Abstract
Phenylalanine ammonia-lyase (PAL) acts as the rate-limiting enzyme for anthocyanin biosynthesis through the phenylpropanoid pathway, a crucial component of plant secondary metabolism. The PAL gene family plays a crucial role in plants’ defense and stress responses, but its in silico identification and expression analyses in Brassica oleracea under different abiotic stresses remain unexplored. In this study, nine BolPAL, seven BrPAL, four AtPAL, and seventeen BnPAL genes were obtained from the genomes of B. oleracea, Brassica rapa, Arabidopsis thaliana, and Brassica napus, respectively. Segmental duplication and purifying selection are the causes of the BolPAL gene’s amplification and evolution. The BolPAL genes with comparable intron–exon architectures and motifs were grouped together in the same clade. Three categories comprised the cis-regulatory elements: abiotic stressors, phytohormones, and light. According to the results of the qRT-PCR experiments, the majority of the BolPAL genes were expressed highly under MeJA, a low temperature, and a high temperature, and they were downregulated under ABA. Under white light (100 µmol m−2 s−1) with 50, 100, or 150 µmol m−2 s−1 far-red (FR), only a small number of the PAL genes were expressed at 50 and 100 µmol m−2 s−1 FR, while the majority of the PAL genes were slightly elevated at 150 µmol m−2 s−1 FR. This work offers a theoretical foundation for molecular breeding research to investigate the role of BolPAL genes and their role in anthocyanin biosynthesis.
1. Introduction
Phenylalanine ammonia-lyase (PAL) controls the rate at which phenylalanine enters the phenylpropanoid metabolic pathway, which in turn influences the formation of the secondary metabolites in plants, including flavonoids, lignin, and hydroxycinnamic acid amide (HCAA) [1,2]. It is a crucial enzyme that catalyzes the initial stage of the phenylpropanoid pathway and serves as an enzyme to connect the primary and the secondary phenylpropanoid metabolisms [3]. These metabolites are key factors for plant growth, development, and stress tolerance [4]. First discovered in 1961 in barley [3], the PAL gene is typically found in plants as a gene family that spans multiple members in various species, including four members in willow [5]. Nevertheless, there is a lack of extensive understanding regarding the complete distribution of the genes related to phenylalanine ammonia-lyase (PAL) throughout the entire genome of B. oleracea, a highly nutritious crop.
A study showed that the transcription factors CsbHLHv and CsMYB regulate the CsPAL gene, which contributes to anthocyanins synthesis [6]. In a study, it was shown that in the shoots of purple-leaved tea plants, the CsPAL4 gene expressed and exhibited a significant positive correlation with anthocyanin content [6]. A different study showed that in red and white grapes, VvPAL1 and VvPAL5 are involved in the production of anthocyanins [7]. In another study, 14 days after wheat stripe rust struck the studied plants, the plants that had had the TaPAL32 and TaPAL42 genes silenced displayed a more severe illness than the control plants [8]. In a different study, the majority of StPALs in potatoes responded to drought and high temperatures [9]. In another study, AtPAL1 and AtPAL2 were strongly expressed in response to temperature changes and nitrogen stress, which resulted in the formation and accumulation of flavonoids in A. thaliana [10]. In a different study, double-knockout mutants of AtPAL1 and AtPAL2 resulted in an increased resistance to drought stress, an increased susceptibility to ultraviolet-B (UV-B) radiation, and a decreased biosynthesis of flavonoids and anthocyanins [10]. These studies collectively show that plant tolerance to external stressors is thus significantly influenced by PAL genes.
In plants, the PAL proteins are comparatively conserved. Plants differ greatly in the amount of PAL protein they have, but their molecular weight is generally constant, ranging from 275 to 330 kDa [11,12]. The PAL family is usually made up of several PALs, and each member of the family responds differently to biotic and abiotic stressors and has a unique pattern of expression [13]. The PAL gene family has been extensively examined in different crops, including wheat (Triticum aestivum L.) [8], walnut (Juglans regia) [14], rice (Oryza sativa) [15], coleus (Solenostemon scutellarioides (L.) Codd) [13], cottonwood (Populus trichocarpa) [16], and A. thaliana [17]. However, the PAL gene family in B. oleracea and its expression under various abiotic stressors have not been identified yet.
Renowned for its delicate stems and nourishing leaves, Chinese kale (B. oleracea), sometimes referred to as kailaan or gai lan, is a widely used leafy vegetable in Chinese cuisine [18,19]. We performed a genome-wide investigation to identify the BolPALs in B. oleracea’s genome to evaluate the evolutionary relationships, conserved motifs, gene structure, and cis-acting regions in the promoter sequences of BolPALs in order to investigate their structural diversity and evolution. Furthermore, we represented the expression profile of BolPALs that were subjected to various treatments such as abscisic acid (ABA), methyl jasmonate (MeJA), a low temperature, a high temperature, and light with varying far-red capacities. These results will not only help us better understand how BolPALs respond to abiotic stress, but they will also make it easier to study the biological roles of an important gene family and identify possible objectives for productive breeding in B. oleracea crops under a variety of abiotic stressors.
2. Results
2.1. Identification and Characterization of the BolPALs
A total of nine BolPAL genes were recognized from the whole genome of B. oleracea via the Brassica database, http://www.brassicadb.cn/ (accessed on 12 May 2024) [20], as shown in Table 1. The names of these genes are BolPAL1-1, BolPAL1-2, BolPAL2-1, BolPAL2-2, BolPAL2-3, BolPAL2-4, BolPAL3-1, BolPAL3-2, and BolPAL4. Further, we verified the PAL gene-specific lyase aromatic domain (PF00221) in all the identified genes via the NCBI conserved domain database, https://www.ncbi.nlm.nih.gov/Structure/cdd/cdd.shtml (accessed on 12 May 2024) (Figure S1). Detailed information on all the BolPALs is given in Table 1. The coding sequences ranged from 801 bp (BolPAL3-1) to 2172 bp (BolPAL2-2 and BolPAL2-4), and the protein sequences ranged from 267 aa to 724 aa, while the predicted molecular weights varied from 29.8 to 78.6 MW/kDa, and the isoelectric points varied from 5.54 (BolPAL2-3) to 7.71 (BolPAL3-1). During subcellular localization prediction, we found that all the BolPAL proteins were located on the cytoplasm and only BolPAL1-1 was found on the plastid. Moreover, 4, 7, and 17 PAL genes from A. thaliana, B. rapa, and B. napus, respectively, were also identified (Table S2).

Table 1.
Detailed information of PAL genes in B. oleracea.
2.2. Phylogenetic Analysis of the PAL Family Genes
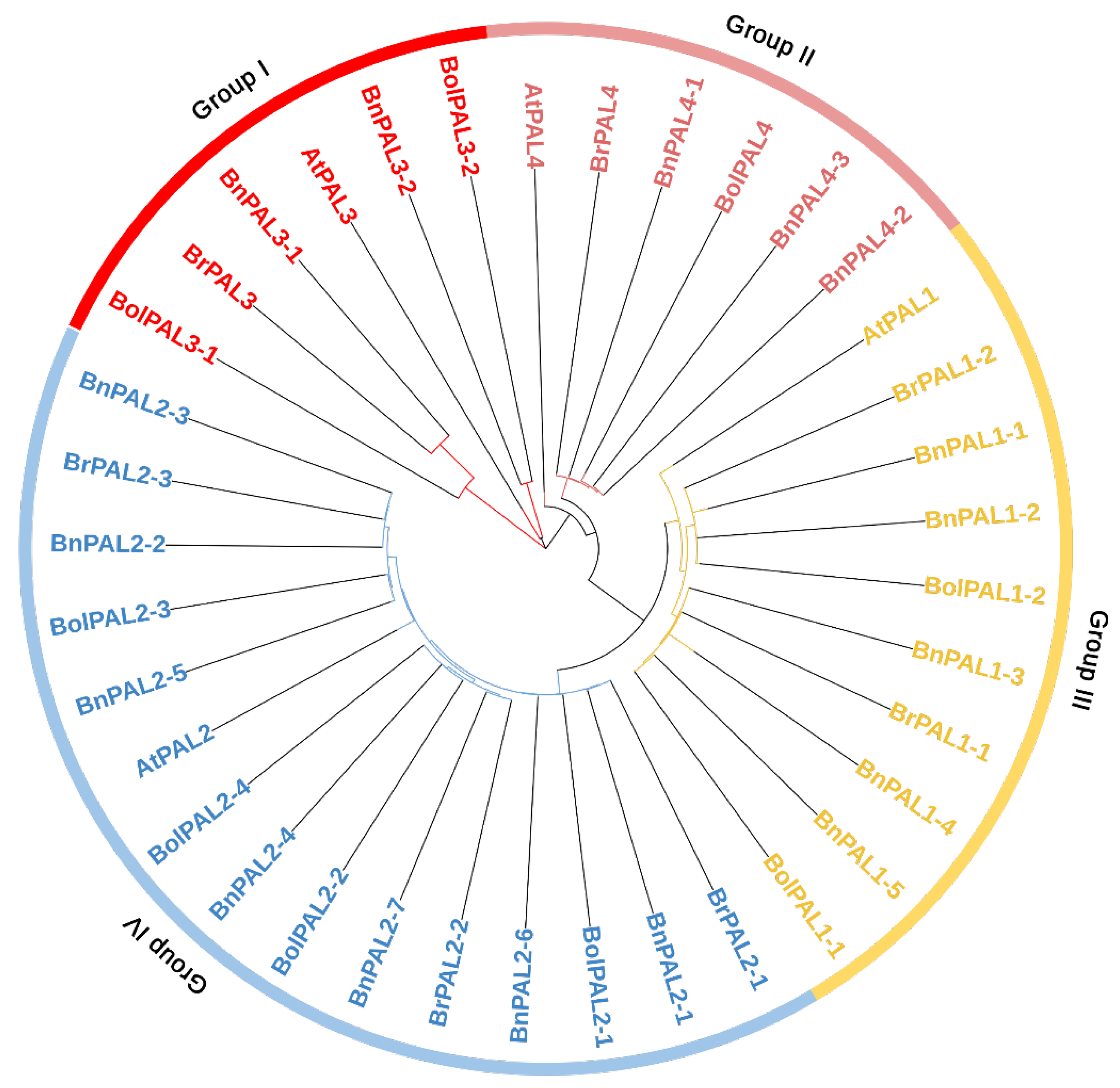
Based on the phylogenetic results, all the PAL genes from B. oleracea, B. rapa, A. thaliana, and B. napus were separated into four major groups (I, II, III, and IV), based on their node distributions, as shown in Figure 1. Group I contained 2 BolPAL (BolPAL3-1 and BolPAL3-2), 1 AtPAL3, 2 BnPAL (BnPAL3-1 and BnPAL3-2), and 1 BrPAL (BrPAL3). Group II consisted of 1 BolPAL (BolPAL4), 1 AtPAL4, 3 BnPAL (BnPAL4-1, BnPAL4-2, and BnPAL4-3), and 1 BrPAL (BrPAL4). Group III contained 2 BolPAL (BolPAL1-1 and BolPAL1-2), 1 AtPAL1, 5 BnPAL (BnPAL1-1 to BnPAL1-5), and 2 BrPAL (BrPAL1-1 and BrPAL1-2). Group IV consisted of 4 BolPAL (BolPAL2-1 to BolPAL2-4), 1 AtPAL2, 7 BnPAL (BnPAL2-1 to BnPAL2-7), and 3 BrPAL (BrPAL2-1 to BrPAL2-3) (Figure 1). The uniqueness of this phylogenetic analysis lies in its thorough comparative methodology, emphasizing specific gene clusters and their preservation across several species. This comprehensive and inclusive study improves the comprehension of PAL genes’ evolution, offering a vital structure for future investigations on plant genetics and breeding within the Brassicaceae family.
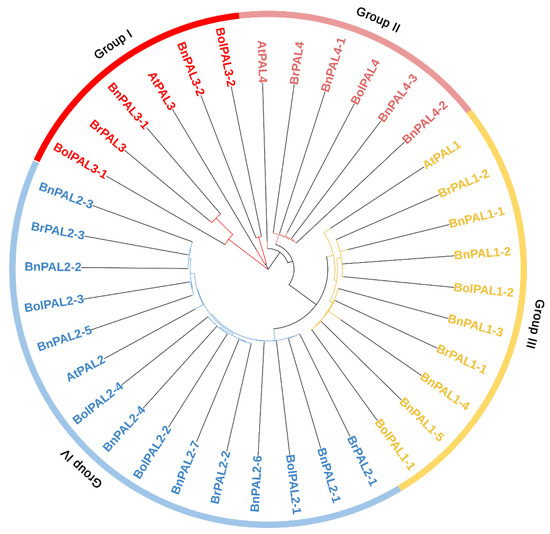
Figure 1.
PAL proteins identified from B. oleracea (9 BolPALs), B. napus (17 BnPALs), B. rapa (7 BrPALs), and A. thaliana (4 AtPALs) were used to create the neighbor-joining phylogenetic tree. Node distributions divided all the PAL genes into four primary groups (I, II, III, and IV) separated by different colors.
2.3. Circos and Synteny Analysis of BolPAL Genes
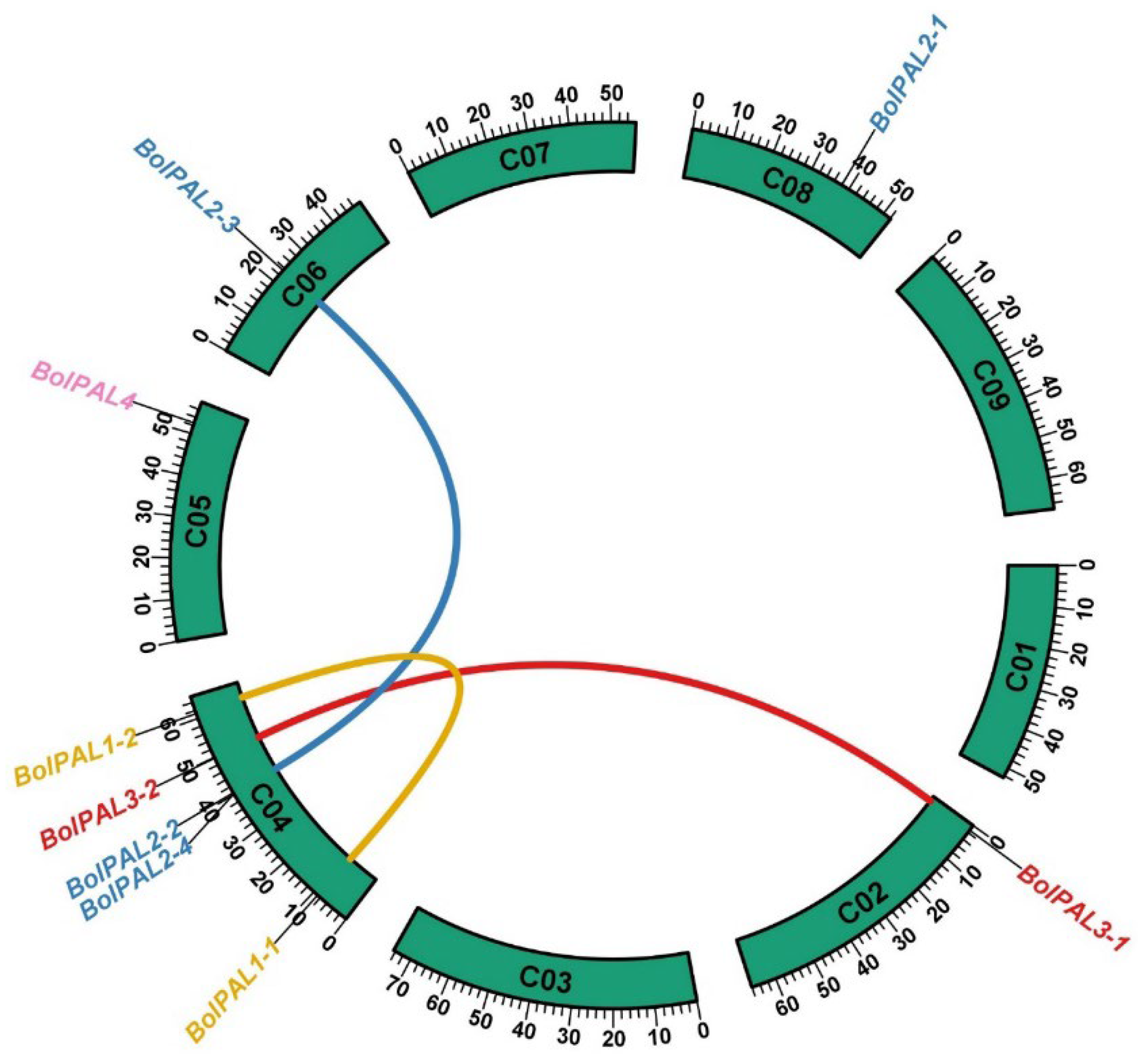
The objective of employing a Circos analysis in the investigation of a gene family is to visually represent the genomic distribution and syntenic connections among the genes across various chromosomes and species [21]. This study aids in the identification of gene duplication, conservation, and rearrangement patterns, which offer constructive understandings of the evolutionary dynamics and functional diversity of the BolPALs. Our Circos analysis revealed that 9 BolPAL genes are distributed unevenly on 5 out of 10 chromosomes of B. oleracea (Figure 2). C04 contains the five (BolPAL1-1, BolPAL1-2, BolPAL2-2, BolPAL2-4, and BolPAL3-2) BolPAL genes, while C02, C05, C06, and C08 contain one gene each (Figure 2). In addition, two paralogous and one homologous gene were identified. Moreover, we found only one tandem and one proximal distribution on chromosome C04. These results indicated that the BolPAL gene family has evolved through not only whole genome triplication, but also through tandem and proximal duplications.
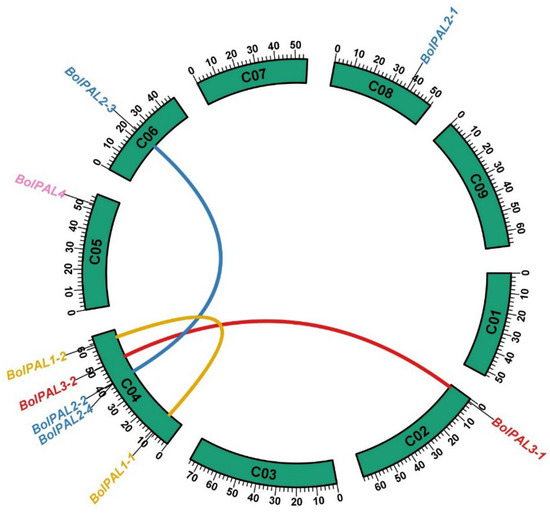
Figure 2.
The chromosomal dispersal and inert-chromosomal interaction of the BolPALs are illustrated in circular form. Lines of different colors represent the PAL genes’ pairs in B. oleracea.
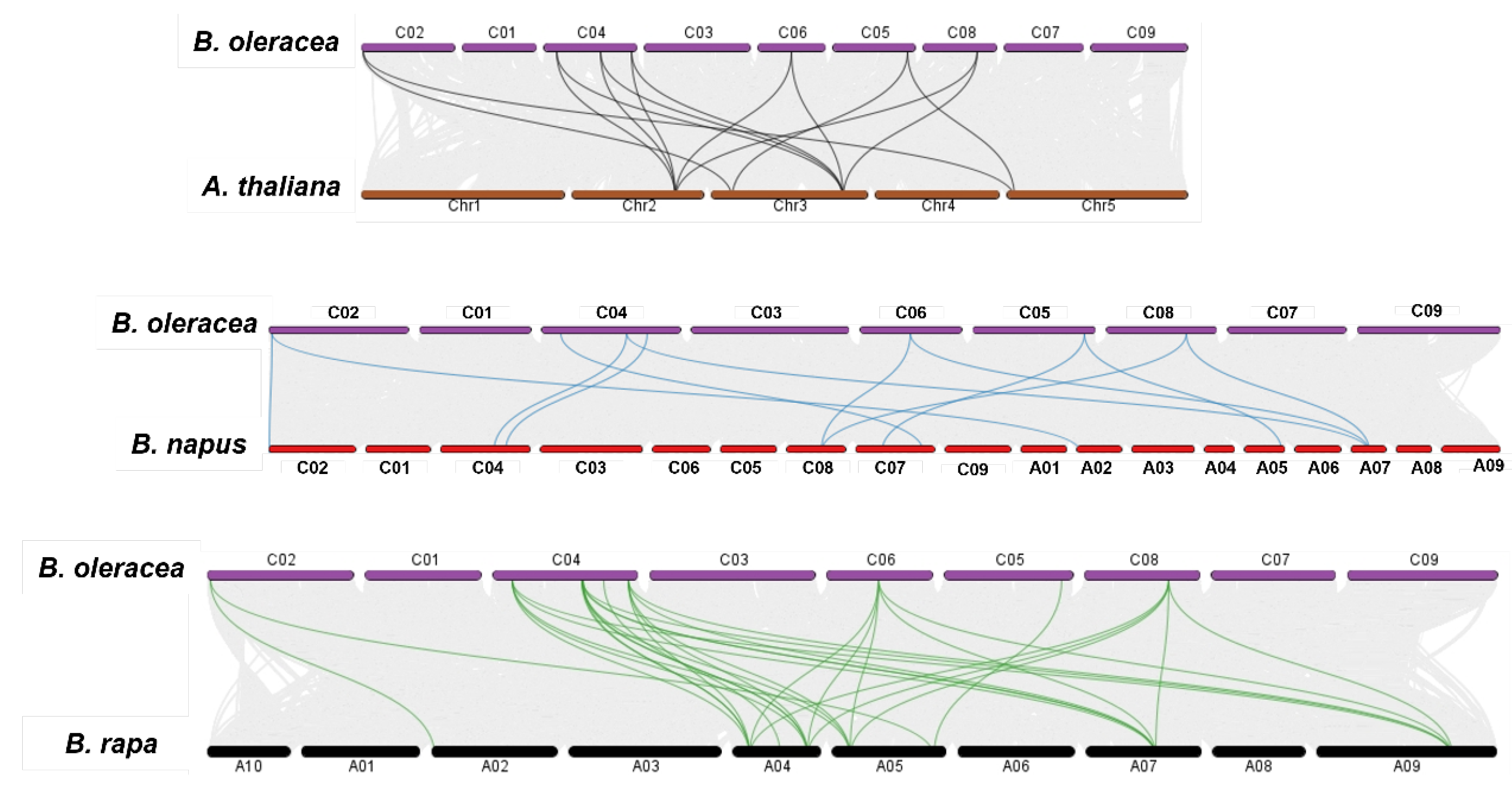
Evolutionary syntenic relations of the BolPAL genes with the A. thaliana, B. napus, and B. oleracea genomes were observed via a collinearity analysis (Figure 3). According to the result, all the BolPAL genes have syntenic connotations with the AtPAL, BnPAL, and BrPAL genes. In detail, the AtPALs from Chr2 are associated to the BolPALs from C04, C05, C06 and C08. The AtPALs from Chr3 made syntenic associations with the BolPALs from C02, C04, C05, and C08. The AtPALs from Chr5 are associated to the BolPALs from C02 and C05. In the same manner, syntenic associations between the PAL genes from B. oleracea, B. rapa, and B. napus are also diverse (Figure 3). These results show that diverse duplication processes in the whole genome contributed to the evolutionary development of the BolPALs.
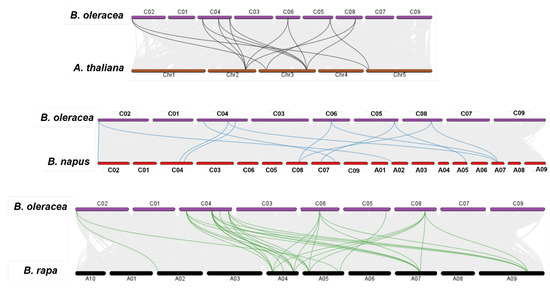
Figure 3.
Graphical illustration of syntenic association of B. oleracea with A. thaliana, B. rapa, and B. napus. Gray lines show all syntenic pairing, while colored lines represent the syntenic association of PAL genes between B. oleracea, A. thaliana, and B. napus genomes.
2.4. Gene Structure and Conserved Motif Analysis
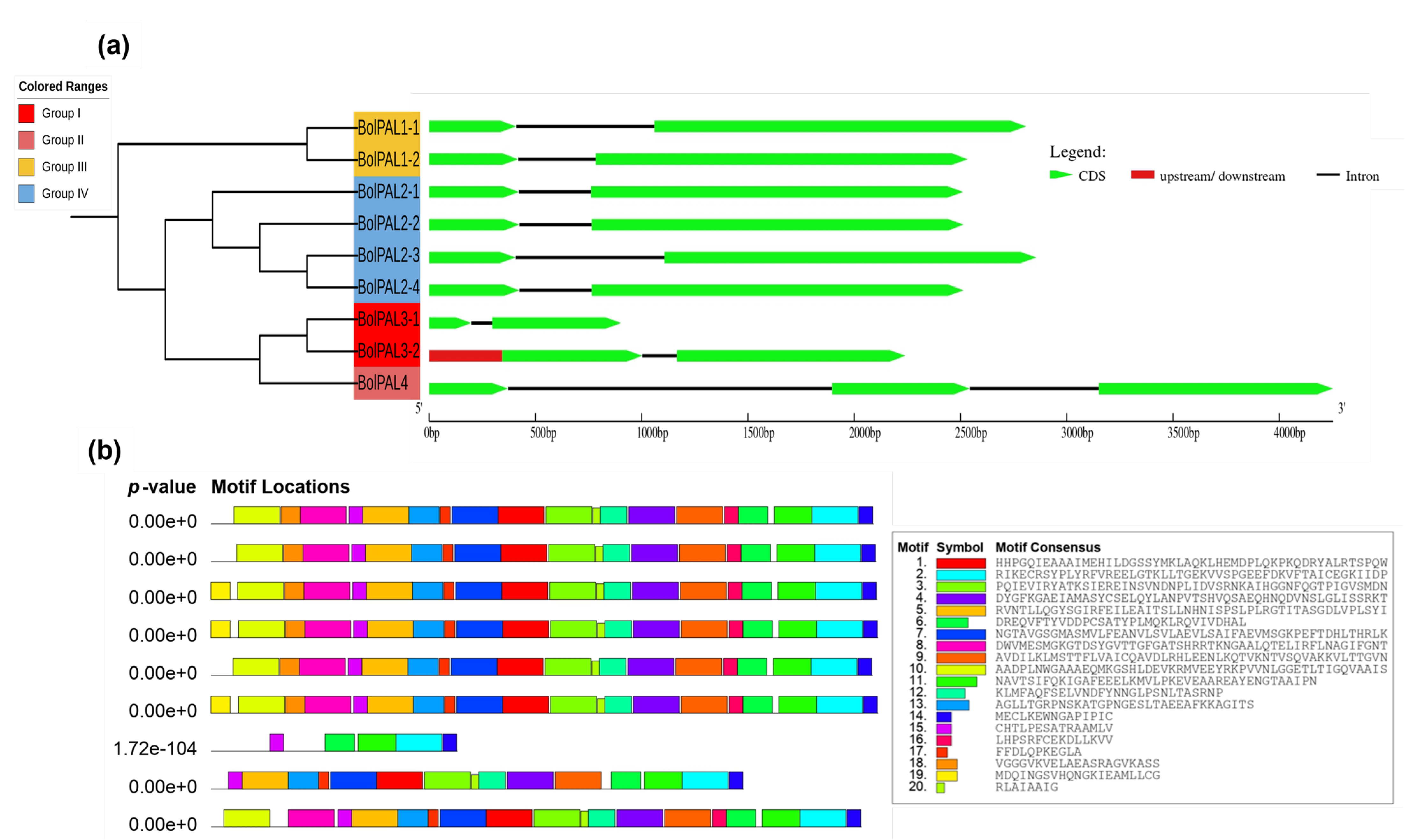
An examination of gene structure and conserved motifs was conducted to gain a better insight into the arrangement and functional components of the genes. This analysis provides valuable insights into the evolutionary history and regulatory processes of the genes. To understand the BolPAL gene family’s development, the exon–intron configuration and the motif distribution were observed (Figure 4a,b). We observed that all the BolPAL genes contain only one intron, except BolPAL4, which contains two introns, while two to three exons were identified in the BolPAL gene family (Figure 4a). Genes from all the groups were found to have a similar exon–intron distribution. In the same manner, all groups possessed 18–19 motifs distributions in their protein regions except BolPAL3-1 from group I, which contains 5 motifs (Figure 4b). Information of the motifs is given in Table S3. The preserved distribution of introns/exons and motifs within and among several classes of BolPAL genes suggests that these genes have retained their structural integrity and functional roles throughout the course of evolution. This evidence indicates that the BolPAL genes have a vital and consistent function in growth and regulation.
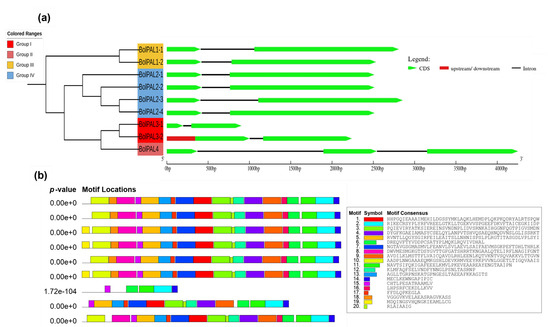
Figure 4.
Structural and conserved motif analysis of the PAL family from B. oleracea. Based on phylogenetic analysis, PAL genes were divided into four different groups (I–IV). (a) Gene structural analysis. (b) The conserved motifs of BolPALs.
2.5. Recognition of Cis-Acting Elements in the Promoter Region of BolPALs
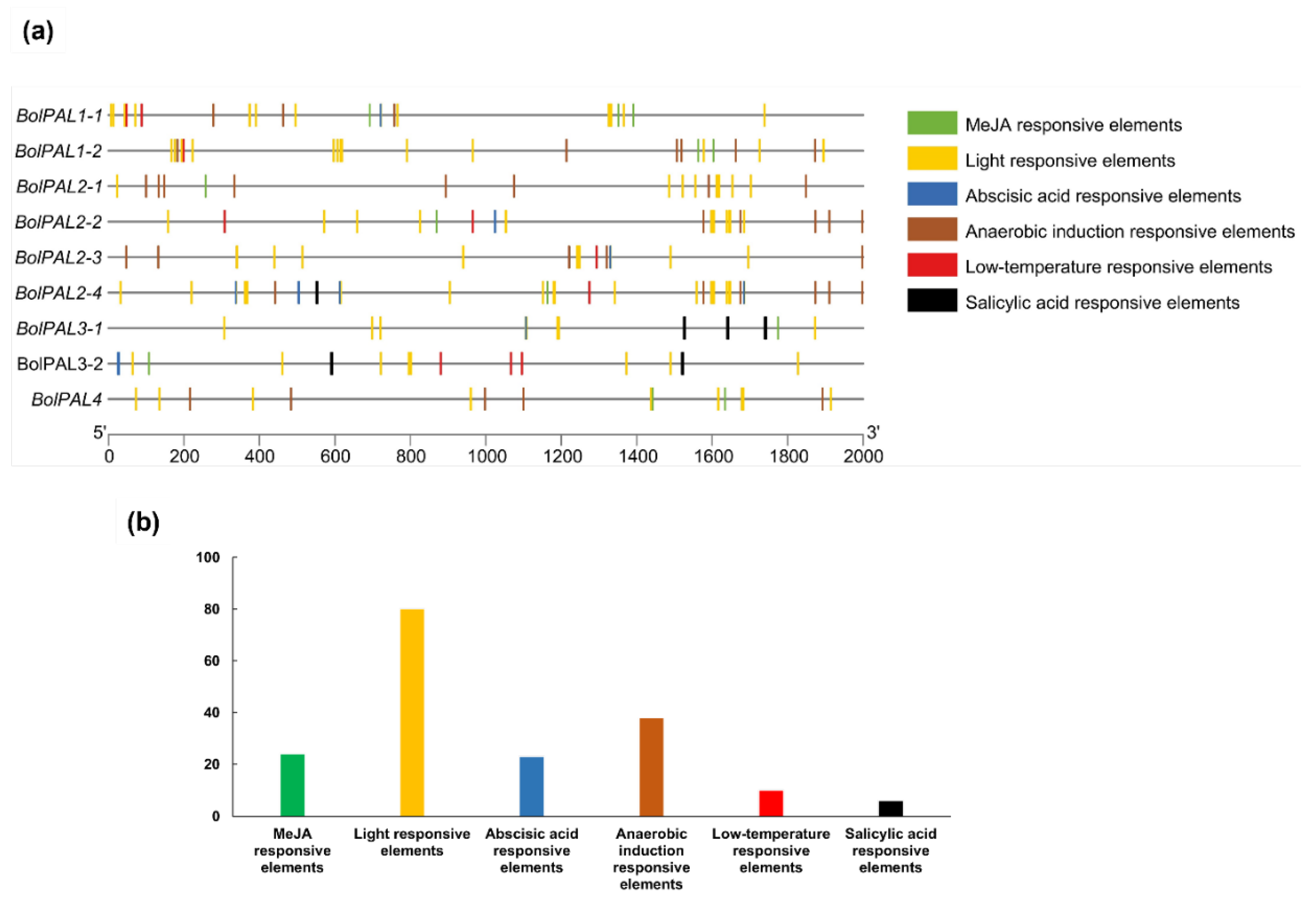
The identification of cis-acting elements in the promoter region involves the recognition of particular DNA sequences that control the transcription of neighboring genes [22]. These elements are essential for comprehending the regulation of gene expression in response to different physiological and environmental signals [23]. A study of the cis-elements present on 2000 base pairs from the upstream region of each BolPAL gene was performed to find out the regulatory pathway and the function of genes (Figure 5; Table S4). Three diverse phytohormone-related regulatory elements were identified in the BolPAL genes, including ABA (ABRE-ACGTG), MeJA (TGACG-motif), and salicylic acid (SA) (TCA-element) (Figure 5a). Aside from the phytohormones, we also discovered other factors, such as anaerobic condition-responsive (ARE-AAACCA), light-responsive (G-box and GT1-motif), and low-temperature-responsive (LTR-CCGAAA) cis-elements, that we expect will further enhance the activity and regulatory pattern of the BolPAL genes (Figure 5a; Table S4).
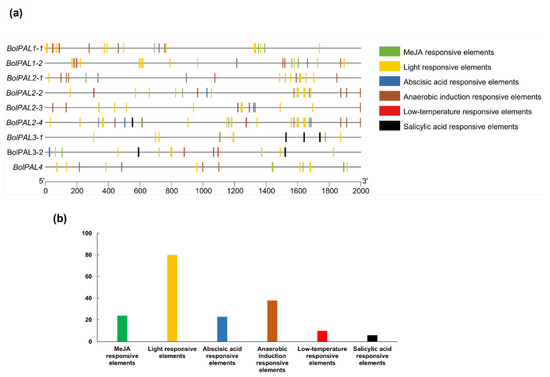
Figure 5.
Identification of cis-acting elements in the BolPAL gene promoters. (a) Different abiotic stress (anaerobic, light, and low temperature)- and phytohormone (ABA, MeJA, and SA)-related cis-regulatory elements in BolPALs. (b) Graphical representation of total number of different phytohormones and other abiotic stress-related elements targeting BolPAL genes.
Moreover, we identified the total number of phytohormone-responsive and abiotic-responsive elements (Figure 5b). The results demonstrated that the promoter region of the BolPAL genes is targeted by MeJA-, ABA-, light-, and anaerobic-responsive elements. These phytohormone-related and different stress-related elements in the BolPALs’ promoter regions predict that transcriptional profiling of these genes may vary due to these stresses.
2.6. Investigation of miRNAs Directing BolPALs
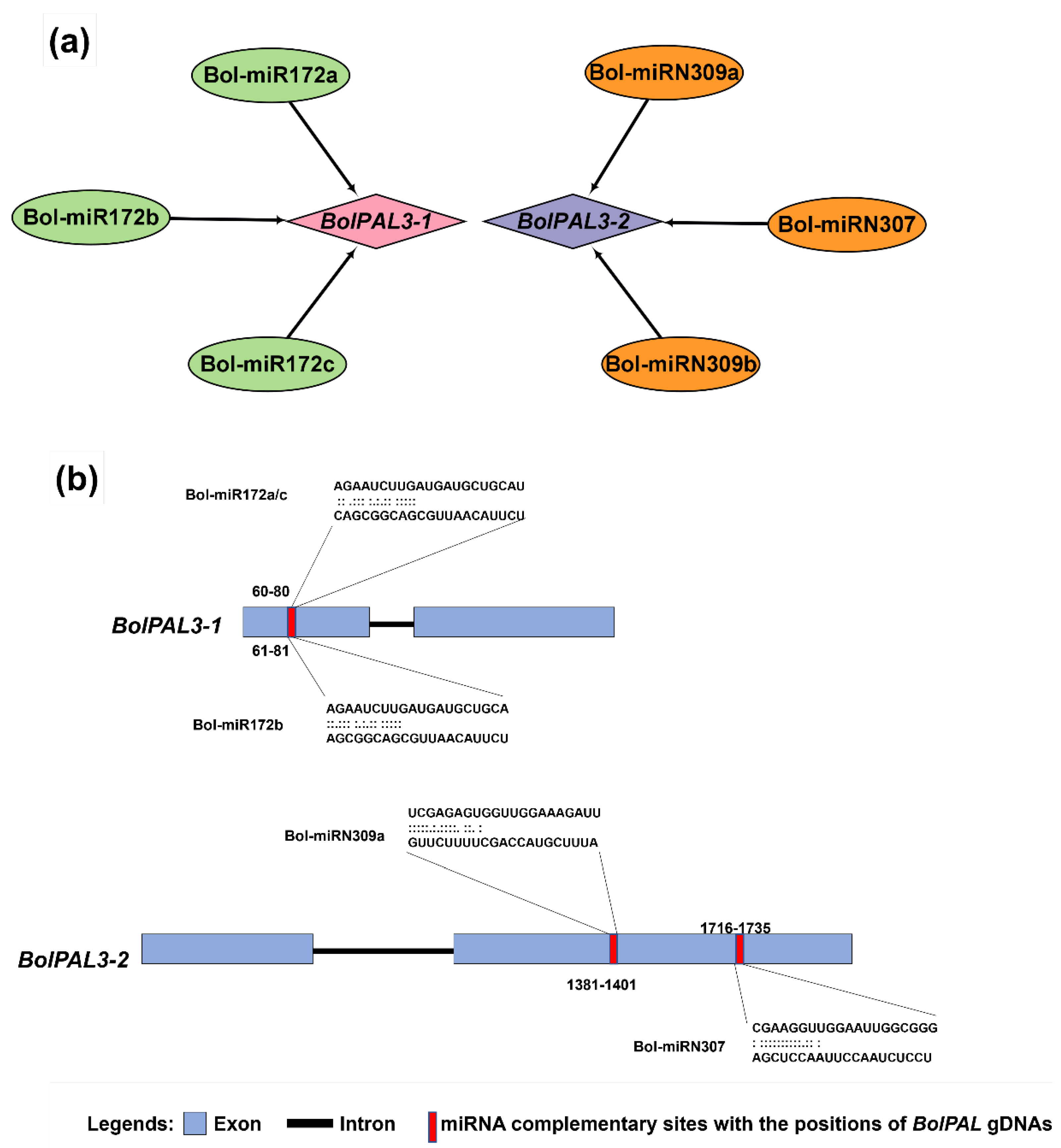
Different biotic stress- and abiotic stress-related responses in many plants were found to have connections with miRNA-directed regulators. Therefore, to enhance our knowledge of the miRNAs controlling the regulation of the BolPALs, we identified the miRNAs targeting different BolPAL genes, as revealed in Figure 6, in which BolPAL3-1 and BolPAL3-2 were found to be targeted by Bol-miR172a-c and BolmiRN309a-c. The conserved miRNA family miR172 is found in many plant species and has been linked to a number of biological processes, such as the regulation of flowering time, the phase transition from vegetative to reproductive growth, and the development of floral organs [24,25,26]. Our findings suggest that the control of BolPAL3-1 and BolPAL3-2 by Bol-miR172a-c may have a similar impact on important developmental processes in B. oleracea, suggesting a wider functional significance of these miRNAs in the growth and development of plants. Detailed information on these putative miRNAs is presented in Table S5.
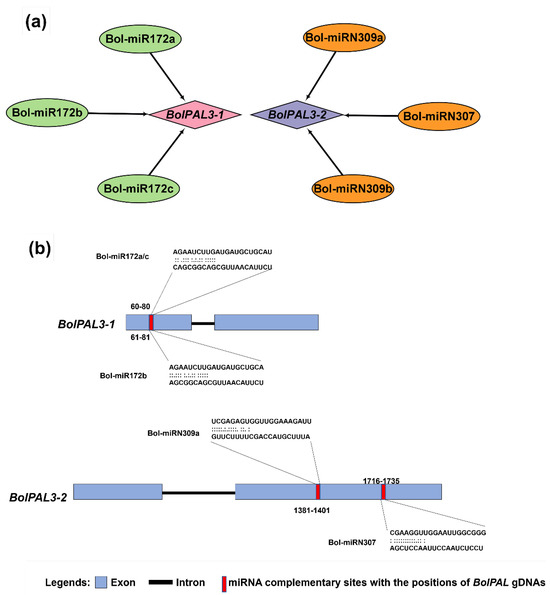
Figure 6.
The miRNA targeting sites in the BolPAL genes (BolPAL3-1 and BolPAL3-2). (a) Network of interaction between BolmiRNAs targeting the BolPAL genes while (b) showing the targeting sites in BolPAL3-1 and BolPAL3-2 genes.
2.7. GO Annotation of BolPAL Gene Family
To predict the functions of the BolPALs, we executed a GO enrichment analysis based on their molecular function (MF), biological process (BP), and cellular component (CC) classes (Figures S2 and S3). These terminologies assist us in predicting the molecular functions of different genes. Complete information on the annotation results is available in Table S6.
The GO-MF enrichment analysis predicted seven enriched terms, ammonia-lyase activity (GO:0016841), catalytic activity (GO:0003824), carbon–oxygen lyase activity (GO:0016835), phenylalanine ammonia-lyase activity (GO:0045548), hydro-lyase activity (GO:0016836), lyase activity (GO:0016829), and transferring alkyl or aryl (other than methyl) groups (GO:0016765). While the GO-BP enrichment analysis found 14 enriched terms, including the L-phenylalanine catabolic process (GO:0006559), the sulfur compound metabolic process (GO:0006790), developmental cell growth (GO:0048588), unidimensional cell growth (GO:0009826), the metabolic process (GO:0008152), cell morphogenesis involved in differentiation (GO:0010770), photoreactive repair (GO:0006281), the alpha-amino acid biosynthetic process (GO:1901607), the developmental process involved in reproduction (GO:0003006), the multicellular organismal process (GO:0032501), fertilization (GO:0007338), localization (GO:0051179), the response to stimulus (GO:0048583), and unidimensional cell growth (GO:0009826).
The GO-CC enrichment annotation examination spotted six enriched terms, cytoplasm (GO:0005737), nucleus (GO:0005634), cellular anatomical entity (GO:0110165), extra cellular space (GO:0005615, cellular anatomical entity (GO:0110165), and protein-containing complex (GO:0032991) (Figure S3). The GO enrichment data suggest that the PAL genes have a crucial function in the formation of anthocyanin and the metabolism of phenylpropanoid.
2.8. Expression Analysis of BolPAL Genes by Transcriptome Analysis
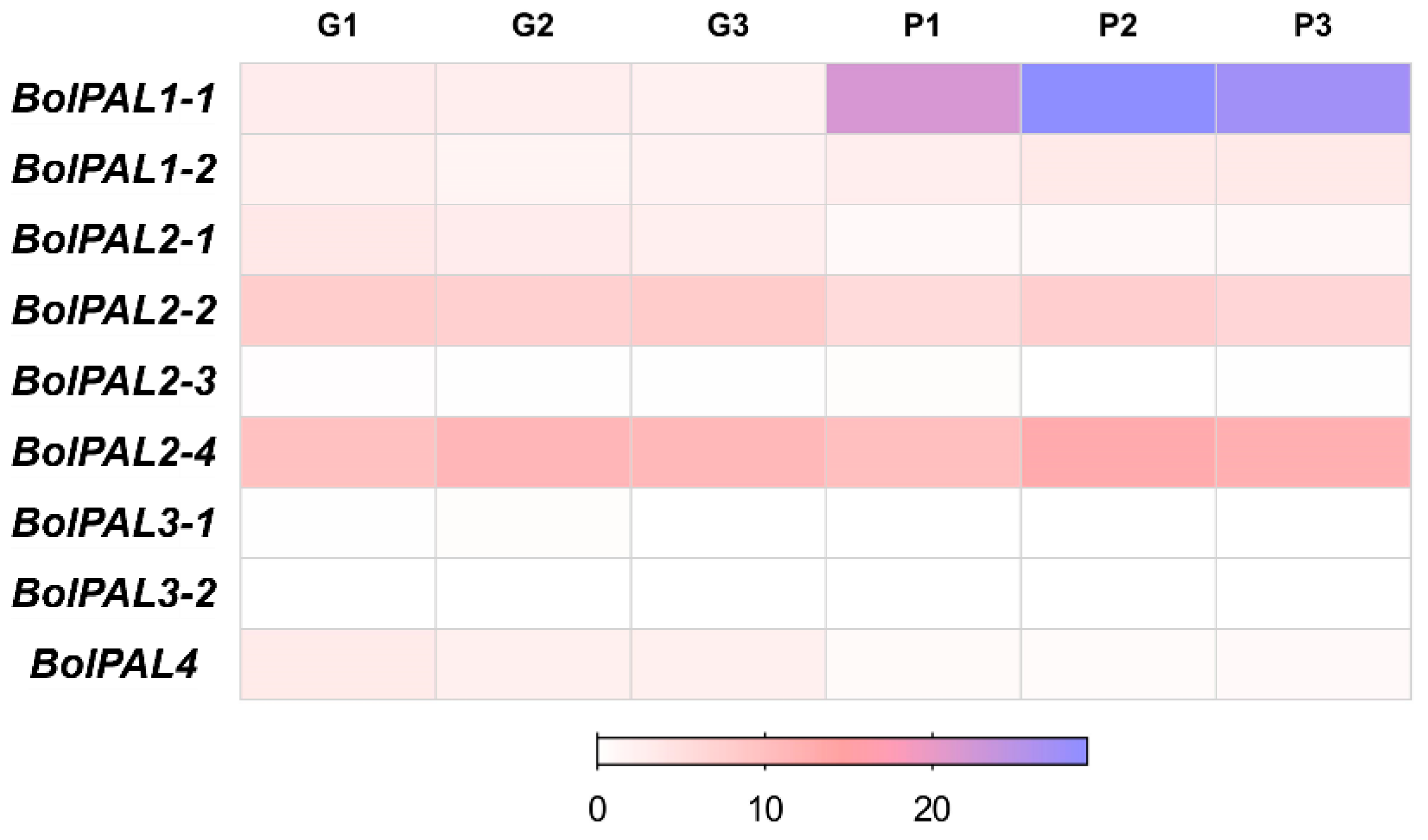
The increased expression of PAL genes can upregulate the production of PAL enzymes, leading to a higher flow of substrates via the phenylpropanoid pathway and finally resulting in greater amounts of anthocyanins [27,28]. In contrast, the reduction or suppression of PAL genes might lead to a decrease in anthocyanin synthesis [29,30]. According to the transcriptome analysis [31], out of nine, only one gene, BolPAL1-1, was upregulated, while two genes, BolPAL2-1 and BolPAL4, were downregulated in purple-stalked Chinese kale (B. oleracea), as compared to green (Figure 7; Table S7). These results suggest that the three PAL genes are essential for the creation of anthocyanin and purple color. It has been reported that PAL genes catalyze the initial step in the phenylpropanoid pathway, which results in the production of phenolic chemicals [31]. These molecules serve as precursors for the process of anthocyanin biosynthesis, which leads to the build-up of pigments that are responsible for the purple coloration in the plant [31].
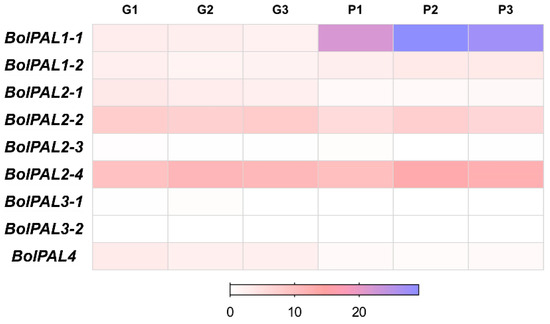
Figure 7.
Heatmap of transcriptome expression analysis of BolPAL genes in green- (G1, G2, G3) and purple-stalked (P1, P2, P3) Chinese kale (B. oleracea).
Gaining understanding of the function of PAL genes in anthocyanin biosynthesis in B. oleracea is crucial for plant breeders and biotechnologists who seek to create cultivars with a heightened nutritional value, aesthetic appeal, and potentially greater resilience to stress by augmenting anthocyanin levels. Modifying the expression of the PAL gene using genetic engineering or selective breeding techniques could be employed to accomplish these objectives.
2.9. Expression Analysis of BolPAL Genes under Phytohormone, Abiotic, and Light Treatments
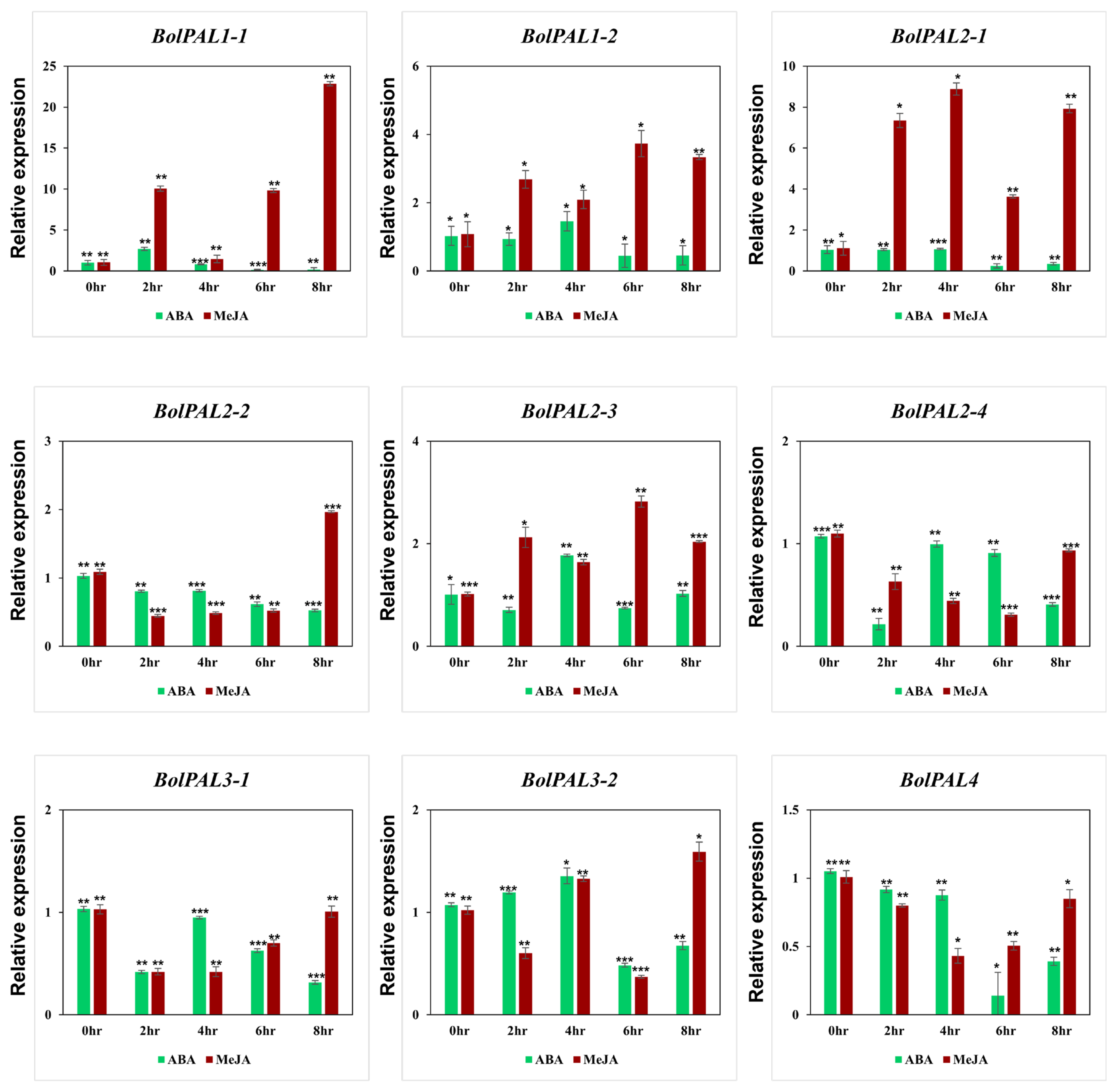
Under different phytohormones, the growth of plants can be affected at physiological, molecular, and biochemical levels. Our cis-element identification revealed the presence of ABA-, MeJA-, and SA-responsive elements in the BolPAL genes. Therefore, we applied ABA and MeJA phytohormones to observe their role in the expression of the BolPAL genes (Figure 8). Six members of this gene family were expressed under MeJA treatment at different times, which include BolPAL1-1 at 2 h, 6 h, and 8 h; BolPAL1-2 and BolPAL2-1 at 2 h, 4 h, 6 h, and 8 h; BolPAL2-2 at 8 h; BolPAL2-3 at 2 h, 6 h, and 8 h; and BolPAL3-2 at 8 h, while the remaining three genes were found to be downregulated (Figure 8). Under ABA treatment, almost all the members of this gene family were found to be downregulated, except for a few genes that were partially expressed at different time points, such as BolPAL1-2 and BolPAL2-3 at 4 h, and BolPAL3-2 at 2 h and 4 h (Figure 8). Three genes, named BolPAL1-2, BolPAL2-3, and BolPAL3-2, are common genes that displayed elevated expression under both the phytohormones (MeJA and ABA), which predicts that these three genes are the most important for further studies and can be helpful in future breeding programs of B. oleracea crops.
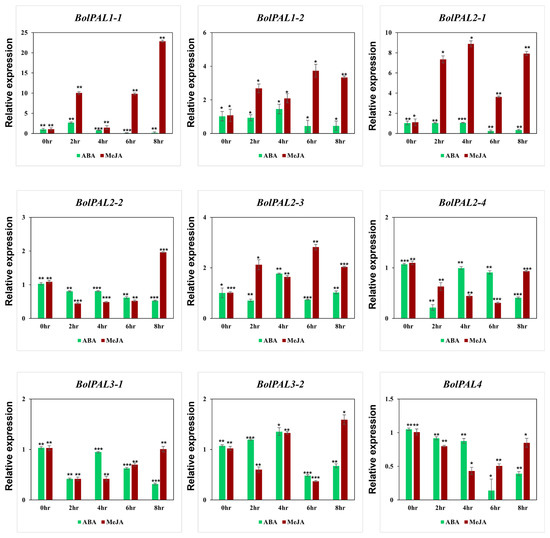
Figure 8.
Graphical demonstration of the expression profile of the BolPAL genes in B. oleracea under phytohormonal stresses (ABA and MeJA) (0 h as control, 2 h, 4 h, 6 h, and 8 h). The 2−∆∆CT method was used to examine the results. (*, p < 0.01; **, p < 0.005; ***, p < 0.001).
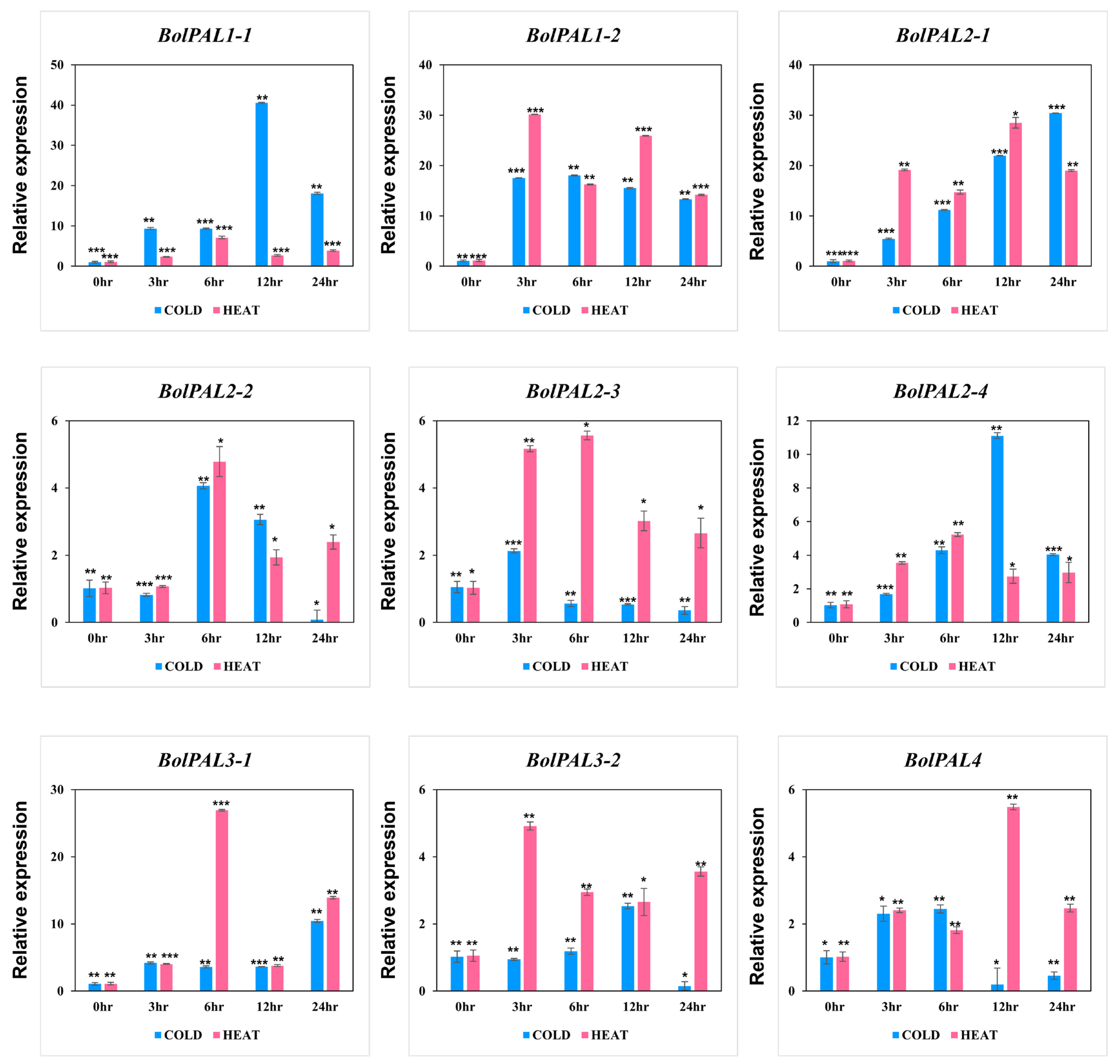
Surprisingly, all the BolPAL genes were regulated under cold and heat stress at different time points (Figure 9). In detail, under cold stress, BolPAL1-1, BolPAL2-4, BolPAL3-2 at 12 h, BolPAL1-2, BolPAL2-2, and BolPAL4 at 6 h; BolPAL2-1 and BolPAL3-1 at 24 h; and BolPAL2-3 at 3 h expressed strongly. Under heat stress, five genes, including BolPAL1-1, BolPAL2-2, BolPAL2-3, BolPAL2-4, and BolPAL3-1 expressed at 6 h; BolPAL1-2 and BolPAL3-2 at 3 h; and BolPAL2-1 and BolPAL4 at 12 h showed elevated expressions. Significant changes in the expression of the PAL genes could result in the promoted synthesis of phenolic compounds, which function as antioxidants and enhance resistance to low and high temperatures [32,33]. These results indicate that PAL genes are crucial in response to studied stress conditions, especially temperature-related stresses.
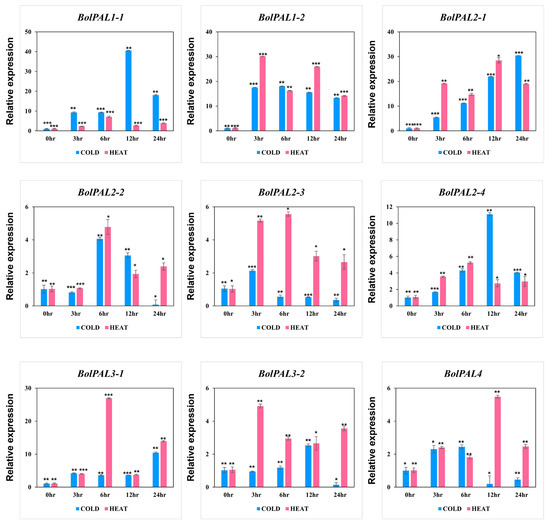
Figure 9.
Graphical presentation of BolPAL genes under low and high temperature stresses at different time points (0 h as control, 2 h, 4 h, 6 h, and 8 h). The 2−∆∆CT method was used to examine the results. (*, p < 0.01; **, p < 0.005; ***, p < 0.001).
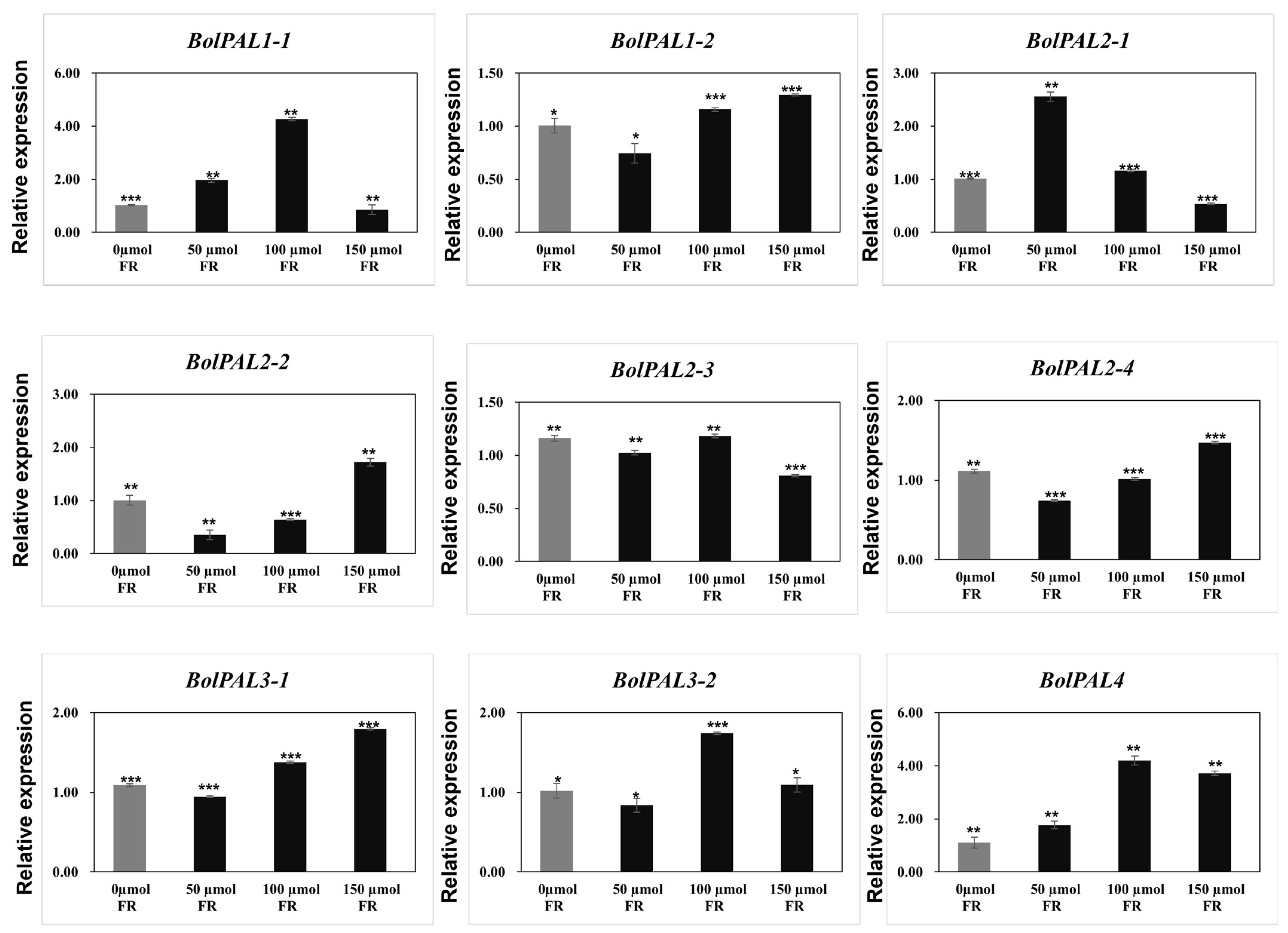
For light stress, white light with a 100 µmol m−2 s−1 intensity as a control, plus 50, 100, and 150 µmol m−2 s−1 FR (FR) were used as stresses. Four genes (BolPAL1-2, BolPAL2-2, BolPAL2-4, and BolPAL3-1) were upregulated at 150 µmol m−2 s−1 FR, whereas BolPAL1-1, BolPAL3-2, and BolPAL4 were expressed at 100 µmol m-2 s-1 FR, and only BolPAL2-1 was upregulated at 50 µmol m−2 s−1 FR (Figure 10). Far-red (FR) light stress activates phytochrome signaling, triggering hormonal pathways and transcription factors that upregulate gene expression [34]. This increases phenolic compound production, helping plants manage oxidative stress and reinforce structural defenses [35,36]. Our results indicate that BolPAL genes play a major role in stress responses and adaptation, as evidenced by the increased expression of these genes under MeJA, ABA, heat, cold, and light stresses. These outcomes can be helpful in developing new cultivars that are more resilient and are stress-tolerant. Additionally, understanding the regulation of BolPAL genes under various stress conditions might aid future studies that aim to enhance agricultural performance and stability in dynamic environmental changes.
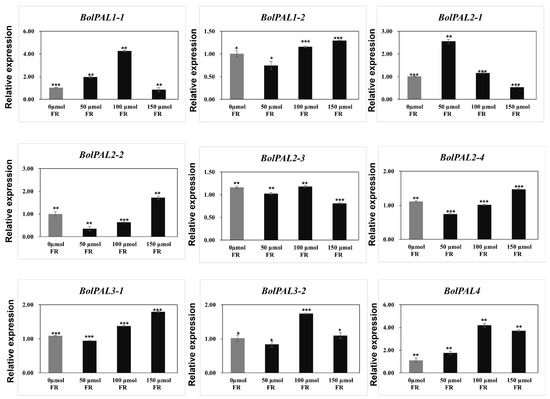
Figure 10.
Graphical illustration of BolPAL genes under white light (100 µmol m−2 s−1), plus 50, 100, and 150 µmol m−2 s−1 far-red (FR). The 2−∆∆CT method was used to examine the results. (*, p < 0.01; **, p < 0.005; ***, p < 0.001).
3. Discussion
Plants use the phenylpropanoid pathway as a major metabolic process to produce a wide range of secondary metabolites, including phenolic acids, lignin, and flavonoids [27]. The enzyme phenylalanine ammonia-lyase (PAL) catalyzes the deamination of the amino acid phenylalanine, which is the first step in this route [31,37]. Phenylalanine is transformed by PAL into trans-cinnamic acid, which thereafter proceeds via a sequence of enzymatic processes to yield different phenylpropanoids [27]. These substances are essential for UV protection, pigmentation, structural integrity, and pathogen defense systems in plants [27,38]. Specifically, plants get their red, blue, and purple hues from anthocyanins, which are generated from flavonoids [31,39,40]. Because they catalyze the first and rate-limiting step in this route, the PAL genes are essential for controlling the flow of metabolites into the phenylpropanoid biosynthesis [27]. Plant growth, stress responses, and general adaptability are all impacted by the activity and expression levels of the PAL genes, which in turn directly affect the synthesis of phenolic chemicals [41,42].
The PAL enzyme plays a crucial role in the secondary phenylpropanoid pathway, enabling the production of lignin, which provides mechanical support, the synthesis of pigments like anthocyanins and flavonoid nodule factors, and defense against biotic and abiotic stresses [43,44]. Thus, a great deal of research has gone into figuring out the signal transduction pathways connected to these mechanisms. As an example, in A. thaliana, overexpressed GmPAL1.1 improved seed maturity under stress conditions of high temperatures and humidity [45]. PtPAL1 expression in poplar was upregulated in response to leaf damage [11]. Furthermore, pepper plants that had CaPAL1 silenced were more vulnerable to Xcv susceptibility [11]. Quadruple mutants of all thte AtPAL genes (AtPAL1-AtPAL4) also showed abnormalities and a significant decrease in the quantities of stored salicylic acid [17].
In this study, we used the Brassica database to identify nine BolPAL genes that aligned with the PAL genes found in other plants, such as cucumber [46], hickory nut [47], and tea tree [6]. These BolPAL genes are notable for having conserved motifs and similar gene structures. These conserved motifs may govern molecular processes in PAL genes in several plant species [11]. The PAL genes from B. oleracea, A. thaliana, B. rapa, and B. napus were analyzed using a phylogenetic analysis, which showed that the BolPAL showed strong affinities with A. thaliana and two additional Brassica crops, B. napus and B. oleracea. The BolPALs exhibited a high degree of genetic variety, which may be explained by the genome-wide triploidization processes that occurred in Brassica after their separation from the A. thaliana lineage [48,49]. The BolPAL family members are unevenly distributed among five chromosomes in B. oleracea, which is comparable to the PAL genes, which are dispersed on a few chromosomes in other species [11]. It has been found that, during the evolution of plants, procreation and ancestor modification can produce many family members, which can be grouped together on a single or multiple chromosomes [8,50], indicating that the BolPAL family’s growth is being driven by chromosomal duplication, while the diverse range of developments among Brassica species may be explained by differences in the gene families’ sizes [51]. Furthermore, it is possible that functional redundancy among close relatives can be quantified, which may lead to the development of more potent methods for detecting traits that are deficient in loss-of-function research involving B. oleracea breeding.
The growth and activation of the PAL gene’s expression are also governed by the promoter activity of the gene, and the promoter region often includes a range of cis-regulatory elements. The cis-acting components of the BolPAL promoter were predicted and analyzed, and the results suggest that the BolPALs may react to light, abiotic stress, and several phytohormones. Based on phytohormone, abiotic, and light treatments, five members of the BolPAL gene family (BolPAL1-1, BolPAL1-2, BolPAL2-1, BolPAL2-2, and BolPAL3-2) were found to have high expression at various times under MeJA treatment (Figure 8). Almost all the members of this gene family were shown to be suppressed when treated with ABA, with the exception of a few genes, such as BolPAL1-2, BolPAL2-3, and BolPAL3-2, which merely expressed highly at different times (Figure 8). Interestingly, at different times, all the BolPAL genes were substantially regulated under heat and cold stresses (Figure 9). White light with 100 µmol m−2 s−1 intensity, plus 50, 100, and 150 µmol m−2 s−1 far-red (FR), were utilized as stressors to further investigate the possible role of the BolPAL genes. At 150 µmol m−2 s−1 FR, the majority of the genes were slightly elevated; however, only a small number of genes were expressed at 50 and 100 umol m−2 s−1 FR (Figure 10). These findings suggest that the PAL genes are essential for adapting to the stressors being studied, especially temperature-related stress.
Similar findings were also discovered in studies conducted on other species [17,52,53,54]. The elevation of the BolPAL expression levels at low temperatures in this study is consistent with the upregulation of CsPAL gene expression levels during cold stress [46]. After receiving exogenous ABA treatment, CsPAL2 and SlPAL5s’ expression was dropped [55]; in the same study, BolPAL’s expression was likewise downregulated; following MeJA treatment, six potato PAL genes were considerably upregulated and four were dramatically downregulated [9]. In this study, under MeJA treatment, four BolPAL genes dramatically downregulated, whereas five of them showed considerable upregulation. BrPAL genes were downregulated under 60 umol m−2 s−1 FR [52]. In our results, only BolPAL1-1 and BolPAL2-1 were slightly upregulated under 50 umol m−2 s−1 FR. In conclusion, under a variety of abiotic stressors and phytohormone treatments, the expression of most BolPAL genes were regulated at different time points. Members of the BolPAL gene family can be differentially induced to express themselves, depending on when stress and phytohormone treatments occur. These variations in gene expression can be beneficial to plants as they may enable them to have varying degrees of stress resistance and play a significant role in hormone signaling.
4. Materials and Methods
4.1. Identification of BolPALs in B. oleracea
The Brassica database (BRAD) http://www.brassicadb.cn/ (accessed on 12 May 2024) was used to identify and obtain all the sequences of the BolPALs [20], while the protein sequences of A. thaliana were downloaded from the Arabidopsis Information Resource (TAIR) at http://www.arabidopsis.org/ (accessed on 12 May 2024) [56]. To verify all the PAL genes of B. oleracea, we performed an analysis to find the PAL gene-specific lyase aromatic domain (PF00221) in all the protein sequences by using the NCBI conserved domain database, https://www.ncbi.nlm.nih.gov/Structure/cdd/wrpsb.cgi/ (accessed on 12 May 2024), followed by the TBtools software V 1.098: https://github.com/CJ-Chen/TBtools (accessed on 12 May 2024) [57]. An online tool, ExPASy (http://web.expasy.org/protparam/) (accessed on 12 May 2024), was employed to measure the physiochemical properties (molecular weight and isoelectric points) of the BolPALs. The gene structure of the BolPALs was identified and constructed by accessing the Gene Structure Display Server 2.0, http://gsds.gao-lab.org/ (accessed on 12 May 2024), while a conserved motif analysis of the BolPAL proteins was carried out using MEME (V 4.11.4; accessed on 12 May 2024).
4.2. Phylogenetic Tree and Synteny Investigation of BolPALs
To observe their evolutionary relationships, the protein sequences of the PAL family from B. oleracea, B. rapa, A. thaliana, and B. napus were used. The neighbor-joining (NJ) method, with 1000 bootstrap replicates, was used to align and construct the peptide sequences using MEGA-X (accessed on 12 May 2024). Collinearity and synteny analysis of all PAL genes were performed through TBtools software V 1.098; https://github.com/CJ-Chen/TBtools/ (accessed on 12 May 2024) [57].
4.3. Cis-Elements Investigation in the Promoter Region of BolPALs
To detect cis-regulatory elements, two thousand base pairs from upstream of the genome sequences of each BolPAL were taken from BRAD, http://www.brassicadb.cn/ (accessed on 12 May 2024) [20]. An online tool called PlantCARE, http://bioinformatics.psb.ugent.be/webtools/plantcare/html/ (accessed on 12 May 2024), was accessed to analyze the promoter regions and the prediction of cis-regulatory elements. The obtained results were presented using TBtools software V 1.098, https://github.com/CJ-Chen/TBtools/ (accessed on 12 May 2024) [57].
4.4. Prediction of Putative miRNA Targeting BolPALs
The genomic sequences of the BolPALs were compiled to categorize possible miRNAs by the psRNATarget database, http://plantgrn.noble.org/psRNATarget/ (accessed on 12 May 2024). The Cytoscape V3.8.2, https://cytoscape.org/download.html/ (accessed on 12 May 2024), software program was used to construct the network among the predicted miRNAs and the corresponding target BolPALs [58].
4.5. Expression Profiling of BolPALs by Transcriptome Data
We used RNA-seq data [31] to highlight the BolPAL genes’ expression in green-stalked (ZSJL) and purple-stalked (HJJL, R) Chinese kale (B. oleracea). FPKMs (fragments per kilobase of exon model per million mapped reads) were used to analyze the expression values. The heatmap of the BolPAL genes’ expression was created using GraphPad Prism 9.0.0 software, accessible at https://www.graphpad.com/, (accessed on 12 May 2024).
4.6. Plant Materials and Stress Conditions
In this study, green-stalked (ZSJL), wild-type Chinese kale (B. oleracea) that was obtained from the Vegetable Research Institute, Guangdong Academy of Agricultural Sciences, was used for different stress treatments. The vigorous seeds that had a 100% germination rate were selected and sterilized with a 10% hypochlorous acid solution for 5 min. Cycling conditions of 25 °C and 16 h/8 h light/dark were used to grow seeds in the growth chamber. To determine the effect of the phytohormones, three-week-old seedlings were treated with 100 µM abscisic acid (ABA) and 100 µM MeJA by exogenously spraying [59]. All the leaves were collected at 0 h as the control, as well as 2 h, 4 h, 6 h, and 8 h after the treatment was applied. For light stress, white LED light with 100 µmol m−2 s−1 intensity as the control and white light plus 50, 100, and 150 µmol m−2 s−1 far-red light (FR) were applied for three weeks, and all the leaves were collected [52]. To analyze the temperature stress, 4 °C for cold stress and 38 °C for heat stress were applied to the three-week-old seedlings under 16 h light/8 h dark conditions, and all the leaves were collected at 0 h as the control, as well as 3 h, 6 h, 12 h, and 24 h after the treatment. Three biological replicates of each treatment were taken and rapidly frozen in liquid nitrogen and stored at −80 °C.
4.7. RNA Extraction and qRT-PCR Analysis
The total RNA from the leaf was isolated using the Tiangen RNA Extraction Kit (Beijing, China). The Agilent Bioanalyzer 2100 system confirmed the RNA quantity (Agilent Technologies, Beijing, China). In detail, the leaf samples were ground into a fine powder using a mortar and pestle, followed by adding the lysis buffer, ensured thorough homogenization. Further, ethanol was added to facilitate the RNA binding to the column, and the sample was then transferred to the spin column, then it was centrifuged to bind the RNA to the membrane, then the column was washed with wash buffers to remove contaminants, then the purified RNA was eluted in RNase-free water by centrifugation. The cDNA was synthesized by a cDNA Synthesis SuperMix kit (TransGen Biotech, Beijing, China) and then diluted 10× by adding distilled deionized water. The qRT-PCR was performed with a CFX Connect Real-Time System (Bio-Rad, Hercules, CA, USA) using a SYBER® Green Supermix (Vazyme, Nanjing, China). BolActin primers were used as a control to analyze the results. The qRT-PCR reaction was performed as follows: 94 °C for 10 min, followed by 40 cycles of 94 °C for 15 s, 60 °C for 30 s, and 72 °C for 10 s. Three biological replicates were used for each reaction, and then all the data were analyzed using the 2−∆∆CT method. Table S1 contains information on all the primers for the BolPAL genes.
5. Conclusions
In this study, we identified nine BolPAL members that play a crucial role in the metabolic pathway of anthocyanin biosynthesis, located on five different chromosomes. Through a comprehensive and systematic analysis, including conserved structural domains, chromosomal positioning, phylogenetic relationships, and gene expression analyses under various stresses, we observed a strong correlation between the promoter cis-elements of the BolPAL genes and their expression in response to abiotic stresses, light, and plant hormones. These findings offer novel insights into the functional role of BolPALs and contribute to the ongoing development of the regulatory network governing anthocyanin biosynthesis in B. oleracea.
Supplementary Materials
The following supporting information can be downloaded at: https://www.mdpi.com/article/10.3390/ijms251910276/s1.
Author Contributions
U.K. and M.F. designed and carried out the experiments and wrote the original draft; S.J. performed the visualization; M.L. and I.K. performed the validation; and J.G. and G.L. proofread the draft and gave the final approval for the publication of the manuscript. All authors have read and agreed to the published version of the manuscript.
Funding
This research was funded by the 2023 Provincial Rural Revitalization Strategy Special Seed Industry Revitalization Action Project (2023-NBH-00-008) and the 2022 Provincial Rural Revitalization Strategy Special Fund Seed Industry Revitalization Project (2022-NJS-03-001).
Institutional Review Board Statement
Not applicable.
Informed Consent Statement
Not applicable.
Data Availability Statement
The data presented in this study are available on request from the corresponding author.
Conflicts of Interest
The authors declare no conflicts of interest.
References
- Jones, D.H. Phenylalanine ammonia-lyase: Regulation of its induction, and its role in plant development. Phytochemistry 1984, 23, 1349–1359. [Google Scholar] [CrossRef]
- Vogt, T. Phenylpropanoid biosynthesis. Mol. Plant 2010, 3, 2–20. [Google Scholar] [CrossRef] [PubMed]
- Koukol, J.; Conn, E.E. The metabolism of aromatic compounds in higher plants: IV. Purification and properties of the phenylalanine deaminase of Hordeum vulgare. J. Biol. Chem. 1961, 236, 2692–2698. [Google Scholar] [CrossRef]
- Blake, V.C.; Kling, J.G.; Hayes, P.M.; Jannink, J.L.; Jillella, S.R.; Lee, J.; Matthews, D.E.; Chao, S.; Close, T.J.; Muehlbauer, G.J. The Hordeum toolbox: The barley coordinated agricultural project genotype and phenotype resource. Plant Genome 2012, 5, 81–91. [Google Scholar] [CrossRef]
- de Jong, F.; Hanley, S.J.; Beale, M.H.; Karp, A. Characterisation of the willow phenylalanine ammonia-lyase (PAL) gene family reveals expression differences compared with poplar. Phytochemistry 2015, 117, 90–97. [Google Scholar] [CrossRef]
- Chen, X.; Wang, P.; Gu, M.; Hou, B.; Zhang, C.; Zheng, Y.; Sun, Y.; Jin, S.; Ye, N. Identification of PAL genes related to anthocyanin synthesis in tea plants and its correlation with anthocyanin content. Hortic. Plant J. 2022, 8, 381–394. [Google Scholar] [CrossRef]
- Zhao, T.; Li, R.; Yao, W.; Wang, Y.; Zhang, C.; Li, Y. Genome-wide identification and characterisation of phenylalanine ammonia-lyase gene family in grapevine. J. Hortic. Sci. Biotechnol. 2021, 96, 456–468. [Google Scholar] [CrossRef]
- Zhan, C.; Li, Y.; Li, H.; Wang, M.; Gong, S.; Ma, D.; Li, Y. Phylogenomic analysis of phenylalanine ammonia-lyase (PAL) multigene family and their differential expression analysis in wheat (Triticum aestivum L.) suggested their roles during different stress responses. Front. Plant Sci. 2022, 13, 982457. [Google Scholar] [CrossRef]
- Mo, F.; Li, L.; Zhang, C.; Yang, C.; Chen, G.; Niu, Y.; Si, J.; Liu, T.; Sun, X.; Wang, S. Genome-wide analysis and expression profiling of the phenylalanine ammonia-lyase gene family in Solanum tuberosum. Int. J. Mol. Sci. 2022, 23, 6833. [Google Scholar] [CrossRef]
- Olsen, K.M.; Lea, U.S.; Slimestad, R.; Verheul, M.; Lillo, C. Differential expression of four Arabidopsis PAL genes; PAL1 and PAL2 have functional specialization in abiotic environmental-triggered flavonoid synthesis. J. Plant Physiol. 2008, 165, 1491–1499. [Google Scholar] [CrossRef]
- Rawal, H.; Singh, N.; Sharma, T. Conservation, divergence, and genome-wide distribution of PAL and POX A gene families in plants. Int. J. Genom. 2013, 2013, 678969. [Google Scholar]
- Zhang, X.; Liu, C. Multifaceted regulations of gateway enzyme phenylalanineammonia-lyase in the biosynthesis of phenylpropanoids. Mol. Plant 2015, 8, 17–27. [Google Scholar] [CrossRef] [PubMed]
- Zhu, Q.; Xie, X.; Lin, H.; Sui, S.; Shen, R.; Yang, Z.; Lu, K.; Li, M.; Liu, Y.-G. Isolation and functional characterization of a phenylalanine ammonia-lyase gene (SsPAL1) from coleus (Solenostemon scutellarioides (L.) Codd). Molecules 2015, 20, 16833–16851. [Google Scholar] [CrossRef]
- Yan, F.; Li, H.; Zhao, P. Genome-Wide Identification and transcriptional expression of the PAL Gene family in common Walnut (Juglans regia L.). Genes 2019, 10, 46. [Google Scholar] [CrossRef]
- Zeng, J.; Ouyang, L.; Liu, J.; He, H.; Zhu, C.; Peng, X.; He, X.; Fu, J.; Chen, X.; Bian, J. Whole genome analysis and stress expression research of PAL gene in rice. Genom. Appl. Biol. 2018, 37, 3881–3888. [Google Scholar]
- Shi, R.; Shuford, C.M.; Wang, J.P.; Sun, Y.-H.; Yang, Z.; Chen, H.-C.; Tunlaya-Anukit, S.; Li, Q.; Liu, J.; Muddiman, D.C. Regulation of phenylalanine ammonia-lyase (PAL) gene family in wood forming tissue of Populus trichocarpa. Planta 2013, 238, 487–497. [Google Scholar] [CrossRef]
- Huang, J.; Gu, M.; Lai, Z.; Fan, B.; Shi, K.; Zhou, Y.-H.; Yu, J.-Q.; Chen, Z. Functional analysis of the Arabidopsis PAL gene family in plant growth, development, and response to environmental stress. Plant Physiol. 2010, 153, 1526–1538. [Google Scholar] [CrossRef]
- Larkcom, J. Oriental Vegetables: The Complete Guide for Garden and Kitchen; John Murray Ltd.: London, UK, 1991; ISBN 0719547814. [Google Scholar]
- Karamat, U.; Sun, X.; Li, N.; Zhao, J. Genetic regulators of leaf size in Brassica crops. Hortic. Res. 2021, 8, 91. [Google Scholar] [CrossRef]
- Chen, H.; Wang, T.; He, X.; Cai, X.; Lin, R.; Liang, J.; Wu, J.; King, G.; Wang, X. BRAD V3. 0: An upgraded Brassicaceae database. Nucleic Acids Res. 2022, 50, D1432–D1441. [Google Scholar] [CrossRef]
- Yu, H.; Li, Y.; Han, W.; Bao, L.; Liu, F.; Ma, Y.; Pu, Z.; Zeng, Q.; Zhang, L.; Bao, Z. Pan-evolutionary and regulatory genome architecture delineated by an integrated macro-and microsynteny approach. Nat. Protoc. 2024, 19, 1623–1678. [Google Scholar] [CrossRef]
- Schmitz, R.J.; Grotewold, E.; Stam, M. Cis-regulatory sequences in plants: Their importance, discovery, and future challenges. Plant Cell 2022, 34, 718–741. [Google Scholar] [CrossRef] [PubMed]
- Marand, A.P.; Eveland, A.L.; Kaufmann, K.; Springer, N.M. cis-Regulatory elements in plant development, adaptation, and evolution. Annu. Rev. Plant Biol. 2023, 74, 111–137. [Google Scholar] [CrossRef]
- Luo, Y.; Guo, Z.; Li, L. Evolutionary conservation of microRNA regulatory programs in plant flower development. Dev. Biol. 2013, 380, 133–144. [Google Scholar] [CrossRef] [PubMed]
- Zhu, Q.-H.; Helliwell, C.A. Regulation of flowering time and floral patterning by miR172. J. Exp. Bot. 2011, 62, 487–495. [Google Scholar] [CrossRef] [PubMed]
- Tang, M.; Bai, X.; Niu, L.-J.; Chai, X.; Chen, M.-S.; Xu, Z.-F. miR172 regulates both vegetative and reproductive development in the perennial woody plant Jatropha curcas. Plant Cell Physiol. 2018, 59, 2549–2563. [Google Scholar] [CrossRef]
- Dong, N.Q.; Lin, H.X. Contribution of phenylpropanoid metabolism to plant development and plant–environment interactions. J. Integr. Plant Biol. 2021, 63, 180–209. [Google Scholar] [CrossRef] [PubMed]
- Tak, Y.; Kaur, M.; Gautam, C.; Kumar, R.; Tilgam, J.; Natta, S. Phenolic biosynthesis and metabolic pathways to alleviate stresses in plants. In Plant Phenolics in Abiotic Stress Management; Springer: Berlin/Heidelberg, Germany, 2023; pp. 63–87. [Google Scholar]
- Mattus-Araya, E.; Guajardo, J.; Herrera, R.; Moya-León, M.A. ABA speeds up the progress of color in developing F. chiloensis fruit through the activation of PAL, CHS and ANS, key genes of the phenylpropanoid/flavonoid and anthocyanin pathways. Int. J. Mol. Sci. 2022, 23, 3854. [Google Scholar] [CrossRef]
- Liu, H.; Liu, Z.; Wu, Y.; Zheng, L.; Zhang, G. Regulatory mechanisms of anthocyanin biosynthesis in apple and pear. Int. J. Mol. Sci. 2021, 22, 8441. [Google Scholar] [CrossRef]
- Tang, K.; Karamat, U.; Li, G.; Guo, J.; Jiang, S.; Fu, M.; Yang, X. Integrated metabolome and transcriptome analyses reveal the role of BoGSTF12 in anthocyanin accumulation in Chinese kale (Brassica oleracea var. alboglabra). BMC Plant Biol. 2024, 24, 335. [Google Scholar] [CrossRef]
- Sharma, A.; Shahzad, B.; Rehman, A.; Bhardwaj, R.; Landi, M.; Zheng, B. Response of phenylpropanoid pathway and the role of polyphenols in plants under abiotic stress. Molecules 2019, 24, 2452. [Google Scholar] [CrossRef]
- Rivero, R.M.; Ruiz, J.M.; Garcıa, P.C.; Lopez-Lefebre, L.R.; Sánchez, E.; Romero, L. Resistance to cold and heat stress: Accumulation of phenolic compounds in tomato and watermelon plants. Plant Sci. 2001, 160, 315–321. [Google Scholar] [CrossRef] [PubMed]
- Madhusmita, P.; Neelofar, M. Low-light and its effects on crop yield: Genetic and genomic implications. J. Biosci. 2020, 45, 102. [Google Scholar]
- Liu, L.; Xie, Y.; Yahaya, B.S.; Wu, F. GIGANTEA Unveiled: Exploring Its Diverse Roles and Mechanisms. Genes 2024, 15, 94. [Google Scholar] [CrossRef]
- Pazuki, A.; Aflaki, F.; Pessarakli, M. Ultraviolet Radiation Effects on Plants: Harmful or Beneficial? In Handbook of Plant and Crop Stress, Fourth Edition; CRC Press: Boca Raton, FL, USA, 2019; pp. 451–466. [Google Scholar]
- Barros, J.; Dixon, R.A. Plant phenylalanine/tyrosine ammonia-lyases. Trends Plant Sci. 2020, 25, 66–79. [Google Scholar] [CrossRef] [PubMed]
- Yadav, V.; Wang, Z.; Wei, C.; Amo, A.; Ahmed, B.; Yang, X.; Zhang, X. Phenylpropanoid pathway engineering: An emerging approach towards plant defense. Pathogens 2020, 9, 312. [Google Scholar] [CrossRef]
- Liu, Y.; Feng, X.; Zhang, Y.; Zhou, F.; Zhu, P. Simultaneous changes in anthocyanin, chlorophyll, and carotenoid contents produce green variegation in pink–leaved ornamental kale. BMC Genom. 2021, 22, 455. [Google Scholar] [CrossRef]
- Zou, J.; Gong, Z.; Liu, Z.; Ren, J.; Feng, H. Investigation of the Key Genes Associated with Anthocyanin Accumulation during Inner Leaf Reddening in Ornamental Kale (Brassica oleracea L. var. acephala). Int. J. Mol. Sci. 2023, 24, 2837. [Google Scholar] [CrossRef] [PubMed]
- Hajam, Y.A.; Lone, R.; Kumar, R. Role of plant phenolics against reactive oxygen species (ROS) induced oxidative stress and biochemical alterations. In Plant Phenolics in Abiotic Stress Management; Springer: Berlin/Heidelberg, Germany, 2023; pp. 125–147. [Google Scholar]
- Yadav, S.; Chattopadhyay, D. Lignin: The building block of defense responses to stress in plants. J. Plant Growth Regul. 2023, 42, 6652–6666. [Google Scholar] [CrossRef]
- MacDonald, M.J.; D’Cunha, G.B. A modern view of phenylalanine ammonia lyase. Biochem. Cell Biol. 2007, 85, 273–282. [Google Scholar] [CrossRef]
- Ferrer, J.-L.; Austin, M.; Stewart, C., Jr.; Noel, J. Structure and function of enzymes involved in the biosynthesis of phenylpropanoids. Plant Physiol. Biochem. 2008, 46, 356–370. [Google Scholar] [CrossRef]
- Zhang, X.; Shen, Y.; Mu, K.; Cai, W.; Zhao, Y.; Shen, H.; Wang, X.; Ma, H. Phenylalanine ammonia lyase GmPAL1. 1 promotes seed vigor under high-temperature and-humidity stress and enhances seed germination under salt and drought stress in transgenic Arabidopsis. Plants 2022, 11, 3239. [Google Scholar] [CrossRef] [PubMed]
- Shang, Q.-M.; Li, L.; Dong, C.-J. Multiple tandem duplication of the phenylalanine ammonia-lyase genes in Cucumis sativus L. Planta 2012, 236, 1093–1105. [Google Scholar] [CrossRef] [PubMed]
- Shi, R.; Sun, Y.-H.; Li, Q.; Heber, S.; Sederoff, R.; Chiang, V.L. Towards a systems approach for lignin biosynthesis in Populus trichocarpa: Transcript abundance and specificity of the monolignol biosynthetic genes. Plant Cell Physiol. 2010, 51, 144–163. [Google Scholar] [CrossRef]
- Liu, S.; Liu, Y.; Yang, X.; Tong, C.; Edwards, D.; Parkin, I.A.; Zhao, M.; Ma, J.; Yu, J.; Huang, S. The Brassica oleracea genome reveals the asymmetrical evolution of polyploid genomes. Nat. Commun. 2014, 5, 3930. [Google Scholar] [CrossRef]
- He, Z.; Ji, R.; Havlickova, L.; Wang, L.; Li, Y.; Lee, H.T.; Song, J.; Koh, C.; Yang, J.; Zhang, M. Genome structural evolution in Brassica crops. Nat. Plants 2021, 7, 757–765. [Google Scholar] [CrossRef]
- Hu, B.; Jin, J.; Guo, A.-Y.; Zhang, H.; Luo, J.; Gao, G. GSDS 2.0: An upgraded gene feature visualization server. Bioinformatics 2015, 31, 1296–1297. [Google Scholar] [CrossRef] [PubMed]
- The Brassica rapa Genome Sequencing Project Consortium; Wang, X.; Wang, H.; Wang, J.; Sun, R.; Wu, J.; Liu, S.; Bai, Y.; Mun, J.-H.; Bancroft, I.; et al. The genome of the mesopolyploid crop species Brassica rapa. Nat. Genet. 2011, 43, 1035–1039. [Google Scholar] [CrossRef]
- Tian, J.; Xu, R.; Chang, K.; Yuan, S.; Huang, C.; Wang, J.; Li, S.; Liu, F.; Zhong, F. Identification of PAL gene in purple cabbage and functional analysis related to anthocyanin synthesis. Horticulturae 2023, 9, 469. [Google Scholar] [CrossRef]
- Feng, Y.; Huang, Q.; Zhang, R.; Li, J.; Luo, K.; Chen, Y. Molecular characterisation of PAL gene family reveals their role in abiotic stress response in lucerne (Medicago sativa). Crop Pasture Sci. 2022, 73, 300–311. [Google Scholar] [CrossRef]
- He, Y.; Zhong, X.; Jiang, X.; Cong, H.; Sun, H.; Qiao, F. Characterisation, expression and functional analysis of PAL gene family in Cephalotaxus hainanensis. Plant Physiol. Biochem. 2020, 156, 461–470. [Google Scholar] [CrossRef]
- Guo, J.; Wang, M.-H. Characterization of the phenylalanine ammonia-lyase gene (SlPAL5) from tomato (Solanum lycopersicum L.). Mol. Biol. Rep. 2009, 36, 1579–1585. [Google Scholar] [CrossRef] [PubMed]
- Poole, R.L. The TAIR database. In Plant Bioinformatics: Methods and Protocols; Humana Press: Totowa, NJ, USA, 2007; pp. 179–212. [Google Scholar]
- Chen, C.; Wu, Y.; Li, J.; Wang, X.; Zeng, Z.; Xu, J.; Liu, Y.; Feng, J.; Chen, H.; He, Y.; et al. TBtools-II: A “one for all, all for one” bioinformatics platform for biological big-data mining. Mol. Plant 2023, 16, 1733–1742. [Google Scholar] [CrossRef] [PubMed]
- Kohl, M.; Wiese, S.; Warscheid, B. Cytoscape: Software for visualization and analysis of biological networks. In Data Mining in Proteomics: From Standards to Applications; Springer Science+Business Media, LLC: Totowa, NJ, USA, 2011; pp. 291–303. [Google Scholar]
- Karamat, U.; Tabusam, J.; Khan, M.K.U.; Awan, M.J.A.; Zulfiqar, S.; Du, W.; Farooq, M.A. Genome-wide identification, characterization, and expression profiling of eukaryotic-specific UBP Family genes in Brassica rapa. J. Plant Growth Regul. 2023, 42, 3552–3567. [Google Scholar] [CrossRef]
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).