

New Small-Molecule SERCA Inhibitors Enhance Treatment Efficacy in Lenvatinib-Resistant Papillary Thyroid Cancer



Abstract
1. Introduction
2. Results
2.1. Patient-Derived PTC Cell Lines and Their Properties
2.2. Distinctions in Genetic Alterations and Activated Signaling Pathways between Patient-Derived Lenvatinib-Sensitive and -Resistant PTC Cell Lines
2.3. Identification of Therapeutic Molecules, Candidates 40 and 42, Based on SERCA Structure through In-Silico Screening for Suppression of Lenvatinib-Resistant PTC
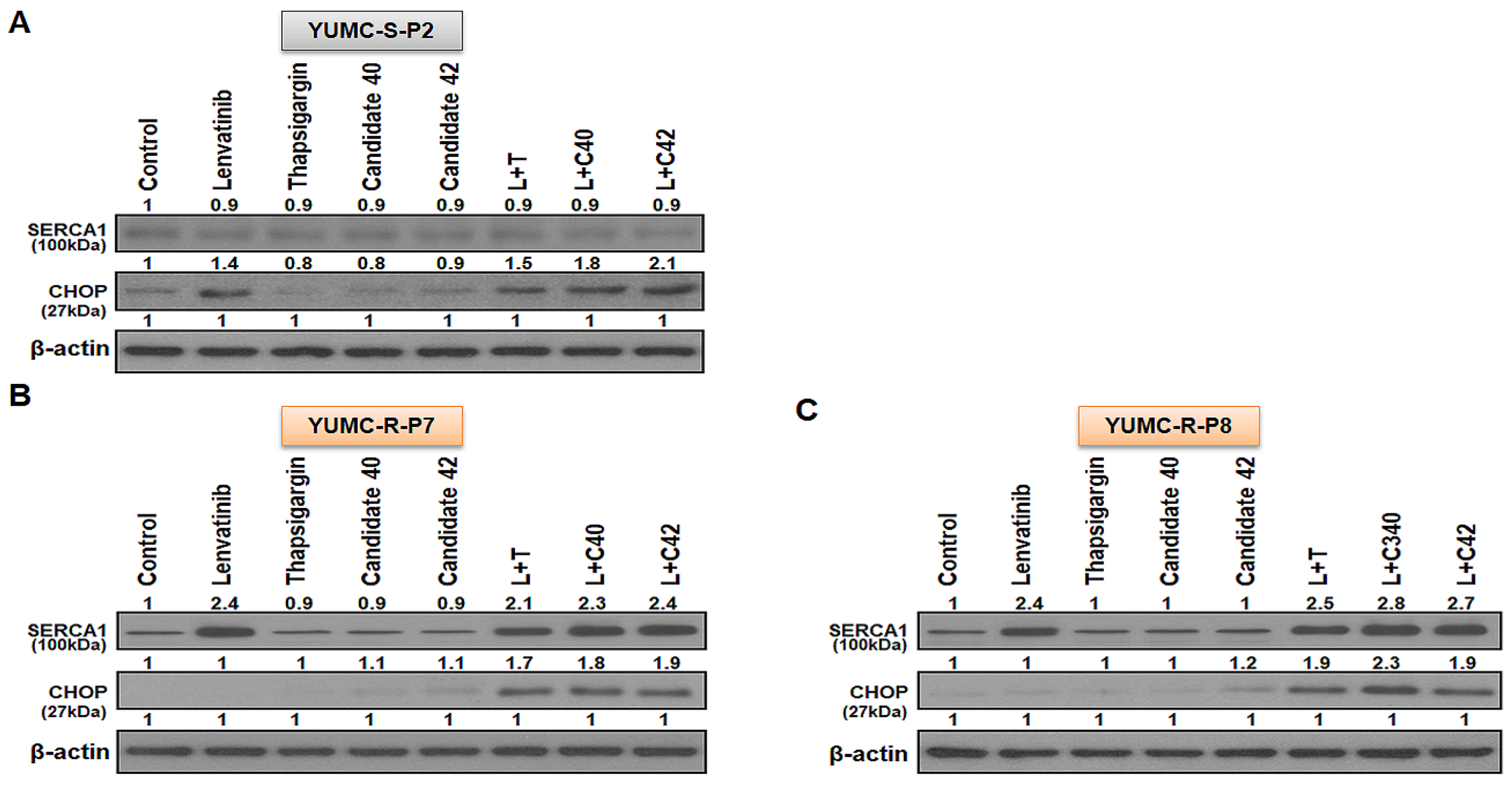
2.4. SERCA1 as a Key Player in Lenvatinib-Resistant PTC Cells for Prolonging Survival under Lenvatinib Treatment
2.5. Targeted Therapy In Vivo Treatment with Candidates 40 and 42 in a Patient-Derived Lenvatinib-Resistant PTC Cell Mouse Xenograft Model
3. Discussion
4. Materials and Methods
4.1. Study Design and Ethical Considerations
4.2. Patients
4.2.1. Patient 1
4.2.2. Patients 2 and 3
4.3. Patient Tissue Specimens
4.4. Primary Culture and Cancer Cell Isolation
4.5. mRNA-seq Data
4.6. Statistical Analysis of Gene Expression Level
4.7. Hierarchical Clustering
4.8. Total RNA Extraction and Quantitative Reverse Transcription-Polymerase Chain Reaction
4.9. Cell Viability Assay
4.10. Immunoblot Analysis
4.11. Human PTC Cell Xenograft
4.12. Statistical Analysis
4.13. Virtual Screening with Chemical Binding Similarity
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Nguyen, Q.T.; Lee, E.J.; Huang, M.G.; Park, Y.I.; Khullar, A.; Plodkowski, R.A. Diagnosis and treatment of patients with thyroid cancer. Am. Health Drug Benefits 2015, 8, 30–40. [Google Scholar] [PubMed]
- Su, Z.; Bao, W.; Yang, G.; Liu, J.; Zhao, B. SOX12 Promotes Thyroid Cancer Cell Proliferation and Invasion by Regulating the Expression of POU2F1 and POU3F1. Yonsei Med. J. 2022, 63, 591–600. [Google Scholar] [CrossRef] [PubMed]
- Owonikoko, T.K.; Chowdry, R.P.; Chen, Z.; Kim, S.; Saba, N.F.; Shin, D.M.; Khuri, F.R. Clinical Efficacy of Targeted Biologic Agents as Second-Line Therapy of Advanced Thyroid Cancer. Oncol. 2013, 18, 1262–1269. [Google Scholar] [CrossRef] [PubMed]
- Raue, F.; Frank-Raue, K. Thyroid Cancer: Risk-Stratified Management and Individualized Therapy. Clin. Cancer Res. 2016, 22, 5012–5021. [Google Scholar] [CrossRef]
- Ibrahimpasic, T.; Ghossein, R.; Shah, J.P.; Ganly, I. Poorly Differentiated Carcinoma of the Thyroid Gland: Current Status and Future Prospects. Thyroid® 2019, 29, 311–321. [Google Scholar] [CrossRef]
- de Castroneves, L.A.; Negrão, M.V.; de Freitas, R.M.C.; Papadia, C.; Lima, J.V.; Fukushima, J.T.; Simão, E.F.; Kulcsar, M.A.V.; Tavares, M.R.; Jorge, A.A.d.L.; et al. Sorafenib for the Treatment of Progressive Metastatic Medullary Thyroid Cancer: Efficacy and Safety Analysis. Thyroid® 2016, 26, 414–419. [Google Scholar] [CrossRef]
- Ito, Y.; Onoda, N.; Ito, K.-I.; Sugitani, I.; Takahashi, S.; Yamaguchi, I.; Kabu, K.; Tsukada, K. Sorafenib in Japanese Patients with Locally Advanced or Metastatic Medullary Thyroid Carcinoma and Anaplastic Thyroid Carcinoma. Thyroid® 2017, 27, 1142–1148. [Google Scholar] [CrossRef]
- Sun, Y.; Hao, G.; Zhuang, M.; Lv, H.; Liu, C.; Su, K. MEG3 LncRNA from Exosomes Released from Cancer-Associated Fibroblasts Enhances Cisplatin Chemoresistance in SCLC via a MiR-15a-5p/CCNE1 Axis. Yonsei Med. J. 2022, 63, 229–240. [Google Scholar] [CrossRef]
- D’andrea, A.D. Mechanisms of PARP inhibitor sensitivity and resistance. DNA Repair 2018, 71, 172–176. [Google Scholar] [CrossRef]
- Li, Y.-J.; Lei, Y.-H.; Yao, N.; Wang, C.-R.; Hu, N.; Ye, W.-C.; Zhang, D.-M.; Chen, Z.-S. Autophagy and multidrug resistance in cancer. Chin. J. Cancer 2017, 36, 52. [Google Scholar] [CrossRef]
- Ozaki, T.; Yu, M.; Yin, D.; Sun, D.; Zhu, Y.; Bu, Y.; Sang, M. Impact of RUNX2 on drug-resistant human pancreatic cancer cells with p53 mutations. BMC Cancer 2018, 18, 309. [Google Scholar] [CrossRef] [PubMed]
- He, Y. Systematic response of staurosporine scaffold-based inhibitors to drug-resistant cancer kinase mutations. Arch. Der Pharm. 2020, 353, e1900320. [Google Scholar] [CrossRef] [PubMed]
- Coca-Pelaz, A.; Shah, J.P.; Hernandez-Prera, J.C.; Ghossein, R.A.; Rodrigo, J.P.; Hartl, D.M.; Olsen, K.D.; Shaha, A.R.; Zafereo, M.; Suarez, C.; et al. Papillary Thyroid Cancer—Aggressive Variants and Impact on Management: A Narrative Review. Adv. Ther. 2020, 37, 3112–3128. [Google Scholar] [CrossRef] [PubMed]
- Lam, A.K.-Y.; Lo, C.-Y.; Lam, K.S.-L. Papillary Carcinoma of Thyroid: A 30-yr Clinicopathological Review of the Histological Variants. Endocr. Pathol. 2005, 16, 323–330. [Google Scholar] [CrossRef]
- Liu, J.; Liu, Y.; Lin, Y.; Liang, J. Radioactive Iodine-Refractory Differentiated Thyroid Cancer and Redifferentiation Therapy. Endocrinol. Metab. 2019, 34, 215–225. [Google Scholar] [CrossRef]
- Bo, W.; Chen, Y. Lenvatinib resistance mechanism and potential ways to conquer. Front. Pharmacol. 2023, 14, 1153991. [Google Scholar] [CrossRef]
- Brini, M.; Carafoli, E. Calcium Pumps in Health and Disease. Physiol. Rev. 2009, 89, 1341–1378. [Google Scholar] [CrossRef]
- Cui, C.; Merritt, R.; Fu, L.; Pan, Z. Targeting calcium signaling in cancer therapy. Acta Pharm. Sin. B 2017, 7, 3–17. [Google Scholar] [CrossRef]
- Denmeade, S.R.; Isaacs, J.T. The SERCA pump as a therapeutic target: Making a “smart bomb” for prostate cancer. Cancer Biol. Ther. 2005, 4, 21–29. [Google Scholar] [CrossRef]
- Chemaly, E.R.; Troncone, L.; Lebeche, D. SERCA control of cell death and survival. Cell Calcium 2018, 69, 46–61. [Google Scholar] [CrossRef]
- Kim, Y.; Yun, H.J.; Choi, K.H.; Kim, C.W.; Lee, J.H.; Weicker, R.; Kim, S.-M.; Park, K.C. Discovery of New Anti-Cancer Agents against Patient-Derived Sorafenib-Resistant Papillary Thyroid Cancer. Int. J. Mol. Sci. 2023, 24, 16413. [Google Scholar] [CrossRef] [PubMed]
- Chang, H.-S.; Kim, Y.; Lee, S.Y.; Yun, H.J.; Chang, H.-J.; Park, K.C. Anti-Cancer SERCA Inhibitors Targeting Sorafenib-Resistant Human Papillary Thyroid Carcinoma. Int. J. Mol. Sci. 2023, 24, 7069. [Google Scholar] [CrossRef] [PubMed]
- Park, K.C.; Kim, J.M.; Kim, S.Y.; Kim, S.-M.; Lim, J.H.; Kim, M.K.; Fang, S.; Kim, Y.; Mills, G.B.; Noh, S.H.; et al. PMCA inhibition reverses drug resistance in clinically refractory cancer patient-derived models. BMC Med. 2023, 21, 38. [Google Scholar] [CrossRef] [PubMed]
- Park, K.C.; Kim, S.W.; Jeon, J.Y.; Jo, A.R.; Choi, H.J.; Kim, J.; Lee, H.G.; Kim, Y.; Mills, G.B.; Noh, S.H.; et al. Survival of Cancer Stem-Like Cells Under Metabolic Stress via CaMK2α-mediated Upregulation of Sarco/Endoplasmic Reticulum Calcium ATPase Expression. Clin. Cancer Res. 2018, 24, 1677–1690. [Google Scholar] [CrossRef]
- Yun, H.-J.; Lim, J.-H.; Kim, S.-Y.; Kim, S.-M.; Park, K.-C. Discovery of Pharmaceutical Composition for Prevention and Treatment in Patient-Derived Metastatic Medullary Thyroid Carcinoma Model. Biomedicines 2022, 10, 1901. [Google Scholar] [CrossRef]
- Phi, L.T.H.; Sari, I.N.; Yang, Y.-G.; Lee, S.-H.; Jun, N.; Kim, K.S.; Lee, Y.K.; Kwon, H.Y. Cancer Stem Cells (CSCs) in Drug Resistance and their Therapeutic Implications in Cancer Treatment. Stem Cells Int. 2018, 2018, 5416923. [Google Scholar] [CrossRef]
- Prieto-Vila, M.; Takahashi, R.-U.; Usuba, W.; Kohama, I.; Ochiya, T. Drug Resistance Driven by Cancer Stem Cells and Their Niche. Int. J. Mol. Sci. 2017, 18, 2574. [Google Scholar] [CrossRef]
- Yang, L.; Shi, P.; Zhao, G.; Xu, J.; Peng, W.; Zhang, J.; Zhang, G.; Wang, X.; Dong, Z.; Chen, F.; et al. Targeting Cancer Stem Cell Pathways for Cancer Therapy. Signal Transduct. Target. Ther. 2020, 5, 8. [Google Scholar] [CrossRef]
- Kim, S.-M.; Park, K.; Lim, J.H.; Yun, H.J.; Kim, S.Y.; Choi, K.H.; Kim, C.W.; Lee, J.H.; Weicker, R.; Pan, C.-H.; et al. Potential Therapeutic Agents against Paclitaxel—And Sorafenib-Resistant Papillary Thyroid Carcinoma. Int. J. Mol. Sci. 2022, 23, 10378. [Google Scholar] [CrossRef]
- Lim, J.H.; Choi, K.H.; Kim, S.Y.; Park, C.S.; Kim, S.-M.; Park, K.C. Patient-Derived, Drug-Resistant Colon Cancer Cells Evade Chemotherapeutic Drug Effects via the Induction of Epithelial-Mesenchymal Transition-Mediated Angiogenesis. Int. J. Mol. Sci. 2020, 21, 7469. [Google Scholar] [CrossRef]
- A Pimentel, A.; Benaim, G. Ca2+ and sphingolipids as modulators for apoptosis and cancer. Invest. Clin. 2012, 53, 84–110. [Google Scholar] [PubMed]
- Seo, J.-A.; Kim, B.; Dhanasekaran, D.N.; Tsang, B.K.; Song, Y.S. Curcumin induces apoptosis by inhibiting sarco/endoplasmic reticulum Ca2+ ATPase activity in ovarian cancer cells. Cancer Lett. 2015, 371, 30–37. [Google Scholar] [CrossRef] [PubMed]
- Lim, J.H.; Park, K.; Choi, K.H.; Kim, C.W.; Lee, J.H.; Weicker, R.; Pan, C.-H.; Kim, S.-M.; Park, K.C. Drug Discovery Using Evolutionary Similarities in Chemical Binding to Inhibit Patient-Derived Hepatocellular Carcinoma. Int. J. Mol. Sci. 2022, 23, 7971. [Google Scholar] [CrossRef] [PubMed]
- Limonta, P.; Moretti, R.M.; Marzagalli, M.; Fontana, F.; Raimondi, M.; Marelli, M.M. Role of Endoplasmic Reticulum Stress in the Anticancer Activity of Natural Compounds. Int. J. Mol. Sci. 2019, 20, 961. [Google Scholar] [CrossRef]
- Bergdorf, K.; Ferguson, D.C.; Mehrad, M.; Ely, K.; Stricker, T.; Weiss, V.L. Papillary thyroid carcinoma behavior: Clues in the tumor microenvironment. Endocr.-Relat. Cancer 2019, 26, 601–614. [Google Scholar] [CrossRef]
- Lewiński, A.; Adamczewski, Z. Papillary thyroid carcinoma: A cancer with an extremely diverse genetic background and prognosis. Pol. Arch. Intern. Med. 2017, 127, 388–389. [Google Scholar] [CrossRef]
- Giuffrida, R.; Adamo, L.; Iannolo, G.; Vicari, L.; Giuffrida, D.; Eramo, A.; Gulisano, M.; Memeo, L.; Conticello, C. Resistance of papillary thyroid cancer stem cells to chemotherapy. Oncol. Lett. 2016, 12, 687–691. [Google Scholar] [CrossRef]
- de Groot, J.W.B.; Links, T.P.; Plukker, J.T.M.; Lips, C.J.M.; Hofstra, R.M.W. RET as a Diagnostic and Therapeutic Target in Sporadic and Hereditary Endocrine Tumors. Endocr. Rev. 2006, 27, 535–560. [Google Scholar] [CrossRef]
- Russo, D.; Damante, G.; Puxeddu, E.; Durante, C.; Filetti, S. Epigenetics of thyroid cancer and novel therapeutic targets. J. Mol. Endocrinol. 2011, 46, R73–R81. [Google Scholar] [CrossRef]
- Kerr, A.J.; Dodwell, D.; McGale, P.; Holt, F.; Duane, F.; Mannu, G.; Darby, S.C.; Taylor, C.W. Adjuvant and neoadjuvant breast cancer treatments: A systematic review of their effects on mortality. Cancer Treat. Rev. 2022, 105, 102375. [Google Scholar] [CrossRef]
- Ettrich, T.J.; Sturm, N.; Güthle, M.; Hüttner, F.J.; Perkhofer, L. Pancreatic Cancer: Current Multimodality Treatment Options and the Future Impact of Molecular Biological Profiling. Visc. Med. 2022, 38, 20–29. [Google Scholar] [CrossRef] [PubMed]
- Akateh, C.; Black, S.M.; Conteh, L.; Miller, E.D.; Noonan, A.; Elliott, E.; Pawlik, T.M.; Tsung, A.; Cloyd, J.M. Neoadjuvant and adjuvant treatment strategies for hepatocellular carcinoma. World J. Gastroenterol. 2019, 25, 3704–3721. [Google Scholar] [CrossRef] [PubMed]
- Foerster, F.; Galle, P.R. The Current Landscape of Clinical Trials for Systemic Treatment of HCC. Cancers 2021, 13, 1962. [Google Scholar] [CrossRef] [PubMed]
- O’Brien, J.; Etherington; Salmon, G.; McNally, S. The influence of nicardipine on platelet tests in patients with claudication. Thromb. Res. 1990, 58, 75–80. [Google Scholar] [CrossRef]
- Jin, M.; Wang, J.; Ji, X.; Cao, H.; Zhu, J.; Chen, Y.; Yang, J.; Zhao, Z.; Ren, T.; Xing, J. MCUR1 facilitates epithelial-mesenchymal transition and metastasis via the mitochondrial calcium dependent ROS/Nrf2/Notch pathway in hepatocellular carcinoma. J. Exp. Clin. Cancer Res. 2019, 38, 136. [Google Scholar] [CrossRef]
- Sun, J.; Ailiman, M. Regulation of calcium pump through Notch/Jagged/Hes signaling pathway in canine model of chronic atrial fibrillation. Int. J. Clin. Exp. Pathol. 2019, 12, 4034–4040. [Google Scholar]
- Pagliaro, L.; Marchesini, M.; Roti, G. Targeting oncogenic Notch signaling with SERCA inhibitors. J. Hematol. Oncol. 2021, 14, 8. [Google Scholar] [CrossRef]
- Marchesini, M.; Gherli, A.; Montanaro, A.; Patrizi, L.; Sorrentino, C.; Pagliaro, L.; Rompietti, C.; Kitara, S.; Heit, S.; Olesen, C.E.; et al. Blockade of Oncogenic NOTCH1 with the SERCA Inhibitor CAD204520 in T Cell Acute Lymphoblastic Leukemia. Cell Chem. Biol. 2020, 27, 678–697.e13. [Google Scholar] [CrossRef]
- Yun, H.J.; Kim, M.; Kim, S.Y.; Fang, S.; Kim, Y.; Chang, H.-S.; Chang, H.-J.; Park, K.C. Effects of Anti-Cancer Drug Sensitivity-Related Genetic Differences on Therapeutic Approaches in Refractory Papillary Thyroid Cancer. Int. J. Mol. Sci. 2022, 23, 699. [Google Scholar] [CrossRef]
- Kim, D.; Langmead, B.; Salzberg, S.L. HISAT: A fast spliced aligner with low memory requirements. Nat. Methods 2015, 12, 357–360. [Google Scholar] [CrossRef]
- Pertea, M.; Pertea, G.M.; Antonescu, C.M.; Chang, T.-C.; Mendell, J.T.; Salzberg, S.L. StringTie enables improved reconstruction of a transcriptome from RNA-seq reads. Nat. Biotechnol. 2015, 33, 290–295. [Google Scholar] [CrossRef] [PubMed]
- Pertea, M.; Kim, D.; Pertea, G.M.; Leek, J.T.; Salzberg, S.L. Transcript-level expression analysis of RNA-seq experiments with HISAT, StringTie and Ballgown. Nat. Protoc. 2016, 11, 1650–1667. [Google Scholar] [CrossRef] [PubMed]
- Park, K.; Ko, Y.-J.; Durai, P.; Pan, C.-H. Machine learning-based chemical binding similarity using evolutionary relationships of target genes. Nucleic Acids Res. 2019, 47, e128. [Google Scholar] [CrossRef] [PubMed]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| YUMC-S-P2 | YUMC-R-P7 | YUMC-R-P8 | |
|---|---|---|---|
| Age at Diagnosis | 53 | 57 | 52 |
| Gender | Male | Female | Male |
| Primary Disease Site | Thyroid | Thyroid | Thyroid |
| Stage | T4aN1bM0 | T4aN1bM1 | T4aN1bM1 |
| Primary Pathology | Papillary thyroid cancer | Papillary thyroid cancer (Recurrence & Metastasis after lenvatinib treatment) | Papillary thyroid cancer (Recurrence & Metastasis after lenvatinib treatment) |
| Classification of specimen used for culture | Fresh tumor | Fresh tumor | Fresh tumor |
| Obtained from | Severance Hospital, Seoul, Republic of Korea | Severance Hospital, Seoul, Republic of Korea | Severance Hospital, Seoul, Republic of Korea |
| Cell Line | Histopathology | Animal | Cell Proliferation IC50 (μM) | |||
|---|---|---|---|---|---|---|
| Lenvatinib | L + T | L + C40 | S + C42 | |||
| YUMC-S-P2 | Thyroid, Papillary | Human | 12 (±0.1) | 12 (±0.2) | 12 (±0.2) | 12 (±0.2) |
| YUMC-R-P7 | Thyroid, Papillary | Human | ─ | 12 (±0.3) | 25 (±0.2) | 23 (±0.1) |
| YUMC-R-P8 | Thyroid, Papillary | Human | ─ | 23 (±0.2) | 13 (±0.2) | 13 (±0.4) |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Kim, J.; Chang, H.-S.; Yun, H.J.; Chang, H.-J.; Park, K.C. New Small-Molecule SERCA Inhibitors Enhance Treatment Efficacy in Lenvatinib-Resistant Papillary Thyroid Cancer. Int. J. Mol. Sci. 2024, 25, 10646. https://doi.org/10.3390/ijms251910646
Kim J, Chang H-S, Yun HJ, Chang H-J, Park KC. New Small-Molecule SERCA Inhibitors Enhance Treatment Efficacy in Lenvatinib-Resistant Papillary Thyroid Cancer. International Journal of Molecular Sciences. 2024; 25(19):10646. https://doi.org/10.3390/ijms251910646
Chicago/Turabian StyleKim, Jungmin, Hang-Seok Chang, Hyeok Jun Yun, Ho-Jin Chang, and Ki Cheong Park. 2024. "New Small-Molecule SERCA Inhibitors Enhance Treatment Efficacy in Lenvatinib-Resistant Papillary Thyroid Cancer" International Journal of Molecular Sciences 25, no. 19: 10646. https://doi.org/10.3390/ijms251910646
APA StyleKim, J., Chang, H.-S., Yun, H. J., Chang, H.-J., & Park, K. C. (2024). New Small-Molecule SERCA Inhibitors Enhance Treatment Efficacy in Lenvatinib-Resistant Papillary Thyroid Cancer. International Journal of Molecular Sciences, 25(19), 10646. https://doi.org/10.3390/ijms251910646