
Transcriptomic Analysis of the CNL Gene Family in the Resistant Rice Cultivar IR28 in Response to Ustilaginoidea virens Infection
Abstract
1. Introduction
2. Results
2.1. Disease Symptoms of U. virens-Inoculated IR28 and WX98 Cultivars
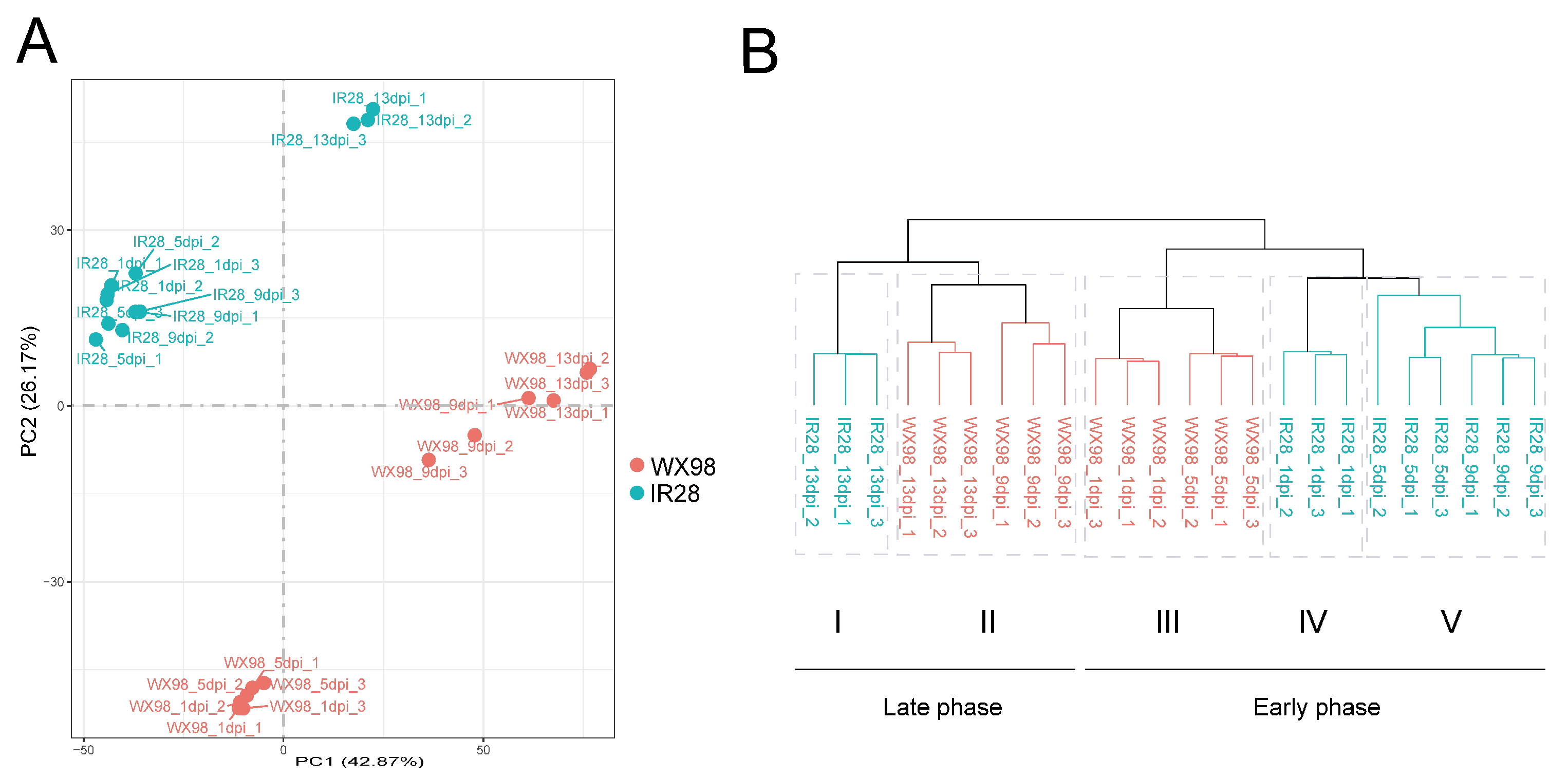
2.2. Transcriptome Profiling and Identification of Differentially Expressed Genes
2.3. GO Enrichment Analysis of Differentially Expressed Genes (DEGs)
2.4. KEGG Pathway Enrichment Analysis of DEGs in Plant–Pathogen Interactions
2.5. Analysis of CNL Genes in Rice in Response to U. virens Infection
2.6. Validation of Candidate Genes Via qRT-PCR
3. Discussion
4. Materials and Methods
4.1. Plant Material and Fungal Strains and U. virens Inoculation
4.2. RNA-Seq of IR28 and WX98 Following U. virens Inoculation
4.3. Differentially Expressed Genes, GO Enrichment, and KEGG Enrichment
4.4. qRT-PCR Evaluation of Resistance-Related Genes and Candidate CNLs
4.5. Principal Component Analysis (PCA) and Heatmap
4.6. Identification of CNL Gene Family Members
4.7. Gene Structure Analysis, Chromosomal Distribution, and Gene Duplication
4.8. Statistical Analyses
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Song, J.H.; Wei, W.; Lv, B.; Lin, Y.; Yin, W.X.; Peng, Y.L.; Schnabel, G.; Huang, J.B.; Jiang, D.H.; Luo, C.X. Rice false smut fungus hijacks the rice nutrients supply by blocking and mimicking the fertilization of rice ovary. Environ. Microbiol. 2016, 18, 3840–3849. [Google Scholar] [CrossRef] [PubMed]
- Wen Ming, W.; Jing, F.; John Martin Jerome, J. Rice false smut: An increasing threat to grain yield and quality. In Protecting Rice Grains in the Post-Genomic Era; Yulin, J., Ed.; IntechOpen: Rijeka, Croatia, 2019; p. Ch.7. [Google Scholar]
- Sun, Q.; Liu, H.; Zhang, Y.; Yi, X.E.; Kong, R.; Cheng, S.; Man, J.; Zheng, L.; Huang, J.; Su, G.; et al. Global distribution of ustiloxins in rice and their male-biased hepatotoxicity. Environ. Pollut. 2022, 301, 118992. [Google Scholar] [CrossRef] [PubMed]
- Zhang, Y.; Xu, Q.; Sun, Q.; Kong, R.; Liu, H.; Yi, X.e.; Liang, Z.; Letcher, R.J.; Liu, C. Ustiloxin a inhibits proliferation of renal tubular epithelial cells in vitro and induces renal injury in mice by disrupting structure and respiratory function of mitochondria. J. Hazard. Mater. 2023, 448, 130791. [Google Scholar] [CrossRef] [PubMed]
- Fan, J.; Yang, J.; Wang, Y.Q.; Li, G.B.; Li, Y.; Huang, F.; Wang, W.M. Current understanding on villosiclava virens, a unique flower-infecting fungus causing rice false smut disease. Mol. Plant Pathol. 2016, 17, 1321–1330. [Google Scholar] [CrossRef] [PubMed]
- Fan, J.; Guo, X.Y.; Li, L.; Huang, F.; Sun, W.X.; Li, Y.; Huang, Y.Y.; Xu, Y.J.; Shi, J.; Lei, Y.; et al. Infection of ustilaginoidea virens intercepts rice seed formation but activates grain-filling-related genes. J. Integr. Plant Biol. 2015, 57, 577–590. [Google Scholar] [CrossRef]
- Zhang, Y.; Zhang, K.; Fang, A.; Han, Y.; Yang, J.; Xue, M.; Bao, J.; Hu, D.; Zhou, B.; Sun, X.; et al. Specific adaptation of ustilaginoidea virens in occupying host florets revealed by comparative and functional genomics. Nat. Commun. 2014, 5, 3849. [Google Scholar] [CrossRef]
- Zhang, M.; Wang, S.; Yuan, M. An update on molecular mechanism of disease resistance genes and their application for genetic improvement of rice. Mol. Breed. 2019, 39, 154. [Google Scholar] [CrossRef]
- Sha, G.; Sun, P.; Kong, X.; Han, X.; Sun, Q.; Fouillen, L.; Zhao, J.; Li, Y.; Yang, L.; Wang, Y.; et al. Genome editing of a rice cdp-dag synthase confers multipathogen resistance. Nature 2023, 618, 1017–1023. [Google Scholar] [CrossRef]
- Li, G.B.; He, J.X.; Wu, J.L.; Wang, H.; Zhang, X.; Liu, J.; Hu, X.H.; Zhu, Y.; Shen, S.; Bai, Y.F. Overproduction of osrack1a, an effector-targeted scaffold protein promoting osrbohb-mediated ros production, confers rice floral resistance to false smut disease without yield penalty. Mol. Plant 2022, 15, 1790–1806. [Google Scholar] [CrossRef]
- Chen, X.; Liu, C.; Wang, H.; Liu, Q.; Yue, Y.; Duan, Y.; Wang, Z.; Zheng, L.; Chen, X.; Wang, Y.; et al. Ustilaginoidea virens-secreted effector uv1809 suppresses rice immunity by enhancing ossrt2-mediated histone deacetylation. Plant Biotechnol. J. 2024, 22, 148–164. [Google Scholar] [CrossRef]
- Li, Q.; Wang, B.; Yu, J.; Dou, D. Pathogen-informed breeding for crop disease resistance. J. Integr. Plant. Bio. 2021, 63, 305–311. [Google Scholar] [CrossRef] [PubMed]
- Andargie, M.; Li, L.; Feng, A.; Zhu, X.; Li, J. Mapping of the quantitative trait locus (qtl) conferring resistance to rice false smut disease. Curr. Plant Biol. 2018, 15, 38–43. [Google Scholar] [CrossRef]
- Zhou, Y.L.; Xie, X.W.; Zhang, F.; Wang, S.; Liu, X.Z.; Zhu, L.H.; Xu, J.L.; Gao, Y.M.; Li, Z.K. Detection of quantitative resistance loci associated with resistance to rice false smut (ustilaginoidea virens) using introgression lines. Plant Pathol. 2014, 63, 365–372. [Google Scholar] [CrossRef]
- Han, Y.; Li, D.; Yang, J.; Huang, F.; Sheng, H.; Sun, W. Mapping quantitative trait loci for disease resistance to false smut of rice. Phytopathol. Res. 2020, 2, 20. [Google Scholar] [CrossRef]
- Fu, R.; Zhao, L.; Chen, C.; Wang, J.; Lu, D. Conjunctive analysis of bsa-seq and ssr markers unveil the candidate genes for resistance to rice false smut. Biomolecules 2024, 14, 79. [Google Scholar] [CrossRef] [PubMed]
- Neelam, K.; Kumar, K.; Kaur, A.; Kishore, A.; Kaur, P.; Babbar, A.; Kaur, G.; Kamboj, I.; Lore, J.S.; Vikal, Y.; et al. High-resolution mapping of the quantitative trait locus (qtls) conferring resistance to false smut disease in rice. J. Appl. Genet. 2022, 63, 35–45. [Google Scholar] [CrossRef] [PubMed]
- Wang, J.; Chai, J. Molecular actions of nlr immune receptors in plants and animals. Sci. China Life Sci. 2020, 63, 1303–1316. [Google Scholar] [CrossRef]
- Maruta, N.; Burdett, H.; Lim, B.Y.J.; Hu, X.; Desa, S.; Manik, M.K.; Kobe, B. Structural basis of nlr activation and innate immune signalling in plants. Immunogenetics 2022, 74, 5–26. [Google Scholar] [CrossRef]
- Lapin, D.; Johanndrees, O. Molecular innovations in plant tir-based immunity signaling. Plant Cell 2022, 34, 1479–1496. [Google Scholar] [CrossRef]
- Shao, Z.Q.; Xue, J.Y.; Wang, Q.; Wang, B.; Chen, J.Q. Revisiting the origin of plant nbs-lrr genes. Trends Plant Sci. 2019, 24, 9–12. [Google Scholar] [CrossRef]
- Förderer, A.; Li, E.; Lawson, A.W.; Deng, Y.N.; Sun, Y.; Logemann, E.; Zhang, X.; Wen, J.; Han, Z.; Chang, J.; et al. A wheat resistosome defines common principles of immune receptor channels. Nature 2022, 610, 532–539. [Google Scholar] [CrossRef] [PubMed]
- Zhao, H.; Wang, X.; Jia, Y.; Minkenberg, B.; Wheatley, M.; Fan, J.; Jia, M.H.; Famoso, A.; Edwards, J.D.; Wamishe, Y. The rice blast resistance gene ptr encodes an atypical protein required for broad-spectrum disease resistance. Nat. Commun. 2018, 9, 2039. [Google Scholar] [CrossRef]
- Ning, X.; Yunyu, W.; Aihong, L. Strategy for use of rice blast resistance genes in rice molecular breeding. Rice Sci. 2020, 27, 263–277. [Google Scholar] [CrossRef]
- Yu, M.; Zhou, Z.; Liu, X.; Yin, D.; Li, D.; Zhao, X.; Li, X.; Li, S.; Chen, R.; Lu, L.; et al. The osspk1–osrac1–rai1 defense signaling pathway is shared by two distantly related nlr proteins in rice blast resistance. Plant Physiol. 2021, 187, 2852–2864. [Google Scholar] [CrossRef]
- Zhang, B.; Zhang, H.; Li, F.; Ouyang, Y.; Yuan, M.; Li, X.; Xiao, J.; Wang, S. Multiple alleles encoding atypical nlrs with unique central tandem repeats in rice confer resistance to xanthomonas oryzae pv. Oryzae. Plant Commun. 2020, 1, 100088. [Google Scholar] [CrossRef] [PubMed]
- Fan, J.; Liu, J.; Gong, Z.Y.; Xu, P.Z.; Hu, X.H.; Wu, J.L.; Li, G.B.; Yang, J.; Wang, Y.Q.; Zhou, Y.F.; et al. The false smut pathogen ustilaginoidea virens requires rice stamens for false smut ball formation. Environ. Microbiol. 2020, 22, 646–659. [Google Scholar] [CrossRef]
- Desmedt, W.; Jonckheere, W.; Nguyen, V.H.; Ameye, M.; De Zutter, N.; De Kock, K.; Debode, J.; Van Leeuwen, T.; Audenaert, K.; Vanholme, B.; et al. The phenylpropanoid pathway inhibitor piperonylic acid induces broad-spectrum pest and disease resistance in plants. Plant Cell Environ. 2021, 44, 3122–3139. [Google Scholar] [CrossRef] [PubMed]
- Ma, H.; Chen, J.; Zhang, Z.; Ma, L.; Yang, Z.; Zhang, Q.; Li, X.; Xiao, J.; Wang, S. Mapk kinase 10.2 promotes disease resistance and drought tolerance by activating different mapks in rice. Plant J. 2017, 92, 557–570. [Google Scholar] [CrossRef]
- Nabavi, S.M.; Šamec, D.; Tomczyk, M.; Milella, L.; Russo, D.; Habtemariam, S.; Suntar, I.; Rastrelli, L.; Daglia, M.; Xiao, J.; et al. Flavonoid biosynthetic pathways in plants: Versatile targets for metabolic engineering. Biotechnol. Adv. 2020, 38, 107316. [Google Scholar] [CrossRef]
- Chen, X.; Tang, J.; Pei, Z.; Liu, H.; Huang, J.; Luo, C.; Tom, H.; Zheng, L. The ‘pears and lemons’ protein uvpal1 regulates development and virulence of ustilaginoidea virens. Environ. Microbiol. 2020, 22, 5414–5432. [Google Scholar] [CrossRef]
- Li, G.-B.; Liu, J.; He, J.-X.; Li, G.-M.; Zhao, Y.-D.; Liu, X.-L.; Hu, X.-H.; Zhang, X.; Wu, J.-L.; Shen, S.; et al. Rice false smut virulence protein subverts host chitin perception and signaling at lemma and palea for floral infection. Plant Cell 2024, 36, 2000–2020. [Google Scholar] [CrossRef] [PubMed]
- Mesa, A.; Basterrech, S.; Guerberoff, G.; Alvarez-Valin, F. Hidden Markov models for gene sequence classification: Classifying the VSG gene in the Trypanosoma brucei genome. Pattern Anal. Appl. 2016, 19, 793–805. [Google Scholar] [CrossRef]
- Wang, Y.; Tang, H.; Debarry, J.D.; Tan, X.; Li, J.; Wang, X.; Lee, T.H.; Jin, H.; Marler, B.; Guo, H.; et al. Mcscanx: A toolkit for detection and evolutionary analysis of gene synteny and collinearity. Nucleic Acids Res. 2012, 40, e49. [Google Scholar] [CrossRef] [PubMed]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Number of Smut Balls | Infected Flower Ratio (%) | |||
|---|---|---|---|---|
| HWD-2 | JS60-2 | HWD-2 | JS60-2 | |
| WX98 | 38.70 ± 24.40 *** | 30.60 ± 24.96 *** | 23.10 ± 13.76 *** | 16.53 ± 10.98 ** |
| IR28 | 6.65 ± 8.13 | 6.60 ± 6.31 | 5.54 ± 6.58 | 5.54 ± 5.33 |
| KEGG ID | Pathway | Gene RATIO | p-Value | Adjusted p-Value | Count |
|---|---|---|---|---|---|
| osa00940 | Phenylpropanoid biosynthesis | 38/302 | 7.10 × 10−8 | 7.24 × 10−6 | 38 |
| osa00941 | Flavonoid biosynthesis | 11/302 | 2.10 × 10−5 | 1.073 × 10−3 | 11 |
| osa04626 | Plant–pathogen interaction | 26/302 | 4.03 × 10−4 | 1.37 × 10−2 | 26 |
| Gene IDs | Gene_Description | Domains | 1 Day Post Inoculation | 5 Days Post Inoculation | ||
|---|---|---|---|---|---|---|
| log2FoldChange | Adjusted p-Value | log2FoldChange | Adjusted p-Value | |||
| CNGCs: | ||||||
| BGIOSGA020131 | cyclic nucleotide-binding protein | CAP_ED | 4.98 | 2.74 × 10−21 | 5.02 | 6.59 × 10−23 |
| CDPK: | ||||||
| BGIOSGA014554 | calcium-dependent protein kinase | PKc_like | 1.76 | 3.73 × 10−13 | 2.04 | 1.65 × 10−11 |
| Rboh: | ||||||
| BGIOSGA004406 | NADPH_Ox and Efh protein | NADPH_Ox | 0.65 | 2.48 × 10−3 | 1.13 | 4.46 × 10−4 |
| BGIOSGA001663 | Respiratory burst oxidase homolog protein B | NADPH_Ox, FNR_like, FAD_binding_1, | 1.10 | 5.50 × 10−19 | 1.14 | 4.71 × 10−6 |
| CaMCML: | ||||||
| BGIOSGA000087 | EF-hand | PTZ00183 | 3.05 | 1.95 × 10−90 | 4.24 | 4.88 × 10−35 |
| BGIOSGA000109 | EF-hand | PTZ00184 | 1.57 | 4.49 × 10−31 | 1.75 | 1.20 × 10−18 |
| BGIOSGA002446 | EF-hand | PTZ00184 | 2.26 | 2.86 × 10−52 | 2.38 | 4.50 × 10−22 |
| BGIOSGA017499 | EF-hand | PTZ00184 | 3.86 | 2.05 × 10−61 | 2.85 | 4.18 × 10−9 |
| BGIOSGA018321 | EF-hand | PTZ00184 | 0.13 | 9.02 × 10−1 | 1.32 | 1.294 × 10−2 |
| BGIOSGA018556 | EF-hand | PTZ00184 | 1.13 | 3.89 × 10−20 | 1.10 | 5.40 × 10−6 |
| BGIOSGA018153 | EF-hand | PTZ00184 | 1.50 | 2.97 × 10−27 | 1.63 | 1.02 × 10−6 |
| BGIOSGA034766 | EF-hand | Efh | 0.98 | 2.42 × 10−6 | 1.00 | 9.81 × 10−3 |
| MPK3/6: | ||||||
| BGIOSGA010959 | Mitogen-activated protein kinase | PKc_like | 2.23 | 3.80 × 10−56 | 1.01 | 3.98 × 10−5 |
| WRKY33: | ||||||
| BGIOSGA030067 | WRKY transcription factor | WRKY | 2.68 | 1.67 × 10−4 | 3.35 | 2.29 × 10−4 |
| WRKY22: | ||||||
| BGIOSGA001178 | WRKY transcription factor | WRKY | 1.54 | 8.68 × 10−7 | 1.39 | 1.93 × 10-4 |
| PR1: | ||||||
| BGIOSGA001599 | pathogenesis-related family 1 (PR-1) | CAP | 11.32 | 7.16 × 10−40 | 12.56 | 3.64 × 10−31 |
| BGIOSGA025083 | pathogenesis-related family 1 (PR-1) | CAP | 9.71 | 9.94 × 10−15 | 2.70 | 5.06 × 10−7 |
| BGIOSGA025088 | pathogenesis-related family 1 (PR-1) | CAP | 1.95 | 8.07 × 10−47 | 1.51 | 4.71 × 10−8 |
| BGIOSGA025078 | pathogenesis-related family 1 (PR-1) | CAP | 5.01 | 5.52 × 10−112 | 2.82 | 8.57 × 10−5 |
| RPM1: | ||||||
| BGIOSGA036090 | NB-LRR disease-resistance protein | RX-CC, NB-ARC, LRR | 9.88 | 2.50 × 10−15 | 9.51 | 1.46 × 10−13 |
| BGIOSGA015563 | NB-LRR disease-resistance protein | RX-CC, NB-ARC | 4.84 | 1.11 × 10−2 | 5.58 | 2.34 × 10−3 |
| BGIOSGA030404 | NB-LRR disease resistance | RX-CC, NB-ARC, LRR | 0.40 | 2.70 × 10−1 | 1.94 | 1.49 × 10−6 |
| HSP90: | ||||||
| BGIOSGA031149 | HSP90 | 7.51 | 3.15 × 10−8 | 7.74 | 1.50 × 10−8 | |
| KCS1/10: | ||||||
| BGIOSGA011802 | PLN02192 | 8.59 | 2.27 × 10−11 | 8.69 | 1.28 × 10−10 | |
| BGIOSGA023203 | PLN02192 | 9.29 | 6.89 × 10−14 | 7.26 | 8.30 × 10−15 | |
| BGIOSGA010624 | type III polyketide synthase | CHS | 3.72 | 1.89 × 10−2 | 3.35 | 8.87 × 10−7 |
| Gene IDs | Chr No. | Gene Length | Subcellular Location | 1 Day Post Inoculation | 5 Days Post Inoculation | ||
|---|---|---|---|---|---|---|---|
| log2FoldChange | Adjusted p-Value | log2FoldChange | Adjusted p-Value | ||||
| BGIOSGA003230 | 1 | 2916 | cyto * | 4.93 | 9.03 × 10−27 | 2.37 | 1.97 × 10−7 |
| BGIOSGA003329 | 1 | 2400 | cyto | 5.26 | 6.44 × 10−74 | 5.14 | 6.66 × 10−40 |
| BGIOSGA028077 | 8 | 3954 | plas | 6.56 | 2.15 × 10−78 | 1.87 | 2.36 × 10−18 |
| BGIOSGA029433 | 9 | 2937 | cyto | 1.28 | 9.39 × 10−9 | 1.24 | 2.84 × 10−5 |
| BGIOSGA032258 | 10 | 2415 | plas | 9.64 | 2.32 × 10−14 | 2.06 | 2.64 × 10−5 |
| BGIOSGA032326 | 10 | 2187 | nucl | 2.35 | 8.25 × 10−24 | 2.32 | 3.26 × 10−10 |
| BGIOSGA032859 | 10 | 1080 | cyto | 1.06 | 2.27 × 10−6 | 1.34 | 1.68 × 10−5 |
| BGIOSGA033963 | 11 | 3246 | nucl | 11.19 | 7.76 × 10−20 | 10.36 | 5.71 × 10−16 |
| BGIOSGA035581 | 11 | 2940 | nucl | 2.66 | 1.28 × 10−57 | 1.89 | 2.17 × 10−17 |
| BGIOSGA035675 | 11 | 3912 | chlo | 2.05 | 3.07 × 10−24 | 2.11 | 2.17 × 10−20 |
| BGIOSGA036090 | 12 | 3243 | cyto | 9.88 | 2.5 × 10−15 | 9.51 | 1.46 × 10−13 |
| BGIOSGA036720 | 12 | 3993 | mito | 4.86 | 1.6 × 10−83 | 2.63 | 5.23 × 10−20 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Wang, Z.-Q.; Wang, Y.-F.; Xu, T.; Li, X.-Y.; Zhang, S.; Chang, X.-Q.; Yang, X.-L.; Meng, S.; Lv, L. Transcriptomic Analysis of the CNL Gene Family in the Resistant Rice Cultivar IR28 in Response to Ustilaginoidea virens Infection. Int. J. Mol. Sci. 2024, 25, 10655. https://doi.org/10.3390/ijms251910655
Wang Z-Q, Wang Y-F, Xu T, Li X-Y, Zhang S, Chang X-Q, Yang X-L, Meng S, Lv L. Transcriptomic Analysis of the CNL Gene Family in the Resistant Rice Cultivar IR28 in Response to Ustilaginoidea virens Infection. International Journal of Molecular Sciences. 2024; 25(19):10655. https://doi.org/10.3390/ijms251910655
Chicago/Turabian StyleWang, Zuo-Qian, Yu-Fu Wang, Ting Xu, Xin-Yi Li, Shu Zhang, Xiang-Qian Chang, Xiao-Lin Yang, Shuai Meng, and Liang Lv. 2024. "Transcriptomic Analysis of the CNL Gene Family in the Resistant Rice Cultivar IR28 in Response to Ustilaginoidea virens Infection" International Journal of Molecular Sciences 25, no. 19: 10655. https://doi.org/10.3390/ijms251910655
APA StyleWang, Z.-Q., Wang, Y.-F., Xu, T., Li, X.-Y., Zhang, S., Chang, X.-Q., Yang, X.-L., Meng, S., & Lv, L. (2024). Transcriptomic Analysis of the CNL Gene Family in the Resistant Rice Cultivar IR28 in Response to Ustilaginoidea virens Infection. International Journal of Molecular Sciences, 25(19), 10655. https://doi.org/10.3390/ijms251910655