
Molecular Characterization of the MoxR AAA+ ATPase of Synechococcus sp. Strain NKBG15041c

 ,
,  and
and
Abstract
1. Introduction
2. Results
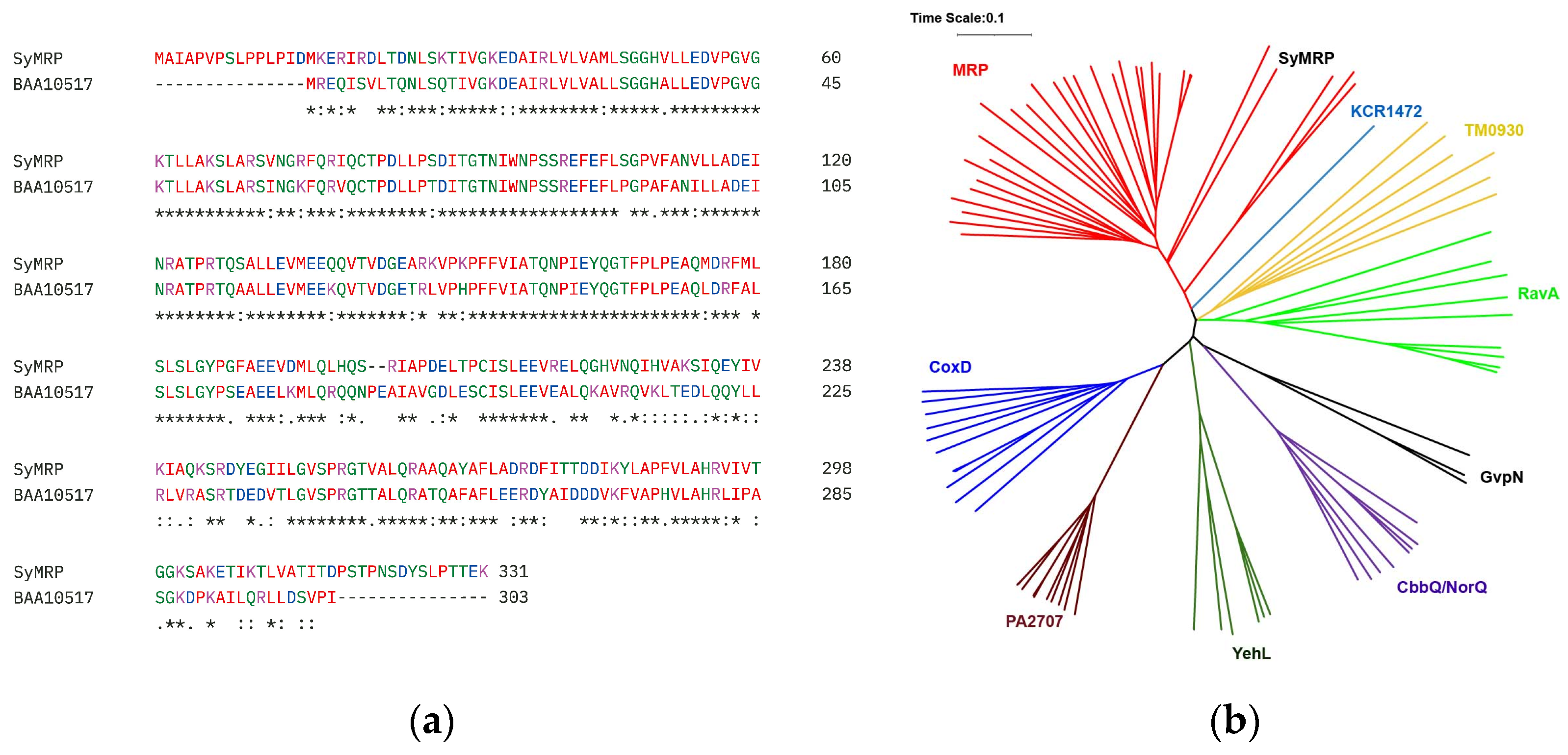
2.1. Screening of a Gene Responsible for the Stress Tolerance of Synechococcus sp. Strain NKBG15041c
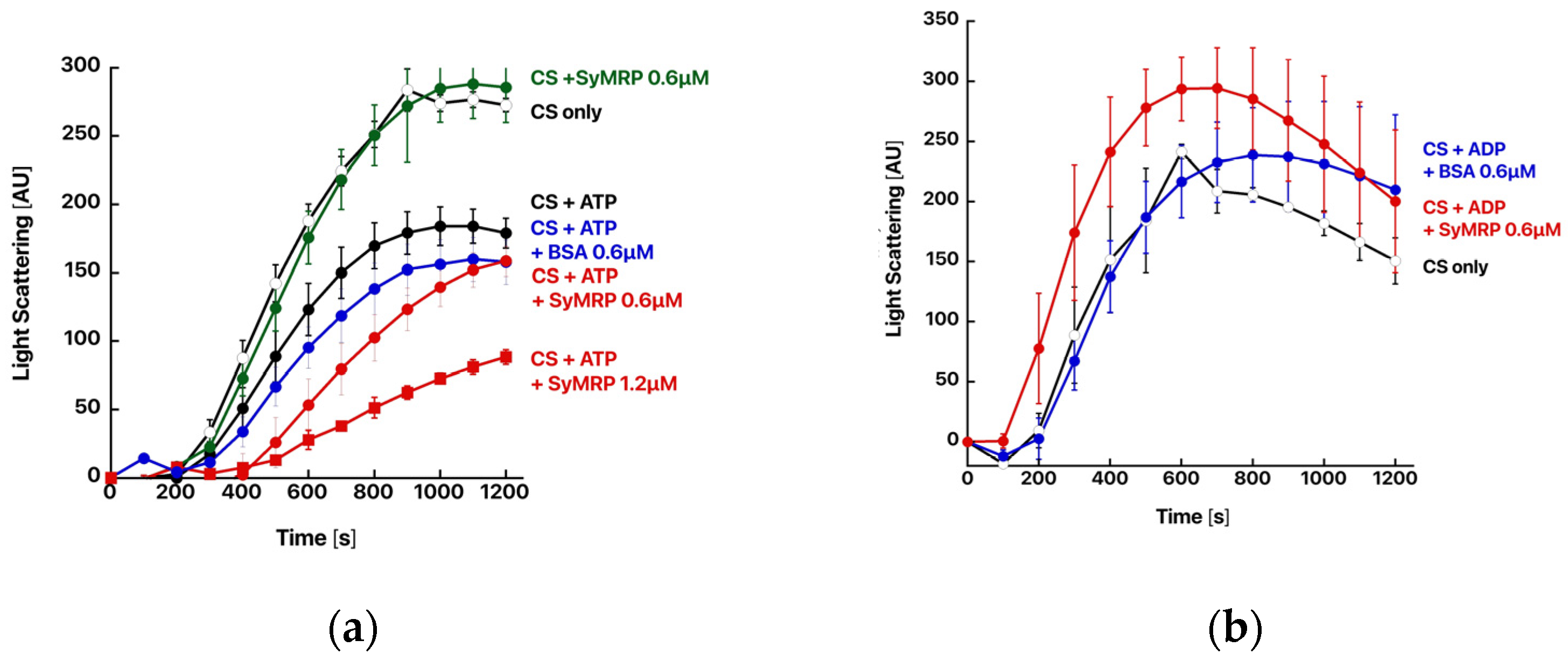
2.2. Expression and Functional Characterization of SyMRP
2.3. Oligomeric Structure of SyMRP
3. Discussion
4. Materials and Methods
4.1. Strains and Media
4.2. Genomic DNA Library Construction
4.3. Evaluation of Stress Tolerance in E. coli Expressing SyMRP and BAA10517
4.4. Expression and Purification of SyMRP
4.5. ATPase Activity
4.6. Protein Aggregation Measurements
4.7. Size Exclusion Chromatography on HPLC
4.8. Analytical Ultracentrifugation (AUC) Measurements
4.9. Small-Angle X-ray Scattering (SAXS) Measurements
4.10. Molecular Modeling
4.11. AFM Observation
4.12. Electron Microscopy
Supplementary Materials
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Ayarpadikannan, S.; Chung, E.; Cho, C.W.; So, H.A.; Kim, S.O.; Jeon, J.M.; Kwak, M.H.; Lee, S.W.; Lee, J.H. Exploration for the salt stress tolerance genes from a salt-treated halophyte, Suaeda asparagoides. Plant Cell Rep. 2012, 31, 35–48. [Google Scholar] [CrossRef] [PubMed]
- Ellis, J. Proteins as molecular chaperones. Nature 1987, 328, 378–379. [Google Scholar] [CrossRef]
- Gething, M.J.; Sambrook, J. Protein folding in the cell. Nature 1992, 355, 33–45. [Google Scholar] [CrossRef]
- Sode, K.; Tatara, M.; Takeyama, H.; Burgess, J.G.; Matsunaga, T. Conjugative gene transfer in marine cyanobacteria: Synechococcus sp., Synechocystis sp. and Pseudanabaena sp. Appl. Microbiol. Biotechnol. 1992, 37, 369–373. [Google Scholar] [CrossRef] [PubMed]
- Yoshino, T.; Honda, T.; Tanaka, M.; Tanaka, T. Draft Genome Sequence of Marine Cyanobacterium Synechococcus sp. Strain NKBG15041c. Genome Announc. 2013, 1, e00954-13. [Google Scholar] [CrossRef] [PubMed]
- Yamada, A.; Sekiguchi, M.; Mimura, T.; Ozeki, Y. The role of plant CCTalpha in salt- and osmotic-stress tolerance. Plant Cell Physiol. 2002, 43, 1043–1048. [Google Scholar] [CrossRef]
- Bhandari, V.; Van Ommen, D.A.J.; Wong, K.S.; Houry, W.A. Analysis of the Evolution of the MoxR ATPases. J. Phys. Chem. A 2022, 126, 4734–4746. [Google Scholar] [CrossRef]
- Tsai, Y.C.; Liew, L.; Guo, Z.; Liu, D.; Mueller-Cajar, O. The CbbQO-type rubisco activases encoded in carboxysome gene clusters can activate carboxysomal form IA rubiscos. J. Biol. Chem. 2022, 298, 101476. [Google Scholar] [CrossRef]
- Bhat, J.Y.; Thieulin-Pardo, G.; Hartl, F.U.; Hayer-Hartl, M. Rubisco Activases: AAA+ Chaperones Adapted to Enzyme Repair. Front. Mol. Biosci. 2017, 4, 20. [Google Scholar] [CrossRef]
- Wong, K.S.; Bhandari, V.; Janga, S.C.; Houry, W.A. The RavA-ViaA Chaperone-like System Interacts with and Modulates the Activity of the Fumarate Reductase Respiratory Complex. J. Mol. Biol. 2017, 429, 324–344. [Google Scholar] [CrossRef]
- Bhandari, V.; Reichheld, S.E.; Houliston, S.; Lemak, A.; Arrowsmith, C.H.; Sharpe, S.; Houry, W.A. The RavA-ViaA chaperone complex modulates bacterial persistence through its association with the fumarate reductase enzyme. J. Biol. Chem. 2023, 299, 105199. [Google Scholar] [CrossRef] [PubMed]
- Dieppedale, J.; Sobral, D.; Dupuis, M.; Dubail, I.; Klimentova, J.; Stulik, J.; Postic, G.; Frapy, E.; Meibom, K.L.; Barel, M.; et al. Identification of a putative chaperone involved in stress resistance and virulence in Francisella tularensis. Infect. Immun. 2011, 79, 1428–1439. [Google Scholar] [CrossRef] [PubMed]
- Mironov, K.S.; Sinetova, M.A.; Shumskaya, M.; Los, D.A. Universal Molecular Triggers of Stress Responses in Cyanobacterium Synechocystis. Life 2019, 9, 67. [Google Scholar] [CrossRef] [PubMed]
- Cerveny, J.; Sinetova, M.A.; Zavrel, T.; Los, D.A. Mechanisms of High Temperature Resistance of Synechocystis sp. PCC 6803: An Impact of Histidine Kinase 34. Life 2015, 5, 676–699. [Google Scholar] [CrossRef]
- Nishizawa, M.; Walinda, E.; Morimoto, D.; Kohn, B.; Scheler, U.; Shirakawa, M.; Sugase, K. Effects of Weak Nonspecific Interactions with ATP on Proteins. J. Am. Chem. Soc. 2021, 143, 11982–11993. [Google Scholar] [CrossRef] [PubMed]
- Putnam, C.D.; Hammel, M.; Hura, G.L.; Tainer, J.A. X-ray solution scattering (SAXS) combined with crystallography and computation: Defining accurate macromolecular structures, conformations and assemblies in solution. Q. Rev. Biophys. 2007, 40, 191–285. [Google Scholar] [CrossRef] [PubMed]
- Hura, G.L.; Menon, A.L.; Hammel, M.; Rambo, R.P.; Poole, F.L., 2nd; Tsutakawa, S.E.; Jenney, F.E., Jr.; Classen, S.; Frankel, K.A.; Hopkins, R.C.; et al. Robust, high-throughput solution structural analyses by small angle X-ray scattering (SAXS). Nat. Methods 2009, 6, 606–612. [Google Scholar] [CrossRef]
- Snider, J.; Houry, W.A. MoxR AAA+ ATPases: A novel family of molecular chaperones? J. Struct. Biol. 2006, 156, 200–209. [Google Scholar] [CrossRef]
- Kahle, M.; Appelgren, S.; Elofsson, A.; Carroni, M.; Adelroth, P. Insights into the structure-function relationship of the NorQ/NorD chaperones from Paracoccus denitrificans reveal shared principles of interacting MoxR AAA+/VWA domain proteins. BMC Biol. 2023, 21, 47. [Google Scholar] [CrossRef]
- Wang, J.; Huang, X.; Ge, H.; Wang, Y.; Chen, W.; Zheng, L.; Huang, C.; Yang, H.; Li, L.; Sui, N.; et al. The quantitative proteome atlas of a model cyanobacterium. J. Genet. Genom. 2022, 49, 96–108. [Google Scholar] [CrossRef]
- Schuck, P. Size-distribution analysis of macromolecules by sedimentation velocity ultracentrifugation and lamm equation modeling. Biophys. J. 2000, 78, 1606–1619. [Google Scholar] [CrossRef] [PubMed]
- Shimizu, N.; Yatabe, K.; Nagatani, Y.; Saijyo, S.; Kosuge, T.; Igarashi, N. Software development for analysis of small-angle X-ray scattering data. AIP Conf. Proc. 2016, 1741, 050017. [Google Scholar] [CrossRef]
- Abramson, J.; Adler, J.; Dunger, J.; Evans, R.; Green, T.; Pritzel, A.; Ronneberger, O.; Willmore, L.; Ballard, A.J.; Bambrick, J.; et al. Accurate structure prediction of biomolecular interactions with AlphaFold 3. Nature 2024, 630, 493–500. [Google Scholar] [CrossRef] [PubMed]
- Franke, D.; Petoukhov, M.V.; Konarev, P.V.; Panjkovich, A.; Tuukkanen, A.; Mertens, H.D.T.; Kikhney, A.G.; Hajizadeh, N.R.; Franklin, J.M.; Jeffries, C.M.; et al. ATSAS 2.8: A comprehensive data analysis suite for small-angle scattering from macromolecular solutions. J. Appl. Crystallogr. 2017, 50 Pt 4, 1212–1225. [Google Scholar] [CrossRef] [PubMed]
- Ando, T.; Kodera, N.; Takai, E.; Maruyama, D.; Saito, K.; Toda, A. A high-speed atomic force microscope for studying biological macromolecules. Proc. Natl. Acad. Sci. USA 2001, 98, 12468–12472. [Google Scholar] [CrossRef]
- Uchihashi, T.; Watanabe, Y.H.; Nakazaki, Y.; Yamasaki, T.; Watanabe, H.; Maruno, T.; Ishii, K.; Uchiyama, S.; Song, C.; Murata, K.; et al. Dynamic structural states of ClpB involved in its disaggregation function. Nat. Commun. 2018, 9, 2147. [Google Scholar] [CrossRef]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| s20,w/S | M/kDa | w/% | |
|---|---|---|---|
| Without ATP | 4.18 | 90.4 | 4.0 |
| 7.60 | 221 | 91.4 | |
| 9.87 | 327 | 4.6 | |
| With ATP | 4.26 | 93.2 | 2.6 |
| 7.53 | 218 | 90.9 | |
| 9.72 | 320 | 6.5 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Mano, K.; Noi, K.; Oe, K.; Mochizuki, T.; Morishima, K.; Inoue, R.; Sugiyama, M.; Noguchi, K.; Shinohara, K.; Yohda, M.; et al. Molecular Characterization of the MoxR AAA+ ATPase of Synechococcus sp. Strain NKBG15041c. Int. J. Mol. Sci. 2024, 25, 9955. https://doi.org/10.3390/ijms25189955
Mano K, Noi K, Oe K, Mochizuki T, Morishima K, Inoue R, Sugiyama M, Noguchi K, Shinohara K, Yohda M, et al. Molecular Characterization of the MoxR AAA+ ATPase of Synechococcus sp. Strain NKBG15041c. International Journal of Molecular Sciences. 2024; 25(18):9955. https://doi.org/10.3390/ijms25189955
Chicago/Turabian StyleMano, Kota, Kentaro Noi, Kumiko Oe, Takahiro Mochizuki, Ken Morishima, Rintaro Inoue, Masaaki Sugiyama, Keiichi Noguchi, Kyosuke Shinohara, Masafumi Yohda, and et al. 2024. "Molecular Characterization of the MoxR AAA+ ATPase of Synechococcus sp. Strain NKBG15041c" International Journal of Molecular Sciences 25, no. 18: 9955. https://doi.org/10.3390/ijms25189955
APA StyleMano, K., Noi, K., Oe, K., Mochizuki, T., Morishima, K., Inoue, R., Sugiyama, M., Noguchi, K., Shinohara, K., Yohda, M., & Yamada, A. (2024). Molecular Characterization of the MoxR AAA+ ATPase of Synechococcus sp. Strain NKBG15041c. International Journal of Molecular Sciences, 25(18), 9955. https://doi.org/10.3390/ijms25189955