
The Effect of Prenatal and Neonatal Fluoride Exposure to Morphine-Induced Neuroinflammation

 , , , , , , , and
, , , , , , , and
Abstract
1. Introduction
2. Results
2.1. Gene Expression—rT-PCR Analysis
2.1.1. COX-1
2.1.2. COX-2
2.1.3. GFAP
2.1.4. Iba1
2.2. Protein Expression—Western Blot Analysis
2.2.1. COX-1
2.2.2. COX-2
2.2.3. GFAP Protein Expression
2.2.4. Iba1
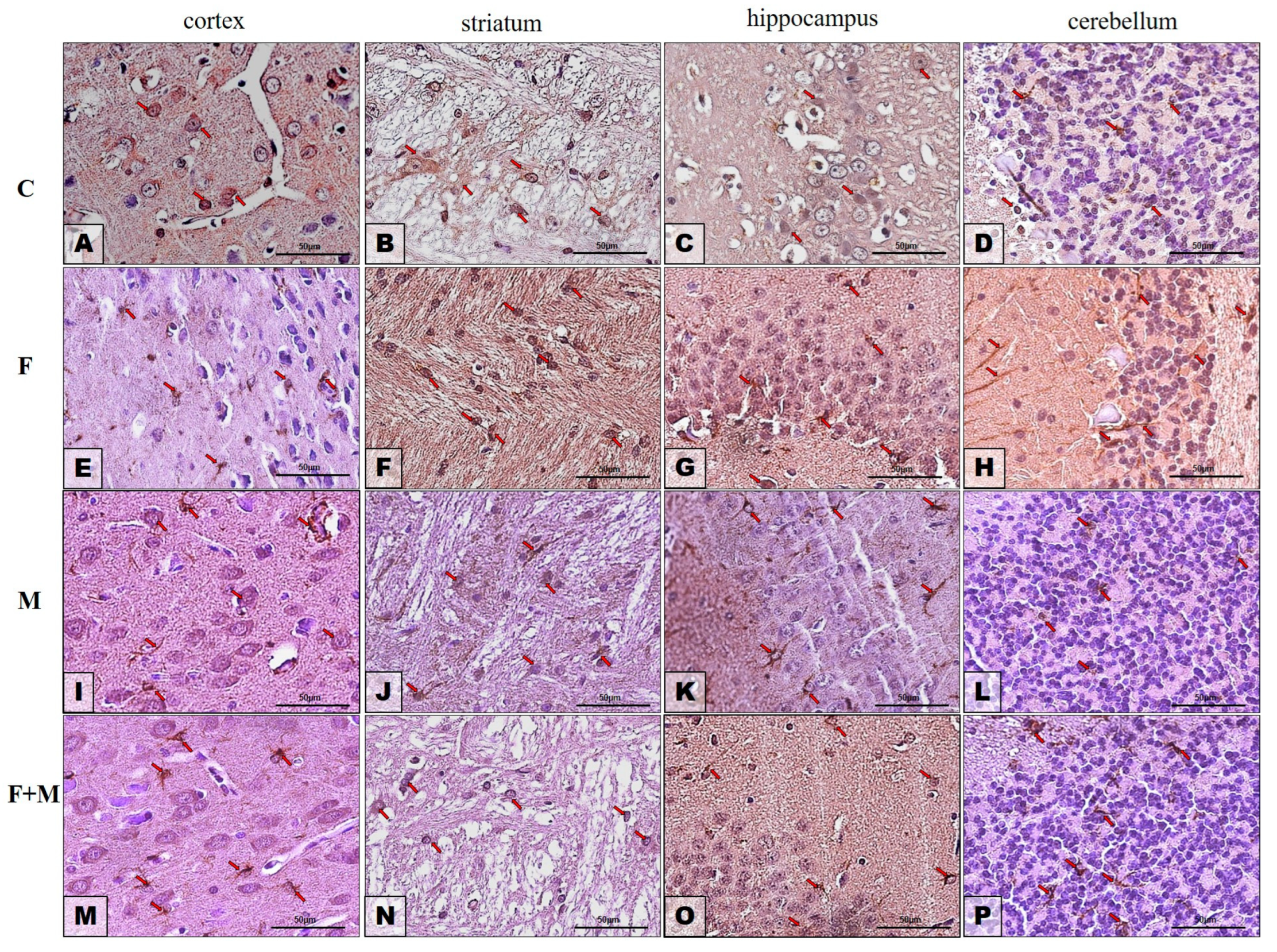
2.3. Protein Expression—Immunohistochemistry Analysis
2.3.1. GFAP
2.3.2. Iba1
3. Discussion
3.1. COX-1 Dependent Inflammatory Response in Morphine Dependence Is Influenced by Perinatal Fluoride Exposure
3.2. COX-2 Expression Changes May Affect Response to Morphine, thus Modulating Dependence and Tolerance to Morphine in Perinatal Fluoride Exposure
3.3. GFAP Increased Expression Indicates Active Astrogliosis in Pre- and Neonatal Fluoride Exposure
3.4. Morphine Treatment Causes Structure-Dependent Iba1 Expression Changes That Are Influenced by Fluoride Pre-Exposure
4. Materials and Methods
4.1. Animal Model
4.2. Gene Expression Analysis
- − GAPDH forward: ATGACTCTACCCACGGCAAG, reverse: CTGGAAGATGGT GATGGGTT
- − Iba1 forward: GATTTGCAGGGAGGAAAAGCT, reverse: AACCCCAAGTTTCTCCAGCAT
- − GFAP forward: GGTGGAGAGGGACAATCTCA, reverse: CCAGCTGCTCCTGGAGTTCT
- − COX-1 forward: GTTCACAGGAGAGAAGGAGATG, reverse: GGAGCCCCCATCTCTATCATGC
- − COX-2 forward: AATGAGTACCGCAAACGCTTCT, reverse: AGCCATTTCTTTCTCTCCTGTAAG.
4.3. Western Blot Analysis
4.4. Immunohistochemical Analysis
4.5. Statistical Analysis
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Kleber, H.D. Pharmacologic treatments for opioid dependence: Detoxification and maintenance options. Dialogues Clin. Neurosci. 2007, 9, 455–470. [Google Scholar] [CrossRef] [PubMed]
- Sharfstein, J.M.; Olsen, Y. Making Amends for the Opioid Epidemic. JAMA—J. Am. Med. Assoc. 2019, 321, 1446–1447. [Google Scholar] [CrossRef] [PubMed]
- Listos, J.; Łupina, M.; Talarek, S.; Mazur, A.; Orzelska-Górka, J.; Kotlińska, J. The Mechanisms Involved in Morphine Addiction: An Overview. Int. J. Mol. Sci. 2019, 20, 4302. [Google Scholar] [CrossRef] [PubMed]
- Marcellino, D.; Ferré, S.; Casadó, V.; Cortés, A.; Le Foll, B.; Mazzola, C.; Drago, F.; Saur, O.; Stark, H.; Soriano, A.; et al. Identification of dopamine D1–D3 receptor heteromers: Indications for a role of synergistic D1–D3 receptor interactions in the striatum. J. Biol. Chem. 2008, 283, 26016–26025. [Google Scholar] [CrossRef] [PubMed]
- Iranpour, M.; Torkzadeh-Tabrizi, S.; Khatoon-Asadi, Z.; Malekpour-Afshar, R. Immunohistochemical Assessment of Inflammation and Regeneration in Morphine-Dependent Rat Brain. Addict. Health 2018, 10, 156. [Google Scholar] [CrossRef] [PubMed]
- Namba, M.D.; Leyrer-Jackson, J.M.; Nagy, E.K.; Olive, M.F.; Neisewander, J.L. Neuroimmune Mechanisms as Novel Treatment Targets for Substance Use Disorders and Associated Comorbidities. Front. Neurosci. 2021, 15, 650785. [Google Scholar] [CrossRef]
- Dinda, A.; Gitman, M.; Singhal, P.C. Immunomodulatory effect of morphine: Therapeutic implications. Expert Opin. Drug Saf. 2005, 4, 669–675. [Google Scholar] [CrossRef]
- Feehan, A.K.; Zadina, J.E. Morphine immunomodulation prolongs inflammatory and postoperative pain while the novel analgesic ZH853 accelerates recovery and protects against latent sensitization. J. Neuroinflammation 2019, 16, 100. [Google Scholar] [CrossRef]
- Kohno, M.; Link, J.; Dennis, L.E.; McCready, H.; Huckans, M.; Hoffman, W.F.; Loftis, J.M. Neuroinflammation in addiction: A review of neuroimaging studies and potential immunotherapies. Pharmacol. Biochem. Behav. 2019, 179, 34–42. [Google Scholar] [CrossRef]
- Lacagnina, M.J.; Rivera, P.D.; Bilbo, S.D. Glial and Neuroimmune Mechanisms as Critical Modulators of Drug Use and Abuse. Neuropsychopharmacology 2016, 42, 156–177. [Google Scholar] [CrossRef]
- Hutchinson, M.R.; Bland, S.T.; Johnson, K.W.; Rice, K.C.; Maier, S.F.; Watkins, L.R. Opioid-induced glial activation: Mechanisms of activation and implications for opioid analgesia, dependence, and reward. Sci. World J. 2007, 7, 98–111. [Google Scholar] [CrossRef] [PubMed]
- Wang, X.; Loram, L.C.; Ramos, K.; De Jesus, A.J.; Thomas, J.; Cheng, K.; Reddy, A.; Somogyi, A.A.; Hutchinson, M.R.; Watkins, L.R.; et al. Morphine activates neuroinflammation in a manner parallel to endotoxin. Proc. Natl. Acad. Sci. USA 2012, 109, 6325–6330. [Google Scholar] [CrossRef] [PubMed]
- Eidson, L.N.; Murphy, A.Z. Inflammatory mediators of opioid tolerance: Implications for dependency and addiction. Peptides 2019, 115, 51–58. [Google Scholar] [CrossRef] [PubMed]
- Watkins, L.R.; Hutchinson, M.R.; Johnston, I.N.; Maier, S.F. Glia: Novel counter-regulators of opioid analgesia. Trends Neurosci. 2005, 28, 661–669. [Google Scholar] [CrossRef] [PubMed]
- Mika, J.; Wawrzczak-Bargiela, A.; Osikowicz, M.; Makuch, W.; Przewlocka, B. Attenuation of morphine tolerance by minocycline and pentoxifylline in naive and neuropathic mice. Brain Behav. Immun. 2009, 23, 75–84. [Google Scholar] [CrossRef]
- Davis, M. Modulation of microglia can attenuate neuropathic pain symptoms and enhance morphine effectiveness. Pharmacol. Rep. 2008, 60, 175–176. [Google Scholar] [CrossRef]
- Guth, S.; Hüser, S.; Roth, A.; Degen, G.; Diel, P.; Edlund, K.; Eisenbrand, G.; Engel, K.H.; Epe, B.; Grune, T.; et al. Toxicity of fluoride: Critical evaluation of evidence for human developmental neurotoxicity in epidemiological studies, animal experiments and in vitro analyses. Arch. Toxicol. 2020, 94, 1375–1415. [Google Scholar] [CrossRef]
- Iamandii, I.; De Pasquale, L.; Giannone, M.E.; Veneri, F.; Generali, L.; Consolo, U.; Birnbaum, L.S.; Castenmiller, J.; Halldorsson, T.I.; Filippini, T.; et al. Does fluoride exposure affect thyroid function? A systematic review and dose-response meta-analysis. Environ. Res. 2023, 242, 117759. [Google Scholar] [CrossRef]
- Dec, K.; Łukomska, A.; Baranowska-Bosiacka, I.; Pilutin, A.; Maciejewska, D.; Skonieczna-Żydecka, K.; Derkacz, R.; Goschorska, M.; Wąsik, A.; Rębacz-Maron, E.; et al. Pre-and postnatal exposition to fluorides induce changes in rats liver morphology by impairment of antioxidant defense mechanisms and COX induction. Chemosphere 2018, 211, 112–119. [Google Scholar] [CrossRef]
- European Commission Directive 2003/40/EC of 16 May 2003 Establishing the List, Concentration Limits and Labelling Requirements for the Constituents of Natural Mineral Waters and the Conditions for Using Ozone-Enriched Air for the Treatment of Natural Mineral Waters and Spring Waters. Available online: https://eur-lex.europa.eu/LexUriServ/LexUriServ.do?uri=OJ:L:2003:126:0034:0039:EN:PDF (accessed on 31 December 2023).
- Aoun, A.; Darwiche, F.; Al Hayek, S.; Doumit, J. The Fluoride Debate: The Pros and Cons of Fluoridation. Prev. Nutr. Food Sci. 2018, 23, 171–180. [Google Scholar] [CrossRef]
- Viswanathan, G.; Gopalakrishnan, S.; Siva Ilango, S. Assessment of water contribution on total fluoride intake of various age groups of people in fluoride endemic and non-endemic areas of Dindigul District, Tamil Nadu, South India. Water Res. 2010, 44, 6186–6200. [Google Scholar] [CrossRef] [PubMed]
- Viswanathan, G.; Jaswanth, A.; Gopalakrishnan, S.; Ilango, S.S. Mapping of fluoride endemic areas and assessment of fluoride exposure. Sci. Total Environ. 2009, 407, 1579–1587. [Google Scholar] [CrossRef] [PubMed]
- Gopalakrishnan, S.B.; Viswanathan, G.; Ilango, S.S. Prevalence of fluorosis and identification of fluoride endemic areas in Manur block of Tirunelveli District, Tamil Nadu, South India. Appl. Water Sci. 2012, 2, 235–243. [Google Scholar] [CrossRef]
- Anasuya, A.; Bapurao, S.; Paranjape, P.K. Fluoride and silicon intake in normal and endemic fluorotic areas. J. Trace Elem. Med. Biol. Organ Soc. Miner. Trace Elem. GMS 1996, 10, 149–155. [Google Scholar] [CrossRef] [PubMed]
- Dec, K.; Łukomska, A.; Maciejewska, D.; Jakubczyk, K.; Baranowska-Bosiacka, I.; Chlubek, D.; Wąsik, A.; Gutowska, I. The Influence of Fluorine on the Disturbances of Homeostasis in the Central Nervous System. Biol. Trace Elem. Res. 2017, 177, 224–234. [Google Scholar] [CrossRef] [PubMed]
- Grandjean, P. Developmental fluoride neurotoxicity: An updated review. Environ. Health Glob. Access Sci. Source 2019, 18, 110. [Google Scholar] [CrossRef] [PubMed]
- Kaindl, A.M.; Degos, V.; Peineau, S.; Gouadon, E.; Chhor, V.; Loron, G.; Le Charpentier, T.; Josserand, J.; Ali, C.; Vivien, D.; et al. Activation of microglial N-methyl-D-aspartate receptors triggers inflammation and neuronal cell death in the developing and mature brain. Ann. Neurol. 2012, 72, 536–549. [Google Scholar] [CrossRef]
- Dec, K.; Łukomska, A.; Skonieczna-Żydecka, K.; Kolasa-Wołosiuk, A.; Tarnowski, M.; Baranowska-Bosiacka, I.; Gutowska, I. Long-term exposure to fluoride as a factor promoting changes in the expression and activity of cyclooxygenases (COX1 and COX2) in various rat brain structures. Neurotoxicology 2019, 74, 81–90. [Google Scholar] [CrossRef]
- Shivarajashankara, Y.M.; Shivashankara, A.R.; Gopalakrishna Bhat, P.; Muddanna Rao, S.; Hanumanth Rao, S. Histological changes in the brain of young fluoride-intoxicated rats. Fluoride 2002, 35, 12–21. [Google Scholar]
- Souza-Monteiro, D.; Ferreira, M.K.M.; Bittencourt, L.O.; Aragão, W.A.B.; de Oliveira, I.G.; Maia, C.S.F.; Freire, M.A.M.; Zohoori, F.V.; Buzalaf, M.A.R.; Lima, R.R. Intrauterine and Postnatal Exposure to High Levels of Fluoride Is Associated with Motor Impairments, Oxidative Stress, and Morphological Damage in the Cerebellum of Offspring Rats. Int. J. Mol. Sci. 2022, 23, 8556. [Google Scholar] [CrossRef]
- Pannunzio, A.; Coluccia, M. Cyclooxygenase-1 (COX-1) and COX-1 Inhibitors in Cancer: A Review of Oncology and Medicinal Chemistry Literature. Pharmaceuticals 2018, 11, 101. [Google Scholar] [CrossRef] [PubMed]
- López, D.E.; Ballaz, S.J. The Role of Brain Cyclooxygenase-2 (Cox-2) Beyond Neuroinflammation: Neuronal Homeostasis in Memory and Anxiety. Mol. Neurobiol. 2020, 57, 5167–5176. [Google Scholar] [CrossRef] [PubMed]
- Mandwie, M.; Piper, J.; Gorrie, C.; Keay, K.; Musumeci, G.; Al-Badri, G.; Castorina, A. Rapid GFAP and Iba1 expression changes in the female rat brain following spinal cord injury. Neural Regen. Res. 2022, 17, 378. [Google Scholar] [CrossRef] [PubMed]
- Fukui, M.; Tanaka, M.; Toda, H.; Asano, M.; Yamazaki, M.; Hasegawa, G.; Imai, S.; Fujinami, A.; Ohta, M.; Nakamura, N. The serum concentration of allograft inflammatory factor-1 is correlated with metabolic parameters in healthy subjects. Metabolism 2012, 61, 1021–1025. [Google Scholar] [CrossRef]
- Silver, J.; Miller, J.H. Regeneration beyond the glial scar. Nat. Rev. Neurosci. 2004, 5, 146–156. [Google Scholar] [CrossRef]
- Baranowska-Bosiacka, I.; Listos, J.; Gutowska, I.; Machoy-Mokrzyńska, A.; Kolasa-Wołosiuk, A.; Tarnowski, M.; Puchałowicz, K.; Prokopowicz, A.; Talarek, S.; Listos, P.; et al. Effects of perinatal exposure to lead (Pb) on purine receptor expression in the brain and gliosis in rats tolerant to morphine analgesia. Toxicology 2016, 339, 19–33. [Google Scholar] [CrossRef]
- Listos, J.; Baranowska-Bosiacka, I.; Wąsik, A.; Talarek, S.; Tarnowski, M.; Listos, P.; Łupina, M.; Antkiewicz-Michaluk, L.; Gutowska, I.; Tkacz, M.; et al. The adenosinergic system is involved in sensitization to morphine withdrawal signs in rats-neurochemical and molecular basis in dopaminergic system. Pharmaceuticals 2016, 233, 2383–2397. [Google Scholar] [CrossRef]
- Listos, J.; Baranowska-Bosiacka, I.; Talarek, S.; Listos, P.; Orzelska, J.; Fidecka, S.; Gutowska, I.; Kolasa, A.; Rybicka, M.; Chlubek, D. The effect of perinatal lead exposure on dopamine receptor D2 expression in morphine dependent rats. Toxicology 2013, 310, 73–83. [Google Scholar] [CrossRef]
- Kobayashi, C.A.N.; de L. Leite, A.; da Silva, T.L.; dos Santos, L.D.; Nogueira, F.C.S.; Santos, K.S.; de Oliveira, R.C.; Palma, M.S.; Domont, G.B.; Buzalaf, M.A.R. Proteomic analysis of urine in rats chronically exposed to fluoride. J. Biochem. Mol. Toxicol. 2011, 25, 8–14. [Google Scholar] [CrossRef]
- Morales-González, J.A.; Gutiérrez-Salinas, J.; García-Ortiz, L.; del C. Chima-Galán, M.; Madrigal-Santillán, E.; Esquivel-Soto, J.; Esquivel-Chirino, C.; González-Rubio, M.G.L.Y. Effect of sodium fluoride ingestion on malondialdehyde concentration and the activity of antioxidant enzymes in rat erythrocytes. Int. J. Mol. Sci. 2010, 11, 2443–2452. [Google Scholar] [CrossRef]
- Łukomska, A.; Baranowska-Bosiacka, I.; Dec, K.; Pilutin, A.; Tarnowski, M.; Jakubczyk, K.; Żwierełło, W.; Skórka-Majewicz, M.; Chlubek, D.; Gutowska, I. Changes in Gene and Protein Expression of Metalloproteinase-2 and -9 and Their Inhibitors TIMP2 and TIMP3 in Different Parts of Fluoride-Exposed Rat Brain. Int. J. Mol. Sci. 2020, 22, 391. [Google Scholar] [CrossRef] [PubMed]
- Zhang, Y.; Zhou, B.H.; Tan, P.P.; Chen, Y.; Miao, C.Y.; Wang, H.W. Key Role of Pro-inflammatory Cytokines in the Toxic Effect of Fluoride on Hepa1-6 Cells. Biol. Trace Elem. Res. 2020, 197, 115–122. [Google Scholar] [CrossRef] [PubMed]
- Ishijima, T.; Nakajima, K. Inflammatory cytokines TNFα, IL-1β, and IL-6 are induced in endotoxin- stimulated microglia through different signaling cascades. Sci. Prog. 2021, 104. [Google Scholar] [CrossRef]
- Kupnicka, P.; Listos, J.; Tarnowski, M.; Kolasa-Wołosiuk, A.; Wąsik, A.; Łukomska, A.; Barczak, K.; Gutowska, I.; Chlubek, D.; Baranowska-Bosiacka, I. Fluoride Affects Dopamine Metabolism and Causes Changes in the Expression of Dopamine Receptors (D1R and D2R) in Chosen Brain Structures of Morphine-Dependent Rats. Int. J. Mol. Sci. 2020, 21, 2361. [Google Scholar] [CrossRef] [PubMed]
- Pereira, M.; Dombrowski, P.A.; Losso, E.M.; Chioca, L.R.; Da Cunha, C.; Andreatini, R. Memory impairment induced by sodium fluoride is associated with changes in brain monoamine levels. Neurotox. Res. 2011, 19, 55–62. [Google Scholar] [CrossRef]
- Ghazanfari, N.; van Waarde, A.; Dierckx, R.A.J.O.; Doorduin, J.; de Vries, E.F.J. Is cyclooxygenase-1 involved in neuroinflammation? J. Neurosci. Res. 2021, 99, 2976–2998. [Google Scholar] [CrossRef] [PubMed]
- Aïd, S.; Bosetti, F. Targeting cyclooxygenases-1 and -2 in neuroinflammation: Therapeutic implications. Biochimie 2011, 93, 46. [Google Scholar] [CrossRef] [PubMed]
- Koob, G.F.; Ahmed, S.H.; Boutrel, B.; Chen, S.A.; Kenny, P.J.; Markou, A.; O’Dell, L.E.; Parsons, L.H.; Sanna, P.P. Neurobiological mechanisms in the transition from drug use to drug dependence. Neurosci. Biobehav. Rev. 2004, 27, 739–749. [Google Scholar] [CrossRef]
- Gasbarri, A.; Sulli, A.; Packard, M.G. The dopaminergic mesencephalic projections to the hippocampal formation in the rat. Prog. Neuro-Psychopharmacol. Biol. Psychiatry 1997, 21, 1–22. [Google Scholar] [CrossRef]
- Lisman, J.E.; Grace, A.A. The hippocampal-VTA loop: Controlling the entry of information into long-term memory. Neuron 2005, 46, 703–713. [Google Scholar] [CrossRef]
- Schott, B.H.; Sellner, D.B.; Lauer, C.J.; Habib, R.; Frey, J.U.; Guderian, S.; Heinze, H.J.; Düzel, E. Activation of midbrain structures by associative novelty and the formation of explicit memory in humans. Learn. Mem. 2004, 11, 383–387. [Google Scholar] [CrossRef] [PubMed]
- Kim, J.; Ham, S.; Hong, H.; Moon, C.; Im, H.-I. Brain Reward Circuits in Morphine Addiction. Mol. Cells 2016, 39, 645–653. [Google Scholar] [CrossRef] [PubMed]
- Ranjbar, H.; Soti, M.; Banazadeh, M.; Saleki, K.; Kohlmeier, K.A.; Shabani, M. Addiction and the cerebellum with a focus on actions of opioid receptors. Neurosci. Biobehav. Rev. 2021, 131, 229–247. [Google Scholar] [CrossRef] [PubMed]
- Etkin, A.; Prater, K.E.; Schatzberg, A.F.; Menon, V.; Greicius, M.D. Disrupted amygdalar subregion functional connectivity and evidence of a compensatory network in generalized anxiety disorder. Arch. Gen. Psychiatry 2009, 66, 1361–1372. [Google Scholar] [CrossRef] [PubMed]
- Chen, C.H.; Fremont, R.; Arteaga-Bracho, E.E.; Khodakhah, K. Short latency cerebellar modulation of the basal ganglia. Nat. Neurosci. 2014, 17, 1767–1775. [Google Scholar] [CrossRef]
- Badura, A.; Verpeut, J.L.; Metzger, J.W.; Pereira, T.D.; Pisano, T.J.; Deverett, B.; Bakshinskaya, D.E.; Wang, S.S.H. Normal cognitive and social development require posterior cerebellar activity. eLife 2018, 7, e36401. [Google Scholar] [CrossRef] [PubMed]
- Farley, S.J.; Radley, J.J.; Freeman, J.H. Amygdala Modulation of Cerebellar Learning. J. Neurosci. 2016, 36, 2190–2201. [Google Scholar] [CrossRef]
- Babayan, B.M.; Watilliaux, A.; Viejo, G.; Paradis, A.L.; Girard, B.; Rondi-Reig, L. A hippocampo-cerebellar centred network for the learning and execution of sequence-based navigation. Sci. Rep. 2017, 7, e36401. [Google Scholar] [CrossRef]
- Schwarz, L.A.; Miyamichi, K.; Gao, X.J.; Beier, K.T.; Weissbourd, B.; Deloach, K.E.; Ren, J.; Ibanes, S.; Malenka, R.C.; Kremer, E.J.; et al. Viral-genetic tracing of the input-output organization of a central noradrenaline circuit. Nature 2015, 524, 88–92. [Google Scholar] [CrossRef]
- Carta, I.; Chen, C.H.; Schott, A.L.; Dorizan, S.; Khodakhah, K. Cerebellar modulation of the reward circuitry and social behavior. Science 2019, 363, eaav0581. [Google Scholar] [CrossRef]
- Moulton, E.A.; Elman, I.; Becerra, L.R.; Goldstein, R.Z.; Borsook, D. The cerebellum and addiction: Insights gained from neuroimaging research. Addict. Biol. 2014, 19, 317–331. [Google Scholar] [CrossRef] [PubMed]
- Ruan, C.H.; So, S.P.; Ruan, K.H. Inducible COX-2 dominates over COX-1 in prostacyclin biosynthesis: Mechanisms of COX-2 inhibitor risk to heart disease. Life Sci. 2011, 88, 24–30. [Google Scholar] [CrossRef] [PubMed]
- Zielinski, M.R.; Krueger, J.M. Inflammation and Sleep. In Therapy in Sleep Medicine; Elsevier Inc.: Amsterdam, The Netherlands, 2012; pp. 607–616. ISBN 9781437717037. [Google Scholar]
- Hewett, S.J.; Bell, S.C.; Hewett, J.A. Contributions of cyclooxygenase-2 to neuroplasticity and neuropathology of the central nervous system. Pharmacol. Ther. 2006, 112, 335–357. [Google Scholar] [CrossRef] [PubMed]
- Slanina, K.A.; Schweitzer, P. Inhibition of cyclooxygenase-2 elicits a CB1-mediated decrease of excitatory transmission in rat CA1 hippocampus. Neuropharmacology 2005, 49, 653–659. [Google Scholar] [CrossRef] [PubMed]
- Chan, M.M.-Y.; Moore, A.R. Resolution of inflammation in murine autoimmune arthritis is disrupted by cyclooxygenase-2 inhibition and restored by prostaglandin E2-mediated lipoxin A4 production. J. Immunol. 2010, 184, 6418–6426. [Google Scholar] [CrossRef] [PubMed]
- Farooqui, M.; Li, Y.; Rogers, T.; Poonawala, T.; Griffin, R.J.; Song, C.W.; Gupta, K. COX-2 inhibitor celecoxib prevents chronic morphine-induced promotion of angiogenesis, tumour growth, metastasis and mortality, without compromising analgesia. Br. J. Cancer 2007, 97, 1523. [Google Scholar] [CrossRef] [PubMed]
- Hutchinson, M.R.; Zhang, Y.; Shridhar, M.; Evans, J.H.; Buchanan, M.M.; Zhao, T.X.; Slivka, P.F.; Coats, B.D.; Rezvani, N.; Wieseler, J.; et al. Evidence that opioids may have toll-like receptor 4 and MD-2 effects. Brain. Behav. Immun. 2010, 24, 83–95. [Google Scholar] [CrossRef] [PubMed]
- Lewis, S.S.; Hutchinson, M.R.; Rezvani, N.; Loram, L.C.; Zhang, Y.; Maier, S.F.; Rice, K.C.; Watkins, L.R. Evidence that intrathecal morphine-3-glucuronide may cause pain enhancement via toll-like receptor 4/MD-2 and interleukin-1β. Neuroscience 2010, 165, 569–583. [Google Scholar] [CrossRef]
- Maragakis, N.J.; Rothstein, J.D. Mechanisms of Disease: Astrocytes in neurodegenerative disease. Nat. Clin. Pract. Neurol. 2006, 2, 679–689. [Google Scholar] [CrossRef]
- Stichel, C.C.; Müller, H.W. The CNS lesion scar: New vistas on an old regeneration barrier. Cell Tissue Res. 1998, 294, 1–9. [Google Scholar] [CrossRef]
- Sami, A.; Selzer, M.E.; Li, S. Advances in the Signaling Pathways Downstream of Glial-Scar Axon Growth Inhibitors. Front. Cell. Neurosci. 2020, 14, 174. [Google Scholar] [CrossRef]
- Song, P.; Zhao, Z.Q. The involvement of glial cells in the development of morphine tolerance. Neurosci. Res. 2001, 39, 281–286. [Google Scholar] [CrossRef] [PubMed]
- Kaur, K.; Gill, J.S.; Bansal, P.K.; Deshmukh, R. Neuroinflammation—A major cause for striatal dopaminergic degeneration in Parkinson’s disease. J. Neurol. Sci. 2017, 381, 308–314. [Google Scholar] [CrossRef] [PubMed]
- Imai, Y.; Kohsaka, S. Intracellular signaling in M-CSF-induced microglia activation: Role of Iba1. Glia 2002, 40, 164–174. [Google Scholar] [CrossRef] [PubMed]
- Hopperton, K.E.; Mohammad, D.; Trépanier, M.O.; Giuliano, V.; Bazinet, R.P. Markers of microglia in post-mortem brain samples from patients with Alzheimer’s disease: A systematic review. Mol. Psychiatry 2017, 23, 177–198. [Google Scholar] [CrossRef] [PubMed]
- Liu, H.; Leak, R.K.; Hu, X. Neurotransmitter receptors on microglia. Stroke Vasc. Neurol. 2016, 1, 52–58. [Google Scholar] [CrossRef] [PubMed]
- Pocock, J.M.; Kettenmann, H. Neurotransmitter receptors on microglia. Trends Neurosci. 2007, 30, 527–535. [Google Scholar] [CrossRef]
- Peng, J.; Pan, J.; Wang, H.; Mo, J.; Lan, L.; Peng, Y. Morphine-induced microglial immunosuppression via activation of insufficient mitophagy regulated by NLRX1. J. Neuroinflammation 2022, 19, 87. [Google Scholar] [CrossRef]
- Qiu, S.; Feng, Y.; LeSage, G.; Zhang, Y.; Stuart, C.; He, L.; Li, Y.; Caudle, Y.; Peng, Y.; Yin, D. Chronic Morphine-Induced MicroRNA-124 Promotes Microglial Immunosuppression by Modulating P65 and TRAF6. J. Immunol. 2015, 194, 1021–1030. [Google Scholar] [CrossRef]
- Sacerdote, P. Opioid-induced immunosuppression. Curr. Opin. Support. Palliat. Care 2008, 2, 14–18. [Google Scholar] [CrossRef]
- Chen, M.; Zhao, Y.; Yang, H.; Luan, W.; Song, J.; Cui, D.; Dong, Y.; Lai, B.; Ma, L.; Zheng, P. Morphine disinhibits glutamatergic input to VTA dopamine neurons and promotes dopamine neuron excitation. eLife 2015, 4, e09275. [Google Scholar] [CrossRef]
- Horvath, R.J.; DeLeo, J.A. Morphine Enhances Microglial Migration through Modulation of P2X4 Receptor Signaling. J. Neurosci. 2009, 29, 998–1005. [Google Scholar] [CrossRef]
- Horvath, R.J.; Romero-Sandoval, E.A.; Leo, J.A.D. Inhibition of microglial P2X4 receptors attenuates morphine tolerance, Iba1, GFAP and μ opioid receptor protein expression while enhancing perivascular microglial ED2. Pain 2010, 150, 401–413. [Google Scholar] [CrossRef]
- Metryka, E.; Gutowska, I.; Kupnicka, P.; Tarnowski, M.; Tkacz, M.; Listos, J.; Talarek, S.; Barczak, K.; Chlubek, D.; Baranowska-Bosiacka, I. The expression of purinergic P2X4 and P2X7 receptors in selected mesolimbic structures during morphine withdrawal in rats. Brain Res. 2019, 1719, 49–56. [Google Scholar] [CrossRef]
- Dunipace, A.J.; Brizendine, E.J.; Zhang, W.; Wilson, M.E.; Miller, L.L.; Katz, B.P.; Warrick, J.M.; Stookey, G.K. Effect of aging on animal response to chronic fluoride exposure. J. Dent. Res. 1995, 74, 358–368. [Google Scholar] [CrossRef]
- McPherson, C.A.; Zhang, G.; Gilliam, R.; Brar, S.S.; Wilson, R.; Brix, A.; Picut, C.; Harry, G.J. An Evaluation of Neurotoxicity Following Fluoride Exposure from Gestational through Adult Ages in Long-Evans Hooded Rats. Neurotox. Res. 2018, 34, 781–798. [Google Scholar] [CrossRef]
- Fernando, W.B.N.T.; Nanayakkara, N.; Gunarathne, L.; Chandrajith, R. Serum and urine fluoride levels in populations of high environmental fluoride exposure with endemic CKDu: A case-control study from Sri Lanka. Environ. Geochem. Health 2020, 42, 1497–1504. [Google Scholar] [CrossRef]
- Usman, M.; Ali, A.; Jabbar Siddiqui, A.; Iftikhar, F.; Kumari, S.; Sibt-e-Hassan, S.; Musharraf, S.G. Evaluation of the chronic intoxication of fluoride on human serum metabolome using untargeted metabolomics. Arab. J. Chem. 2022, 15, 103928. [Google Scholar] [CrossRef]











{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Prefrontal Cortex | Striatum | Hippocampus | Cerebellum | |||||
|---|---|---|---|---|---|---|---|---|
| Iba1 | GFAP | Iba1 | GFAP | Iba1 | GFAP | Iba1 | GFAP | |
| Control (C) | + | + | ++ | + | + | + | + | + |
| Fluoride (F) | + | ++ | +++ | +++ | ++ | ++ | + | ++ |
| Morphine (M) | +++ | ++ | ++ | + | + | + | + | + |
| Morphine + Fluoride (M + F) | ++ | +++ | ++ | + | + | + | + | ++ |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Kupnicka, P.; Listos, J.; Tarnowski, M.; Kolasa, A.; Kapczuk, P.; Surówka, A.; Kwiatkowski, J.; Janawa, K.; Chlubek, D.; Baranowska-Bosiacka, I. The Effect of Prenatal and Neonatal Fluoride Exposure to Morphine-Induced Neuroinflammation. Int. J. Mol. Sci. 2024, 25, 826. https://doi.org/10.3390/ijms25020826
Kupnicka P, Listos J, Tarnowski M, Kolasa A, Kapczuk P, Surówka A, Kwiatkowski J, Janawa K, Chlubek D, Baranowska-Bosiacka I. The Effect of Prenatal and Neonatal Fluoride Exposure to Morphine-Induced Neuroinflammation. International Journal of Molecular Sciences. 2024; 25(2):826. https://doi.org/10.3390/ijms25020826
Chicago/Turabian StyleKupnicka, Patrycja, Joanna Listos, Maciej Tarnowski, Agnieszka Kolasa, Patrycja Kapczuk, Anna Surówka, Jakub Kwiatkowski, Kamil Janawa, Dariusz Chlubek, and Irena Baranowska-Bosiacka. 2024. "The Effect of Prenatal and Neonatal Fluoride Exposure to Morphine-Induced Neuroinflammation" International Journal of Molecular Sciences 25, no. 2: 826. https://doi.org/10.3390/ijms25020826
APA StyleKupnicka, P., Listos, J., Tarnowski, M., Kolasa, A., Kapczuk, P., Surówka, A., Kwiatkowski, J., Janawa, K., Chlubek, D., & Baranowska-Bosiacka, I. (2024). The Effect of Prenatal and Neonatal Fluoride Exposure to Morphine-Induced Neuroinflammation. International Journal of Molecular Sciences, 25(2), 826. https://doi.org/10.3390/ijms25020826