Abstract
The HNF1B gene, located on chromosome 17q12, encodes a transcription factor essential for the development of several organs. It regulates the expression of multiple genes in renal, pancreatic, hepatic, neurological, and genitourinary tissues during prenatal and postnatal development, influencing processes such as nephrogenesis, cellular polarity, tight junction formation, cilia development, ion transport in the renal tubule, and renal metabolism. Mutations that alter the function of Hnf1b deregulate those processes, leading to various pathologies characterized by both renal and extrarenal manifestations. The main renal diseases that develop are polycystic kidney disease, hypoplastic or dysplastic kidneys, structural abnormalities, Congenital Anomalies of the Kidney and Urinary Tract (CAKUT), and electrolyte imbalances such as hyperuricemia and hypomagnesemia. Extrarenal manifestations include Maturity-Onset Diabetes of the Young (MODY), hypertransaminasemia, genital and urinary tract malformations, Autism Spectrum Disorder (ASD), and other neurodevelopmental disorders. Patients with HNF1B alterations typically carry either punctual mutations or a monoallelic microdeletion in the 17q12 region. Future research on the molecular mechanisms and genotype–phenotype correlations in HNF1B-related conditions will enhance our understanding, leading to improved clinical management, genetic counseling, monitoring, and patient care.
1. Introduction
The hepatocyte nuclear factor 1-beta (HNF1B) gene encodes a member of the superfamily of homeodomain-containing transcription factors [1]. These factors interact with promoter regions, coactivators, and co-repressors to regulate gene expression [2,3,4]. HNF1B plays a crucial role in the development and function of kidneys, intestines, liver, and pancreatic beta cells [1,3,5,6,7]. In renal processes, HNF1B is essential during both embryogenesis (regulating nephrogenesis, cell polarity and cell–cell adhesion) and in the mature kidney (where it regulates electrolyte transport, primary cilia development, and renal metabolism) [3,8,9].
Located on chromosome 17q12, the HNF1B gene lies between the 36,046,434 and 36,105,096 positions (GRCh37/hg19) [10]. The 17q12 region is flanked by repetitive sequences, which promotes errors during DNA recombination. If not corrected, these errors result in structural variants such as rearrangements, deletions, or duplications [11,12,13]. The most common alteration in this region is the recurrent 17q12 microdeletion, found in 50% of patients with HNF1B mutations. The deletion size varies between 1.3 and 1.8 Mb among patients. In addition to HNF1B, this region includes several other genes also affected by the deletion [2,3,14,15], including AATF, ACACA, C17orf78, DDX52, DHRS11, DUSP14, GGNBP2, HNF1B, LHX1, MRM1, MYO19, PIGW, SYNRG, TADA2A, and ZNHIT3 [2,3,14,16]. In 30–50% of patients with HNF1B mutations, these are de novo [14,17].
The protein encoded by HNF1B consists of 557 amino acids, featuring four domains: the dimerization domain, the Pit-Oct-Unc (POU)-specific (POUS) DNA-binding domain, the POU-homeodomain (POUH) DNA-binding domain, and the transactivation C-terminus domain. The latter is crucial for coactivator recruitment and transcriptional regulation [10,18,19] (Figure 1a).
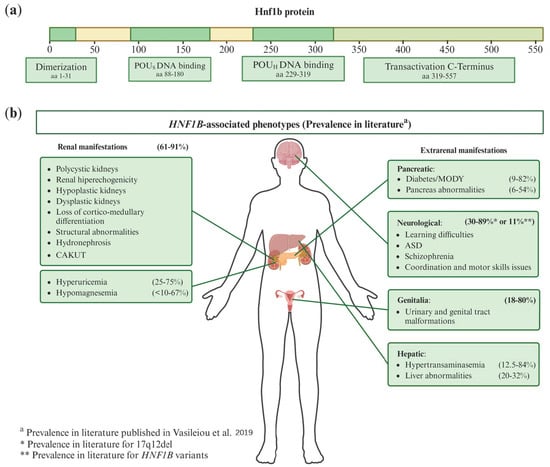
Figure 1.
(a) Scheme of canonical HNF1B transcript (NM_000458.4) protein domains. aa: amino acid; POUS: Pit-Onc-Unc specific; POUH: Pit-Onc-Unc homeodomain. (b) Phenotypic manifestations associated with HNF1B and their prevalence in the literature. There is a substantial variability in both renal and extrarenal manifestations among HNF1B patients. These patients may exhibit various combinations of these phenotypes. The prevalence of each manifestation shown in the figure has been derived from the article by Vasileiou et al. [20], where they reviewed the prevalence ranges for each condition from 82 published studies on HNF1B patients with clinical data. CAKUT: Congenital Anomalies of the Kidney and Urinary Tract; MODY: Maturity-Onset Diabetes of the Young; ASD: Autism Spectrum Disorder.
This gene presents complex animal models, as monoallelic deletion does not produce a clear phenotype in mouse models, whereas biallelic deletion results in a much more severe phenotype than that observed in humans, even causing early embryonic lethality. Niborski L et al. [21] generated a murine model with a human mutation at the intron-2 splice donor site, which exhibits a phenotype more similar to that in humans. Here, the genes previously linked to Hnf1b (Wnt9b, Pax2, Pkd2, Bicc1, Crb3, Kif12, Cys1, Glis2, and Glis3) were either modestly decreased or unaffected. Additionally, they observed that HNF1B-regulated gene expression varied by developmental stage; for example, Umod, Tmem27, and Pkhd1 were significantly downregulated at E17.5 but not at P1 [21]. This reflects the complexity of the animal models for this gene and the need for ongoing research to better understand HNF1B function and its correlation with its phenotype in humans.
This review summarizes the multifaceted roles of the HNF1B gene in renal physiology, providing an overview of the current knowledge and highlighting the gene’s significance in kidney development and function.
2. HNF1B-Associated Diseases and Clinical Significance
HNF1B-associated disease follows an autosomal dominant inheritance [22,23]. While biallelic germ line inactivation of HNF1B is embryonically lethal [24,25], monoallelic pathogenic mutations cause a variety of disorders in various organs. These disorders often manifest as diabetes and renal conditions during prenatal, postnatal, and adulthood stages [14].
The first described HNF1B-associated renal disorder was Renal Cysts And Diabetes syndrome (RCAD), characterized by renal cysts and Maturity-Onset Diabetes of the Young type 5 (MODY5), with high phenotypic variability [2,3]. Renal ultrasound scans of affected patients typically reveal combinations of abnormalities, including polycystic, hyperechogenic, hypoplastic and dysplastic kidneys, loss of cortico-medullary differentiation, structural abnormalities, and hydronephrosis [14,20,22,26,27].
In addition to RCAD, other common renal disorders associated with HNF1B include Congenital Anomalies of the Kidney and Urinary Tract (CAKUT) [2,3] and Autosomal Dominant Tubulointerstitial Kidney Disease type HNF1B (ADTKD-HNF1B) [28] These are frequently associated with electrolyte disturbances such as hyperuricemia and hypomagnesemia [14,15,19,20,22]. Hypomagnesemia can occasionally lead to a misdiagnosis of Gitelman syndrome [29]. A large proportion of patients reach at least chronic kidney disease (CKD) stage 3 in adulthood, with some developing renal failure [19].
Beyond the kidneys, HNF1B mutations can lead to extrarenal manifestations affecting the pancreas (notably MODY [14,15]), liver (hypertransaminasemia, liver cysts), genitalia (genital tract malformations), and neurodevelopment (learning difficulties, autism spectrum disorder, schizophrenia, coordination and motor skills issues or neurodevelopment delay) [15,20,26,30].
Patients may present with various combinations of these phenotypes, as shown in Figure 1b. As illustrated, renal manifestations are the most prevalent, with 61% to 91% of patients displaying at least one renal symptom listed in the figure. The data about prevalence in the literature were derived from the Vasileiou et al. paper [20], where they reviewed 82 published articles containing clinical information on HNF1B patients. Differences in study methodologies and the significant phenotypic variability caused by HNF1B contribute to the wide range of prevalence rates reported [20].
The phenotypic variability in HNF1B-related diseases may be partially explained by the range of biological processes and genes regulated by HNF1B at both prenatal and postnatal stages. Section 4 of this review explores these mechanisms and their role in disease development.
In summary, patients with HNF1B mutations display considerable phenotypic diversity, ranging from CAKUT to electrolyte imbalances and extrarenal conditions. The specific symptomatology remains unpredictable, and it is unclear what drives these differences. One possibility is that the type of genetic alteration influences the affected biological pathways, given the regulatory role of HNF1B over numerous genes involved in diverse biological processes. Thus, different mutations may disrupt distinct pathways, resulting in diverse clinical outcomes. Recent studies have correlated the type of genetic alteration (17q12 deletion or punctual HNF1B variants) with patient symptoms. It has been suggested that patients with punctual HNF1B mutations tend to have more severe renal phenotypes and some additional extrarenal manifestations, whereas those with the 17q12 deletion exhibit a broader range of extrarenal symptoms and lighter renal phenotype [19,20,26,31].
Diagnosing HNF1B-associated diseases is challenging due to its broad phenotypic manifestations and significant intrafamilial variability in renal phenotypes [14,17]. Further research into the mechanisms underlying this phenotypic variability could improve diagnostic strategies and patient management.
3. Hnf1b Protein Domains and Transcriptional Complexes
The structure of the Hnf1b protein consists of four domains (Figure 1a): the dimerization domain (residues 1–31), the Pit-Oct-Unc (POU)-specific DNA-binding domain (POUS) (residues 88–180), the POU-homeodomain (POUH) DNA-binding domain (residues 229–319), and the C-terminal transactivation domain (residues 319–557). The most genetically conserved regions of HNF1B are the dimerization and POU-DNA-binding domains [18,32]. POUS has at least two α-helices. POUH recognizes the consensus sequence 5′-GTTAATNATTAAC-3′ [2,3,4], and forms three α-helices with the third helix functioning as the DNA recognition helix [33]. The POUS and POUH domains cooperate to enhance the DNA-binding affinity and specificity [32,33]. The transactivation C-terminus domain is important for the protein to function [32,33,34]; Tholen L. et al. [34] showed that a Hnf1b lacking this domain failed to induce gene expression due to its inability to bind essential cofactors [34].
To regulate gene expression, Hnf1b forms DNA-binding transcriptional complexes by interacting with other molecules. In these complexes, Hnf1b can function as a homodimer (two Hnf1b molecules) or as a heterodimer with its paralogue Hnf1a [1,23,24], with which it shares 70% sequence homology [35]. Notably, alterations in HNF1A are also associated with diabetes [36,37,38,39]. Depending on the target gene, other molecules may join the complex [40]. Hnf1b’s genetic regulation also depends on the tissue and developmental stage, this can be observed in the transcriptomic analysis of Hnf1b-altered mice of Niborski L et al. [21] during prenatal (E14.5, E15.5 y E17.5) and postnatal (P1) stages. Their findings revealed differences in the genetic regulation between stages [21].
DNA-binding transcriptional complexes are formed by different combinations of key molecules such as Pcbd1, Pcbd2, CBP, P/CAF, Zyxin, NCoR, and Hdac1 [3,40,41,42,43,44,45].
One of these key molecules is Pcbd1, which interacts with Hnf1b in organs such as the kidney, pancreas, and liver during embryogenesis [40,43,44]; an example of a gene regulated by a transcriptional complex that includes Pcbd1 is FXYD2 [40,44].
Another key molecule is Pcbd2, which is mainly expressed in the kidney, lung, spleen, and adipose tissue and regulates genes different from those influenced by Pcbd1, such as KCNJ16 [44].
The key molecules CREB-binding protein (CBP) and P300/CBP-Associated Factor (P/CAF) are coactivators with intrinsic histone acetyltransferase activity, and they are required for Hnf1a- and Hnf1b-mediated transcription [42].
Choi et al. [41] identified zyxin as another key molecule that interacts with Hnf1b in renal epithelial cells [41,45]. It potentially functions as a scaffolding protein that facilitates the assembly of transcription factors and coactivators on gene promoters [41]. Zyxin can interact together with the coactivator CBP, enhancing Hnf1b’s transcriptional activity [41].
The key molecules Nuclear Receptor Corepressor (NCoR) and histone deacetylase 1 (Hdac1) can negatively regulate Hnf1a [42].
Due to the diverse molecular combinations forming its DNA-binding transcriptional complexes, Hnf1b has a broad regulatory scope, controlling the expression of numerous genes with various functions [21]. The composition of these complexes varies depending on the expressed genes, organ, cell type, and developmental stage [21,44,46].
Key molecules forming these complexes could be a significant area of research, potentially leading to the identification of cofactor genes critical for diagnosing HNF1B-related pathologies.
4. Hnf1b Renal Functions in Kidney Development and Physiology
As previously mentioned, Hnf1b is a transcription factor that plays a pivotal role in various aspects of kidney development and physiology. Figure 2 illustrates the genes directly regulated by Hnf1b. The mechanistic action of Hnf1b in renal physiology is described in the following four subsections, listed in Table 1.
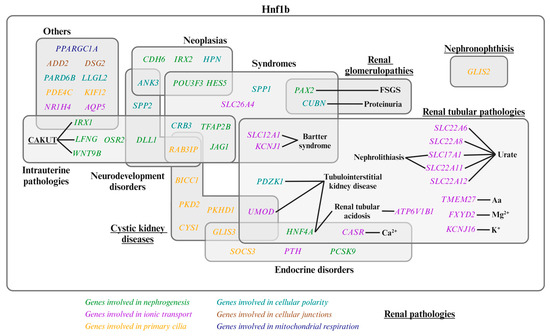
Figure 2.
Genes regulated by Hnf1b and their associated diseases. Genes involved in nephrogenesis, ion transport, primary cilia, cellular polarity, cellular junctions, and mitochondrial respiration are shown in green, purple, orange, light blue, brown, and dark blue, respectively. Underlined pathologies are renal pathologies [1,2,3,23,34,45,47,48,49]. CAKUT: Congenital Anomalies of the Kidney and Urinary Tract; FSGS: Focal Segmental Glomerulosclerosis; Aa: amino acid.

Table 1.
Index of mechanistic action of Hnf1b in renal physiology.
4.1. Hnf1b in Nephrogenesis
Nephrogenesis is the process by which nephrons develop during embryonic stages until their maturation, at which point they become functional. This process begins with the secretion of signaling molecules from the metanephric mesenchyme (MM) to the ureteric bud (UB), triggering UB outgrowth and the formation of the collecting duct system and ureter (Figure 3a) [2,47,48,50]. The steps of this process, illustrated in Figure 3, involve interactions between transcription factors, promoter regions, coactivators, and co-repressors to regulate gene expression [1,2,3,4]: (1) Induction of nephron progenitor cells to form the renal vesicle (RV) [47,48,50]; (2) morphogenesis and patterning of the RV, which differentiates into the comma-shaped body (CSB) and then the S-shaped body (SSB) [47,48,50]; and (3) final differentiation of the SSB into a mature nephron, which includes the proximal segment (subdivided into Bowman’s capsule, podocyte, and proximal tubule); the intermediate tubule (containing the loop of Henle); and the distal tubule (Figure 3b,c) [48].
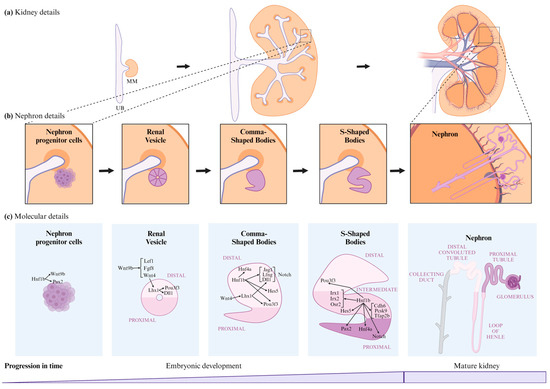
Figure 3.
Nephrogenesis stages and Hnf1b’s role in this process. (a) Kidney details of renal development from embryonic to mature kidneys. (b) Nephron details of renal development from embryonic to mature nephrons. (c) Molecular details and Hnf1b’s role in renal development from embryonic to mature nephrons. UB, Ureteric Bud; MM, Metanephric Mesenchyme.
- 1.
- Induction of nephron progenitor cells to form the RV: Hnf1b is implicated in several stages of renal development, regulating a significant proportion of the mechanisms underlying these processes (Figure 3c). Hnf1b conditional inactivation in murine nephron progenitors showed that Hnf1b acts upstream of Wnt9b in mice [3,48]. However, research by Niborski L et al. [21] showed minimal impact on Wnt9b expression in a mouse model with an identified HNF1B human mutation at the intron-2 splice donor site [21]. Wnt9b plays a crucial role in nephrogenesis, particularly in inducing the mesenchyme-to-epithelium transition (MET). Wnt9b is expressed uniformly in the UB epithelium, with increased expression in areas where RVs will form [47,51]. MET is responsible for forming the RV from nephron progenitor cells [2,9,47,51,52,53,54] and involves a stepwise assembly of intercellular junctions and de novo establishment of apical–basal polarity to form the RV, the first polarized epithelial precursor of the nephron [55]. This suggests that Hnf1b plays a crucial role in initiating nephrogenesis [3,48].Wnt9b signaling activates the expression of the differentiation markers Lef1, Fgf8, and Wnt4 in the surrounding MM [47]. Lef1 and Wnt4 display polarized expression patterns in the distal RV, regulating its early polarity [47]; Wnt4 also triggers the expression of Lhx1, the next transcriptional regulator [47,52,56,57]. Lhx1 drives RV progression to the CSB by activating Dll1 and Pou3f3 (also known as Brn1) expression in the RV. Lhx1 is also involved in the proximo-distal differentiation of the RV, CSB, and SSB [47,48,58]. Hnf1b also regulates PAX2, a transcription factor critical for the MET of nephron progenitors, maintaining nephric duct epithelial polarity and SSB differentiation [3,8,47,48,59,60,61,62]. PAX2 defects have been linked to focal segmental glomerulosclerosis (FSGS) [23].
- 2.
- Differentiation into CSB and its progression to SSB: in the CSB, Pou3f3 expression is regulated by Lhx1 in the distal RV and by Hnf1b in the distal CSB regions, as well as the proximal and bulge regions of the SSB [8,47,52]. Pou3f3 is involved in elongating and differentiating the loop of Henle and forming the distal convoluted tubule (DCT) [8,48,52]. Hnf1b also activates Notch pathway components such as Dll1, Jag1, Lfng [52], and Hes5 [8], which are crucial for inducing differentiation and polarization in nephrogenesis [8,47]. Dll1 and Jag1 are ligands for Notch receptors, Dll1 expression is regulated by both Lhx1 and Hnf1b [8,47,48,52], and Jag1 is regulated by Hnf1a and Hnf1b [8,52]. Lfng, regulated by Hnf1b, enhances its expression in the distal region of the CSB and in the proximal region of the SSB [47,48,52]. Defective expression in Notch components significantly reduces proximal tubule formation [8]. In addition, Hes5 expression was observed to change in murine Hnf1b mutants [8], and it is specially expressed in the CSB and epithelial cells that form the bulge between mid and lower limb of SSBs [8,48] (Figure 3c).
- 3.
- Progression from SSB to mature nephron: Hnf1b regulates the expression of IRX1, IRX2, and OSR2 genes in the SSB intermediate region, these are involved in tubule differentiation and expansion [8,47,48,52]. Hnf1b binds to the promoter regions of genes expressed in the proximal/intermediate tubule of SSB, including CDH6, PCSK9, and TFAP2B [48]. CDH6 is similarly expressed in RV and proximal tubule precursor cells [8]. Hnf1b activates HNF4A transcription in the distal region of the CSB and the proximal region of the SSB [2,8,48,52]. HNF4A encodes a nuclear transcription factor required for proximal tubule development [48,52]. In the Hnf1b-altered model, Hnf4a is downregulated during all nephrogenesis stages and in mature kidneys [21]. This gene has been associated with diabetes and renal cyst development [36,38,63,64].In the Heliot C. et al. [48] study, the conditional inactivation of Hnf1b in murine nephron progenitors led to rudimentary nephrons comprising a glomerulus connected to the collecting system by a short tubule with distal fates. This defect was preceded by strong downregulation of the Notch pathway components (Lfng, Dll1, and Jag1) and Irx1/2 factors, which are potential regulators of proximal and loop of Henle segment fates (Figure 3c).
In summary, transcription factors such as Hnf1b do not regulate single genes but entire gene networks [34], and Hnf1b plays a central role in initiating nephrogenesis and driving the differentiation and expansion of the RV into a mature nephron [48,60].
4.2. Hnf1b Implication in Apical-Basolateral Polarity, Tight Junctions, Primary Cilia Development and Cyst Formation
Hnf1b-regulated components discussed previously are essential for the process of nephrogenesis, and some of them also play a role in maintaining polarity in different metanephron regions.
Nephrons are composed of renal epithelial cells organized in a monolayer, connected by adherent and tight junctions. Their proper functioning depends on maintaining apical–basolateral polarity [55], which ensures the correct positioning of cytoskeletal and junctional components [34]. Massa et al. [8] observed defects in the CSB and SSB stages in the absence of Hnf1b, highlighting its role in epithelial maturation and maintenance [8]. Moreover, altered polarization, differentiation, and organization of renal epithelial cells were noted in Hnf1b-inactivated models [34,60,65].
Par and Crumbs complexes are constituted by core proteins involved in establishing apical polarity, and are evolutionarily conserved [34,66]. The Par complex consists of Par3, Par6, Cdc42, and aPKC, while the Crumbs complex includes Crb3, Pals1, and Patj [66,67,68]. Basolateral polarity, on the other hand, is primarily regulated by the Scribble proteins: Scrib, Dlg, and Llgl [34,68].
Tholen et al. [34] identified Hnf1b-regulated genes involved in cell polarity, cell–cell junctions, and cytoskeletal integrity using a mouse DCT cell line. These genes include PARD6B, CRB3, LLGL2, ANK3, ADD2, RAB3IP, and DSG2 [34]. Other Hnf1b-regulated genes contributing to apical–basal polarity include ATP6V1B1, SLC26A4, PDZK1, AQP5, and HPN [21,60]. They also reported reduced tight junction integrity and changes in markers of polarized epithelium such as Cdh16, Pkhd1, and Cys1 [60], alongside aberrant localization of Muc1, basal Laminin, and apical aPKC [34,48,60].
In the study by Niborski L et al. [21], reduced Hnf1b expression led to the downregulation of Spp1 and Spp2, proteins crucial for early PT stages, particularly in cell adhesion and the extracellular environment [1,5,21]. Another early PT marker downregulated under these conditions is Cubn, a gene linked to chronic benign proteinuria and Imerslund–Grasbeck syndrome [1,21,23,69].
Several studies suggest that defects in renal cell polarity can affect primary cilia maintenance, and ciliary dysfunction is closely associated with renal cyst formation and failure [2,3,34,60,70,71]. Indeed, renal cysts are frequently observed in HNF1B patients and animal models [14,19,72]. Otherwise, Hnf1b regulates genes implicated in primary cilium development [2]; they influence cell proliferation, migration, apoptosis, planar cell polarity, and differentiation via pathways such as Wnt, cAMP, and mTOR [3,34,72,73]. Key primary cilium genes regulated by Hnf1b include:
- PKD2: encodes Polycystin 2, a Ca2+-permeable cation channel [1,2]. PKD2 mutations cause Autosomal Dominant Polycystic Kidney Disease (ADPKD) [3,23]. This is due to reduced Ca2+ entry and activation of the Ca2+-inhibitable adenylyl cyclases (AC5 and AC6), which elevates cAMP levels. This rise in cAMP stimulates cell proliferation and fluid secretion, promoting cyst growth [3,74].
- PKHD1: encodes a protein located in the primary cilia [2,3]; Hnf1b regulates this gene, particularly in collecting ducts (CD) [3]. PKHD1 is associated with Autosomal Recessive Polycystic Kidney Disease (ARPKD) [3,23].
- PDE4C: catabolizes cAMP in the primary cilia and is downregulated in HNF1B mutant cells, leading to increased cAMP levels and subsequent cyst formation [3,74].
- UMOD: involved in electrolyte transport in the thick ascending limb (TAL) [2]; UMOD is linked to ADTKD and ciliary function [3].
- KIF12: part of the microtubule cytoskeleton [1,2,49], expressed in primary cilia and mitotic spindles [2,34]. It is downregulated in HNF1B patients [49], although no significant effect was observed in the murine model by Niborski L et al. [21].
- HNF4A: plays a role in various kidney processes, including PT development and cystogenesis, by modulating PKD1 expression [63].
- SOCS3: negatively regulated by Hnf1b; its upregulation impairs tubule formation [3] and is highly upregulated in human polycystic kidneys [3,75].
- BICC1: an RNA-binding protein that modulates translation during embryonic development and is associated with renal cystic dysplasia [1,23,76].
- CYS1: involved in congenital polycystic kidney disease [1,21,69].
- GLIS2: encodes a transcription factor linked to nephronophthisis [3,23].
- GLIS3: implicated in pancreatic beta cells, thyroid, eye, liver, and kidney development [1]. It is also involved in polycystic disease affecting both the kidneys [21,69,77] and pancreas [78].
4.3. Hnf1b Regulates Ion Transport in Kidney
In mature kidneys, Hnf1b regulates genes responsible for solute transport [23]. Alterations in HNF1B often lead to ADTKD-HNF1B, which is characterized by electrolyte imbalances such as hypomagnesemia, hyperparathyroidism, hyperuricemia, and hypocalciuria. These imbalances arise from Hnf1b’s transcriptional regulation of ion transport gene networks (Figure 4) [2,3,11,19,79].
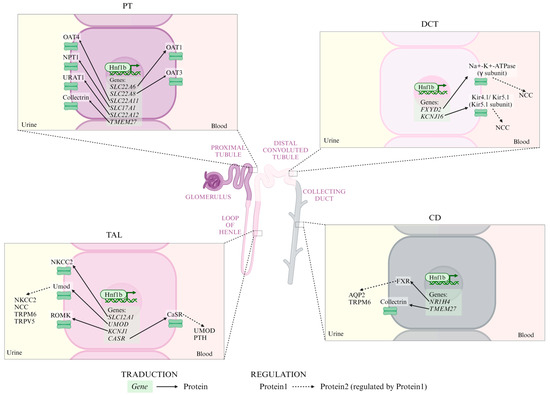
Figure 4.
Ionic transporters regulated by Hnf1b in each tubular segment. Continuous arrows represent transduction processes from gene to protein; dashed arrows represent regulation processes from protein1 to protein2. PT: proximal tubule; TAL: thick ascending limb; DCT: distal convoluted tubule; CD: collecting duct; OAT4: organic anion transporter 4; NPT1: Na+-phosphate transporter 1; URAT1: urate transporter; OAT1: organic anion transporter 1; OAT3: organic anion transporter 3; NKCC2: Na+-K+-Cl- co-transporter 2; Umod: uromodulin; ROMK: renal outer medullary potassium channel; NCC: Na+-Cl- co-transporter; TRPM6: transient receptor potential melastatin type 6; TRPV5: TRP vanilloid type 5; CaSR: calcium-sensing receptor; PTH: parathyroid hormone; FXR: farnesoid X receptor; and AQP2: aquaporin 2.
In the proximal tubule (PT), Hnf1b regulates expression of:
- Organic anion transporters (SLC22A6, SLC22A8, and SLC22A11 genes) [2,3,80,81,82].
- Na+-phosphate transporter 1 (SLC17A1 gene) [2,3,83].
- Renal urate transporter (SLC22A12 gene) [2,3,84].
- TMEM27, which encodes collectrin and enhances amino acid transporter surface expression. TMEM27 is also expressed in the CD [85]. Its expression is downregulated in Hnf1b-altered mice [21]. Tmem27 disruption in mice leads to a severe defect in renal amino acid uptake [3].
Most HNF1B patients do not show a strong PT phenotype. This may be due to Hnf1a compensation, which is expressed exclusively in the PT; other nephron segments do not benefit from this compensatory action [2,86].
In the TAL, Hnf1b regulates:
- SLC12A1, encoding the Na+-K+-Cl- co-transporter 2 (NKCC2) [2,48], which is essential for Na+ and Cl- transport across the apical membrane [3] and paracellular divalent cation transport [2]. Mutations in this gene are linked to Bartter syndrome [23].
- UMOD, which encodes uromodulin, activates NKCC2, Na+-Cl- co-transporter (NCC) [87], transient receptor potential melastatin type 6 (TRPM6) [88] and TRP vanilloid type 5 (TRPV5) [89]. Reduced uromodulin expression is associated with medullary cyst formation and renal electrolyte imbalances [2,90].
- KCNJ1, which encodes a K+ channel and is associated with Bartter syndrome [1,21,23].
- CASR (calcium-sensing receptor, CaSR), a negative regulator of UMOD [2,91]. Decreased CaSR expression is expected to elevate blood calcium levels [2,3,11,92]. In the parathyroid gland, PTH expression can be repressed by Hnf1b or CaSR [2,93]. PTH also inhibits uric acid secretion via ABCG2 downregulation [2].
In the DCT, Hnf1b regulates:
- FXYD2 encodes the γ subunit of the Na+-K+-ATPase. Mutations in HNF1B and FXYD2 mainly result in hypomagnesemia, hypocalciuria, and basolateral membrane depolarization, leading to increased intracellular chloride levels and inhibiting NCC [2,3,14,24,79,94,95].
- KCNJ16, encoding the Kir5.1 subunit of the basolateral Kir4.1/Kir5.1 K+ channel, crucial for K+ recycling and Na+-K+-ATPase activity. Dysfunction of this channel leads to basolateral membrane depolarization and reduced NCC activity [2,96,97].
In the CD, Hnf1b regulates:
- NR1H4, encoding farnesoid X receptor (FXR), essential for urine concentration by regulating AQP2 expression [2,98]. FXR also directly activates TRPM6 expression, contributing to Mg homeostasis [2,99].
- TMEM27, which encodes an amino acid transporter (collectrin) which is also expressed in PT [85].
4.4. Role of Hnf1b in Intrarenal Metabolism: Mitochondrial Respiration and Cholesterol
A study by Casemayou et al. [49] identified PPARGC1A as a gene directly regulated by Hnf1b, as chromatin immunoprecipitation confirmed Hnf1b binding to PPARGC1A promoter, and inhibition or overexpression of Hnf1b led to decreased [45] or increased PPARGC1A expression, respectively [49]. PPARGC1A is a transcriptional coactivator that regulates mitochondrial biogenesis [1]. Its implication in the mitochondrial biogenesis mechanism provides an explanation for the observed metabolic change with Hnf1b inactivation or deletion in mouse PT cells [45,49], human cells [45], and kidney samples from patients with pathogenic HNF1B alterations [14]. The metabolic changes observed are similar to the Warburg effect seen under hypoxic conditions, and are characterized by increased lipid accumulation, oxidative stress modulators, lactate production, and decreased Ppargc1a and choline kinase-alpha expression, along with reduced ATP production [45,49].
This way, Hnf1b regulates the bioenergetic metabolism and mitochondrial morphology of renal tubular epithelial cells [2,3,45,49]. Given the kidney’s high energy demands to sustain basal metabolism and the electrochemical gradients necessary for active ion transport [2,100,101,102,103], compromised ATP production due to HNF1B pathogenic alterations can severely disrupt transporter function. This disruption can lead to energy metabolism alteration, fibrosis, acute kidney injury, electrolyte imbalances resembling a Gitelman-like phenotype, and progression to CKD [102,104].
Hnf1b also plays a role in renal cholesterol metabolism by directly regulating genes involved in cholesterol biosynthesis [2,3,105], such as PCSK9, a key gene which is crucial for nephrogenesis and internalization of low-density lipoprotein (LDL) receptors [45,106]. Hnf1b inactivation reduces cholesterol biosynthesis while increasing cholesterol uptake in the kidneys, potentially altering lipid metabolism [2,3,105].
5. Conclusions and Future Perspectives
Throughout this review, we have highlighted the diverse renal functions regulated by Hnf1b, including nephrogenesis, tubular elongation, epithelial cell polarization, tight junctions, primary cilia structure, ion transport, mitochondrial respiration, and cholesterol metabolism.
Genetic variants altering Hnf1b function lead to dysregulation of its target genes, resulting in a spectrum of renal and extrarenal pathologies. Figure 2 illustrates the Hnf1b-regulated genes with their associated diseases.
The variability in phenotype manifestation, progression, and severity is notable among individuals with HNF1B alterations. Studies have shown that both 17q12 deletions and point mutations in HNF1B cause MODY and renal anomalies (Figure 2). However, point mutations tend to result in a more severe and earlier renal phenotype, while 17q12 deletions are associated with a broader range of renal and non-renal disorders, including liver and psychiatric conditions [12,14,19,26,27,30,31].
Investigating the molecules that constitute the Hnf1b transcriptional complex in renal tissues is also essential. Even if HNF1B itself is unaltered, incomplete transcription complexes may still cause dysregulation and associated manifestations. Identifying these components and incorporating them into genetic screening could improve diagnostic accuracy and patient outcomes.
In conclusion, HNF1B is a key regulator, especially in prenatal and postnatal kidneys, controlling genes involved in several physiological functions mentioned previously. Genetic HNF1B alterations can affect all of these processes, triggering different renal and extrarenal diseases. Further research into animal models and genotype–phenotype correlations in HNF1B-related disease is crucial. Understanding these correlations can help elucidate the full spectrum of the disease and guide more personalized treatment approaches.
Author Contributions
Conceptualization, N.C. and M.Á.G.-G.; formal analysis, E.S.-C. and N.C.; investigation, E.S.-C.; resources, M.Á.G.-G.; writing—original draft preparation, E.S.-C.; writing—review and editing, N.C. and M.Á.G.-G.; visualization, E.S.-C., N.C. and M.Á.G.-G.; supervision, N.C. and M.Á.G.-G.; project administration, M.Á.G.-G.; funding acquisition, M.Á.G.-G. All authors have read and agreed to the published version of the manuscript.
Funding
This study was supported by the Instituto de Salud Carlos III (ISCIII) (REDinREN fund RD016/0009, to M.Á.G.G.; RICORS2040 RD21/0005/0020 funded by European Union—NextGenerationEU, Mecanismo para la Recuperación y la Resiliencia (MRR) to M.Á.G.G.; and projects PI15/01467, PI18/0037 and PI22/00227 co-funded by the European Union. to M.Á.G.G.); the Senefro Foundation (SEN2021_2 to M.Á.G.G.); the Xunta de Galicia/GAIN (Grupos con potencial de crecemento-GPC fund IN607B 2016/020 and IN607B 2023/07 to M.Á.G.G.); and salary support from the Xunta de Galicia (Predoctoral fellowships, to E.S.C.) and ISCIII through PMP21-000109 (Predoctoral research, to E.S.C.).
Institutional Review Board Statement
Not applicable.
Informed Consent Statement
Not applicable.
Data Availability Statement
No new data were created or analyzed in this study. Data sharing is not applicable to this article.
Acknowledgments
The authors express their gratitude to PKDcore for providing materials that contributed to the knowledge of cystic disease together with translational research, and to the Art for Dent association for their donations supporting our research group.
Conflicts of Interest
The authors declare no conflicts of interest.
References
- GeneCards—Human Genes|Gene Database|Gene Search. Available online: https://www.genecards.org/ (accessed on 23 May 2024).
- Tholen, L.E.; Hoenderop, J.G.J.; de Baaij, J.H.F. Mechanisms of ion transport regulation by HNF1β in the kidney: Beyond transcriptional regulation of channels and transporters. Pflugers Arch. 2022, 474, 901–916. [Google Scholar] [CrossRef] [PubMed]
- Ferrè, S.; Igarashi, P. New insights into the role of HNF-1β in kidney (patho)physiology. Pediatr. Nephrol. 2019, 34, 1325–1335. [Google Scholar] [CrossRef] [PubMed]
- Rauluseviciute, I.; Riudavets-Puig, R.; Blanc-Mathieu, R.; Castro-Mondragon, J.A.; Ferenc, K.; Kumar, V.; Lemma, R.B.; Lucas, J.; Chèneby, J.; Baranasic, D.; et al. JASPAR 2024: 20th anniversary of the open-access database of transcription factor binding profiles. Nucleic Acids Res. 2024, 52, D174–D182. [Google Scholar] [CrossRef] [PubMed]
- Gene Ontology Resource. Available online: https://geneontology.org/ (accessed on 9 May 2024).
- Coffinier, C.; Barra, J.; Babinet, C.; Yaniv, M. Expression of the vHNF1/HNF1β homeoprotein gene during mouse organogenesis. Mech. Dev. 1999, 89, 211–213. [Google Scholar] [CrossRef]
- Coffinier, C.; Gresh, L.; Fiette, L.; Tronche, F.; Schütz, G.; Babinet, C.; Pontoglio, M.; Yaniv, M.; Barra, J. Bile system morphogenesis defects and liver dysfunction upon targeted deletion of HNF1β. Development 2002, 129, 1829–1838. [Google Scholar] [CrossRef]
- Massa, F.; Garbay, S.; Bouvier, R.; Sugitani, Y.; Noda, T.; Gubler, M.-C.; Heidet, L.; Pontoglio, M.; Fischer, E. Hepatocyte nuclear factor 1β controls nephron tubular development. Development 2013, 140, 886–896. [Google Scholar] [CrossRef]
- Lokmane, L.; Heliot, C.; Garcia-Villalba, P.; Fabre, M.; Cereghini, S. vHNF1 functions in distinct regulatory circuits to control ureteric bud branching and early nephrogenesis. Development 2010, 137, 347–357. [Google Scholar] [CrossRef]
- National Center for Biotechnology Information (NCBI) Gene: HNF1B HNF1 Homeobox B [Homo sapiens (human)]. Available online: https://www.ncbi.nlm.nih.gov/gene/6928 (accessed on 20 May 2024).
- Viering, D.H.H.M.; de Baaij, J.H.F.; Walsh, S.B.; Kleta, R.; Bockenhauer, D. Genetic causes of hypomagnesemia, a clinical overview. Pediatr. Nephrol. 2017, 32, 1123–1135. [Google Scholar] [CrossRef]
- Mefford, H.C.; Clauin, S.; Sharp, A.J.; Moller, R.S.; Ullmann, R.; Kapur, R.; Pinkel, D.; Cooper, G.M.; Ventura, M.; Ropers, H.H.; et al. Recurrent Reciprocal Genomic Rearrangements of 17q12 Are Associated with Renal Disease, Diabetes, and Epilepsy. Am. J. Hum. Genet. 2007, 81, 1057–1069. [Google Scholar] [CrossRef]
- Bellanné-Chantelot, C.; Clauin, S.; Chauveau, D.; Collin, P.; Daumont, M.; Douillard, C.; Dubois-Laforgue, D.; Dusselier, L.; Gautier, J.-F.; Jadoul, M.; et al. Large Genomic Rearrangements in the Hepatocyte Nuclear Factor-1β (TCF2) Gene Are the Most Frequent Cause of Maturity-Onset Diabetes of the Young Type 5. Diabetes 2005, 54, 3126–3132. [Google Scholar] [CrossRef]
- Faguer, S.; Decramer, S.; Chassaing, N.; Bellanné-Chantelot, C.; Calvas, P.; Beaufils, S.; Bessenay, L.; Lengelé, J.P.; Dahan, K.; Ronco, P.; et al. Diagnosis, management, and prognosis of HNF1B nephropathy in adulthood. Kidney Int. 2011, 80, 768–776. [Google Scholar] [CrossRef] [PubMed]
- Dubois-Laforgue, D.; Cornu, E.; Saint-Martin, C.; Coste, J.; Bellanné-Chantelot, C.; Timsit, J. Diabetes, Associated Clinical Spectrum, Long-term Prognosis, and Genotype/Phenotype Correlations in 201 Adult Patients With Hepatocyte Nuclear Factor 1B ( HNF1B ) Molecular Defects. Diabetes Care 2017, 40, 1436–1443. [Google Scholar] [CrossRef] [PubMed]
- Nassar, L.R.; Barber, G.P.; Benet-Pagès, A.; Casper, J.; Clawson, H.; Diekhans, M.; Fischer, C.; Gonzalez, J.N.; Hinrichs, A.S.; Lee, B.T.; et al. The UCSC Genome Browser database: 2023 update. Nucleic Acids Res. 2023, 51, D1188–D1195. [Google Scholar] [CrossRef] [PubMed]
- Bockenhauer, D.; Jaureguiberry, G. HNF1B-associated clinical phenotypes: The kidney and beyond. Pediatr. Nephrol. 2016, 31, 707–714. [Google Scholar] [CrossRef] [PubMed]
- Hojny, J.; Bartu, M.; Krkavcova, E.; Nemejcova, K.; Sevcik, J.; Cibula, D.; Fryba, V.; Plincelnerova, L.; Dundr, P.; Struzinska, I. Identification of novel HNF1B mRNA splicing variants and their qualitative and semi-quantitative profile in selected healthy and tumour tissues. Sci. Rep. 2020, 10, 6958. [Google Scholar] [CrossRef]
- Buffin-Meyer, B.; Richard, J.; Guigonis, V.; Weber, S.; König, J.; Heidet, L.; Moussaoui, N.; Vu, J.-P.; Faguer, S.; Casemayou, A.; et al. Renal and extra-renal phenotypes in patients with HNF1B variants and chromosome 17q12 micro deletions. Kidney Int. Rep. 2024, 8, 2514–2526. [Google Scholar] [CrossRef]
- Vasileiou, G.; Hoyer, J.; Thiel, C.T.; Schaefer, J.; Zapke, M.; Krumbiegel, M.; Kraus, C.; Zweier, M.; Uebe, S.; Ekici, A.B.; et al. Prenatal diagnosis of HNF1B-associated renal cysts: Is there a need to differentiate intragenic variants from 17q12 microdeletion syndrome? Prenat. Diagn. 2019, 39, 1136–1147. [Google Scholar] [CrossRef]
- Niborski, L.L.; Paces-Fessy, M.; Ricci, P.; Bourgeois, A.; Magalhães, P.; Kuzma-Kuzniarska, M.; Lesaulnier, C.; Reczko, M.; Declercq, E.; Zürbig, P.; et al. Hnf1b haploinsufficiency differentially affects developmental target genes in a new renal cysts and diabetes mouse model. Dis. Model Mech. 2021, 14, dmm047498. [Google Scholar] [CrossRef]
- Clissold, R.L.; Hamilton, A.J.; Hattersley, A.T.; Ellard, S.; Bingham, C. HNF1B-associated renal and extra-renal disease—An expanding clinical spectrum. Nat. Rev. Nephrol. 2015, 11, 102–112. [Google Scholar] [CrossRef]
- Home—OMIM. Available online: https://www.omim.org/ (accessed on 19 April 2024).
- Grand, K.; Stoltz, M.; Rizzo, L.; Röck, R.; Kaminski, M.M.; Salinas, G.; Getwan, M.; Naert, T.; Pichler, R.; Lienkamp, S.S. HNF1B Alters an Evolutionarily Conserved Nephrogenic Program of Target Genes. J. Am. Soc. Nephrol. 2023, 34, 412–432. [Google Scholar] [CrossRef]
- Karczewski, K.J.; Francioli, L.C.; Tiao, G.; Cummings, B.B.; Alföldi, J.; Wang, Q.; Collins, R.L.; Laricchia, K.M.; Ganna, A.; Birnbaum, D.P.; et al. The mutational constraint spectrum quantified from variation in 141,456 humans. Nature 2020, 581, 434–443. [Google Scholar] [CrossRef] [PubMed]
- Verscaj, C.P.; Velez-Bartolomei, F.; Bodle, E.; Chan, K.; Lyons, M.J.; Thorson, W.; Tan, W.; Rodig, N.; Graham, J.M.; Peron, A.; et al. Characterization of the prenatal renal phenotype associated with 17q12, HNF1B, microdeletions. Prenat. Diagn. 2024, 44, 237–246. [Google Scholar] [CrossRef] [PubMed]
- Cleper, R.; Reches, A.; Shapira, D.; Simchoni, S.; Reisman, L.; Ben-Sira, L.; Yaron, Y.; Wolman, I.; Malinger, G.; Brabbing-Goldstein, D.; et al. Improving renal phenotype and evolving extra-renal features of 17q12 deletion encompassing the HNF1B gene. Transl. Pediatr. 2021, 10, 3130–3139. [Google Scholar] [CrossRef] [PubMed]
- Eckardt, K.U.; Alper, S.L.; Antignac, C.; Bleyer, A.J.; Chauveau, D.; Dahan, K.; Deltas, C.; Hosking, A.; Kmoch, S.; Rampoldi, L.; et al. Autosomal dominant tubulointerstitial kidney disease: Diagnosis, classification, and management—A KDIGO consensus report. Kidney Int. 2015, 88, 676–683. [Google Scholar] [CrossRef]
- Adalat, S.; Hayes, W.N.; Bryant, W.A.; Booth, J.; Woolf, A.S.; Kleta, R.; Subtil, S.; Clissold, R.; Colclough, K.; Ellard, S.; et al. HNF1B Mutations Are Associated with a Gitelman-like Tubulopathy That Develops During Childhood. Kidney Int. Rep. 2019, 4, 1304–1311. [Google Scholar] [CrossRef]
- Moreno-De-Luca, D.; Mulle, J.G.; Kaminsky, E.B.; Sanders, S.J.; Myers, S.M.; Adam, M.P.; Pakula, A.T.; Eisenhauer, N.J.; Uhas, K.; Weik, L.; et al. Deletion 17q12 Is a Recurrent Copy Number Variant that Confers High Risk of Autism and Schizophrenia. Am. J. Hum. Genet. 2010, 87, 618–630. [Google Scholar] [CrossRef]
- Lim, S.H.; Kim, J.H.; Han, K.H.; Ahn, Y.H.; Kang, H.G.; Ha, I.-S.; Cheong, H., II. Genotype and Phenotype Analyses in Pediatric Patients with HNF1B Mutations. J. Clin. Med. 2020, 9, 2320. [Google Scholar] [CrossRef]
- Kato, T.; Tanaka, D.; Muro, S.; Jambaljav, B.; Mori, E.; Yonemitsu, S.; Oki, S.; Inagaki, N. A Novel p.L145Q Mutation in the HNF1B Gene in a Case of Maturity-onset Diabetes of the Young Type 5 (MODY5). Intern. Med. 2018, 57, 2035–2039. [Google Scholar] [CrossRef] [PubMed]
- Lu, P.; Rha, G.B.; Chi, Y.-I. Structural Basis of Disease-Causing Mutations in Hepatocyte Nuclear Factor 1β. Biochemistry 2007, 46, 12071–12080. [Google Scholar] [CrossRef]
- Tholen, L.E.; Latta, F.; Martens, J.H.A.; Hoenderop, J.G.J.; de Baaij, J.H.F. Transcription factor HNF1β controls a transcriptional network regulating kidney cell structure and tight junction integrity. Am. J. Physiol. Ren. Physiol. 2023, 324, F211–F224. [Google Scholar] [CrossRef]
- Wiedmann, M.M.; Aibara, S.; Spring, D.R.; Stewart, M.; Brenton, J.D. Structural and calorimetric studies demonstrate that the hepatocyte nuclear factor 1β (HNF1β) transcription factor is imported into the nucleus via a monopartite NLS sequence. J. Struct. Biol. 2016, 195, 273–281. [Google Scholar] [CrossRef] [PubMed]
- Bouvet, D.; Blondel, A.; de Sainte Agathe, J.-M.; Leroy, G.; Saint-Martin, C.; Bellanné-Chantelot, C. Evaluation in Monogenic Diabetes of the Impact of GCK, HNF1A, and HNF4A Variants on Splicing through the Combined Use of In Silico Tools and Minigene Assays. Hum. Mutat. 2023, 2023, 6661013. [Google Scholar] [CrossRef]
- Yamagata, K.; Oda, N.; Kaisaki, P.J.; Menzel, S.; Furuta, H.; Vaxillaire, M.; Southam, L.; Cox, R.D.; Lathrop, G.M.; Boriraj, V.V.; et al. Mutations in the hepatocyte nuclear factor-1α gene in maturity-onset diabetes of the young (MODY3). Nature 1996, 384, 455–458. [Google Scholar] [CrossRef] [PubMed]
- Locke, J.M.; Ellard, S.; Norwood, V.F.; Harries, L.W. Variants in the isoform-specific coding regions of the HNF1A, HNF4A and HNF1B genes are not a common cause of familial, young-onset diabetes or renal cysts and diabetes (RCAD). Diabet. Med. 2009, 26, 569–570. [Google Scholar] [CrossRef] [PubMed]
- Garcia-Gonzalez, M.A.; Carette, C.; Bagattin, A.; Chiral, M.; Makinistoglu, M.P.; Garbay, S.; Prévost, G.; Madaras, C.; Hérault, Y.; Leibovici, M.; et al. A suppressor locus for MODY3-diabetes. Sci. Rep. 2016, 6, 33087. [Google Scholar] [CrossRef] [PubMed]
- Ferrè, S.; de Baaij, J.H.F.; Ferreira, P.; Germann, R.; de Klerk, J.B.C.; Lavrijsen, M.; van Zeeland, F.; Venselaar, H.; Kluijtmans, L.A.J.; Hoenderop, J.G.J.; et al. Mutations in PCBD1 Cause Hypomagnesemia and Renal Magnesium Wasting. J. Am. Soc. Nephrol. 2014, 25, 574–586. [Google Scholar] [CrossRef]
- Choi, Y.-H.; McNally, B.T.; Igarashi, P. Zyxin regulates migration of renal epithelial cells through activation of hepatocyte nuclear factor-1β. Am. J. Physiol. Ren. Physiol. 2013, 305, F100–F110. [Google Scholar] [CrossRef]
- Soutoglou, E. Transcription factor-dependent regulation of CBP and P/CAF histone acetyltransferase activity. EMBO J. 2001, 20, 1984–1992. [Google Scholar] [CrossRef]
- Simaite, D.; Kofent, J.; Gong, M.; Rüschendorf, F.; Jia, S.; Arn, P.; Bentler, K.; Ellaway, C.; Kühnen, P.; Hoffmann, G.F.; et al. Recessive Mutations in PCBD1 Cause a New Type of Early-Onset Diabetes. Diabetes 2014, 63, 3557–3564. [Google Scholar] [CrossRef]
- Tholen, L.E.; Bos, C.; Jansen, P.W.T.C.; Venselaar, H.; Vermeulen, M.; Hoenderop, J.G.J.; de Baaij, J.H.F. Bifunctional protein PCBD2 operates as a co-factor for hepatocyte nuclear factor 1β and modulates gene transcription. FASEB J. 2021, 35, e21366. [Google Scholar] [CrossRef]
- Piedrafita, A.; Balayssac, S.; Casemayou, A.; Saulnier-Blache, J.; Lucas, A.; Iacovoni, J.S.; Breuil, B.; Chauveau, D.; Decramer, S.; Malet-Martino, M.; et al. Hepatocyte nuclear factor-1β shapes the energetic homeostasis of kidney tubule cells. FASEB J. 2021, 35, e21931. [Google Scholar] [CrossRef] [PubMed]
- D’Angelo, A.; Bluteau, O.; Garcia-Gonzalez, M.A.; Gresh, L.; Doyen, A.; Garbay, S.; Robine, S.; Pontoglio, M. Hepatocyte nuclear factor 1α and β control terminal differentiation and cell fate commitment in the gut epithelium. Development 2010, 137, 1573–1582. [Google Scholar] [CrossRef]
- O’Brien, L.L.; McMahon, A.P. Induction and patterning of the metanephric nephron. Semin. Cell Dev. Biol. 2014, 36, 31–38. [Google Scholar] [CrossRef] [PubMed]
- Heliot, C.; Desgrange, A.; Buisson, I.; Prunskaite-Hyyryläinen, R.; Shan, J.; Vainio, S.; Umbhauer, M.; Cereghini, S. HNF1B controls proximal-intermediate nephron segment identity in vertebrates by regulating Notch signalling components and Irx1/2. Development 2013, 140, 873–885. [Google Scholar] [CrossRef] [PubMed]
- Casemayou, A.; Fournel, A.; Bagattin, A.; Schanstra, J.; Belliere, J.; Decramer, S.; Marsal, D.; Gillet, M.; Chassaing, N.; Huart, A.; et al. Hepatocyte Nuclear Factor-1β Controls Mitochondrial Respiration in Renal Tubular Cells. J. Am. Soc. Nephrol. 2017, 28, 3205–3217. [Google Scholar] [CrossRef] [PubMed]
- Costantini, F. Genetic controls and cellular behaviors in branching morphogenesis of the renal collecting system. WIREs Dev. Biol. 2012, 1, 693–713. [Google Scholar] [CrossRef]
- Carroll, T.J.; Park, J.-S.; Hayashi, S.; Majumdar, A.; McMahon, A.P. Wnt9b Plays a Central Role in the Regulation of Mesenchymal to Epithelial Transitions Underlying Organogenesis of the Mammalian Urogenital System. Dev. Cell 2005, 9, 283–292. [Google Scholar] [CrossRef]
- Home—Reactome Pathway Database. Available online: https://reactome.org/ (accessed on 25 April 2024).
- Desgrange, A.; Cereghini, S. Nephron Patterning: Lessons from Xenopus, Zebrafish, and Mouse Studies. Cells 2015, 4, 483–499. [Google Scholar] [CrossRef]
- Park, J.-S.; Valerius, M.T.; McMahon, A.P. Wnt/β-catenin signaling regulates nephron induction during mouse kidney development. Development 2007, 134, 2533–2539. [Google Scholar] [CrossRef]
- Ng-Blichfeldt, J.-P.; Stewart, B.J.; Clatworthy, M.R.; Williams, J.M.; Röper, K. Identification of a core transcriptional program driving the human renal mesenchymal-to-epithelial transition. Dev. Cell 2024, 59, 595–612.e8. [Google Scholar] [CrossRef]
- Brown, A.C.; Muthukrishnan, S.D.; Guay, J.A.; Adams, D.C.; Schafer, D.A.; Fetting, J.L.; Oxburgh, L. Role for compartmentalization in nephron progenitor differentiation. Proc. Natl. Acad. Sci. USA 2013, 110, 4640–4645. [Google Scholar] [CrossRef] [PubMed]
- Kobayashi, A.; Kwan, K.-M.; Carroll, T.J.; McMahon, A.P.; Mendelsohn, C.L.; Behringer, R.R. Distinct and sequential tissue-specific activities of the LIM-class homeobox gene Lim1 for tubular morphogenesis during kidney development. Development 2005, 132, 2809–2823. [Google Scholar] [CrossRef]
- Georgas, K.; Rumballe, B.; Valerius, M.T.; Chiu, H.S.; Thiagarajan, R.D.; Lesieur, E.; Aronow, B.J.; Brunskill, E.W.; Combes, A.N.; Tang, D.; et al. Analysis of early nephron patterning reveals a role for distal RV proliferation in fusion to the ureteric tip via a cap mesenchyme-derived connecting segment. Dev. Biol. 2009, 332, 273–286. [Google Scholar] [CrossRef]
- Naylor, R.W.; Davidson, A.J. Hnf1beta and nephron segmentation. Pediatr. Nephrol. 2014, 29, 659–664. [Google Scholar] [CrossRef] [PubMed]
- Desgrange, A.; Heliot, C.; Skovorodkin, I.; Akram, S.U.; Heikkilä, J.; Ronkainen, V.-P.; Miinalainen, I.; Vainio, S.J.; Cereghini, S. HNF1B controls epithelial organization and cell polarity during ureteric bud branching and collecting duct morphogenesis. Development 2017, 144, 4704–4719. [Google Scholar] [CrossRef]
- Snoek, R.; Nguyen, T.Q.; Van Der Zwaag, B.; Van Zuilen, A.D.; Kruis, H.M.E.; Van Gils-Verrij, L.A.; Goldschmeding, R.; Knoers, N.V.A.M.; Rookmaaker, M.B.; Van Eerde, A.M. Importance of Genetic Diagnostics in Adult-Onset Focal Segmental Glomerulosclerosis. Nephron 2019, 142, 351–358. [Google Scholar] [CrossRef] [PubMed]
- Paces-Fessy, M.; Fabre, M.; Lesaulnier, C.; Cereghini, S. Hnf1b and Pax2 cooperate to control different pathways in kidney and ureter morphogenesis. Hum. Mol. Genet. 2012, 21, 3143–3155. [Google Scholar] [CrossRef]
- Menezes, L.F.; Zhou, F.; Patterson, A.D.; Piontek, K.B.; Krausz, K.W.; Gonzalez, F.J.; Germino, G.G. Network Analysis of a Pkd1-Mouse Model of Autosomal Dominant Polycystic Kidney Disease Identifies HNF4α as a Disease Modifier. PLoS Genet. 2012, 8, e1003053. [Google Scholar] [CrossRef]
- Yamagata, K.; Furuta, H.; Oda, N.; Kaisaki, P.J.; Menzel, S.; Cox, N.J.; Fajans, S.S.; Signorini, S.; Stoffel, M.; Bell, G.I. Mutations in the hepatocyte nuclear factor-4α gene in maturity-onset diabetes of the young (MODY1). Nature 1996, 384, 458–460. [Google Scholar] [CrossRef]
- Mae, S.-I.; Ryosaka, M.; Sakamoto, S.; Matsuse, K.; Nozaki, A.; Igami, M.; Kabai, R.; Watanabe, A.; Osafune, K. Expansion of Human iPSC-Derived Ureteric Bud Organoids with Repeated Branching Potential. Cell Rep. 2020, 32, 107963. [Google Scholar] [CrossRef]
- Martin, E.; Girardello, R.; Dittmar, G.; Ludwig, A. New insights into the organization and regulation of the apical polarity network in mammalian epithelial cells. FEBS J. 2021, 288, 7073–7095. [Google Scholar] [CrossRef] [PubMed]
- Tan, B.; Yatim, S.M.J.M.; Peng, S.; Gunaratne, J.; Hunziker, W.; Ludwig, A. The Mammalian Crumbs Complex Defines a Distinct Polarity Domain Apical of Epithelial Tight Junctions. Curr. Biol. 2020, 30, 2791–2804.e6. [Google Scholar] [CrossRef] [PubMed]
- Pieczynski, J.; Margolis, B. Protein complexes that control renal epithelial polarity. Am. J. Physiol. Ren. Physiol. 2011, 300, F589–F601. [Google Scholar] [CrossRef] [PubMed]
- De Vas, M.G.; Kopp, J.L.; Heliot, C.; Sander, M.; Cereghini, S.; Haumaitre, C. Hnf1b controls pancreas morphogenesis and the generation of Ngn3+ endocrine progenitors. Development 2015, 142, 871–882. [Google Scholar] [CrossRef]
- Veikkolainen, V.; Naillat, F.; Railo, A.; Chi, L.; Manninen, A.; Hohenstein, P.; Hastie, N.; Vainio, S.; Elenius, K. ErbB4 Modulates Tubular Cell Polarity and Lumen Diameter during Kidney Development. J. Am. Soc. Nephrol. 2012, 23, 112–122. [Google Scholar] [CrossRef]
- Wilson, P.D. Apico-basal polarity in polycystic kidney disease epithelia. Biochim. Et Biophys. Acta (BBA) Mol. Basis Dis. 2011, 1812, 1239–1248. [Google Scholar] [CrossRef] [PubMed]
- Shao, A.; Chan, S.C.; Igarashi, P. Role of transcription factor hepatocyte nuclear factor-1β in polycystic kidney disease. Cell Signal 2020, 71, 109568. [Google Scholar] [CrossRef]
- Chan, S.C.; Zhang, Y.; Pontoglio, M.; Igarashi, P. Hepatocyte nuclear factor-1β regulates Wnt signaling through genome-wide competition with β-catenin/lymphoid enhancer binding factor. Proc. Natl. Acad. Sci. USA 2019, 116, 24133–24142. [Google Scholar] [CrossRef]
- Choi, Y.-H.; Suzuki, A.; Hajarnis, S.; Ma, Z.; Chapin, H.C.; Caplan, M.J.; Pontoglio, M.; Somlo, S.; Igarashi, P. Polycystin-2 and phosphodiesterase 4C are components of a ciliary A-kinase anchoring protein complex that is disrupted in cystic kidney diseases. Proc. Natl. Acad. Sci. USA 2011, 108, 10679–10684. [Google Scholar] [CrossRef]
- Song, X.; Di Giovanni, V.; He, N.; Wang, K.; Ingram, A.; Rosenblum, N.D.; Pei, Y. Systems biology of autosomal dominant polycystic kidney disease (ADPKD): Computational identification of gene expression pathways and integrated regulatory networks. Hum. Mol. Genet. 2009, 18, 2328–2343. [Google Scholar] [CrossRef]
- Verdeguer, F.; Le Corre, S.; Fischer, E.; Callens, C.; Garbay, S.; Doyen, A.; Igarashi, P.; Terzi, F.; Pontoglio, M. A mitotic transcriptional switch in polycystic kidney disease. Nat. Med. 2010, 16, 106–110. [Google Scholar] [CrossRef]
- Kang, H.S.; Beak, J.Y.; Kim, Y.-S.; Herbert, R.; Jetten, A.M. Glis3 Is Associated with Primary Cilia and Wwtr1/TAZ and Implicated in Polycystic Kidney Disease. Mol. Cell Biol. 2009, 29, 2556–2569. [Google Scholar] [CrossRef] [PubMed][Green Version]
- Kang, H.S.; Kim, Y.-S.; ZeRuth, G.; Beak, J.Y.; Gerrish, K.; Kilic, G.; Sosa-Pineda, B.; Jensen, J.; Foley, J.; Jetten, A.M. Transcription Factor Glis3, a Novel Critical Player in the Regulation of Pancreatic β-Cell Development and Insulin Gene Expression. Mol. Cell Biol. 2009, 29, 6366–6379. [Google Scholar] [CrossRef] [PubMed]
- Schlingmann, K.P.; de Baaij, J.H.F. The genetic spectrum of Gitelman(-like) syndromes. Curr. Opin. Nephrol. Hypertens. 2022, 31, 508–515. [Google Scholar] [CrossRef]
- Saji, T.; Kikuchi, R.; Kusuhara, H.; Kim, I.; Gonzalez, F.J.; Sugiyama, Y. Transcriptional Regulation of Human and Mouse Organic Anion Transporter 1 by Hepatocyte Nuclear Factor 1 α/β. J. Pharmacol. Exp. Ther. 2008, 324, 784–790. [Google Scholar] [CrossRef] [PubMed]
- Jin, L.; Kikuchi, R.; Saji, T.; Kusuhara, H.; Sugiyama, Y. Regulation of Tissue-Specific Expression of Renal Organic Anion Transporters by Hepatocyte Nuclear Factor 1 α/β and DNA Methylation. J. Pharmacol. Exp. Ther. 2012, 340, 648–655. [Google Scholar] [CrossRef]
- Kikuchi, R.; Kusuhara, H.; Hattori, N.; Shiota, K.; Kim, I.; Gonzalez, F.J.; Sugiyama, Y. Regulation of the Expression of Human Organic Anion Transporter 3 by Hepatocyte Nuclear Factor 1α/β and DNA Methylation. Mol. Pharmacol. 2006, 70, 887–896. [Google Scholar] [CrossRef] [PubMed]
- Cheret, C.; Doyen, A.; Yaniv, M.; Pontoglio, M. Hepatocyte Nuclear Factor 1 α Controls Renal Expression of the Npt1-Npt4 Anionic Transporter Locus. J. Mol. Biol. 2002, 322, 929–941. [Google Scholar] [CrossRef]
- Kikuchi, R.; Kusuhara, H.; Hattori, N.; Kim, I.; Shiota, K.; Gonzalez, F.J.; Sugiyama, Y. Regulation of Tissue-Specific Expression of the Human and Mouse Urate Transporter 1 Gene by Hepatocyte Nuclear Factor 1 α/β and DNA Methylation. Mol. Pharmacol. 2007, 72, 1619–1625. [Google Scholar] [CrossRef]
- Fukui, K.; Yang, Q.; Cao, Y.; Takahashi, N.; Hatakeyama, H.; Wang, H.; Wada, J.; Zhang, Y.; Marselli, L.; Nammo, T.; et al. The HNF-1 target Collectrin controls insulin exocytosis by SNARE complex formation. Cell Metab. 2005, 2, 373–384. [Google Scholar] [CrossRef]
- Terryn, S.; Tanaka, K.; Lengelé, J.-P.; Olinger, E.; Dubois-Laforgue, D.; Garbay, S.; Kozyraki, R.; Van Der Smissen, P.; Christensen, E.I.; Courtoy, P.J.; et al. Tubular proteinuria in patients with HNF1α mutations: HNF1α drives endocytosis in the proximal tubule. Kidney Int. 2016, 89, 1075–1089. [Google Scholar] [CrossRef] [PubMed]
- Tokonami, N.; Takata, T.; Beyeler, J.; Ehrbar, I.; Yoshifuji, A.; Christensen, E.I.; Loffing, J.; Devuyst, O.; Olinger, E.G. Uromodulin is expressed in the distal convoluted tubule where it is critical for regulation of the sodium chloride cotransporter, N.C.C. Kidney Int. 2018, 94, 701–715. [Google Scholar] [CrossRef]
- Nie, M.; Bal, M.S.; Liu, J.; Yang, Z.; Rivera, C.; Wu, X.-R.; Hoenderop, J.G.J.; Bindels, R.J.M.; Marciano, D.K.; Wolf, M.T.F. Uromodulin regulates renal magnesium homeostasis through the ion channel transient receptor potential melastatin 6 (TRPM6). J. Biol. Chem. 2018, 293, 16488–16502. [Google Scholar] [CrossRef] [PubMed]
- Wolf, M.T.F.; Wu, X.-R.; Huang, C.-L. Uromodulin upregulates TRPV5 by impairing caveolin-mediated endocytosis. Kidney Int. 2013, 84, 130–137. [Google Scholar] [CrossRef]
- Trudu, M.; Janas, S.; Lanzani, C.; Debaix, H.; Schaeffer, C.; Ikehata, M.; Citterio, L.; Demaretz, S.; Trevisani, F.; Ristagno, G.; et al. Common noncoding UMOD gene variants induce salt-sensitive hypertension and kidney damage by increasing uromodulin expression. Nat. Med. 2013, 19, 1655–1660. [Google Scholar] [CrossRef]
- Tokonami, N.; Olinger, E.; Debaix, H.; Houillier, P.; Devuyst, O. The excretion of uromodulin is modulated by the calcium-sensing receptor. Kidney Int. 2018, 94, 882–886. [Google Scholar] [CrossRef]
- Kompatscher, A.; de Baaij, J.H.F.; Aboudehen, K.; Farahani, S.; van Son, L.H.J.; Milatz, S.; Himmerkus, N.; Veenstra, G.C.; Bindels, R.J.M.; Hoenderop, J.G.J. Transcription factor HNF1β regulates expression of the calcium-sensing receptor in the thick ascending limb of the kidney. Am. J. Physiol. Ren. Physiol. 2018, 315, F27–F35. [Google Scholar] [CrossRef] [PubMed]
- Ferrè, S.; Bongers, E.M.H.F.; Sonneveld, R.; Cornelissen, E.A.M.; van der Vlag, J.; van Boekel, G.A.J.; Wetzels, J.F.M.; Hoenderop, J.G.J.; Bindels, R.J.M.; Nijenhuis, T. Early Development of Hyperparathyroidism Due to Loss of PTH Transcriptional Repression in Patients With HNF1β Mutations? J. Clin. Endocrinol. Metab. 2013, 98, 4089–4096. [Google Scholar] [CrossRef]
- Ferrè, S.; Veenstra, G.J.C.; Bouwmeester, R.; Hoenderop, J.G.J.; Bindels, R.J.M. HNF-1B specifically regulates the transcription of the γa-subunit of the Na+/K+-ATPase. Biochem. Biophys. Res. Commun. 2011, 404, 284–290. [Google Scholar] [CrossRef]
- de Baaij, J.H.F.; Dorresteijn, E.M.; Hennekam, E.A.M.; Kamsteeg, E.-J.; Meijer, R.; Dahan, K.; Muller, M.; van den Dorpel, M.A.; Bindels, R.J.M.; Hoenderop, J.G.J.; et al. Recurrent FXYD2 p.Gly41Arg mutation in patients with isolated dominant hypomagnesaemia. Nephrol. Dial. Transplant. 2015, 30, 952–957. [Google Scholar] [CrossRef]
- Kompatscher, A.; de Baaij, J.H.F.; Aboudehen, K.; Hoefnagels, A.P.W.M.; Igarashi, P.; Bindels, R.J.M.; Veenstra, G.J.C.; Hoenderop, J.G.J. Loss of transcriptional activation of the potassium channel Kir5.1 by HNF1β drives autosomal dominant tubulointerstitial kidney disease. Kidney Int. 2017, 92, 1145–1156. [Google Scholar] [CrossRef] [PubMed]
- Schlingmann, K.P.; Renigunta, A.; Hoorn, E.J.; Forst, A.-L.; Renigunta, V.; Atanasov, V.; Mahendran, S.; Barakat, T.S.; Gillion, V.; Godefroid, N.; et al. Defects in KCNJ16 Cause a Novel Tubulopathy with Hypokalemia, Salt Wasting, Disturbed Acid-Base Homeostasis, and Sensorineural Deafness. J. Am. Soc. Nephrol. 2021, 32, 1498–1512. [Google Scholar] [CrossRef] [PubMed]
- Zhang, X.; Huang, S.; Gao, M.; Liu, J.; Jia, X.; Han, Q.; Zheng, S.; Miao, Y.; Li, S.; Weng, H.; et al. Farnesoid X receptor (FXR) gene deficiency impairs urine concentration in mice. Proc. Natl. Acad. Sci. USA 2014, 111, 2277–2282. [Google Scholar] [CrossRef] [PubMed]
- Kim, E.Y.; Lee, J.M. Transcriptional Control of Trpm6 by the Nuclear Receptor, F.X.R. Int. J. Mol. Sci. 2022, 23, 1980. [Google Scholar] [CrossRef]
- Bhargava, P.; Schnellmann, R.G. Mitochondrial energetics in the kidney. Nat. Rev. Nephrol. 2017, 13, 629–646. [Google Scholar] [CrossRef]
- Thomas, J.L.; Pham, H.; Li, Y.; Hall, E.; Perkins, G.A.; Ali, S.S.; Patel, H.H.; Singh, P. Hypoxia-inducible factor-1α activation improves renal oxygenation and mitochondrial function in early chronic kidney disease. Am. J. Physiol. Ren. Physiol. 2017, 313, F282–F290. [Google Scholar] [CrossRef]
- Viering, D.; Schlingmann, K.P.; Hureaux, M.; Nijenhuis, T.; Mallett, A.; Chan, M.M.Y.; van Beek, A.; van Eerde, A.M.; Coulibaly, J.-M.; Vallet, M.; et al. Gitelman-Like Syndrome Caused by Pathogenic Variants in mtDNA. J. Am. Soc. Nephrol. 2022, 33, 305–325. [Google Scholar] [CrossRef]
- Wang, Z.; Ying, Z.; Bosy-Westphal, A.; Zhang, J.; Schautz, B.; Later, W.; Heymsfield, S.B.; Müller, M.J. Specific metabolic rates of major organs and tissues across adulthood: Evaluation by mechanistic model of resting energy expenditure. Am. J. Clin. Nutr. 2010, 92, 1369–1377. [Google Scholar] [CrossRef]
- Kang, H.M.; Ahn, S.H.; Choi, P.; Ko, Y.-A.; Han, S.H.; Chinga, F.; Park, A.S.D.; Tao, J.; Sharma, K.; Pullman, J.; et al. Defective fatty acid oxidation in renal tubular epithelial cells has a key role in kidney fibrosis development. Nat. Med. 2015, 21, 37–46. [Google Scholar] [CrossRef]
- Aboudehen, K.; Kim, M.S.; Mitsche, M.; Garland, K.; Anderson, N.; Noureddine, L.; Pontoglio, M.; Patel, V.; Xie, Y.; DeBose-Boyd, R.; et al. Transcription Factor Hepatocyte Nuclear Factor–1β Regulates Renal Cholesterol Metabolism. J. Am. Soc. Nephrol. 2016, 27, 2408–2421. [Google Scholar] [CrossRef]
- Horton, J.D.; Cohen, J.C.; Hobbs, H.H. PCSK9: A convertase that coordinates LDL catabolism. J. Lipid Res. 2009, 50, S172–S177. [Google Scholar] [CrossRef] [PubMed]




Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).