
Combined Integrative RNA-Seq and Serological sIgE Analysis Enhances Understanding of Fish Allergen Profiles and Diagnostic Strategy for Fish Allergy
 , , ,
, , ,  , , , , , , , ,
, , , , , , , ,  add
Show full author list
add
Show full author list
Abstract
:
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
1. Introduction
2. Results
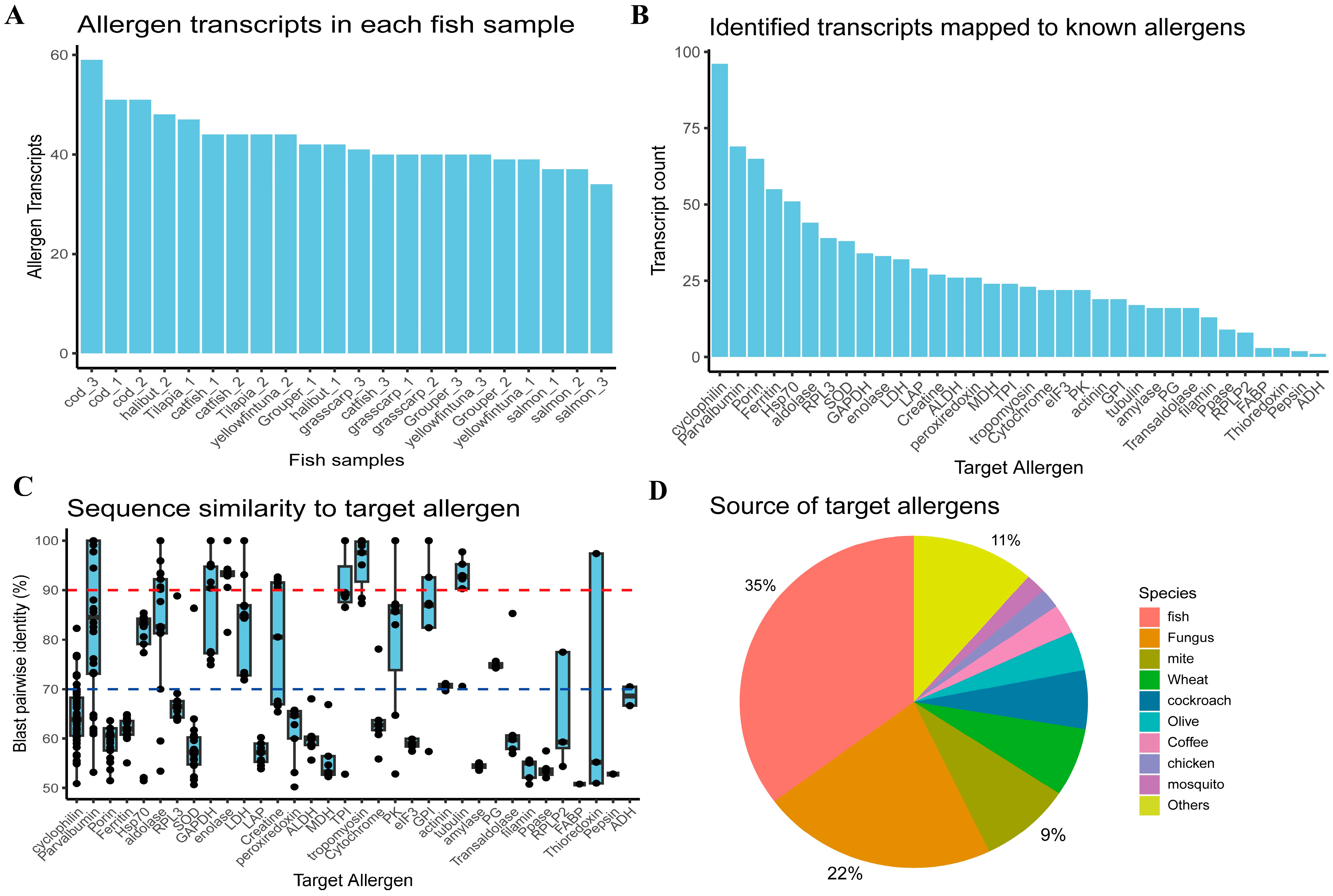
2.1. Overview of Potential Allergen Transcripts Identified by Fish RNA-Seq Analysis
2.2. Overall Fish Allergen Profiles and Top Expressed Allergens Distinguish Fish into Two Categories
2.3. Fish Allergen Profiles Improve Interpretation of sIgE Sensitization Patterns
2.4. Diagnosis Performance of sIgE Data Indicated Rational CRD Designs for Different Fish Allergy
3. Discussion
4. Materials and Methods
4.1. Collection of Fish RNA-Seq and ImmunoCap sIgE Data
4.2. RNA-Seq Data Analysis and Allergen Identification
4.3. Parvalbumin Conservation Score Calculation
4.4. ROC Analysis
4.5. Statistics and Figure Plots
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Lin, H.Y.; Wright, S.; Costello, M.J. Numbers of fish species, higher taxa, and phylogenetic similarity decrease with latitude and depth, and deep-sea assemblages are unique. PeerJ 2023, 11, e16116. [Google Scholar] [CrossRef]
- Chang, F.; Eng, L.; Chang, C. Food Allergy Labeling Laws: International Guidelines for Residents and Travelers. Clin. Rev. Allerg. Immu. 2023, 65, 148–165. [Google Scholar] [CrossRef] [PubMed]
- Moonesinghe, H.; Mackenzie, H.; Venter, C.; Kilburn, S.; Turner, P.; Weir, K.; Dean, T. Prevalence of fish and shellfish allergy: A systematic review. Ann. Allergy Asthma Immunol. 2016, 117, 264–272.e4. [Google Scholar] [CrossRef] [PubMed]
- Connett, G.J.; Gerez, I.; Cabrera-Morales, E.A.; Yuenyongviwat, A.; Ngamphaiboon, J.; Chatchatee, P.; Sangsupawanich, P.; Soh, S.E.; Yap, G.C.; Shek, L.P.; et al. A population-based study of fish allergy in the Philippines, Singapore and Thailand. Int. Arch. Allergy Immunol. 2012, 159, 384–390. [Google Scholar] [CrossRef]
- Wai, C.Y.Y.; Leung, N.Y.H.; Leung, A.S.Y.; Wong, G.W.K.; Leung, T.F. Seafood Allergy in Asia: Geographical Specificity and Beyond. Front. Allergy 2021, 2, 676903. [Google Scholar] [CrossRef]
- Okamoto, M.; Takafuji, S.; Inoue, S.; Tanaka, Y. Fish allergy tolerance 16 months after diagnosis. Allergol Immunopathol. 2021, 49, 25–27. [Google Scholar] [CrossRef]
- Sharp, M.F.; Lopata, A.L. Fish allergy: In review. Clin. Rev. Allergy Immunol. 2014, 46, 258–271. [Google Scholar] [CrossRef]
- Tong, W.S.; Yuen, A.W.; Wai, C.Y.; Leung, N.Y.; Chu, K.H.; Leung, P.S. Diagnosis of fish and shellfish allergies. J. Asthma Allergy 2018, 11, 247–260. [Google Scholar] [CrossRef]
- Yuk, J.E.; Lee, J.; Jeong, K.Y.; Park, K.H.; Kim, J.D.; Kim, J.T.; Lee, J.H.; Park, J.W. Allergenicity and Stability of 6 New Korean Bony Fish Extracts. Allergy Asthma Immunol. Res. 2021, 13, 623–637. [Google Scholar] [CrossRef]
- Armentia, A.; Santos, J.; Serrano, Z.; Martin, B.; Martin, S.; Barrio, J.; Fernandez, S.; Gonzalez-Sagrado, M.; Pineda, F.; Palacios, R. Molecular diagnosis of allergy to Anisakis simplex and Gymnorhynchus gigas fish parasites. Allergol. Immunopathol. 2017, 45, 463–472. [Google Scholar] [CrossRef]
- Klueber, J.; Schrama, D.; Rodrigues, P.; Dickel, H.; Kuehn, A. Fish Allergy Management: From Component-Resolved Diagnosis to Unmet Diagnostic Needs. Curr. Treat. Options Allergy 2019, 6, 322–337. [Google Scholar] [CrossRef]
- Leung, A.S.-Y.; Fusayasu, N.; Álvarez, L.A.; Gu, Y.; Ebisawa, M.; Wong, G.W.-K. Fish allergy management: Should fish be completely avoided? The pros and cons debate. J. Allergy Hypersensitivity Dis. 2024, 2, 100008. [Google Scholar] [CrossRef]
- Dramburg, S.; Hilger, C.; Santos, A.F.; de Las Vecillas, L.; Aalberse, R.C.; Acevedo, N.; Aglas, L.; Altmann, F.; Arruda, K.L.; Asero, R.; et al. EAACI Molecular Allergology User’s Guide 2.0. Pediatr. Allergy Immunol. 2023, 34 (Suppl. 28), e13854. [Google Scholar] [CrossRef] [PubMed]
- Seth, D.; Poowutikul, P.; Pansare, M.; Kamat, D. Food Allergy: A Review. Pediatr. Ann. 2020, 49, e50–e58. [Google Scholar] [CrossRef]
- Karnaneedi, S.; Huerlimann, R.; Johnston, E.B.; Nugraha, R.; Ruethers, T.; Taki, A.C.; Kamath, S.D.; Wade, N.M.; Jerry, D.R.; Lopata, A.L. Novel Allergen Discovery through Comprehensive De Novo Transcriptomic Analyses of Five Shrimp Species. Int. J. Mol. Sci. 2020, 22, 32. [Google Scholar] [CrossRef]
- Vo, T.T.M.; Amoroso, G.; Ventura, T.; Elizur, A. Histological and transcriptomic analysis of muscular atrophy associated with depleted flesh pigmentation in Atlantic salmon (Salmo salar) exposed to elevated seawater temperatures. Sci. Rep. 2023, 13, 4218. [Google Scholar] [CrossRef]
- Vo, T.T.M.; Nguyen, T.V.; Amoroso, G.; Ventura, T.; Elizur, A. Deploying new generation sequencing for the study of flesh color depletion in Atlantic Salmon (Salmo salar). BMC Genom. 2021, 22, 545. [Google Scholar] [CrossRef]
- Wang, X.; Liu, G.; Xie, S.; Pan, L.; Tan, Q. Growth and Meat Quality of Grass Carp (Ctenopharyngodon idellus) Responded to Dietary Protein (Soybean meal) Level Through the Muscle Metabolism and Gene Expression of Myosin Heavy Chains. Front. Nutr. 2022, 9, 833924. [Google Scholar] [CrossRef]
- Lin, G.; Thevasagayam, N.M.; Wan, Z.Y.; Ye, B.Q.; Yue, G.H. Transcriptome Analysis Identified Genes for Growth and Omega-3/-6 Ratio in Saline Tilapia. Front. Genet. 2019, 10, 244. [Google Scholar] [CrossRef]
- Goodman, R.E.; Ebisawa, M.; Ferreira, F.; Sampson, H.A.; van Ree, R.; Vieths, S.; Baumert, J.L.; Bohle, B.; Lalithambika, S.; Wise, J.; et al. AllergenOnline: A peer-reviewed, curated allergen database to assess novel food proteins for potential cross-reactivity. Mol. Nutr. Food Res. 2016, 60, 1183–1198. [Google Scholar] [CrossRef]
- Aalberse, R.C. Structural biology of allergens. J. Allergy Clin. Immunol. 2000, 106, 228–238. [Google Scholar] [CrossRef] [PubMed]
- Nugraha, R.; Kamath, S.D.; Johnston, E.; Zenger, K.R.; Rolland, J.M.; O’Hehir, R.E.; Lopata, A.L. Rapid and comprehensive discovery of unreported shellfish allergens using large-scale transcriptomic and proteomic resources. J. Allergy Clin. Immunol. 2018, 141, 1501–1504.e8. [Google Scholar] [CrossRef] [PubMed]
- Hamada, Y.; Nagashima, Y.; Shiomi, K. Identification of collagen as a new fish allergen. Biosci. Biotechnol. Biochem. 2001, 65, 285–291. [Google Scholar] [CrossRef]
- Kleinnijenhuis, A.J.; van Holthoon, F.L. Domain-Specific Proteogenomic Analysis of Collagens to Evaluate De Novo Sequencing Results and Database Information. J. Mol. Evol. 2018, 86, 293–302. [Google Scholar] [CrossRef]
- Wai, C.Y.Y.; Leung, N.Y.H.; Leung, A.S.Y.; Fusayasu, N.; Sato, S.; Xu, K.J.Y.; Yau, Y.S.; Rosa Duque, J.S.; Kwan, M.Y.W.; Cheng, J.; et al. Differential patterns of fish sensitization in Asian populations: Implication for precision diagnosis. Allergol. Int. 2023, 72, 458–465. [Google Scholar] [CrossRef]
- Kuehn, A.; Hilger, C.; Lehners-Weber, C.; Codreanu-Morel, F.; Morisset, M.; Metz-Favre, C.; Pauli, G.; de Blay, F.; Revets, D.; Muller, C.P.; et al. Identification of enolases and aldolases as important fish allergens in cod, salmon and tuna: Component resolved diagnosis using parvalbumin and the new allergens. Clin. Exp. Allergy 2013, 43, 811–822. [Google Scholar] [CrossRef]
- Ruethers, T.; Kamath, S.; Taki, A.; Le, T.; Karnaneedi, S.; Nugraha, R.; Cao, T.; Nie, S.; Williamson, N.; Mehr, S.; et al. Tropomyosin Is a Novel Major Fish Allergen of Unrecognized Importance. J. Allergy Clin. Immun. 2020, 145, Ab226. [Google Scholar] [CrossRef]
- Liu, R.; Krishnan, H.B.; Xue, W.; Liu, C. Characterization of allergens isolated from the freshwater fish blunt snout bream (Megalobrama amblycephala). J. Agric. Food Chem. 2011, 59, 458–463. [Google Scholar] [CrossRef]
- Kobayashi, A.; Tanaka, H.; Hamada, Y.; Ishizaki, S.; Nagashima, Y.; Shiomi, K. Comparison of allergenicity and allergens between fish white and dark muscles. Allergy 2006, 61, 357–363. [Google Scholar] [CrossRef]
- Li, S.; Chu, K.H.; Wai, C.Y.Y. Genomics of Shrimp Allergens and Beyond. Genes 2023, 14, 2145. [Google Scholar] [CrossRef]
- Li, J.; Li, Z.; Kong, D.; Li, S.; Yu, Y.; Li, H. IgE and IgG4 responses to shrimp allergen tropomyosin and its epitopes in patients from coastal areas of northern China. Mol. Med. Rep. 2020, 22, 371–379. [Google Scholar] [CrossRef] [PubMed]
- Leung, A.S.Y.; Wai, C.Y.Y.; Leung, N.Y.H.; Ngai, N.A.; Chua, G.T.; Ho, P.K.; Lam, I.C.S.; Cheng, J.; Chan, O.M.; Li, P.F.; et al. Real-World Sensitization and Tolerance Pattern to Seafood in Fish-Allergic Individuals. J. Allergy Clin. Immunol. Pract. 2023, 12, 633–642. [Google Scholar] [CrossRef] [PubMed]
- Kuehn, A.; Scheuermann, T.; Hilger, C.; Hentges, F. Important variations in parvalbumin content in common fish species: A factor possibly contributing to variable allergenicity. Int. Arch. Allergy Immunol. 2010, 153, 359–366. [Google Scholar] [CrossRef] [PubMed]
- Kalic, T.; Morel-Codreanu, F.; Radauer, C.; Ruethers, T.; Taki, A.C.; Swoboda, I.; Hilger, C.; Hoffmann-Sommergruber, K.; Ollert, M.; Hafner, C.; et al. Patients Allergic to Fish Tolerate Ray Based on the Low Allergenicity of Its Parvalbumin. J. Allergy Clin. Immunol. Pract. 2019, 7, 500–508.e11. [Google Scholar] [CrossRef]
- Ruethers, T.; Taki, A.C.; Nugraha, R.; Cao, T.T.; Koeberl, M.; Kamath, S.D.; Williamson, N.A.; O’Callaghan, S.; Nie, S.; Mehr, S.S.; et al. Variability of allergens in commercial fish extracts for skin prick testing. Allergy 2019, 74, 1352–1363. [Google Scholar] [CrossRef]
- Kuehn, A.; Codreanu-Morel, F.; Lehners-Weber, C.; Doyen, V.; Gomez-Andre, S.A.; Bienvenu, F.; Fischer, J.; Ballardini, N.; van Hage, M.; Perotin, J.M.; et al. Cross-reactivity to fish and chicken meat—A new clinical syndrome. Allergy 2016, 71, 1772–1781. [Google Scholar] [CrossRef]
- Hilger, C.; Thill, L.; Grigioni, F.; Lehners, C.; Falagiani, P.; Ferrara, A.; Romano, C.; Stevens, W.; Hentges, F. IgE antibodies of fish allergic patients cross-react with frog parvalbumin. Allergy 2004, 59, 653–660. [Google Scholar] [CrossRef]
- Leung, N.Y.H.; Leung, A.S.Y.; Xu, K.J.Y.; Wai, C.Y.Y.; Lam, C.Y.; Wong, G.W.K.; Leung, T.F. Molecular and immunological characterization of grass carp (Ctenopharyngodon idella) parvalbumin Cten i 1: A major fish allergen in Hong Kong. Pediatr. Allergy Immunol. 2020, 31, 792–804. [Google Scholar] [CrossRef]
- Kuehn, A.; Swobode, I.; Arumugam, K.; Hilger, C.; Hentges, F. Fish allergens at a glance: Variable allergenicity of parvalbumins, the major fish allergens. Front. Immunol. 2014, 5, 179. [Google Scholar] [CrossRef]
- Ruethers, T.; Taki, A.C.; Karnaneedi, S.; Nie, S.; Kalic, T.; Dai, D.Y.; Daduang, S.; Leeming, M.; Williamson, N.A.; Breiteneder, H.; et al. Expanding the allergen repertoire of salmon and catfish. Allergy 2021, 76, 1443–1453. [Google Scholar] [CrossRef]
- Sun, Y.B.; Luo, Y.Q.; Chen, J.; Liu, X.; Gao, J.Y.; Xie, Y.H.; Chen, H.B. Fish Allergy: A Review of Clinical Characteristics, Mechanism, Allergens, Epitopes, and Cross-Reactivity. Acs Food Sci. Technol. 2024, 4, 304–315. [Google Scholar] [CrossRef]
- Mukherjee, S.; Horka, P.; Zdenkova, K.; Cermakova, E. Parvalbumin: A Major Fish Allergen and a Forensically Relevant Marker. Genes 2023, 14, 223. [Google Scholar] [CrossRef] [PubMed]
- Taki, A.C.; Ruethers, T.; Nugraha, R.; Karnaneedi, S.; Williamson, N.A.; Nie, S.; Leeming, M.G.; Mehr, S.S.; Campbell, D.E.; Lopata, A.L. Thermostable allergens in canned fish: Evaluating risks for fish allergy. Allergy 2023, 78, 3221–3234. [Google Scholar] [CrossRef] [PubMed]
- Thalayasingam, M.; Lee, B.W. Fish and Shellfish Allergy. In Food Allergy: Molecular Basis and Clinical Practice; Karger Publishers: Basel, Switzerland, 2015; Volume 101, pp. 152–161. [Google Scholar]
- Kobayashi, Y.; Yang, T.; Yu, C.T.; Ume, C.; Kubota, H.; Shimakura, K.; Shiomi, K.; Hamada-Sato, N. Quantification of major allergen parvalbumin in 22 species of fish by SDS-PAGE. Food Chem. 2016, 194, 345–353. [Google Scholar] [CrossRef]
- Kalic, T.; Radauer, C.; Lopata, A.L.; Breiteneder, H.; Hafner, C. Fish Allergy Around the World-Precise Diagnosis to Facilitate Patient Management. Front. Allergy 2021, 2, 732178. [Google Scholar] [CrossRef]
- Bernhisel-Broadbent, J.; Strause, D.; Sampson, H.A. Fish hypersensitivity. II: Clinical relevance of altered fish allergenicity caused by various preparation methods. J. Allergy Clin. Immunol. 1992, 90, 622–629. [Google Scholar] [CrossRef]
- Dijkema, D.; Emons, J.A.M.; Van de Ven, A.A.J.M.; Elberink, J.N.G.O. Fish Allergy: Fishing for Novel Diagnostic and Therapeutic Options. Clin. Rev. Allerg. Immu. 2022, 62, 64–71. [Google Scholar] [CrossRef]
- Wang, M.P.; Thomas, G.N.; Ho, S.Y.; Lai, H.K.; Mak, K.H.; Lam, T.H. Fish consumption and mortality in Hong Kong Chinese—The LIMOR study. Ann. Epidemiol. 2011, 21, 164–169. [Google Scholar] [CrossRef]
- Zhao, H.; Wang, M.; Peng, X.; Zhong, L.; Liu, X.; Shi, Y.; Li, Y.; Chen, Y.; Tang, S. Fish consumption in multiple health outcomes: An umbrella review of meta-analyses of observational and clinical studies. Ann. Transl. Med. 2023, 11, 152. [Google Scholar] [CrossRef]
- Schoos, A.M.; Hansen, S.M.; Skov, F.R.; Stokholm, J.; Bonnelykke, K.; Bisgaard, H.; Chawes, B.L. Allergen Specificity in Specific IgE Cutoff. JAMA Pediatr. 2020, 174, 993–995. [Google Scholar] [CrossRef]
- Chen, S.; Zhou, Y.; Chen, Y.; Gu, J. fastp: An ultra-fast all-in-one FASTQ preprocessor. Bioinformatics 2018, 34, i884–i890. [Google Scholar] [CrossRef] [PubMed]
- Song, L.; Florea, L. Rcorrector: Efficient and accurate error correction for Illumina RNA-seq reads. Gigascience 2015, 4, 48. [Google Scholar] [CrossRef]
- Grabherr, M.G.; Haas, B.J.; Yassour, M.; Levin, J.Z.; Thompson, D.A.; Amit, I.; Adiconis, X.; Fan, L.; Raychowdhury, R.; Zeng, Q.; et al. Full-length transcriptome assembly from RNA-Seq data without a reference genome. Nat. Biotechnol. 2011, 29, 644–652. [Google Scholar] [CrossRef] [PubMed]
- Simao, F.A.; Waterhouse, R.M.; Ioannidis, P.; Kriventseva, E.V.; Zdobnov, E.M. BUSCO: Assessing genome assembly and annotation completeness with single-copy orthologs. Bioinformatics 2015, 31, 3210–3212. [Google Scholar] [CrossRef]
- Smith-Unna, R.; Boursnell, C.; Patro, R.; Hibberd, J.M.; Kelly, S. TransRate: Reference-free quality assessment of de novo transcriptome assemblies. Genome Res. 2016, 26, 1134–1144. [Google Scholar] [CrossRef]
- Gasteiger, E.; Gattiker, A.; Hoogland, C.; Ivanyi, I.; Appel, R.D.; Bairoch, A. ExPASy: The proteomics server for in-depth protein knowledge and analysis. Nucleic Acids Res. 2003, 31, 3784–3788. [Google Scholar] [CrossRef]
- Kumar, S.; Stecher, G.; Tamura, K. MEGA7: Molecular Evolutionary Genetics Analysis Version 7.0 for Bigger Datasets. Mol. Biol. Evol. 2016, 33, 1870–1874. [Google Scholar] [CrossRef]
- Bodenhofer, U.; Bonatesta, E.; Horejs-Kainrath, C.; Hochreiter, S. msa: An R package for multiple sequence alignment. Bioinformatics 2015, 31, 3997–3999. [Google Scholar] [CrossRef]
- Crooks, G.E.; Hon, G.; Chandonia, J.M.; Brenner, S.E. WebLogo: A sequence logo generator. Genome Res. 2004, 14, 1188–1190. [Google Scholar] [CrossRef]
- Sperandei, S. Understanding logistic regression analysis. Biochem. Med. 2014, 24, 12–18. [Google Scholar] [CrossRef]
- Zhang, Z. Variable selection with stepwise and best subset approaches. Ann. Transl. Med. 2016, 4, 136. [Google Scholar] [CrossRef]



Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Liu, Z.-Y.; Wai, C.Y.Y.; Leung, A.S.Y.; Chan, W.H.; Rosa Duque, J.S.; Lam, I.C.S.; Cheng, J.W.; Sit, J.K.C.; Ngai, N.A.; Ho, P.K.; et al. Combined Integrative RNA-Seq and Serological sIgE Analysis Enhances Understanding of Fish Allergen Profiles and Diagnostic Strategy for Fish Allergy. Int. J. Mol. Sci. 2024, 25, 10784. https://doi.org/10.3390/ijms251910784
Liu Z-Y, Wai CYY, Leung ASY, Chan WH, Rosa Duque JS, Lam ICS, Cheng JW, Sit JKC, Ngai NA, Ho PK, et al. Combined Integrative RNA-Seq and Serological sIgE Analysis Enhances Understanding of Fish Allergen Profiles and Diagnostic Strategy for Fish Allergy. International Journal of Molecular Sciences. 2024; 25(19):10784. https://doi.org/10.3390/ijms251910784
Chicago/Turabian StyleLiu, Zhong-Yi, Christine Yee Yan Wai, Agnes Sze Yin Leung, Wai Hung Chan, Jaime Sou Rosa Duque, Ivan Cheuk San Lam, James Wesley Cheng, Jason Ka Chun Sit, Noelle Anne Ngai, Po Ki Ho, and et al. 2024. "Combined Integrative RNA-Seq and Serological sIgE Analysis Enhances Understanding of Fish Allergen Profiles and Diagnostic Strategy for Fish Allergy" International Journal of Molecular Sciences 25, no. 19: 10784. https://doi.org/10.3390/ijms251910784
APA StyleLiu, Z.-Y., Wai, C. Y. Y., Leung, A. S. Y., Chan, W. H., Rosa Duque, J. S., Lam, I. C. S., Cheng, J. W., Sit, J. K. C., Ngai, N. A., Ho, P. K., Chua, G. T., Lee, Q. U., Chan, O. M., Yau, Y. S., Wong, J. S. C., Luk, D. C. K., Ho, M. H. K., Kwan, M. Y. W., Tang, M. F., ... Leung, T. F. (2024). Combined Integrative RNA-Seq and Serological sIgE Analysis Enhances Understanding of Fish Allergen Profiles and Diagnostic Strategy for Fish Allergy. International Journal of Molecular Sciences, 25(19), 10784. https://doi.org/10.3390/ijms251910784