
Genome-Wide Identification and Expression Analysis of the ALKB Homolog Gene Family in Potato (Solanum tuberosum L.)

Abstract
1. Introduction
2. Results
2.1. Genome-Wide Identification of ALKBH Gene Family Members in Potato
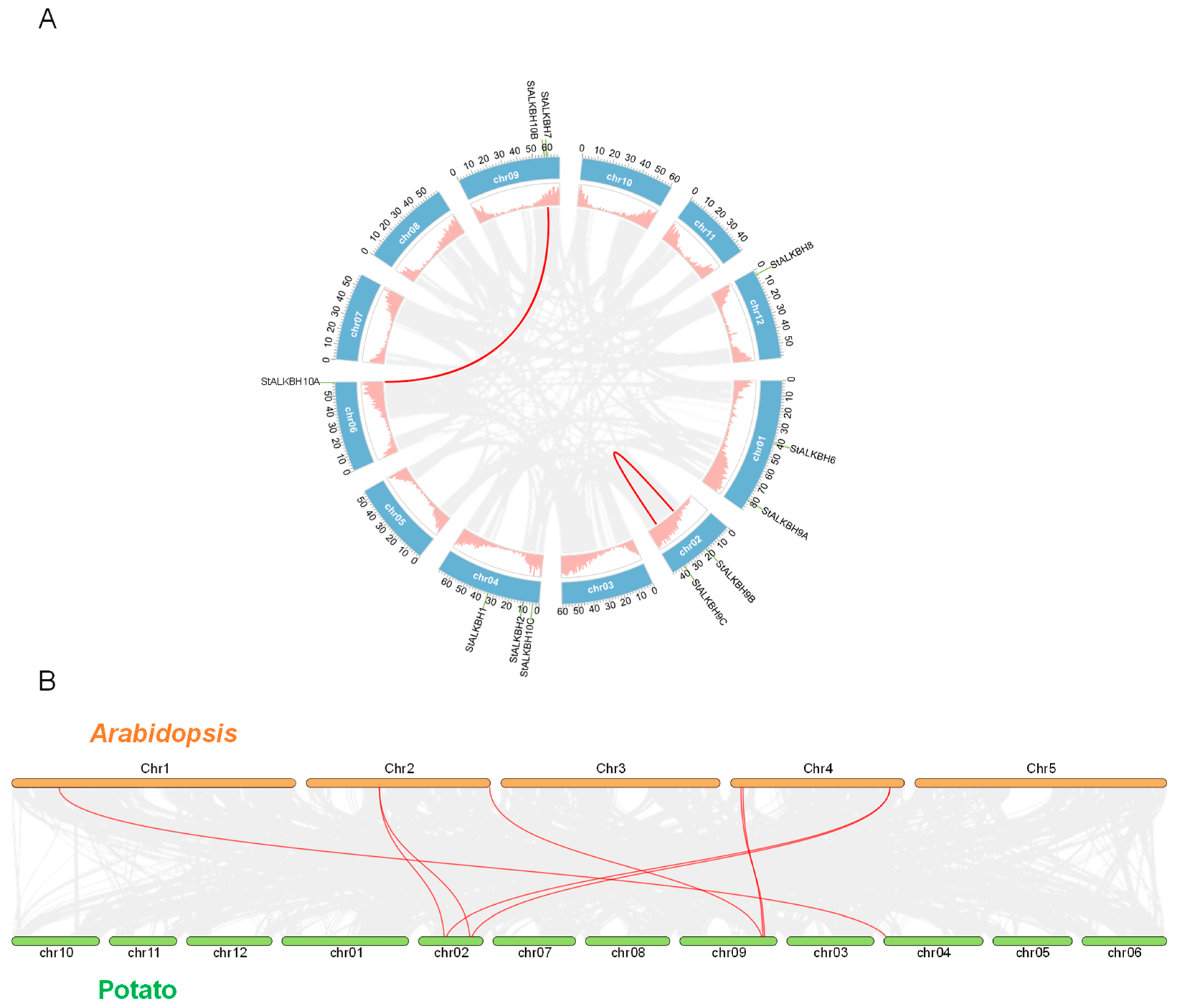
2.2. Chromosomal Location and Collinearity Analysis of ALKBH Family
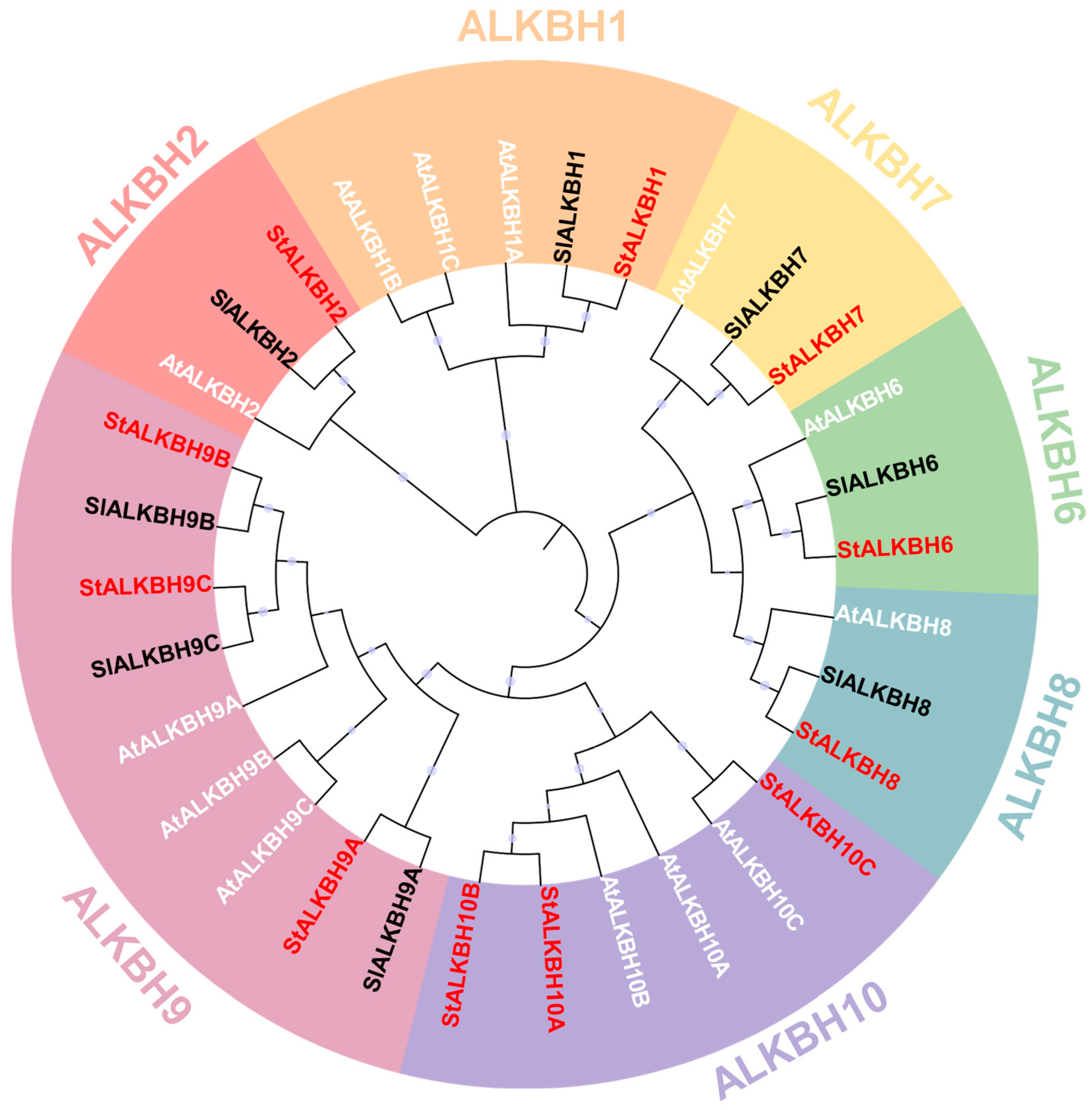
2.3. Evolutionary and Sequence Analyses of StALKBH Family
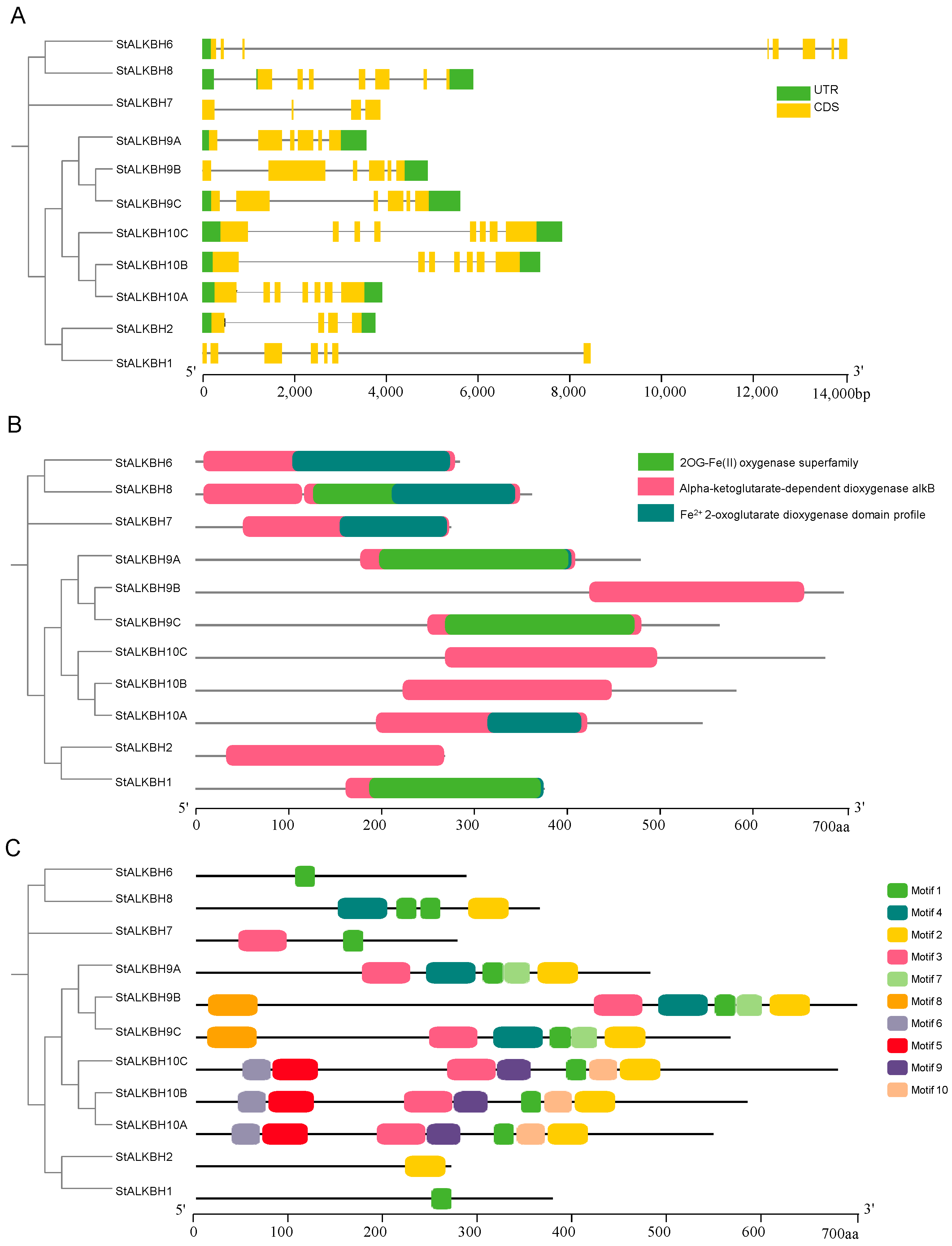
2.4. Gene Structure, Conserved Motifs, and Promoter Analysis of StALKBHs
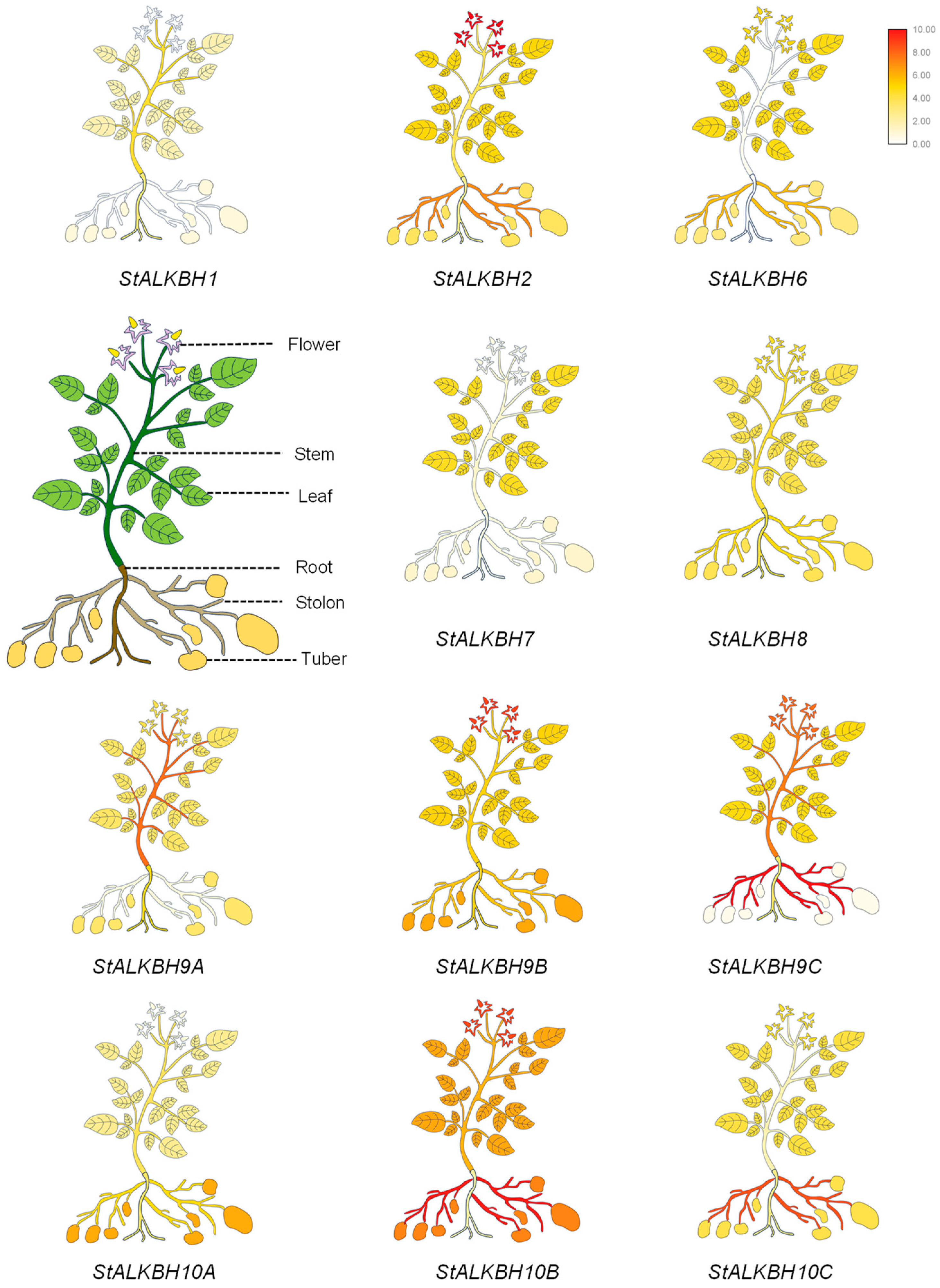
2.5. Tissue Expression of StALKBH Family Genes
2.6. Analysis of StALKBH Family Genes under Abiotic Stress Treatments
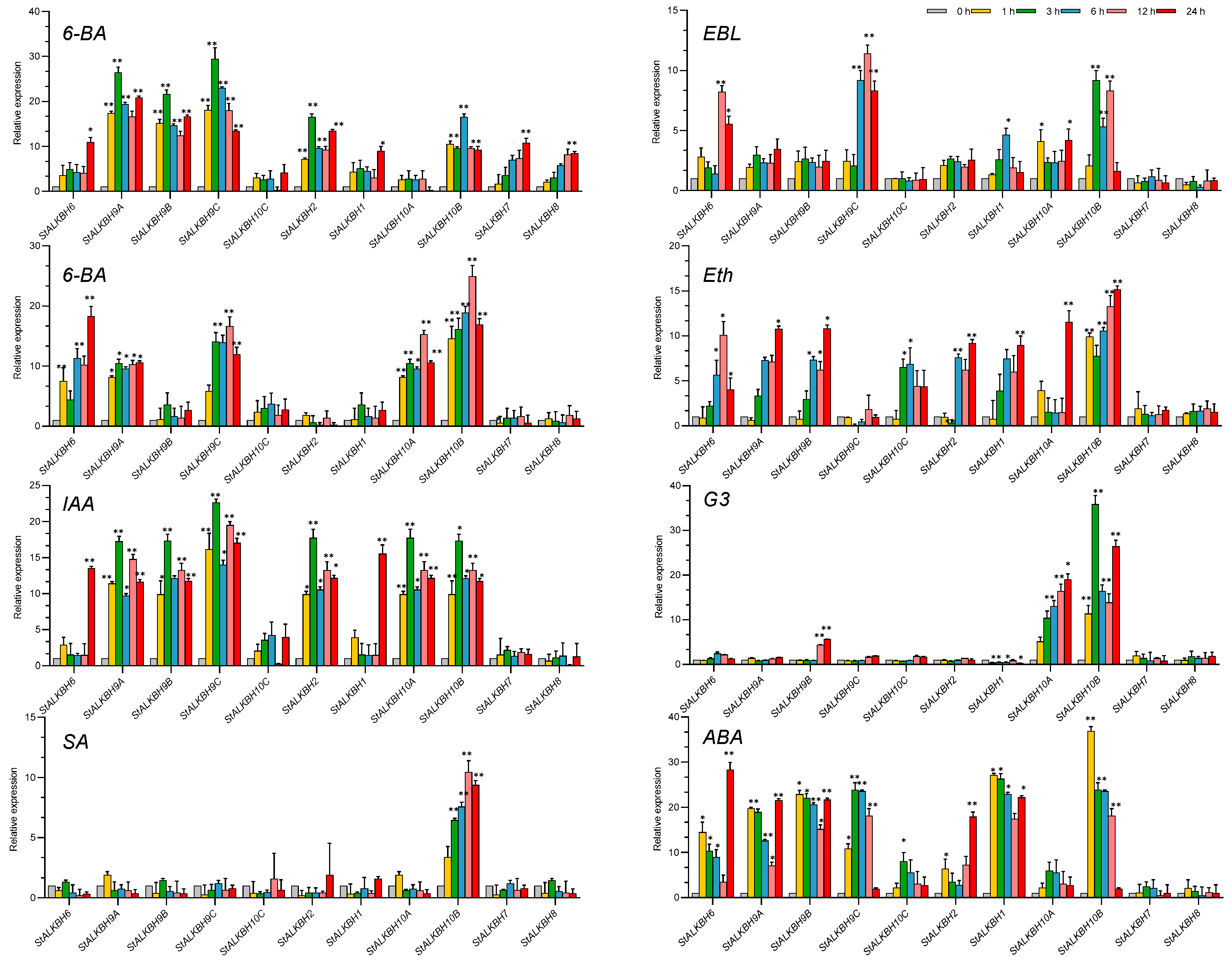
2.7. Analysis of StALKBH Family Genes under Hormone Treatments
3. Discussion
4. Materials and Methods
4.1. Plant Material, Growth Conditions, and Hormone and Stress Treatments
4.2. Identification of StALKBH Genes and Synteny Analysis
4.3. Phylogenetic Analysis, Gene Structure, Protein Motifs, and Promoter Sequence Analysis
4.4. Chromosomal Location and Syntenic Analysis
4.5. Hormone and Stress Treatments and Quantitative Real-Time PCR (qRT-PCR)
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Shao, Y.; Wong, C.E.; Shen, L.; Yu, H. N6-methyladenosine modification underlies messenger RNA metabolism and plant development. Curr. Opin. Plant Biol. 2021, 63, 102047. [Google Scholar] [CrossRef] [PubMed]
- Hu, J.; Cai, J.; Xu, T.; Kang, H. Epitranscriptomic mRNA modifications governing plant stress responses: Underlying mechanism and potential application. Plant Biotechnol. J. 2022, 20, 2245–2257. [Google Scholar] [CrossRef] [PubMed]
- Cai, J.; Hu, J.; Xu, T.; Kang, H. FIONA1-mediated mRNA m6A methylation regulates the response of Arabidopsis to salt stress. Plant Cell Environ. 2024, 47, 900–912. [Google Scholar] [CrossRef] [PubMed]
- Miao, Z.; Zhang, T.; Xie, B.; Qi, Y.; Ma, C. Evolutionary Implications of the RNA N6-Methyladenosine Methylome in Plants. Mol. Biol. Evol. 2022, 39, msab299. [Google Scholar] [CrossRef]
- Shen, L.; Ma, J.; Li, P.; Wu, Y.; Yu, H. Recent advances in the plant epitranscriptome. Genome Biol. 2023, 24, 43. [Google Scholar] [CrossRef] [PubMed]
- Yue, H.; Nie, X.; Yan, Z.; Weining, S. N6-methyladenosine regulatory machinery in plants: Composition, function and evolution. Plant Biotechnol. J. 2019, 17, 1194–1208. [Google Scholar] [CrossRef]
- Tang, J.; Chen, S.; Jia, G. Detection, regulation, and functions of RNA N6-methyladenosine modification in plants. Plant Commun. 2023, 4, 100546. [Google Scholar] [CrossRef]
- Jalal, A.; Sun, J.; Chen, Y.; Fan, C.; Liu, J.; Wang, C. Evolutionary Analysis and Functional Identification of Clock-Associated PSEUDO-RESPONSE REGULATOR (PRRs) Genes in the Flowering Regulation of Roses. Int. J. Mol. Sci. 2022, 23, 7335. [Google Scholar] [CrossRef]
- Bhat, S.S.; Bielewicz, D.; Gulanicz, T.; Bodi, Z.; Yu, X.; Anderson, S.J.; Szewc, L.; Bajczyk, M.; Dolata, J.; Grzelak, N.; et al. mRNA adenosine methylase (MTA) deposits m6a on pri-miRNAs to modulate miRNA biogenesis in Arabidopsis thaliana. Proc. Natl. Acad. Sci. USA 2020, 117, 21785–21795. [Google Scholar] [CrossRef]
- Kataoka, H.; Yamamoto, Y.; Sekiguchi, M. A new gene (alkB) of Escherichia coli that controls sensitivity to methyl methane sulfonate. J. Bacteriol. 1983, 153, 1301–1307. [Google Scholar] [CrossRef]
- Zheng, G.; Dahl, J.A.; Niu, Y.; Fedorcsak, P.; Huang, C.-M.; Li, C.J.; Vågbø, C.B.; Shi, Y.; Wang, W.-L.; Song, S.-H.; et al. ALKBH5 Is a Mammalian RNA Demethylase that Impacts RNA Metabolism and Mouse Fertility. Mol. Cell 2013, 49, 18–29. [Google Scholar] [CrossRef] [PubMed]
- Jia, G.; Fu, Y.; Zhao, X.; Dai, Q.; Zheng, G.; Yang, Y.; Yi, C.; Lindahl, T.; Pan, T.; Yang, Y.-G.; et al. N6-Methyladenosine in nuclear RNA is a major substrate of the obesity-associated FTO. Nat. Chem. Biol. 2011, 7, 885–887. [Google Scholar] [CrossRef] [PubMed]
- Duan, H.-C.; Wei, L.-H.; Zhang, C.; Wang, Y.; Chen, L.; Lu, Z.; Chen, P.R.; He, C.; Jia, G. ALKBH10B Is an RNA N6-Methyladenosine Demethylase Affecting Arabidopsis Floral Transition. Plant Cell 2017, 29, 2995–3011. [Google Scholar] [CrossRef] [PubMed]
- Zhou, L.; Tian, S.; Qin, G. RNA methylomes reveal the m6A-mediated regulation of DNA demethylase gene SlDML2 in tomato fruit ripening. Genome Biol. 2019, 20, 156. [Google Scholar] [CrossRef]
- Merret, R.; Nagarajan, V.K.; Carpentier, M.-C.; Park, S.; Favory, J.-J.; Descombin, J.; Picart, C.; Charng, Y.-Y.; Green, P.J.; Deragon, J.-M.; et al. Heat-induced ribosome pausing triggers mRNA co-translational decay in Arabidopsis thaliana. Nucleic Acids Res. 2015, 43, 4121–4132. [Google Scholar] [CrossRef]
- Tang, J.; Yang, J.; Duan, H.; Jia, G. ALKBH10B, an mRNA m6A Demethylase, Modulates ABA Response During Seed Germination in Arabidopsis. Front. Plant Sci. 2021, 12, 712713. [Google Scholar] [CrossRef]
- Huong, T.T.; Ngoc, L.N.T.; Kang, H. Functional Characterization of a Putative RNA Demethylase ALKBH6 in Arabidopsis Growth and Abiotic Stress Responses. Int. J. Mol. Sci. 2020, 21, 6707. [Google Scholar] [CrossRef]
- Li, B.; Zhang, M.; Sun, W.; Yue, D.; Ma, Y.; Zhang, B.; Duan, L.; Wang, M.; Lindsey, K.; Nie, X.; et al. N6-methyladenosine RNA modification regulates cotton drought response in a Ca2+ and ABA-dependent manner. Plant Biotechnol. J. 2023, 21, 1270–1285. [Google Scholar] [CrossRef]
- Cui, C.; Ma, Z.; Wan, H.; Gao, J.; Zhou, B. GhALKBH10 negatively regulates salt tolerance in cotton. Plant Physiol. Biochem. 2022, 192, 87–100. [Google Scholar] [CrossRef]
- Zhou, L.; Gao, G.; Tang, R.; Wang, W.; Wang, Y.; Tian, S.; Qin, G. m6A-mediated regulation of crop development and stress responses. Plant Biotechnol. J. 2022, 20, 1447–1455. [Google Scholar] [CrossRef]
- Shen, H.; Luo, B.; Wang, Y.; Li, J.; Hu, Z.; Xie, Q.; Wu, T.; Chen, G. Genome-Wide Identification, Classification and Expression Analysis of m6A Gene Family in Solanum lycopersicum. Int. J. Mol. Sci. 2022, 23, 4522. [Google Scholar] [CrossRef] [PubMed]
- Zhao, Y.; Guo, Q.; Cao, S.; Tian, Y.; Han, K.; Sun, Y.; Li, J.; Yang, Q.; Ji, Q.; Sederoff, R.; et al. Genome-wide identification of the AlkB homologs gene family, PagALKBH9B and PagALKBH10B regulated salt stress response in Populus. Front. Plant Sci. 2022, 13, 994154. [Google Scholar] [CrossRef]
- Xu, G.; Guo, C.; Shan, H.; Kong, H. Divergence of duplicate genes in exon–intron structure. Proc. Natl. Acad. Sci. USA 2012, 109, 1187–1192. [Google Scholar] [CrossRef] [PubMed]
- Huang, A.; Wang, Y.; Gu, P.; Yang, Z.; Han, J.; Yi, L. Genome-Wide Identification and Characterization of the AlkB Gene Family in Sweet Orange (Citrus sinensis). Curr. Issues Mol. Biol. 2022, 45, 122–133. [Google Scholar] [CrossRef] [PubMed]
- Fedeles, B.I.; Singh, V.; Delaney, J.C.; Li, D.; Essigmann, J.M. The AlkB Family of Fe(II)/α-Ketoglutarate-dependent Dioxygenases: Repairing Nucleic Acid Alkylation Damage and Beyond. J. Biol. Chem. 2015, 290, 20734–20742. [Google Scholar] [CrossRef]
- Moore, R.C.; Purugganan, M.D. The early stages of duplicate gene evolution. Proc. Natl. Acad. Sci. USA 2003, 100, 15682–15687. [Google Scholar] [CrossRef]
- Kong, H.; Landherr, L.L.; Frohlich, M.W.; Leebens-Mack, J.; Ma, H.; dePamphilis, C.W. Patterns of gene duplication in the plant SKP1 gene family in angiosperms: Evidence for multiple mechanisms of rapid gene birth. Plant J. 2007, 50, 873–885. [Google Scholar] [CrossRef]
- Tang, J.; Lei, D.; Yang, J.; Chen, S.; Wang, X.; Huang, X.; Zhang, S.; Cai, Z.; Zhu, S.; Wan, J.; et al. OsALKBH9-mediated m6A demethylation regulates tapetal PCD and pollen exine accumulation in rice. Plant Biotechnol. J. 2024, 22, 2410–2423. [Google Scholar] [CrossRef] [PubMed]
- Yu, Q.; Liu, S.; Yu, L.; Xiao, Y.; Zhang, S.; Wang, X.; Xu, Y.; Yu, H.; Li, Y.; Yang, J.; et al. RNA demethylation increases the yield and biomass of rice and potato plants in field trials. Nat. Biotechnol. 2021, 39, 1581–1588. [Google Scholar] [CrossRef]
- Huong, T.T.; Yang, Z.; Ngoc, L.N.T.; Kang, H. ALKBH8B, a Putative RNA Demethylase, Plays a Role in the Response of Arabidopsis to Salt Stress and Abscisic Acid. J. Plant Biol. 2022, 65, 319–330. [Google Scholar] [CrossRef]
- Pham, G.M.; Hamilton, J.P.; Wood, J.C.; Burke, J.T.; Zhao, H.; Vaillancourt, B.; Ou, S.; Jiang, J.; Buell, C.R. Construction of a chromosome-scale long-read reference genome assembly for potato. GigaScience 2020, 9, giaa100. [Google Scholar] [CrossRef] [PubMed]
- Chen, C.; Chen, H.; Zhang, Y.; Thomas, H.R.; Frank, M.H.; He, Y.; Xia, R. TBtools: An Integrative Toolkit Developed for Interactive Analyses of Big Biological Data. Mol. Plant 2020, 13, 1194–1202. [Google Scholar] [CrossRef] [PubMed]



{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Gene Name | Gene ID | Arabidopsis Gene | CDS (bp) | Protein Length (aa) | Molecular Weight (KD) | pI | GRAVY | Subcellular Localization |
|---|---|---|---|---|---|---|---|---|
| StALKBH6 | Soltu.DM.01G016630.1 | AT4G20350 | 807 | 268 | 30.01 | 6.29 | −0.17 | Nucleus |
| StALKBH9A | Soltu.DM.01G043090.1 | AT1G48980 | 1356 | 451 | 51.06 | 8.86 | −0.57 | Nucleus |
| StALKBH9B | Soltu.DM.02G005900.1 | AT2G17970 | 1974 | 657 | 72.67 | 6.03 | −0.57 | Nucleus |
| StALKBH9C | Soltu.DM.02G023950.1 | AT4G36090 | 1596 | 531 | 59.60 | 6.22 | −0.65 | Nucleus |
| StALKBH10C | Soltu.DM.04G003750.1 | AT1G14710 | 1917 | 638 | 69.15 | 6.67 | −0.49 | Nucleus |
| StALKBH2 | Soltu.DM.04G010040.1 | AT2G22260 | 762 | 253 | 29.06 | 9.07 | −0.64 | Nucleus |
| StALKBH1 | Soltu.DM.04G016670.2 | AT1G11780 | 1065 | 354 | 39.88 | 5.74 | −0.31 | Nucleus |
| StALKBH10A | Soltu.DM.06G034880.2 | AT2G48080 | 1545 | 514 | 56.77 | 8.06 | −0.26 | Nucleus |
| StALKBH10B | Soltu.DM.09G022420.1 | AT4G02940 | 1647 | 548 | 60.64 | 5.97 | −0.39 | Nucleus |
| StALKBH7 | Soltu.DM.09G023290.2 | AT4G02485 | 780 | 259 | 29.39 | 4.69 | −0.43 | Nucleus |
| StALKBH8 | Soltu.DM.12G004160.1 | AT1G31600 | 1026 | 341 | 38.61 | 6.32 | −0.40 | Nucleus |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Li, Y.; Dong, X.; Ma, J.; Sui, C.; Jian, H.; Lv, D. Genome-Wide Identification and Expression Analysis of the ALKB Homolog Gene Family in Potato (Solanum tuberosum L.). Int. J. Mol. Sci. 2024, 25, 10984. https://doi.org/10.3390/ijms252010984
Li Y, Dong X, Ma J, Sui C, Jian H, Lv D. Genome-Wide Identification and Expression Analysis of the ALKB Homolog Gene Family in Potato (Solanum tuberosum L.). International Journal of Molecular Sciences. 2024; 25(20):10984. https://doi.org/10.3390/ijms252010984
Chicago/Turabian StyleLi, Yan, Xuanming Dong, Jianyu Ma, Chenxin Sui, Hongju Jian, and Dianqiu Lv. 2024. "Genome-Wide Identification and Expression Analysis of the ALKB Homolog Gene Family in Potato (Solanum tuberosum L.)" International Journal of Molecular Sciences 25, no. 20: 10984. https://doi.org/10.3390/ijms252010984
APA StyleLi, Y., Dong, X., Ma, J., Sui, C., Jian, H., & Lv, D. (2024). Genome-Wide Identification and Expression Analysis of the ALKB Homolog Gene Family in Potato (Solanum tuberosum L.). International Journal of Molecular Sciences, 25(20), 10984. https://doi.org/10.3390/ijms252010984