
Biophysical Studies of Amyloid-Binding Fluorophores to Tau AD Core Fibrils Formed without Cofactors




Abstract
1. Introduction
2. Results
2.1. TEM Analysis of Cofactor-Free Tau AD Core (TADC, Tau306–378) Aggregates
2.2. Amyloid-Binding Fluorescent Probes Show Different Sensitivities to TADC Aggregates
2.3. Cofactor-Free Aggregation of TADC Can Be Monitored by Selective Amyloid-Binding Probes
3. Discussion
4. Materials and Methods
4.1. Materials
4.2. Protein Expression and Purification
4.3. Transmission Electron Microscopy
4.4. Fluorescence Measurements
4.5. Fluorescence-Monitored Aggregation Assays
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Zhang, Y.; Wu, K.-M.; Yang, L.; Dong, Q.; Yu, J.-T. Tauopathies: New Perspectives and Challenges. Mol. Neurodegener. 2022, 17, 28. [Google Scholar] [CrossRef] [PubMed]
- Creekmore, B.C.; Watanabe, R.; Lee, E.B. Neurodegenerative Disease Tauopathies. Annu. Rev. Pathol. 2024, 19, 345–370. [Google Scholar] [CrossRef] [PubMed]
- Jeganathan, S.; von Bergen, M.; Brutlach, H.; Steinhoff, H.-J.; Mandelkow, E. Global Hairpin Folding of Tau in Solution. Biochemistry 2006, 45, 2283–2293. [Google Scholar] [CrossRef] [PubMed]
- Wang, Y.; Mandelkow, E. Tau in Physiology and Pathology. Nat. Rev. Neurosci. 2016, 17, 22–35. [Google Scholar] [CrossRef]
- Limorenko, G.; Lashuel, H.A. Revisiting the Grammar of Tau Aggregation and Pathology Formation: How New Insights from Brain Pathology Are Shaping How We Study and Target Tauopathies. Chem. Soc. Rev. 2022, 51, 513–565. [Google Scholar] [CrossRef]
- Goedert, M.; Jakes, R.; Spillantini, M.G.; Hasegawa, M.; Smith, M.J.; Crowther, R.A. Assembly of Microtubule-Associated Protein Tau into Alzheimer-like Filaments Induced by Sulphated Glycosaminoglycans. Nature 1996, 383, 550–553. [Google Scholar] [CrossRef]
- Kampers, T.; Friedhoff, P.; Biernat, J.; Mandelkow, E.-M.; Mandelkow, E. RNA Stimulates Aggregation of Microtubule-Associated Protein Tau into Alzheimer-like Paired Helical Filaments. FEBS Lett. 1996, 399, 344–349. [Google Scholar] [CrossRef]
- Wilson, D.M.; Binder, L.I. Free Fatty Acids Stimulate the Polymerization of Tau and Amyloid Beta Peptides. In Vitro Evidence for a Common Effector of Pathogenesis in Alzheimer’s Disease. Am. J. Pathol. 1997, 150, 2181–2195. [Google Scholar]
- Fichou, Y.; Vigers, M.; Goring, A.K.; Eschmann, N.A.; Han, S. Heparin-Induced Tau Filaments Are Structurally Heterogeneous and Differ from Alzheimer’s Disease Filaments. Chem. Commun. 2018, 54, 4573–4576. [Google Scholar] [CrossRef]
- Zhang, W.; Falcon, B.; Murzin, A.G.; Fan, J.; Crowther, R.A.; Goedert, M.; Scheres, S.H. Heparin-Induced Tau Filaments Are Polymorphic and Differ from Those in Alzheimer’s and Pick’s Diseases. eLife 2019, 8, e43584. [Google Scholar] [CrossRef]
- Aliyan, A.; Cook, N.P.; Martí, A.A. Interrogating Amyloid Aggregates Using Fluorescent Probes. Chem. Rev. 2019, 119, 11819–11856. [Google Scholar] [CrossRef] [PubMed]
- Louros, N.; Ramakers, M.; Michiels, E.; Konstantoulea, K.; Morelli, C.; Garcia, T.; Moonen, N.; D’Haeyer, S.; Goossens, V.; Thal, D.R.; et al. Mapping the Sequence Specificity of Heterotypic Amyloid Interactions Enables the Identification of Aggregation Modifiers. Nat. Commun. 2022, 13, 1351. [Google Scholar] [CrossRef] [PubMed]
- Du, J.-Q.; Luo, W.-C.; Zhang, J.-T.; Li, Q.-Y.; Bao, L.-N.; Jiang, M.; Yu, X.; Xu, L. Machine Learning-Assisted Fluorescence/Fluorescence Colorimetric Sensor Array for Discriminating Amyloid Fibrils. Sens. Actuators B Chem. 2024, 417, 136173. [Google Scholar] [CrossRef]
- Rodriguez Camargo, D.C.; Sileikis, E.; Chia, S.; Axell, E.; Bernfur, K.; Cataldi, R.L.; Cohen, S.I.A.; Meisl, G.; Habchi, J.; Knowles, T.P.J.; et al. Proliferation of Tau 304–380 Fragment Aggregates through Autocatalytic Secondary Nucleation. ACS Chem. Neurosci. 2021, 12, 4406–4415. [Google Scholar] [CrossRef]
- Fitzpatrick, A.W.P.; Falcon, B.; He, S.; Murzin, A.G.; Murshudov, G.; Garringer, H.J.; Crowther, R.A.; Ghetti, B.; Goedert, M.; Scheres, S.H.W. Cryo-EM Structures of Tau Filaments from Alzheimer’s Disease. Nature 2017, 547, 185–190. [Google Scholar] [CrossRef]
- Chakraborty, P.; Rivière, G.; Liu, S.; de Opakua, A.I.; Dervişoğlu, R.; Hebestreit, A.; Andreas, L.B.; Vorberg, I.M.; Zweckstetter, M. Co-Factor-Free Aggregation of Tau into Seeding-Competent RNA-Sequestering Amyloid Fibrils. Nat. Commun. 2021, 12, 4231. [Google Scholar] [CrossRef] [PubMed]
- Carlomagno, Y.; Manne, S.; DeTure, M.; Prudencio, M.; Zhang, Y.-J.; Hanna Al-Shaikh, R.; Dunmore, J.A.; Daughrity, L.M.; Song, Y.; Castanedes-Casey, M.; et al. The AD Tau Core Spontaneously Self-Assembles and Recruits Full-Length Tau to Filaments. Cell Rep. 2021, 34, 108843. [Google Scholar] [CrossRef] [PubMed]
- Fagnen, C.; Giovannini, J.; Catto, M.; Voisin-Chiret, A.S.; Sopkova-de Oliveira Santos, J. On the Tracks of the Aggregation Mechanism of the PHF6 Peptide from Tau Protein: Molecular Dynamics, Energy, and Interaction Network Investigations. ACS Chem. Neurosci. 2022, 13, 2874–2887. [Google Scholar] [CrossRef]
- Ganguly, P.; Do, T.D.; Larini, L.; LaPointe, N.E.; Sercel, A.J.; Shade, M.F.; Feinstein, S.C.; Bowers, M.T.; Shea, J.-E. Tau Assembly: The Dominant Role of PHF6 (VQIVYK) in Microtubule Binding Region Repeat R3. J. Phys. Chem. B 2015, 119, 4582–4593. [Google Scholar] [CrossRef]
- El Mammeri, N.; Dregni, A.J.; Duan, P.; Wang, H.K.; Hong, M. Microtubule-Binding Core of the Tau Protein. Sci. Adv. 2022, 8, eabo4459. [Google Scholar] [CrossRef]
- Arosio, P.; Knowles, T.P.J.; Linse, S. On the Lag Phase in Amyloid Fibril Formation. Phys. Chem. Chem. Phys. 2015, 17, 7606–7618. [Google Scholar] [CrossRef] [PubMed]
- Wegmann, S.; Jung, Y.J.; Chinnathambi, S.; Mandelkow, E.-M.; Mandelkow, E.; Muller, D.J. Human Tau Isoforms Assemble into Ribbon-like Fibrils That Display Polymorphic Structure and Stability*. J. Biol. Chem. 2010, 285, 27302–27313. [Google Scholar] [CrossRef]
- Crowther, R.A. Straight and Paired Helical Filaments in Alzheimer Disease Have a Common Structural Unit. Proc. Natl. Acad. Sci. USA 1991, 88, 2288–2292. [Google Scholar] [CrossRef]
- Kjaergaard, M.; Dear, A.J.; Kundel, F.; Qamar, S.; Meisl, G.; Knowles, T.P.J.; Klenerman, D. Oligomer Diversity during the Aggregation of the Repeat Region of Tau. ACS Chem. Neurosci. 2018, 9, 3060–3071. [Google Scholar] [CrossRef]
- Hammarström, P.; Simon, R.; Nyström, S.; Konradsson, P.; Åslund, A.; Nilsson, K.P.R. A Fluorescent Pentameric Thiophene Derivative Detects in Vitro-Formed Prefibrillar Protein Aggregates. Biochemistry 2010, 49, 6838–6845. [Google Scholar] [CrossRef]
- Lövestam, S.; Li, D.; Wagstaff, J.L.; Kotecha, A.; Kimanius, D.; McLaughlin, S.H.; Murzin, A.G.; Freund, S.M.V.; Goedert, M.; Scheres, S.H.W. Disease-Specific Tau Filaments Assemble via Polymorphic Intermediates. Nature 2024, 625, 119–125. [Google Scholar] [CrossRef] [PubMed]
- Figueira, A.J.; Saavedra, J.; Cardoso, I.; Gomes, C.M. S100B Chaperone Multimers Suppress the Formation of Oligomers during Aβ42 Aggregation. Front. Neurosci. 2023, 17, 1162741. [Google Scholar] [CrossRef] [PubMed]
- Hawe, A.; Sutter, M.; Jiskoot, W. Extrinsic Fluorescent Dyes as Tools for Protein Characterization. Pharm. Res. 2008, 25, 1487–1499. [Google Scholar] [CrossRef]
- Gade Malmos, K.; Blancas-Mejia, L.M.; Weber, B.; Buchner, J.; Ramirez-Alvarado, M.; Naiki, H.; Otzen, D. ThT 101: A Primer on the Use of Thioflavin T to Investigate Amyloid Formation. Amyloid 2017, 24, 1–16. [Google Scholar] [CrossRef]
- Amdursky, N.; Erez, Y.; Huppert, D. Molecular Rotors: What Lies Behind the High Sensitivity of the Thioflavin-T Fluorescent Marker. Acc. Chem. Res. 2012, 45, 1548–1557. [Google Scholar] [CrossRef]
- Biancalana, M.; Koide, S. Molecular Mechanism of Thioflavin-T Binding to Amyloid Fibrils. Biochim. Biophys. Acta 2010, 1804, 1405–1412. [Google Scholar] [CrossRef]
- Sabaté, R.; Lascu, I.; Saupe, S.J. On the Binding of Thioflavin-T to HET-s Amyloid Fibrils Assembled at pH 2. J. Struct. Biol. 2008, 162, 387–396. [Google Scholar] [CrossRef]
- Cloe, A.L.; Orgel, J.P.R.O.; Sachleben, J.R.; Tycko, R.; Meredith, S.C. The Japanese Mutant Aβ (ΔE22-Aβ1−39) Forms Fibrils Instantaneously, with Low-Thioflavin T Fluorescence: Seeding of Wild-Type Aβ1−40 into Atypical Fibrils by ΔE22-Aβ1−39. Biochemistry 2011, 50, 2026–2039. [Google Scholar] [CrossRef]
- Rosen, C.G.; Weber, G. Dimer Formation from 1-Anilino-8-Naphthalenesulfonate Catalyzed by Bovine Serum Albumin. Fluorescent Molecule with Exceptional Binding Properties. Biochemistry 1969, 8, 3915–3920. [Google Scholar] [CrossRef]
- Younan, N.D.; Viles, J.H. A Comparison of Three Fluorophores for the Detection of Amyloid Fibers and Prefibrillar Oligomeric Assemblies. ThT (Thioflavin T); ANS (1-Anilinonaphthalene-8-Sulfonic Acid); and bisANS (4,4’-Dianilino-1,1’-Binaphthyl-5,5’-Disulfonic Acid). Biochemistry 2015, 54, 4297–4306. [Google Scholar] [CrossRef]
- Reinke, A.A.; Abulwerdi, G.A.; Gestwicki, J.E. Quantifying Prefibrillar Amyloids in Vitro by Using a “Thioflavin-Like” Spectroscopic Method. Chembiochem 2010, 11, 1889–1895. [Google Scholar] [CrossRef]
- Zhang, J.; Sandberg, A.; Konsmo, A.; Wu, X.; Nyström, S.; Nilsson, K.P.R.; Konradsson, P.; LeVine III, H.; Lindgren, M.; Hammarström, P. Detection and Imaging of Aβ1-42 and Tau Fibrils by Redesigned Fluorescent X-34 Analogues. Chem.-A Eur. J. 2018, 24, 7210–7216. [Google Scholar] [CrossRef]
- Sipe, J.D.; Benson, M.D.; Buxbaum, J.N.; Ikeda, S.; Merlini, G.; Saraiva, M.J.M.; Westermark, P. Amyloid Fibril Proteins and Amyloidosis: Chemical Identification and Clinical Classification International Society of Amyloidosis 2016 Nomenclature Guidelines. Amyloid 2016, 23, 209–213. [Google Scholar] [CrossRef]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Fluorophore | Structure | λex (nm) | λem Range (nm) |
|---|---|---|---|
| ThT |  | 440 | 450–640 |
| ThS |  | 440 | 450–640 |
| pFTAA |  | 440 | 450–640 |
| hFTAA |  | 440 | 450–640 |
| ANS |  | 375 | 390–630 |
| Bis-ANS |  | 395 | 410–650 |
| X-34 |  | 370 | 390–630 |
| Congo Red | 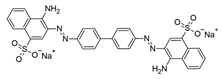 | 497 | 550–700 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Freitas, D.P.; Saavedra, J.; Cardoso, I.; Gomes, C.M. Biophysical Studies of Amyloid-Binding Fluorophores to Tau AD Core Fibrils Formed without Cofactors. Int. J. Mol. Sci. 2024, 25, 9946. https://doi.org/10.3390/ijms25189946
Freitas DP, Saavedra J, Cardoso I, Gomes CM. Biophysical Studies of Amyloid-Binding Fluorophores to Tau AD Core Fibrils Formed without Cofactors. International Journal of Molecular Sciences. 2024; 25(18):9946. https://doi.org/10.3390/ijms25189946
Chicago/Turabian StyleFreitas, Daniela P., Joana Saavedra, Isabel Cardoso, and Cláudio M. Gomes. 2024. "Biophysical Studies of Amyloid-Binding Fluorophores to Tau AD Core Fibrils Formed without Cofactors" International Journal of Molecular Sciences 25, no. 18: 9946. https://doi.org/10.3390/ijms25189946
APA StyleFreitas, D. P., Saavedra, J., Cardoso, I., & Gomes, C. M. (2024). Biophysical Studies of Amyloid-Binding Fluorophores to Tau AD Core Fibrils Formed without Cofactors. International Journal of Molecular Sciences, 25(18), 9946. https://doi.org/10.3390/ijms25189946