRef] [PubMed]
- Kukal, S.; Guin, D.; Rawat, C.; Bora, S.; Mishra, M.K.; Sharma, P.; Paul, P.R.; Kanojia, N.; Grewal, G.K.; Kukreti, S.; et al. Multi-drug Efflux Transporter ABCG2: Expression and Regulation. Cell. Mol. Life Sci. 2021, 78, 6887–6939. [Google Scholar] [CrossRef]
- Qi, X.; Chen, H.; Guan, K.; Wang, R.; Ma, Y. Anti-hyperuricemic and nephroprotective effects of whey protein hydrolysate in potassium oxonate induced hyperuricemic rats. J. Sci. Food Agric. 2021, 101, 4916–4924. [Google Scholar] [CrossRef] [PubMed]
- García-Lino, A.M.; Álvarez-Fernández, I.; Blanco-Paniagua, E.; Merino, G.; Álvarez, A.I. Transporters in the Mammary Gland—Contribution to Presence of Nutrients and Drugs into Milk. Nutrients 2019, 11, 2372. [Google Scholar] [CrossRef]
- Prukała, D.; Sikorska, E.; Koput, J.; Khmelinskii, I.; Karolczak, J.; Gierszewski, M.; Sikorski, M. Acid-Base Equilibriums of Lu-michrome and Its 1-Methyl, 3-Methyl, and 1,3-Dimethyl Derivatives. J. Phys. Chem. A 2012, 116, 7474–7490. [Google Scholar] [CrossRef]
- Ahmad, I.; Fasihullah, Q.; Vaid, F.H.M. A Study of Simultaneous Photolysis and Photoaddition Reactions of Riboflavin in Aqueous Solution. J. Photochem. Photobiol. B Biol. 2004, 75, 13–20. [Google Scholar] [CrossRef]
- Huang, R.; Kim, H.J.; Min, D.B. Photosensitizing Effect of Riboflavin, Lumiflavin, and Lumichrome on the Generation of Volatiles in Soy Milk. J. Agric. Food Chem. 2006, 54, 2359–2364. [Google Scholar] [CrossRef]
- Remucal, C.K.; McNeill, K. Photosensitized Amino Acid Degradation in the Presence of Riboflavin and Its Derivatives. Environ. Sci. Technol. 2011, 45, 5230–5237. [Google Scholar] [CrossRef]
- Bergh, V.J.V.; Tønnesen, H.H. Interaction between the photosensitizer lumichrome and human serum albumin: Effect of excipients. Pharm. Dev. Technol. 2016, 22, 992–1000. [Google Scholar] [CrossRef] [PubMed]
- Hardwick, C.C.; Herivel, T.R.; Hernandez, S.C.; Ruane, P.H.; Goodrich, R.P. Separation, Identification and Quantification of Riboflavin and Its Photoproducts in Blood Products Using High-performance Liquid Chromatography with Fluorescence De-tection: A Method to Support Pathogen Reduction Technology. Photochem. Photobiol. 2004, 80, 609–615. [Google Scholar] [PubMed]
- Jana, R.; Gautam, R.K.; Bapli, A.; Seth, D. Photodynamics of Biological Active Flavin in the Presence of Zwitterionic Surfac-tants. Spectrochim. Acta Part A Mol. Biomol. Spectrosc. 2022, 264, 120304. [Google Scholar] [CrossRef]
- Oka, M.; McCormick, D.B. Urinary Lumichrome-Level Catabolites of Riboflavin are due to Microbial and Photochemical Events and not Rat Tissue Enzymatic Cleavage of the Ribityl Chain. J. Nutr. 1985, 115, 496–499. [Google Scholar] [CrossRef]
- Hamada, K.; Sasaki, M.; Yoshimura, K. Two New Types of Riboflavin-Decomposing Bacteria Isolated from Human Feces. J. Vitaminol. 1956, 2, 307–315. [Google Scholar] [CrossRef]
- Yamamoto, K.; Asano, Y. Efficient Production of Lumichrome by Microbacterium sp. Strain TPU 3598. Appl. Environ. Microbiol. 2015, 81, 7360–7367. [Google Scholar] [CrossRef] [PubMed]
- Foster, J.W. Microbiological Aspects of Riboflavin: III. Oxidation Studies with Pseudomonas Riboflavina. J. Bacteriol. 1944, 48, 97–111. [Google Scholar] [CrossRef]
- Alves, A.J.S.; Pereira, J.A.; Dethoup, T.; Cravo, S.; Mistry, S.; Silva, A.M.S.; Pinto, M.M.M.; Kijjoa, A. A New Meroterpene, A New Benzofuran Derivative and Other Constituents from Cultures of the Marine Sponge-Associated Fungus Acremonium persicinum KUFA 1007 and Their Anticholinesterase Activities. Mar. Drugs 2019, 17, 379. [Google Scholar] [CrossRef]
- Kino, K.; Nakatsuma, A.; Nochi, H.; Kiriyama, Y.; Kurita, T.; Kobayashi, T.; Miyazawa, H. Commentary on the Phototoxicity and Absorption of Vitamin B2 and Its Degradation Product, Lumichrome. Pharm. Anal. Acta 2015, 6, 8–10. [Google Scholar]
- Kino, K.; Kobayashi, T.; Arima, E.; Komori, R.; Kobayashi, T.; Miyazawa, H. Photoirradiation products of flavin derivatives, and the effects of photooxidation on guanine. Bioorg. Med. Chem. Lett. 2009, 19, 2070–2074. [Google Scholar] [CrossRef]
- Seng, F.; Ley, K. A New Synthesis of Lumichrome. Angew. Chem. Int. Ed. Engl. 1972, 11, 1010–1011. [Google Scholar] [CrossRef]
- Huang, R.; Choe, E.; Min, D. Effects of Riboflavin Photosensitized Oxidation on the Volatile Compounds of Soymilk. J. Food Sci. 2004, 69, C733–C738. [Google Scholar] [CrossRef]
- Ahmad, I.; Anwar, Z.; Sheraz, M.A.; Ahmed, S.; Khattak, S.U.R. Stability-indicating spectrofluorimetric method for the assay of riboflavin and photoproducts: Kinetic applications. Luminescence 2018, 33, 1070–1080. [Google Scholar] [CrossRef] [PubMed]
- Marchena, M.; Gil, M.; Martín, C.; Organero, J.A.; Sanchez, F.; Douhal, A. Stability and Photodynamics of Lumichrome Struc-tures in Water at Different PHs and in Chemical and Biological Caging Media. J. Phys. Chem. B 2011, 115, 2424–2435. [Google Scholar] [CrossRef]
- Cunningham, O.; Gore, M.G.; Mantle, T.J. Initial-Rate Kinetics of the Flavin Reductase Reaction Catalysed by Human Biliver-din-IXβ Reductase (BVR-B). Biochem. J. 2000, 345, 393–399. [Google Scholar] [CrossRef]
- Fieschi, F.; Nivière, V.; Frier, C.; Décout, J.-L.; Fontecave, M. The Mechanism and Substrate Specificity of the NADPH:Flavin Oxidoreductase from Escherichia coli. J. Biol. Chem. 1995, 270, 30392–30400. [Google Scholar] [CrossRef] [PubMed]
- Shalloe, F.; Elliott, G.; Ennis, O.; Mantle, T.J. Evidence that biliverdin-IXβ reductase and flavin reductase are identical. Biochem. J. 1996, 316, 385–387. [Google Scholar] [CrossRef]
- Said, H.M.; Ortiz, A.; Ma, T.Y.; McCloud, E. Riboflavin uptake by the human-derived liver cells Hep G2: Mechanism and regulation. J. Cell. Physiol. 1998, 176, 588–594. [Google Scholar] [CrossRef]
- Grønlien, K.G.; Pedersen, M.E.; Rønning, S.B.; Solberg, N.T.; Tønnesen, H.H. Tuning of 2D Cultured Human Fibroblast Behav-ior Using Lumichrome Photocrosslinked Collagen Hydrogels. Mater. Today Commun. 2022, 31, 103635. [Google Scholar] [CrossRef]
- Chantarawong, W.; Kuncharoen, N.; Tanasupawat, S.; Chanvorachote, P. Lumichrome Inhibits Human Lung Cancer Cell Growth and Induces Apoptosis via a p53-Dependent Mechanism. Nutr. Cancer 2019, 71, 1390–1402. [Google Scholar] [CrossRef]
- Etu, S.F.; Alqanthan, A.; Qais, N. 5,10-Dihydro-7,8-Dimethyl Alloxazine as an Anticancer Agent from Lumichrome of Ribofla-vin. J. Drug Des. Dev. 2021, 2, 44–48. [Google Scholar]
- Tan, D.; Ma, A.; Wang, S.; Zhang, Q.; Jia, M.; Kamal-Eldin, A.; Wu, H.; Chen, G. Effects of the Oxygen Content and Light Inten-sity on Milk Photooxidation Using Untargeted Metabolomic Analysis. J. Agric. Food Chem. 2021, 69, 7488–7497. [Google Scholar] [CrossRef] [PubMed]
- Tan, S.; Li, Q.; Guo, C.; Chen, S.; Kamal-Eldin, A.; Chen, G. Reveal the mechanism of hepatic oxidative stress in mice induced by photo-oxidation milk using multi-omics analysis techniques. J. Adv. Res. 2024. [Google Scholar] [CrossRef] [PubMed]
- Fracassetti, D.; Limbo, S.; D’incecco, P.; Tirelli, A.; Pellegrino, L. Development of a HPLC method for the simultaneous analysis of riboflavin and other flavin compounds in liquid milk and milk products. Eur. Food Res. Technol. 2018, 244, 1545–1554. [Google Scholar] [CrossRef]
- Min, D.B.; Boff, J.M. Chemistry and Reaction of Singlet Oxygen in Foods. Compr. Rev. Food Sci. Food Saf. 2002, 1, 58–72. [Google Scholar] [CrossRef]
- Keim, J.P.; Daza, J.; Beltrán, I.; Balocchi, O.A.; Pulido, R.G.; Sepúlveda-Varas, P.; Pacheco, D.; Berthiaume, R. Milk Production Responses, Rumen Fermentation, and Blood Metabolites of Dairy Cows Fed Increasing Concentrations of Forage Rape (Bras-sica Napus Ssp. Biennis). J. Dairy Sci. 2020, 103, 9054–9066. [Google Scholar] [CrossRef]
- van Herwaarden, A.E.; Wagenaar, E.; Merino, G.; Jonker, J.W.; Rosing, H.; Beijnen, J.H.; Schinkel, A.H. Multidrug Transporter ABCG2/Breast Cancer Resistance Protein Secretes Riboflavin (Vitamin B2) into Milk. Mol. Cell. Biol. 2007, 27, 1247–1253. [Google Scholar] [CrossRef]
- Liu, C.; Cao, Z.; Zhang, W.; Tickner, J.; Qiu, H.; Wang, C.; Chen, K.; Wang, Z.; Tan, R.; Dong, S.; et al. Lumichrome inhibits osteoclastogenesis and bone resorption through suppressing RANKL-induced NFAT activation and calcium signaling. J. Cell. Physiol. 2018, 233, 8971–8983. [Google Scholar] [CrossRef]
- Garcia-Lino, A.M.; Blanco-Paniagua, E.; Astorga-Simon, E.N.; Alvarez-Fernandez, L.; Garcia-Mateos, D.; Alvarez-Fernandez, I.; Alvarez, A.I.; Merino, G. Abcg2 transporter affects plasma, milk and tissue levels of meloxicam. Biochem. Pharmacol. 2020, 175, 113924. [Google Scholar] [CrossRef]
- Blanco-Paniagua, E.; Álvarez- Fernández, L.; Garcia-Lino, A.M.; Álvarez, A.I.; Merino, G. Secretion into Milk of the Main Me-tabolites of the Anthelmintic Albendazole Is Mediated by the ABCG2/BCRP Transporter. Antimicrob. Agents Chemother. 2022, 66, e0006222. [Google Scholar] [CrossRef]
- Real, R.; Egido, E.; Pérez, M.; González-Lobato, L.; Barrera, B.; Prieto, J.G.; Álvarez, A.I.; Merino, G. Involvement of breast cancer resistance protein (BCRP/ABCG2) in the secretion of danofloxacin into milk: Interaction with ivermectin. J. Veter Pharmacol. Ther. 2010, 34, 313–321. [Google Scholar] [CrossRef] [PubMed]
- Insińska-Rak, M.; Sikorski, M.; Wolnicka-Glubisz, A. Riboflavin and Its Derivates as Potential Photosensitizers in the Photo-dynamic Treatment of Skin Cancers. Cells 2023, 12, 2304. [Google Scholar] [CrossRef] [PubMed]
- Hira, D.; Terada, T. BCRP/ABCG2 and high-alert medications: Biochemical, pharmacokinetic, pharmacogenetic, and clinical implications. Biochem. Pharmacol. 2018, 147, 201–210. [Google Scholar] [CrossRef] [PubMed]
- Polgar, O.; Robey, R.W.; E Bates, S. ABCG2: Structure, function and role in drug response. Expert. Opin. Drug Metab. Toxicol. 2007, 4, 1–15. [Google Scholar] [CrossRef] [PubMed]
- Wu, C.; Lusvarghi, S.; Hsiao, S.; Liu, T.; Li, Y.; Huang, Y.; Hung, T.; Chang, T.; Memorial, G. Licochalcone A Selectively Resensi-tizes ABCG2-Overexpressing Multidrug-Resistant Cancer Cells to Chemotherapeutic Drugs. J. Nat. Prod. 2020, 83, 1461–1472. [Google Scholar] [CrossRef]
- Perez, M.; Otero, J.A.; Barrera, B.; Prieto, J.G.; Merino, G.; Alvarez, A.I. Inhibition of ABCG2/BCRP Transporter by Soy Isofla-vones Genistein and Daidzein: Effect on Plasma and Milk Levels of Danofloxacin in Sheep. Vet. J. 2013, 196, 203–208. [Google Scholar] [CrossRef]
- Gunes, Y.; Okyar, A.; Krajcsi, P.; Fekete, Z.; Ustuner, O. Modulation of monepantel secretion into milk by soy isoflavones. J. Veter-Pharmacol. Ther. 2022, 46, 185–194. [Google Scholar] [CrossRef]
- Otero, J.A.; García-Mateos, D.; Alvarez-Fernández, I.; García-Villalba, R.; Espín, J.C.; Álvarez, A.I.; Merino, G. Flaxseed-enriched diets change milk concentration of the antimicrobial danofloxacin in sheep. BMC Veter Res. 2018, 14, 14. [Google Scholar] [CrossRef]
- Mao, Q.; Unadkat, J.D. Role of the Breast Cancer Resistance Protein (BCRP/ABCG2) in Drug Transport—An Update. AAPS J. 2014, 17, 65–82. [Google Scholar] [CrossRef]
- Sabet, Z.; Vagiannis, D.; Budagaga, Y.; Zhang, Y.; Novotná, E.; Hanke, I.; Rozkoš, T.; Hofman, J. Talazoparib Does Not Interact with ABCB1 Transporter or Cytochrome P450s, but Modulates Multidrug Resistance Mediated by ABCC1 and ABCG2: An in Vitro and Ex Vivo Study. Int. J. Mol. Sci. 2022, 23, 14338. [Google Scholar] [CrossRef]
- Sharma, S.; Mettu, V.S.; Prasad, B. Interplay of Breast Cancer Resistance Protein (Bcrp/Abcg2), Sex, and Fed State in Oral Pharmacokinetic Variability of Furosemide in Rats. Pharmaceutics 2023, 15, 542. [Google Scholar] [CrossRef] [PubMed]
- Blazquez, A.M.G.; Macias, R.I.R.; Cives-Losada, C.; de la Iglesia, A.; Marin, J.J.G.; Monte, M.J. Lactation during cholestasis: Role of ABC proteins in bile acid traffic across the mammary gland. Sci. Rep. 2017, 7, 7475. [Google Scholar] [CrossRef] [PubMed]
- lvarez-Fernández, L.; Gomez-Gomez, A.; Haro, N.; García-Lino, A.M.; Álvarez, A.I.; Pozo, O.J.; Merino, G. ABCG2 Trans-porter Plays a Key Role in the Biodistribution of Melatonin and Its Main Metabolites. J. Pineal Res. 2023, 74, e12849. [Google Scholar] [CrossRef] [PubMed]
- Egido, E.; Müller, R.; Li-Blatter, X.; Merino, G.; Seelig, A. Predicting Activators and Inhibitors of the Breast Cancer Resistance Protein (ABCG2) and P-Glycoprotein (ABCB1) Based on Mechanistic Considerations. Mol. Pharm. 2015, 12, 4026–4037. [Google Scholar] [CrossRef]
- Riboflavin|C17H20N4O6|CID 493570–PubChem. Available online: https://pubchem.ncbi.nlm.nih.gov/compound/Riboflavin (accessed on 10 March 2024).
- Lumichrome|C12H10N4O2|CID 5326566–PubChem. Available online: https://pubchem.ncbi.nlm.nih.gov/compound/5326566 (accessed on 10 March 2024).
- Fujimura, M.; Yamamoto, S.; Murata, T.; Yasujima, T.; Inoue, K.; Ohta, K.-Y.; Yuasa, H. Functional Characteristics of the Human Ortholog of Riboflavin Transporter 2 and Riboflavin-Responsive Expression of Its Rat Ortholog in the Small Intestine Indicate Its Involvement in Riboflavin Absorption. J. Nutr. 2010, 140, 1722–1727. [Google Scholar] [CrossRef]
- Kubo, Y.; Miki, S.; Akanuma, S.i.; Hosoya, K. ichi Riboflavin Transport Mediated by Riboflavin Transporters (RFVTs/SLC52A) at the Rat Outer Blood-Retinal Barrier. Drug Metab. Pharmacokinet. 2019, 34, 380–386. [Google Scholar] [CrossRef]
- Patel, M.; Vadlapatla, R.K.; Pal, D.; Mitra, A.K. Molecular and Functional Characterization of Riboflavin Specific Transport Sys-tem in Rat Brain Capillary Endothelial Cells. Brain Res. 2012, 1468, 1–10. [Google Scholar] [CrossRef]
- Dallas, S.; Salphati, L.; Gomez-Zepeda, D.; Wanek, T.; Chen, L.; Chu, X.; Kunta, J.; Mezler, M.; Menet, M.-C.; Chasseigneaux, S.; et al. Generation and Characterization of a Breast Cancer Resistance Protein Humanized Mouse Model. Mol. Pharmacol. 2016, 89, 492–504. [Google Scholar] [CrossRef]
- Agarwal, S.; Uchida, Y.; Mittapalli, R.K.; Sane, R.; Terasaki, T.; Elmquist, W.F. Quantitative Proteomics of Transporter Expres-sion in Brain Capillary Endothelial Cells Isolated from P-Glycoprotein (P-Gp), Breast Cancer Resistance Protein (Bcrp), and P-Gp/Bcrp Knockout Mice. Drug Metab. Dispos. 2012, 40, 1164–1169. [Google Scholar] [CrossRef]
- Lagas, J.S.; van Waterschoot, R.A.; Sparidans, R.W.; Wagenaar, E.; Beijnen, J.H.; Schinkel, A.H. Breast Cancer Resistance Protein and P-glycoprotein Limit Sorafenib Brain Accumulation. Mol. Cancer Ther. 2010, 9, 319–326. [Google Scholar] [CrossRef]
- Jonker, J.W.; Brinkhuis, R.F.; Maliepaard, M.; Beijnen, J.H.; Schellens, J.H.M.; Schinkel, A.H. Role of Breast Cancer Resistance Protein in the Bioavailability and Fetal Penetration of Topotecan. JNCI J. Natl. Cancer Inst. 2000, 92, 1651–1656. [Google Scholar] [CrossRef] [PubMed]
- Pavek, P.; Merino, G.; Wagenaar, E.; Bolscher, E.; Novotna, M.; Jonker, J.W.; Schinkel, A.H. Human Breast Cancer Resistance Protein: Interactions with Steroid Drugs, Hormones, the Dietary Carcinogen Transport of Cimetidine. J. Pharmacol. Exp. Ther. 2005, 312, 144–152. [Google Scholar] [CrossRef] [PubMed]
- González-Lobato, L.; Real, R.; Herrero, D.; de la Fuente, A.; Prieto, J.G.; Marqués, M.M.; Álvarez, A.I.; Merino, G. Novel in Vitro Systems for Prediction of Veterinary Drug Residues in Ovine Milk and Dairy Products. Food Addit. Contam.–Part A 2014, 31, 1026–1037. [Google Scholar] [CrossRef] [PubMed]
- Merino, G.; Jonker, J.W.; Wagenaar, E.; van Herwaarden, A.E.; Schinkel, A.H. The Breast Cancer Resistance Protein (BCRP/ABCG2) Affects Pharmacokinetics, Hepatobiliary Excretion, and Milk Secretion of the Antibiotic Nitrofurantoin. Mol. Pharmacol. 2005, 67, 1758–1764. [Google Scholar] [CrossRef]
- Mahnke, H.; Ballent, M.; Baumann, S.; Imperiale, F.; von Bergen, M.; Lanusse, C.; Lifschitz, A.L.; Honscha, W.; Halwachs, S. The ABCG2 Efflux Transporter in the Mammary Gland Mediates Veterinary Drug Secretion across the Blood-Milk Barrier into Milk of Dairy Cows. Drug Metab. Dispos. 2016, 44, 700–708. [Google Scholar] [CrossRef]
- Jonker, J.W.; Buitelaar, M.; Wagenaar, E.; van der Valk, M.A.; Scheffer, G.L.; Scheper, R.J.; Plösch, T.; Kuipers, F.; Elferink, R.P.J.O.; Rosing, H.; et al. The breast cancer resistance protein protects against a major chlorophyll-derived dietary phototoxin and protoporphyria. Proc. Natl. Acad. Sci. USA 2002, 99, 15649–15654. [Google Scholar] [CrossRef]
- Taverniers, I.; De Loose, M.; Van Bockstaele, E. Trends in Quality in the Analytical Laboratory. II. Analytical Method Validation and Quality Assurance. TrAC Trends Anal. Chem. 2004, 23, 535–552. [Google Scholar] [CrossRef]




{kind=link}
{kind=link}
{kind=link}
{kind=link}
| −Ko143 | +Ko143 | |
|---|---|---|
| MDCK-II | 0.85 ± 0.13 | 0.95 ± 0.03 |
| MDCK-II mAbcg2 | 13.81 ± 3.25 * | 1.03 ± 0.04 |
| MDCK-II hABCG2 | 2.57 ± 0.25 * | 1.08 ± 0.08 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Millán-García, A.; Álvarez-Fernández, L.; Blanco-Paniagua, E.; Álvarez, A.I.; Merino, G. The ABCG2 Transporter Affects Plasma Levels, Tissue Distribution and Milk Secretion of Lumichrome, a Natural Derivative of Riboflavin. Int. J. Mol. Sci. 2024, 25, 9884. https://doi.org/10.3390/ijms25189884
Millán-García A, Álvarez-Fernández L, Blanco-Paniagua E, Álvarez AI, Merino G. The ABCG2 Transporter Affects Plasma Levels, Tissue Distribution and Milk Secretion of Lumichrome, a Natural Derivative of Riboflavin. International Journal of Molecular Sciences. 2024; 25(18):9884. https://doi.org/10.3390/ijms25189884
Chicago/Turabian StyleMillán-García, Alicia, Laura Álvarez-Fernández, Esther Blanco-Paniagua, Ana I. Álvarez, and Gracia Merino. 2024. "The ABCG2 Transporter Affects Plasma Levels, Tissue Distribution and Milk Secretion of Lumichrome, a Natural Derivative of Riboflavin" International Journal of Molecular Sciences 25, no. 18: 9884. https://doi.org/10.3390/ijms25189884