
CRISPR/Cas9-Mediated fech Knockout Zebrafish: Unraveling the Pathogenesis of Erythropoietic Protoporphyria and Facilitating Drug Screening

 , , ,
, , ,
Abstract
1. Introduction
2. Results
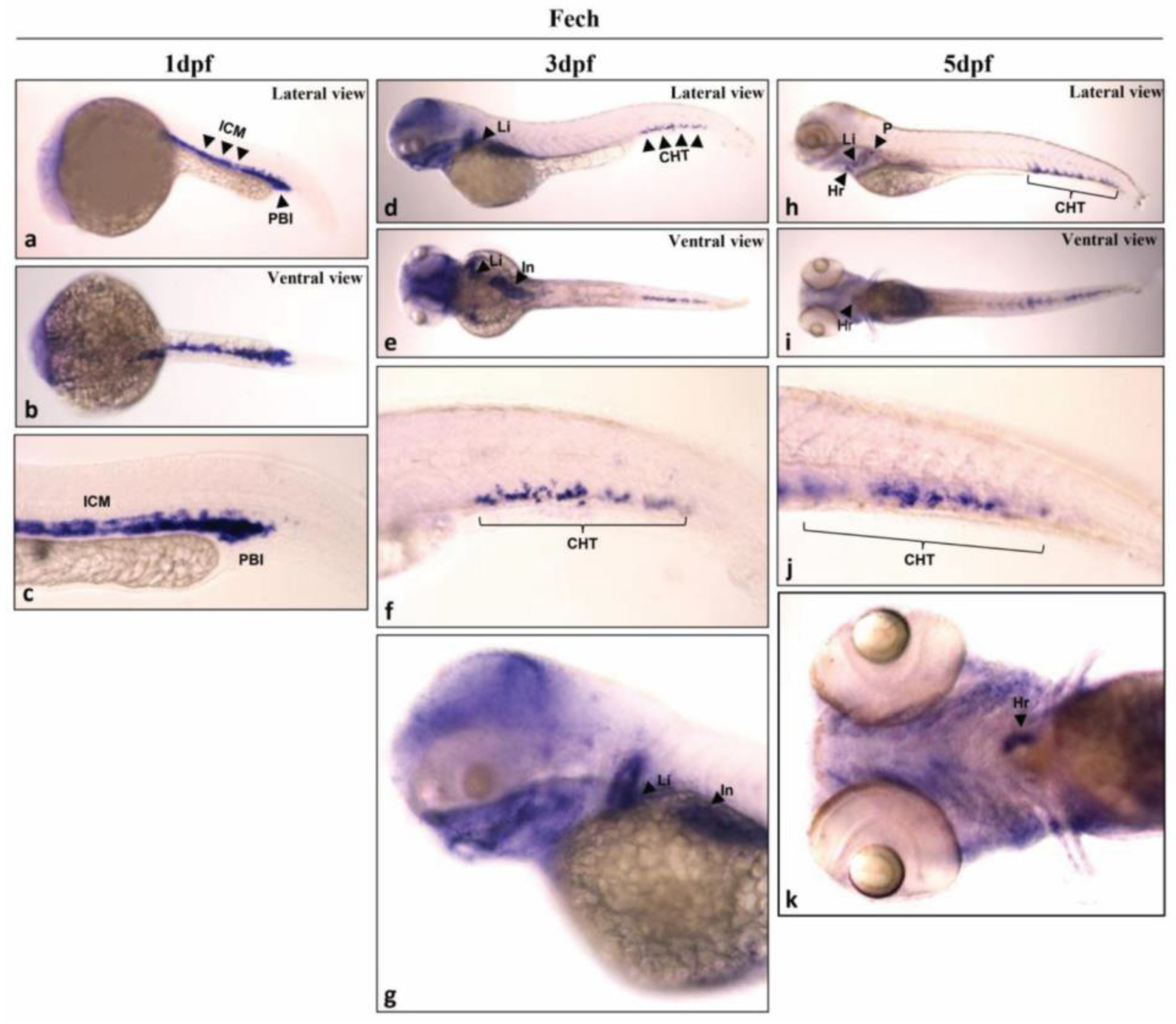
2.1. Analysis of Fech Expression in Different Larval Stages and Tissues of Zebrafish
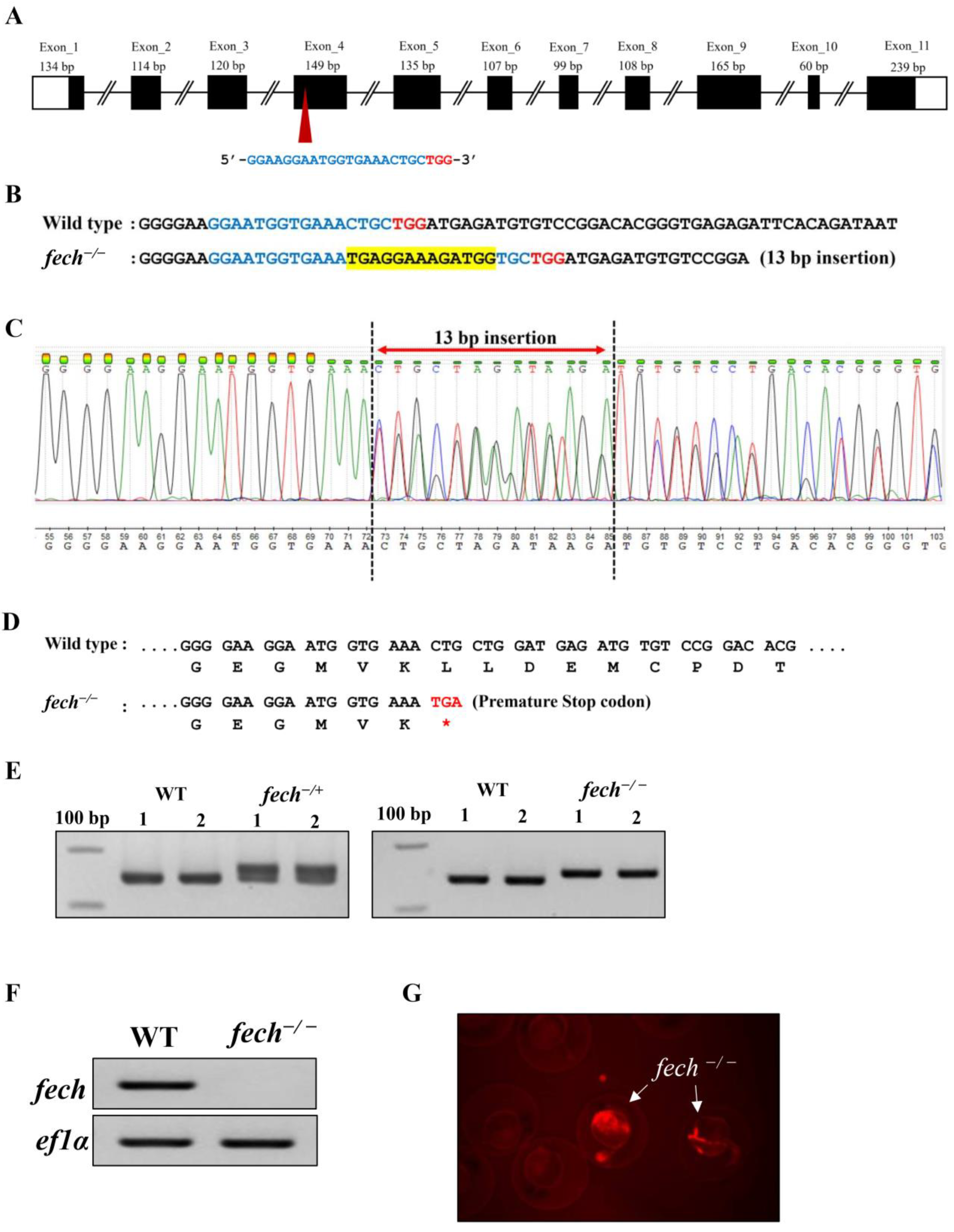
2.2. Generation of a Fech-Knockout Zebrafish Model Using CRISPR/Cas9 Gene Editing
2.3. Phenotypic and Physiological Effects of Fech Deletion in Zebrafish Larvae
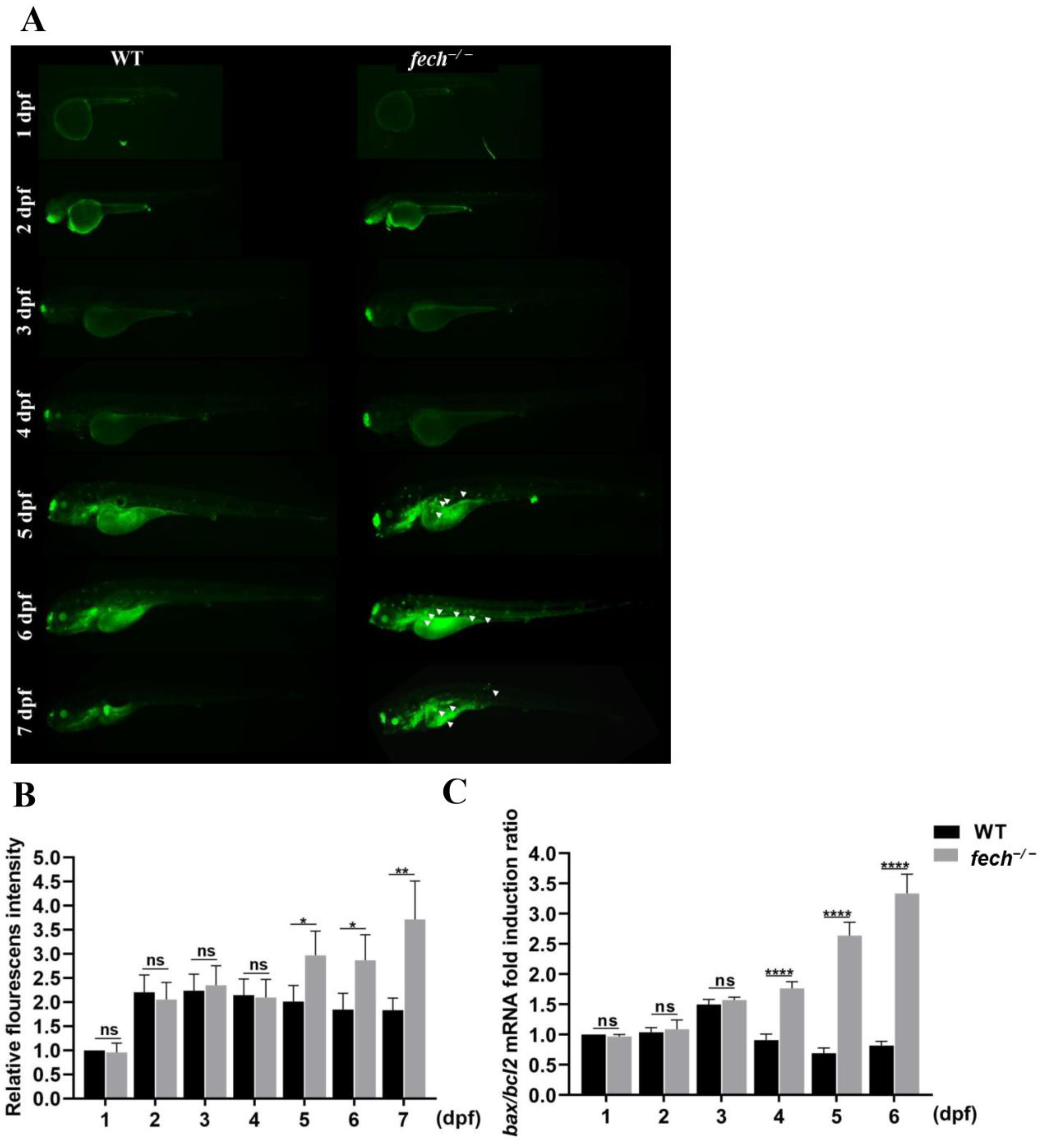
2.4. The Dynamics of PPIX Accumulation in fech−/− Larvae at Various Life Stages
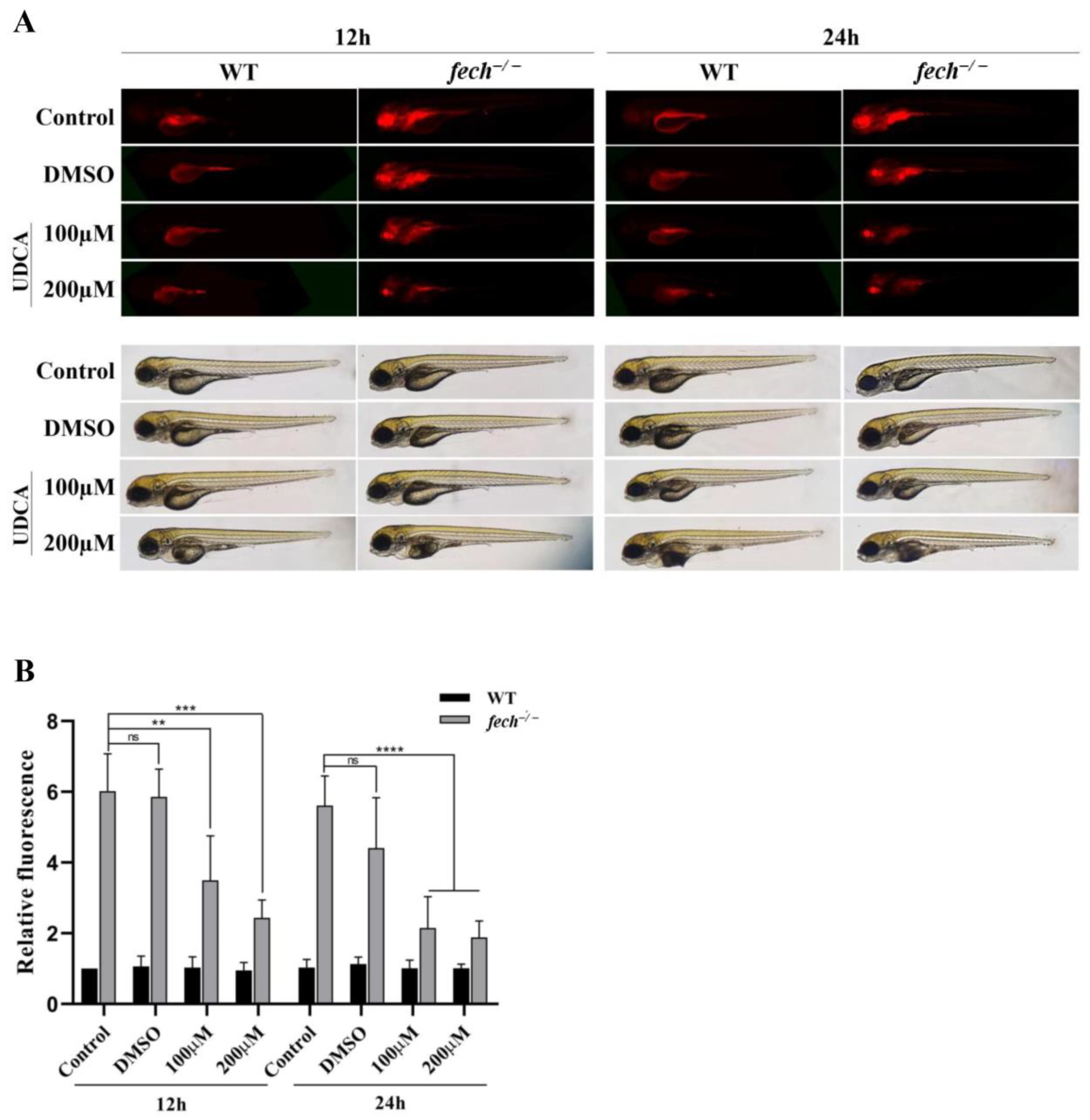
2.5. Attenuation of PPIX Accumulation in fech−/− Larvae following UDCA Treatment
2.6. Suppression of Apoptosis by UDCA Treatment of fech−/− Larvae
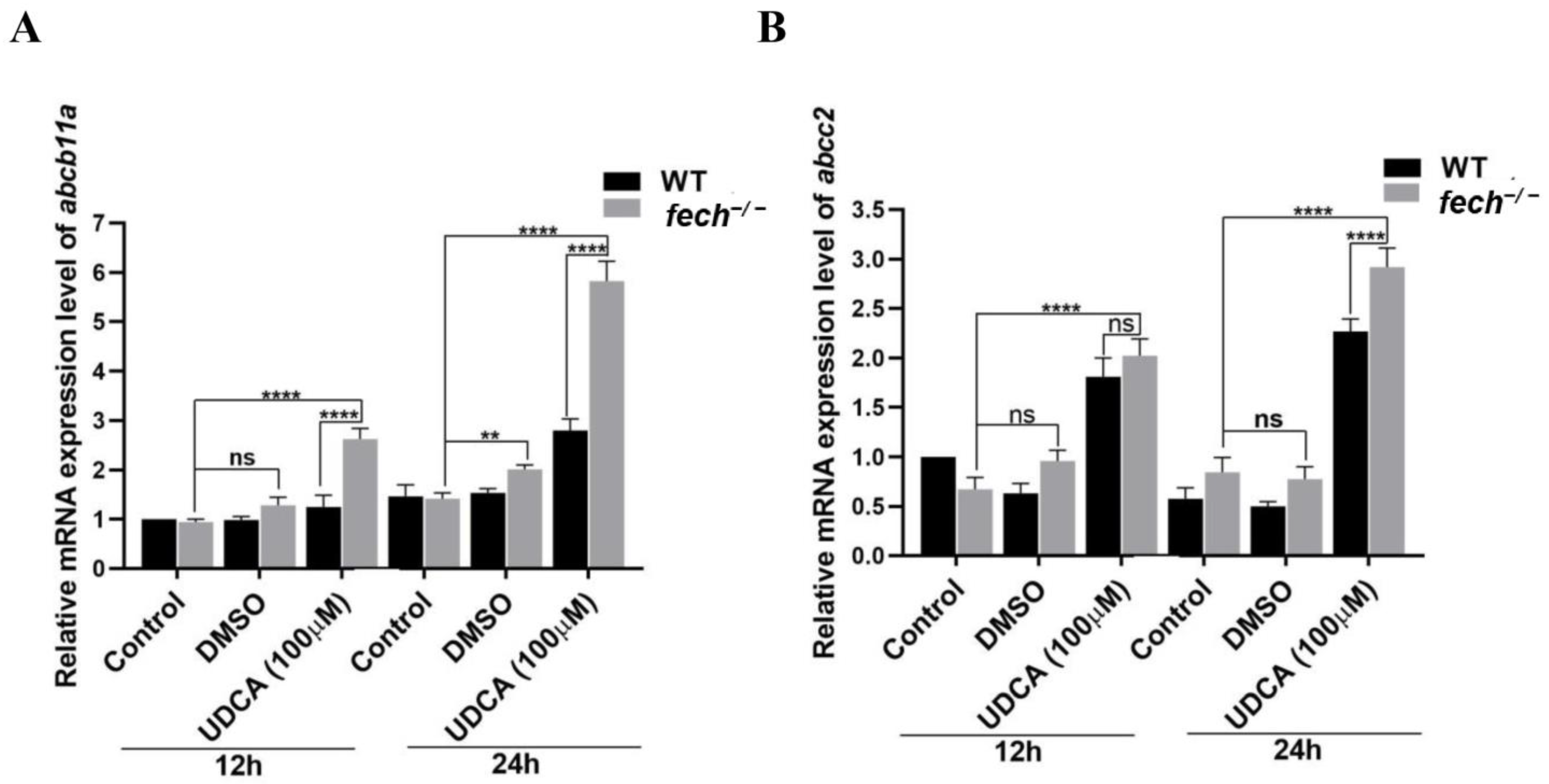
2.7. Activation of Bile Transportation-Related Genes by UDCA Treatment
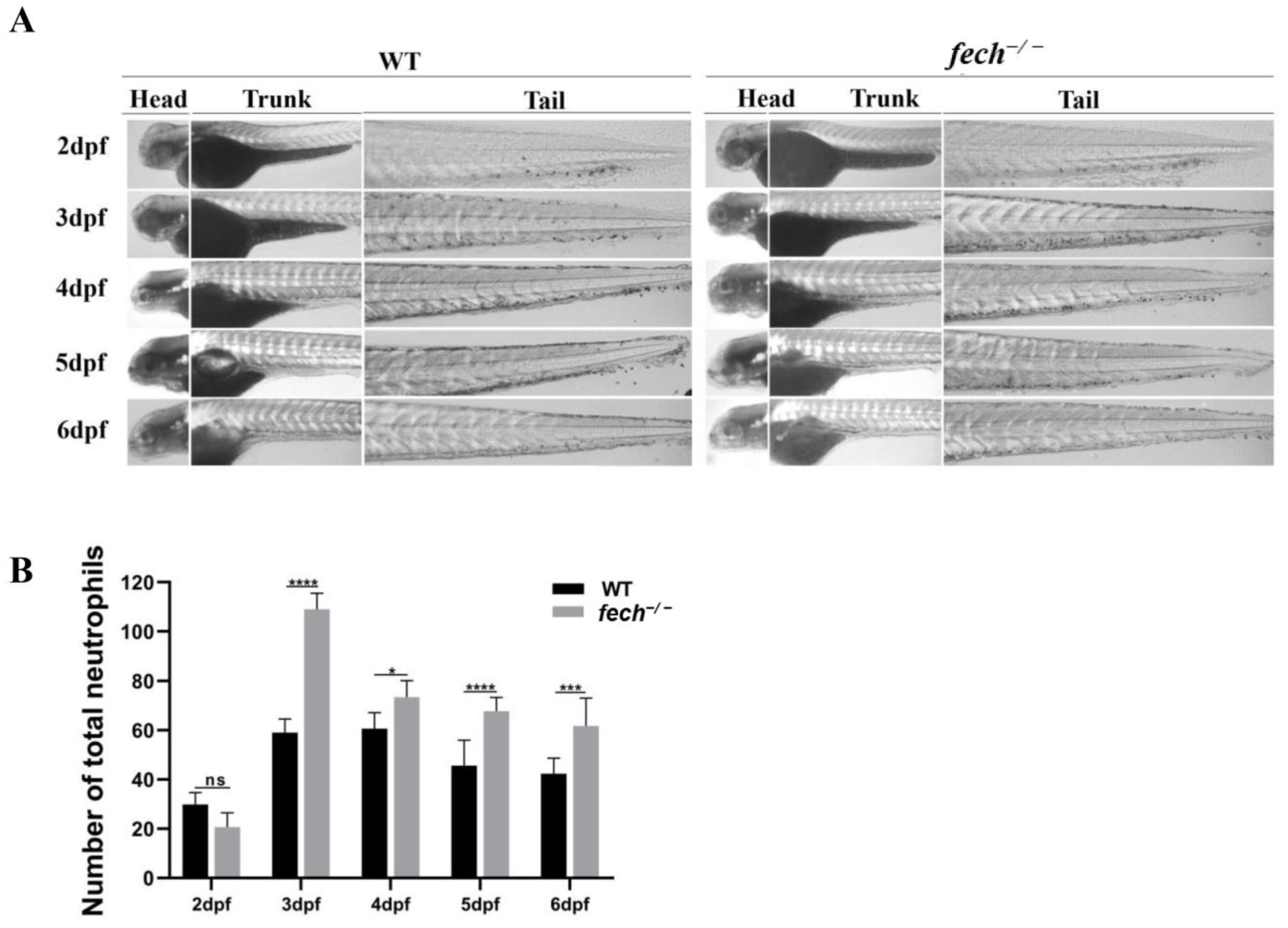
2.8. Temporal Changes in Neutrophil Production and Attenuation of Neutrophil Accumulation via UDCA Treatment in fech−/− Larvae
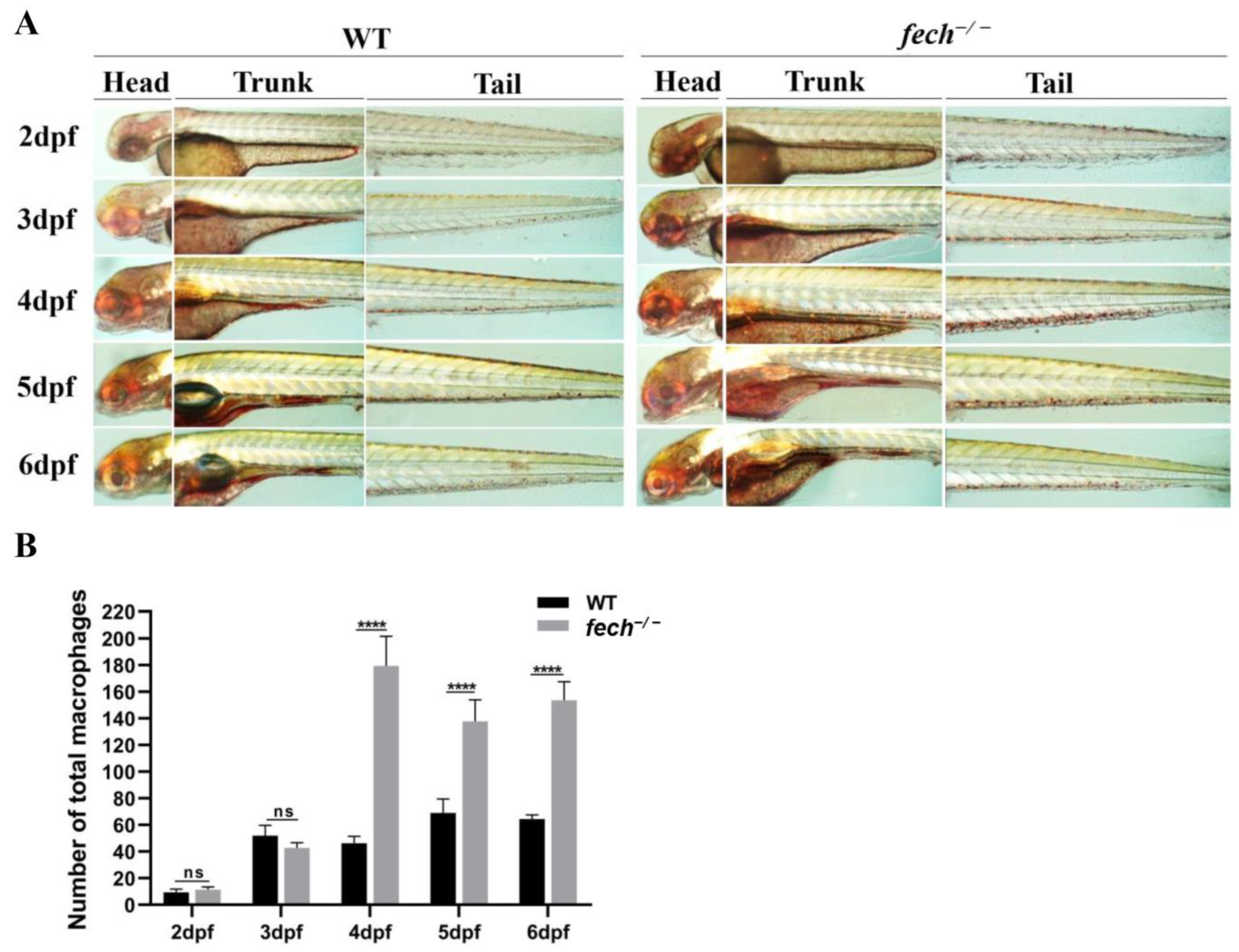
2.9. Temporal Changes in Macrophage Production and Attenuation of Macrophage Accumulation by UDCA Treatment in fech−/− Larvae
3. Discussion
4. Materials and Methods
4.1. Zebrafish Maintenance
4.2. Generation of Fech-Knockout Zebrafish Using CRISPR/Cas9 Technology
4.3. Assessing Phenotypic and Physiological Effects of Fech Deletion in Zebrafish Larvae
4.4. Tissue Collection for Fech Tissue Distribution Analysis
4.5. Total RNA Extraction and RT-qPCR
4.6. Whole-Mount In Situ Hybridization
4.7. Visualization of PPIX Accumulation in fech−/− Zebrafish Embryos and Larvae
4.8. Analyzing the Effect of UDCA on PPIX Accumulation in fech−/− Larvae
4.9. Analyzing the Impact of Fech Deletion in Zebrafish on Apoptosis Using Acridine Orange Staining
4.10. Analyzing the Effect of UDCA on Apoptosis Resulting from PPIX in fech−/− Larvae Using Acridine Orange Staining
4.11. Analyzing the Impact of UDCA Treatment on the Expression of Bile Acid Transporter-Related and Apoptosis-Related Genes
4.12. Analyzing the Effect of Fech Deletion on Neutrophil and Macrophage Production
4.13. Analyzing the Impact of UDCA Treatment on Neutrophil and Macrophage Production in fech−/− Larvae
4.14. Statistical Analysis
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Ardalan, Z.S.; Chandran, S.; Vasudevan, A.; Angus, P.W.; Grigg, A.; He, S.; Macdonald, G.A.; Strasser, S.I.; Tate, C.J.; Kennedy, G.A.; et al. Management of Patients with Erythropoietic Protoporphyria–Related Progressive Liver Disease. Liver Transplant. 2019, 25, 1620–1633. [Google Scholar] [CrossRef]
- Scholnick, P.; Marver, H.S.; Schmid, R. Erythropoietic protoporphyria: Evidence for multiple sites of excess protoporphyrin formation. J. Clin. Investig. 1971, 50, 203–207. [Google Scholar] [CrossRef] [PubMed]
- Goldstein, B.D.; Harber, L.C. Erythropoietic Protoporphyria: Lipid Peroxidation and Red Cell Membrane Damage Associated with Photohemolysis. J. Clin. Investig. 1972, 51, 892–902. [Google Scholar] [CrossRef]
- Wang, P.; Sachar, M.; Guo, G.L.; Shehu, A.I.; Lu, J.; Zhong, X.-B.; Ma, X. Liver metabolomics in a mouse model of erythropoietic protoporphyria. Biochem. Pharmacol. 2018, 154, 474–481. [Google Scholar] [CrossRef] [PubMed]
- Ferreira, G.C.; Franco, R.; Lloyd, S.G.; Moura, I.; Moura, J.J.G.; Huynh, B.H. Structure and function of ferrochelatase. J. Bioenerg. Biomembr. 1995, 27, 221–229. [Google Scholar] [CrossRef] [PubMed]
- Bloomer, J.; Bruzzone, C.; Zhu, L.; Scarlett, Y.; Magness, S.; Brenner, D. Molecular defects in ferrochelatase in patients with protoporphyria requiring liver transplantation. J. Clin. Investig. 1998, 102, 107–114. [Google Scholar] [CrossRef]
- Gouya, L.; Puy, H.; Robreau, A.-M.; Bourgeois, M.; Lamoril, J.; Da Silva, V.; Grandchamp, B.; Deybach, J.-C. The penetrance of dominant erythropoietic protoporphyria is modulated by expression of wildtype FECH. Nat. Genet. 2002, 30, 27–28. [Google Scholar] [CrossRef]
- Ferreira, G.C. Ferrochelatase. Int. J. Biochem. Cell Biol. 1999, 31, 995–1000. [Google Scholar] [CrossRef]
- Taketani, S.; Inazawa, J.; Nakahashi, Y.; Abe, T.; Tokunaga, R. Tokunaga, Structure of the human ferrochelatase gene. Exon/intron gene organization and location of the gene to chromosome 18. Eur. J. Biochem. 1992, 205, 217–222. [Google Scholar] [CrossRef] [PubMed]
- Amaral, J.D.; Viana, R.J.; Ramalho, R.M.; Steer, C.J.; Rodrigues, C.M. Bile acids: Regulation of apoptosis by ursodeoxycholic acid. J. Lipid Res. 2009, 50, 1721–1734. [Google Scholar] [CrossRef]
- Roma, M.G.; Toledo, F.D.; Boaglio, A.C.; Basiglio, C.L.; Crocenzi, F.A.; Pozzi, E.J.S. Ursodeoxycholic acid in cholestasis: Linking action mechanisms to therapeutic applications. Clin. Sci. 2011, 121, 523–544. [Google Scholar] [CrossRef] [PubMed]
- Piffaretti, D.; Burgio, F.; Thelen, M.; Kaelin-Lang, A.; Paganetti, P.; Reinert, M.; D’Angelo, M.L. Protoporphyrin IX tracer fluorescence modulation for improved brain tumor cell lines visualization. J. Photochem. Photobiol. B Biol. 2019, 201, 111640. [Google Scholar] [CrossRef] [PubMed]
- Groß, U.; Frank, M.; Doss, M.O. Hepatic complications of erythropoietic protoporphyria. Photodermatol. Photoimmunol. Photomed. 1998, 14, 52–57. [Google Scholar] [CrossRef]
- Poli, A.; Frieri, C.; Lefebvre, T.; Delforge, J.; Mirmiran, A.; Talbi, N.; Moulouel, B.; Six, M.; Paradis, V.; Parquet, N.; et al. Management of erythropoietic protoporphyria with cholestatic liver disease: A case report. Mol. Genet. Metab. Rep. 2023, 37, 101018. [Google Scholar] [CrossRef] [PubMed]
- Hempfling, W.; Dilger, K.; Beuers, U. Ursodeoxycholic acid—Adverse effects and drug interactions. Aliment. Pharmacol. Ther. 2003, 18, 963–972. [Google Scholar] [CrossRef] [PubMed]
- Streisinger, G.; Walker, C.; Dower, N.; Knauber, D.; Singer, F. Production of clones of homozygous diploid zebra fish (Brachydanio rerio). Nature 1981, 291, 293–296. [Google Scholar] [CrossRef] [PubMed]
- Grunwald, D.J.; Streisinger, G. Induction of recessive lethal and specific locus mutations in the zebrafish with ethyl nitrosourea. Genet. Res. 1992, 59, 103–116. [Google Scholar] [CrossRef]
- Bedell, V.M.; Wang, Y.; Campbell, J.M.; Poshusta, T.L.; Starker, C.G.; Krug, R.G., 2nd; Tan, W.; Penheiter, S.G.; Ma, A.C.; Leung, A.Y.H.; et al. In vivo genome editing using a high-efficiency TALEN system. Nature 2012, 491, 114–118. [Google Scholar] [CrossRef] [PubMed]
- Doyon, Y.; McCammon, J.M.; Miller, J.C.; Faraji, F.; Ngo, C.; Katibah, G.E.; Amora, R.; Hocking, T.D.; Zhang, L.; Rebar, E.J.; et al. Heritable targeted gene disruption in zebrafish using designed zinc-finger nucleases. Nat. Biotechnol. 2008, 26, 702–708. [Google Scholar] [CrossRef] [PubMed]
- Hsu, P.D.; Lander, E.S.; Zhang, F. Development and Applications of CRISPR-Cas9 for Genome Engineering. Cell 2014, 157, 1262–1278. [Google Scholar] [CrossRef]
- Sung, Y.H.; Kim, J.M.; Kim, H.-T.; Lee, J.; Jeon, J.; Jin, Y.; Choi, J.-H.; Ban, Y.H.; Ha, S.-J.; Kim, C.-H.; et al. Highly efficient gene knockout in mice and zebrafish with RNA-guided endonucleases. Genome Res. 2014, 24, 125–131. [Google Scholar] [CrossRef]
- Parng, C.; Seng, W.L.; Semino, C.; McGrath, P. Zebrafish: A Preclinical Model for Drug Screening. ASSAY Drug Dev. Technol. 2002, 1, 41–48. [Google Scholar] [CrossRef] [PubMed]
- Dooley, K.A.; Fraenkel, P.G.; Langer, N.B.; Schmid, B.; Davidson, A.J.; Weber, G.; Chiang, K.; Foott, H.; Dwyer, C.; Wingert, R.A.; et al. montalcino, A zebrafish model for variegate porphyria. Exp. Hematol. 2008, 36, 1132–1142. [Google Scholar] [CrossRef] [PubMed]
- Wang, H.; Long, Q.; Marty, S.D.; Sassa, S.; Lin, S. A zebrafish model for hepatoerythropoietic porphyria. Nat. Genet. 1998, 20, 239–243. [Google Scholar] [CrossRef] [PubMed]
- Childs, S.; Weinstein, B.M.; Mohideen, M.-A.P.; Donohue, S.; Bonkovsky, H.; Fishman, M.C. Zebrafish dracula encodes ferrochelatase and its mutation provides a model for erythropoietic protoporphyria. Curr. Biol. 2000, 10, 1001–1004. [Google Scholar] [CrossRef] [PubMed]
- Kirsch, R.E.; Meissner, P.N.; Hift, R.J. Variegate Porphyria. Semin. Liver Dis. 1998, 18, 33–41. [Google Scholar] [CrossRef]
- Rudnick, S.; Phillips, J. Bonkovsky, Porphyrias consortium of the rare diseases clinical research network, hepatoerythropoietic porphyria. In GeneReviews; Adam, M.P., Feldman, J., Mirzaa, G.M., Pagon, R.A., Wallace, S.E., Bean, L.J., Gripp, K.W., Amemiya, A., Eds.; GeneReviews®, University of Washington, Seattle: Seattle, WA, USA, 1993. Available online: http://www.ncbi.nlm.nih.gov/books/NBK169003/ (accessed on 7 September 2024).
- Song, H.-D.; Sun, X.-J.; Deng, M.; Zhang, G.-W.; Zhou, Y.; Wu, X.-Y.; Sheng, Y.; Chen, Y.; Ruan, Z.; Jiang, C.-L.; et al. Hematopoietic gene expression profile in zebrafish kidney marrow. Proc. Natl. Acad. Sci. USA 2004, 101, 16240–16245. [Google Scholar] [CrossRef]
- Lu, H.; Floris, F.; Rensing, M.; Andersson-Engels, S. Fluorescence Spectroscopy Study of Protoporphyrin IX in Optical Tissue Simulating Liquid Phantoms. Materials 2020, 13, 2105. [Google Scholar] [CrossRef]
- Paumgartner, G.; Beuers, U. Mechanisms of action and therapeutic efficacy of ursodeoxycholic acid in cholestatic liver disease. Clin. Liver Dis. 2004, 8, 67–81. [Google Scholar] [CrossRef]
- Tang, J.; Yan, Z.; Feng, Q.; Yu, L.; Wang, H. The Roles of Neutrophils in the Pathogenesis of Liver Diseases. Front. Immunol. 2021, 12, 625472. [Google Scholar] [CrossRef]
- Thisse, B.; Thisse, C. Fast Release Clones: A High Throughput Expression Analysis. ZFIN Direct Data Submission. 2004. Available online: https://zfin.org/ZDB-PUB-040907-1#summary (accessed on 11 October 2023).
- Thisse, C.; Thisse, B. High-resolution in situ hybridization to whole-mount zebrafish embryos. Nat. Protoc. 2008, 3, 59–69. [Google Scholar] [CrossRef] [PubMed]
- Bertrand, J.Y.; Kim, A.D.; Violette, E.P.; Stachura, D.L.; Cisson, J.L.; Traver, D. Definitive hematopoiesis initiates through a committed erythromyeloid progenitor in the zebrafish embryo. Development 2007, 134, 4147–4156. [Google Scholar] [CrossRef] [PubMed]
- Rasighaemi, P.; Basheer, F.; Liongue, C.; Ward, A.C. Zebrafish as a model for leukemia and other hematopoietic disorders. J. Hematol. Oncol. 2015, 8, 29. [Google Scholar] [CrossRef] [PubMed]
- Dailey, H.A.; Meissner, P.N. Erythroid Heme Biosynthesis and Its Disorders. Cold Spring Harb. Perspect. Med. 2013, 3, a011676. [Google Scholar] [CrossRef]
- Ponka, P. Cell Biology of Heme. Am. J. Med. Sci. 1999, 318, 241–256. [Google Scholar] [CrossRef]
- Zhang, J.; Hamza, I. Zebrafish as a model system to delineate the role of heme and iron metabolism during erythropoiesis. Mol. Genet. Metab. 2018, 128, 204–212. [Google Scholar] [CrossRef]
- Carradice, D.; Lieschke, G.J. Zebrafish in hematology: Sushi or science? Blood 2008, 111, 3331–3342. [Google Scholar] [CrossRef]
- Pelster, B.; Bagatto, B. Respiration. In Fish Physiology; Elsevier: Amsterdam, The Netherlands, 2010; pp. 289–309. [Google Scholar] [CrossRef]
- Wu, X.-J.; Thomas, P.; Zhu, Y. Pgrmc1 Knockout Impairs Oocyte Maturation in Zebrafish. Front. Endocrinol. 2018, 9, 560. [Google Scholar] [CrossRef]
- Atella, G.C.; Gondim, K.C.; Machado, E.A.; Medeiros, M.N.; Silva-Neto, M.A.; Masuda, H. Oogenesis and egg development in triatomines: A biochemical approach. An. Acad. Bras. Cienc. 2005, 77, 405–430. [Google Scholar] [CrossRef]
- Mazur, A. Metabolism of the stimulated rat spleen. J. Clin. Investig. 1968, 47, 2230–2238. [Google Scholar] [CrossRef]
- Thapar, M.; Bonkovsky, H.L. The diagnosis and management of erythropoietic protoporphyria. Gastroenterol. Hepatol. 2008, 4, 561–566. [Google Scholar]
- Kalra, A.; Yetiskul, E.; Wehrle, C.J.; Tuma, F. Physiology, Liver. In StatPearls; StatPearls Publishing: Treasure Island, FL, USA, 2023. Available online: http://www.ncbi.nlm.nih.gov/books/NBK535438/ (accessed on 12 October 2023).
- Balwani, M. Erythropoietic Protoporphyria and X-linked Protoporphyria; National Organization for Rare Disorders: Danbury, Connecticut, 2022; Available online: https://rarediseases.org/rare-diseases/erythropoietic-protoporphyria/#complete-report (accessed on 13 October 2023).
- Farid, Y.; Bowman, N.S.; Lecat, P. Biochemistry, hemoglobin synthesis. In StatPearls; StatPearls Publishing: Treasure Island, FL, USA, 2023. Available online: http://www.ncbi.nlm.nih.gov/books/NBK536912/ (accessed on 13 October 2023).
- Belele, C.L.; English, M.A.; Chahal, J.; Burnetti, A.; Finckbeiner, S.M.; Gibney, G.; Kirby, M.; Sood, R.; Liu, P.P. Differential requirement for Gata1 DNA binding and transactivation between primitive and definitive stages of hematopoiesis in zebrafish. Blood 2009, 114, 5162–5172. [Google Scholar] [CrossRef] [PubMed]
- Brun, A.; Sandberg, S. Mechanisms of photosensitivity in porphyric patients with special emphasis on erythropoietic protoporphyria. J. Photochem. Photobiol. B Biol. 1991, 10, 285–302. [Google Scholar] [CrossRef] [PubMed]
- Sachar, M.; Anderson, K.E.; Ma, X. Protoporphyrin IX: The Good, the Bad, and the Ugly. J. Pharmacol. Exp. Ther. 2015, 356, 267–275. [Google Scholar] [CrossRef] [PubMed]
- Kotb, M.A. Molecular Mechanisms of Ursodeoxycholic Acid Toxicity & Side Effects: Ursodeoxycholic Acid Freezes Regeneration & Induces Hibernation Mode. Int. J. Mol. Sci. 2012, 13, 8882–8914. [Google Scholar] [CrossRef]
- Higuchi, A.; Wakai, E.; Tada, T.; Koiwa, J.; Adachi, Y.; Shiromizu, T.; Goto, H.; Tanaka, T.; Nishimura, Y. Generation of a Transgenic Zebrafish Line for In Vivo Assessment of Hepatic Apoptosis. Pharmaceuticals 2021, 14, 1117. [Google Scholar] [CrossRef]
- Chu, J.; Sadler, K.C. New school in liver development: Lessons from zebrafish. Hepatology 2009, 50, 1656–1663. [Google Scholar] [CrossRef]
- Shimizu, N.; Shiraishi, H.; Hanada, T. Zebrafish as a Useful Model System for Human Liver Disease. Cells 2023, 12, 2246. [Google Scholar] [CrossRef]
- Qiao, L.; Yacoub, A.; Studer, E.; Gupta, S.; Pei, X.Y.; Grant, S.; Hylemon, P.B.; Dent, P. Inhibition of the MAPK and PI3K pathways enhances UDCA-induced apoptosis in primary rodent hepatocytes. Hepatology 2002, 35, 779–789. [Google Scholar] [CrossRef]
- Solá, S.; Aranha, M.M.; Steer, C.J.; Rodrigues, C.M. Game and Players: Mitochondrial Apoptosis and the Therapeutic Potential of Ursodeoxycholic Acid. Curr. Issues Mol. Biol. 2007, 9, 123–138. [Google Scholar] [CrossRef]
- Liu, J.; Lu, H.; Lu, Y.-F.; Lei, X.; Cui, J.Y.; Ellis, E.; Strom, S.C.; Klaassen, C.D. Potency of Individual Bile Acids to Regulate Bile Acid Synthesis and Transport Genes in Primary Human Hepatocyte Cultures. Toxicol. Sci. 2014, 141, 538–546. [Google Scholar] [CrossRef]
- Puy, H.; Gouya, L.; Deybach, J.-C. Porphyrias. Lancet 2010, 375, 924–937. [Google Scholar] [CrossRef] [PubMed]
- Lyoumi, S.; Abitbol, M.; Rainteau, D.; Karim, Z.; Bernex, F.; Oustric, V.; Millot, S.; Lettéron, P.; Heming, N.; Guillmot, L.; et al. Protoporphyrin Retention in Hepatocytes and Kupffer Cells Prevents Sclerosing Cholangitis in Erythropoietic Protoporphyria Mouse Model. Gastroenterology 2011, 141, 1509–1519.e3. [Google Scholar] [CrossRef] [PubMed]
- Anstey, A.V.; Hift, R.J. Liver disease in erythropoietic protoporphyria: Insights and implications for management. Postgrad. Med. J. 2007, 83, 739–748. [Google Scholar] [CrossRef] [PubMed]
- Balwani, M.; Naik, H.; Anderson, K.E.; Bissell, D.M.; Bloomer, J.; Bonkovsky, H.L.; Phillips, J.D.; Overbey, J.R.; Wang, B.; Singal, A.K.; et al. Clinical, Biochemical, and Genetic Characterization of North American Patients with Erythropoietic Protoporphyria and X-linked Protoporphyria. JAMA Dermatol. 2017, 153, 789–796. [Google Scholar] [CrossRef] [PubMed]
- Han, H.; Desert, R.; Das, S.; Song, Z.; Athavale, D.; Ge, X.; Nieto, N. Danger signals in liver injury and restoration of homeostasis. J. Hepatol. 2020, 73, 933–951. [Google Scholar] [CrossRef] [PubMed]
- Pérez-Figueroa, E.; Álvarez-Carrasco, P.; Ortega, E.; Maldonado-Bernal, C. Neutrophils: Many Ways to Die. Front. Immunol. 2021, 12, 631821. [Google Scholar] [CrossRef]
- Tang, L.; Cai, N.; Zhou, Y.; Liu, Y.; Hu, J.; Li, Y.; Yi, S.; Song, W.; Kang, L.; He, H. Acute stress induces an inflammation dominated by innate immunity represented by neutrophils in mice. Front. Immunol. 2022, 13, 1014296. [Google Scholar] [CrossRef]
- Yang, S.; Zhao, M.; Jia, S. Macrophage: Key player in the pathogenesis of autoimmune diseases. Front. Immunol. 2023, 14, 1080310. [Google Scholar] [CrossRef]
- Fujiwara, N.; Kobayashi, K. Macrophages in Inflammation. Curr. Drug Target. Inflamm. Allergy 2005, 4, 281–286. [Google Scholar] [CrossRef]
- Chen, S.; Saeed, A.F.; Liu, Q.; Jiang, Q.; Xu, H.; Xiao, G.G.; Rao, L.; Duo, Y. Macrophages in immunoregulation and therapeutics. Signal Transduct. Target. Ther. 2023, 8, 207. [Google Scholar] [CrossRef] [PubMed]
- Miura, T.; Ouchida, R.; Yoshikawa, N.; Okamoto, K.; Makino, Y.; Nakamura, T.; Morimoto, C.; Makino, I.; Tanaka, H. Functional Modulation of the Glucocorticoid Receptor and Suppression of NF-κB-dependent Transcription by Ursodeoxycholic Acid. J. Biol. Chem. 2001, 276, 47371–47378. [Google Scholar] [CrossRef] [PubMed]
- Yoshikawa, M.; Tsujii, T.; Matsumura, K.; Yamao, J.; Matsumura, Y.; Kubo, R.; Fukui, H.; Ishizaka, S. Immunomodulatory effects of ursodeoxycholic acid on immune responses. Hepatology 1992, 16, 358–364. [Google Scholar] [CrossRef] [PubMed]
- Ishizaki, K.; Iwaki, T.; Kinoshita, S.; Koyama, M.; Fukunari, A.; Tanaka, H.; Tsurufuji, M.; Sakata, K.; Maeda, Y.; Imada, T.; et al. Ursodeoxycholic acid protects concanavalin A-induced mouse liver injury through inhibition of intrahepatic tumor necrosis factor-α and macrophage inflammatory protein-2 production. Eur. J. Pharmacol. 2008, 578, 57–64. [Google Scholar] [CrossRef] [PubMed]
- Avdesh, A.; Chen, M.; Martin-Iverson, M.T.; Mondal, A.; Ong, D.; Rainey-Smith, S.; Taddei, K.; Lardelli, M.; Groth, D.M.; Verdile, G.; et al. Regular Care and Maintenance of a Zebrafish (Danio rerio) Laboratory: An Introduction. JoVE J. Vis. Exp. 2012, 69, e4196. [Google Scholar] [CrossRef]
- Jao, L.-E.; Wente, S.R.; Chen, W. Efficient multiplex biallelic zebrafish genome editing using a CRISPR nuclease system. Proc. Natl. Acad. Sci. USA 2013, 110, 13904–13909. [Google Scholar] [CrossRef]
- Varshney, G.K.; Pei, W.; LaFave, M.C.; Idol, J.; Xu, L.; Gallardo, V.; Carrington, B.; Bishop, K.; Jones, M.; Li, M.; et al. High-throughput gene targeting and phenotyping in zebrafish using CRISPR/Cas9. Genome Res. 2015, 25, 1030–1042. [Google Scholar] [CrossRef]
- Kim, H.J.; Lee, H.J.; Kim, H.; Cho, S.W.; Kim, J.-S. Targeted genome editing in human cells with zinc finger nucleases constructed via modular assembly. Genome Res. 2009, 19, 1279–1288. [Google Scholar] [CrossRef]
- Livak, K.J.; Schmittgen, T.D. Analysis of relative gene expression data using real-time quantitative PCR and the 2−ΔΔCT Method. Methods 2001, 25, 402–408. [Google Scholar] [CrossRef]
- Kimmel, C.B.; Ballard, W.W.; Kimmel, S.R.; Ullmann, B.; Schilling, T.F. Stages of embryonic development of the zebrafish. Dev. Dyn. 1995, 203, 253–310. [Google Scholar] [CrossRef]
- Tucker, B.; Lardelli, M. A Rapid Apoptosis Assay Measuring Relative Acridine Orange Fluorescence in Zebrafish Embryos. Zebrafish 2007, 4, 113–116. [Google Scholar] [CrossRef] [PubMed]
- Rosowski, E.E.; Raffa, N.; Knox, B.P.; Golenberg, N.; Keller, N.P.; Huttenlocher, A. Macrophages inhibit Aspergillus fumigatus germination and neutrophil-mediated fungal killing. PLoS Pathog. 2018, 14, e1007229. [Google Scholar] [CrossRef] [PubMed]
- Herbomel, P.; Levraud, J.-P. Imaging early macrophage differentiation, migration, and behaviors in live zebrafish embryos. Methods Mol. Biol. 2005, 105, 199–214. [Google Scholar] [CrossRef]
- Hubbard, T. The Ensembl genome database project. Nucleic Acids Res. 2002, 30, 38–41. [Google Scholar]
- Benson, D.; Lipman, D.J.; Ostell, J. GenBank. Nucleic Acids Res. 1993, 21, 2963–2965. [Google Scholar]
- Sievers, F.; Higgins, D.G. The clustal omega multiple alignment package. In Multiple Sequence Alignment; Katoh, K., Ed.; Methods in Molecular Biology; Springer: New York, NY, USA, 2021; Volume 2231, pp. 3–16. Available online: http://link.springer.com/10.1007/978-1-0716-1036-7_1 (accessed on 5 October 2023).
- Rice, P.; Longden, I.; Bleasby, A. EMBOSS: The european molecular biology open software suite. Trends Genet. 2000, 16, 276–277. [Google Scholar]
- Paffett-Lugassy, N.N.; Zon, L.T. Analysis of hematopoietic development in the zebrafish. In Developmental Hematopoiesis; Humana Press: Totowa, NJ, USA, 2004; pp. 171–198. Available online: http://link.springer.com/10.1385/1-59259-826-9:171 (accessed on 7 October 2023).






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Application | Primer Sequence (5′ to 3′) | |
|---|---|---|
| sgRNA synthesis | T7-sgRNA (Forward) | GAAATTAATACGACTCACTATAGGAAGGAATGGTGAAACTGCgtttTagagctagaaatagcaagttAaaat |
| Universal reverse primer | gatccgcaccgactcggtgccactttttcaagtTgataaCggactagccttatttTaacttgctatttctag | |
| T7E1 assay | F | ATTGCCAAAAGACGCACCCCAAAGATC |
| R | CTCACCCGTGTCCGGACACATCTCAT | |
| Mutation confirmation (RT-qPCR) | F | GGTGAAACTGCTGGATGAGATGTGT |
| R | ACTGTGGGTACTGTGTGAAGGCC |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Wijerathna, H.M.S.M.; Shanaka, K.A.S.N.; Raguvaran, S.S.; Jayamali, B.P.M.V.; Kim, S.-H.; Kim, M.-J.; Jung, S.; Lee, J. CRISPR/Cas9-Mediated fech Knockout Zebrafish: Unraveling the Pathogenesis of Erythropoietic Protoporphyria and Facilitating Drug Screening. Int. J. Mol. Sci. 2024, 25, 10819. https://doi.org/10.3390/ijms251910819
Wijerathna HMSM, Shanaka KASN, Raguvaran SS, Jayamali BPMV, Kim S-H, Kim M-J, Jung S, Lee J. CRISPR/Cas9-Mediated fech Knockout Zebrafish: Unraveling the Pathogenesis of Erythropoietic Protoporphyria and Facilitating Drug Screening. International Journal of Molecular Sciences. 2024; 25(19):10819. https://doi.org/10.3390/ijms251910819
Chicago/Turabian StyleWijerathna, Hitihami M. S. M., Kateepe A. S. N. Shanaka, Sarithaa S. Raguvaran, Bulumulle P. M. V. Jayamali, Seok-Hyung Kim, Myoung-Jin Kim, Sumi Jung, and Jehee Lee. 2024. "CRISPR/Cas9-Mediated fech Knockout Zebrafish: Unraveling the Pathogenesis of Erythropoietic Protoporphyria and Facilitating Drug Screening" International Journal of Molecular Sciences 25, no. 19: 10819. https://doi.org/10.3390/ijms251910819
APA StyleWijerathna, H. M. S. M., Shanaka, K. A. S. N., Raguvaran, S. S., Jayamali, B. P. M. V., Kim, S.-H., Kim, M.-J., Jung, S., & Lee, J. (2024). CRISPR/Cas9-Mediated fech Knockout Zebrafish: Unraveling the Pathogenesis of Erythropoietic Protoporphyria and Facilitating Drug Screening. International Journal of Molecular Sciences, 25(19), 10819. https://doi.org/10.3390/ijms251910819