
Autumn Olive (Elaeagnus umbellata Thunb.) Berries Improve Lipid Metabolism and Delay Aging in Middle-Aged Caenorhabditis elegans
Abstract
1. Introduction
2. Results
2.1. Confirmation of the Antioxidant Activity and Polyphenol Contents of FAOBs and SAOBs
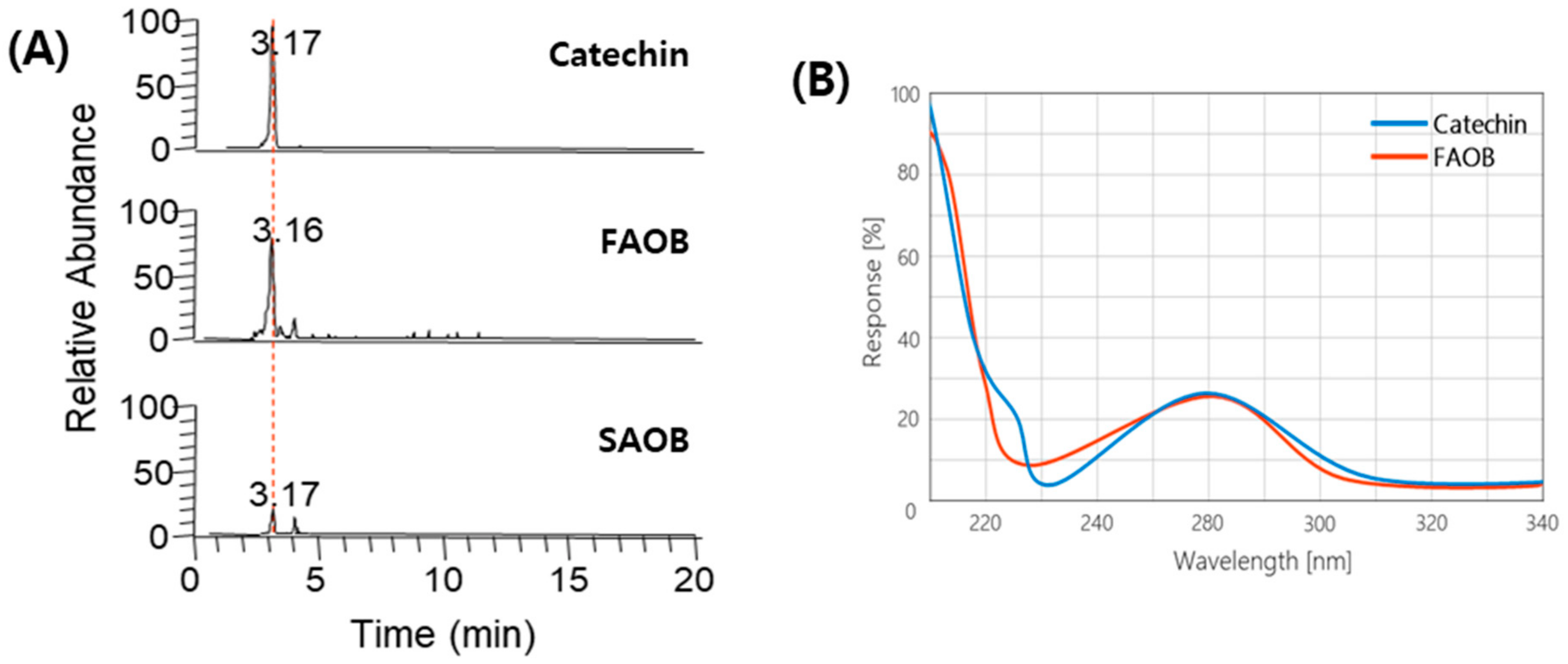
2.2. Catechin Identification
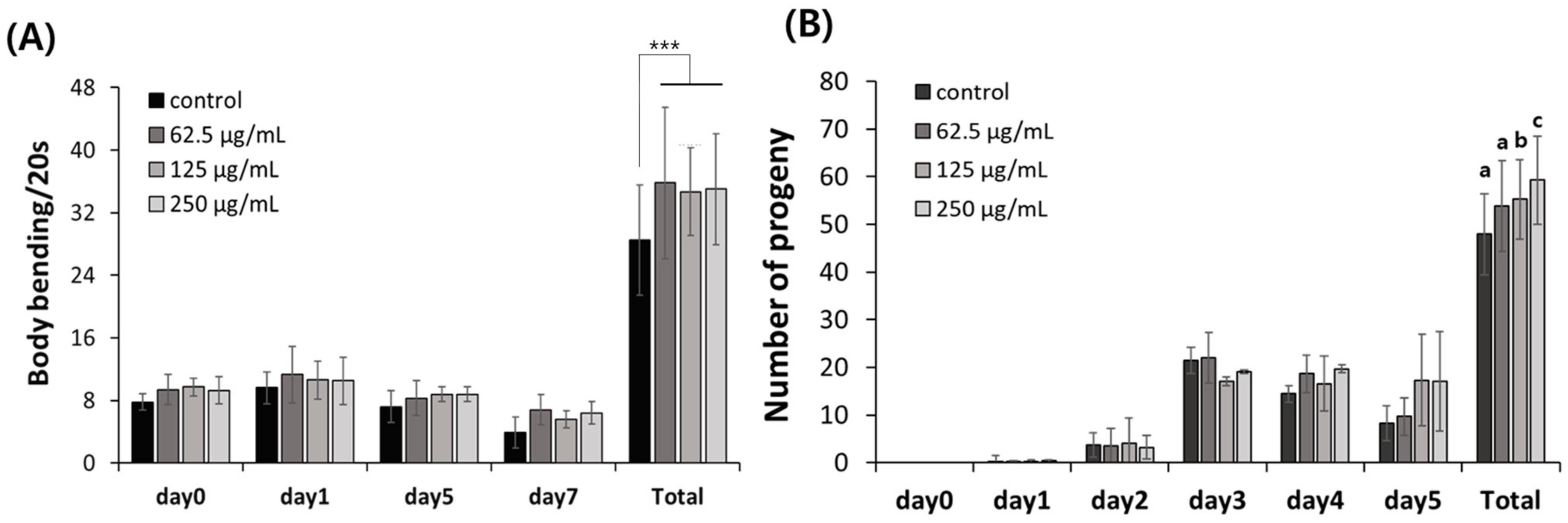
2.3. Safety of FAOB in C. elegans
2.4. FAOB Inhibits Lipid Accumulation in C. elegans
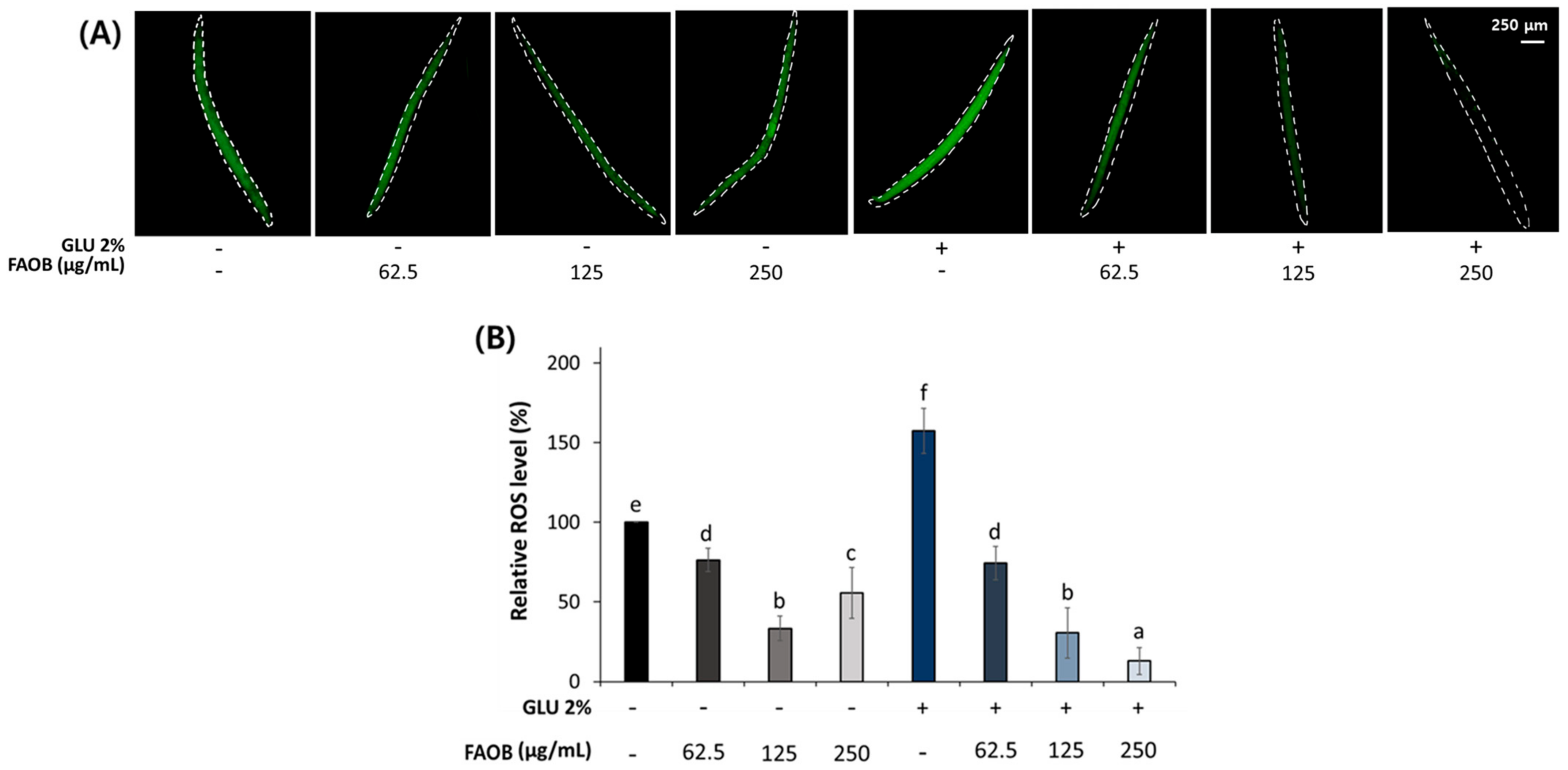
2.5. AOB Reduces Reactive Oxygen Species (ROS) Accumulation in Middle-Aged C. elegans
2.6. FAOB Exhibits an Age-Delaying Effect in C. elegans
2.7. FAOB Regulates Lipid Accumulation by Daf-16 under 2% GLU Dietary Conditions
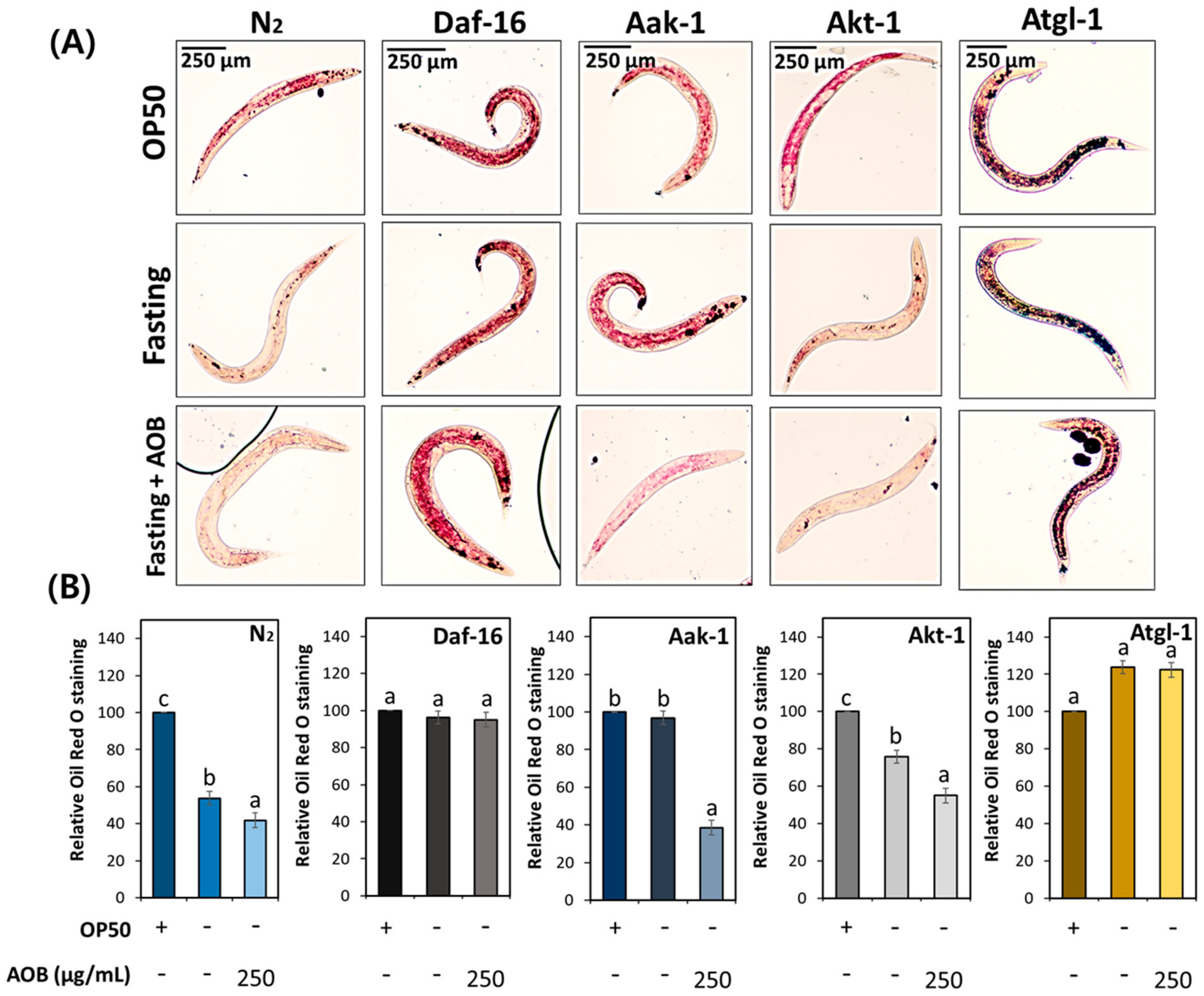
2.8. FAOB Is Involved in Lipolysis Signaling Pathway under Fasting Conditions
3. Discussion
4. Materials and Methods
4.1. Preparation of AOB Extracts
4.2. Reagents
4.3. HPLC Analysis
4.4. Total Phenolic Contents (TPC) of AOB
4.5. DPPH Radical Scavenging Capacity
4.6. Worm Study
4.6.1. Worm Culture
4.6.2. Acute Toxicity
4.6.3. Determination of Stress Resistance
4.6.4. Oil Red O Staining
4.6.5. Triglyceride (TG) Quantification Assay
4.6.6. Reproduction and Pumping Rates
4.6.7. Determination of ROS Level
4.7. Statistical Analysis
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Ng, M.; Fleming, T.; Robinson, M.; Thomson, B.; Graetz, N.; Margono, C.; Mullany, E.C.; Biryukov, S.; Abbafati, C.; Abera, S.F. Global, regional, and national prevalence of overweight and obesity in children and adults during 1980–2013: A systematic analysis for the Global Burden of Disease Study 2013. Lancet 2014, 384, 766–781. [Google Scholar] [CrossRef] [PubMed]
- Geng, J.; Ni, Q.; Sun, W.; Li, L.; Feng, X. The links between gut microbiota and obesity and obesity related diseases. Biomed. Pharmacother. 2022, 147, 112678. [Google Scholar] [CrossRef] [PubMed]
- Lodhi, I.J.; Yin, L.; Jensen-Urstad, A.P.; Funai, K.; Coleman, T.; Baird, J.H.; El Ramahi, M.K.; Razani, B.; Song, H.; Fu-Hsu, F. Inhibiting adipose tissue lipogenesis reprograms thermogenesis and PPARγ activation to decrease diet-induced obesity. Cell Metab. 2012, 16, 189–201. [Google Scholar] [CrossRef] [PubMed]
- Arner, P. Catecholamine-induced lipolysis in obesity. Int. J. Obes. 1999, 23, S10–S13. [Google Scholar] [CrossRef] [PubMed]
- Guarente, L.; Kenyon, C. Genetic pathways that regulate ageing in model organisms. Nature 2000, 408, 255–262. [Google Scholar] [CrossRef]
- Silverman, G.A.; Luke, C.J.; Bhatia, S.R.; Long, O.S.; Vetica, A.C.; Perlmutter, D.H.; Pak, S.C. Modeling molecular and cellular aspects of human disease using the nematode Caenorhabditis elegans. Pediatr. Res. 2009, 65, 10–18. [Google Scholar] [CrossRef] [PubMed]
- Svensk, E.; Devkota, R.; Ståhlman, M.; Ranji, P.; Rauthan, M.; Magnusson, F.; Hammarsten, S.; Johansson, M.; Borén, J.; Pilon, M. Caenorhabditis elegans PAQR-2 and IGLR-2 protect against glucose toxicity by modulating membrane lipid composition. PLoS Genet. 2016, 12, e1005982. [Google Scholar]
- Alcántar-Fernández, J.; González-Maciel, A.; Reynoso-Robles, R.; Pérez Andrade, M.E.; Hernández-Vázquez, A.D.J.; Velázquez-Arellano, A.; Miranda-Ríos, J. High-glucose diets induce mitochondrial dysfunction in Caenorhabditis elegans. PLoS ONE 2019, 14, e0226652. [Google Scholar] [CrossRef]
- Ory, M.G.; Anderson, L.A.; Friedman, D.B.; Pulczinski, J.C.; Eugene, N.; Satariano, W.A. Cancer prevention among adults aged 45–64 years: Setting the stage. Am. J. Prev. Med. 2014, 46, S1–S6. [Google Scholar] [CrossRef]
- Strasser, B.; Arvandi, M.; Pasha, E.; Haley, A.; Stanforth, P.; Tanaka, H. Abdominal obesity is associated with arterial stiffness in middle-aged adults. Nutr. Metab. Cardiovasc. Dis. 2015, 25, 495–502. [Google Scholar] [CrossRef]
- Hodson, L.; Banerjee, R.; Rial, B.; Arlt, W.; Adiels, M.; Boren, J.; Marinou, K.; Fisher, C.; Mostad, I.L.; Stratton, I.M. Menopausal status and abdominal obesity are significant determinants of hepatic lipid metabolism in women. J. Am. Heart Assoc. 2015, 4, e002258. [Google Scholar] [CrossRef] [PubMed]
- Pei, R.; Yu, M.; Bruno, R.; Bolling, B.W. Phenolic and tocopherol content of autumn olive (Elaeagnus umbellate) berries. J. Funct. Foods 2015, 16, 305–314. [Google Scholar] [CrossRef]
- Ahmad, S.D.; Sabir, M.S.; Juma, M.; Asad, H.S. Morphological and biochemical variations in Elaeagnus umbellata Thunb. from mountains of Pakistan. Acta Bot. Croat. 2005, 64, 121–128. [Google Scholar]
- Ishaq, S.; Rathore, H.A.; Sabir, S.M.; Maroof, M.S. Antioxidant properties of Elaeagnus umbellata berry solvent extracts against lipid peroxidation in mice brain and liver tissues. Food Sci. Biotechnol. 2015, 24, 673–679. [Google Scholar] [CrossRef]
- Zannou, O.; Pashazadeh, H.; Ghellam, M.; Hassan, A.M.; Koca, I. Optimization of drying temperature for the assessment of functional and physical characteristics of autumn olive berries. J. Food Process. Preserv. 2021, 45, e15658. [Google Scholar] [CrossRef]
- Fordham, I.M.; Clevidence, B.A.; Wiley, E.R.; Zimmerman, R.H. Fruit of autumn olive: A rich source of lycopene. HortScience 2001, 36, 1136–1137. [Google Scholar] [CrossRef]
- Gamba, G.; Donno, D.; Mellano, M.G.; Riondato, I.; De Biaggi, M.; Randriamampionona, D.; Beccaro, G.L. Phytochemical characterization and bioactivity evaluation of autumn olive (Elaeagnus umbellata Thunb.) pseudodrupes as potential sources of health-promoting compounds. Appl. Sci. 2020, 10, 4354. [Google Scholar] [CrossRef]
- Nazir, N.; Zahoor, M.; Nisar, M.; Khan, I.; Karim, N.; Abdel-Halim, H.; Ali, A. Phytochemical analysis and antidiabetic potential of Elaeagnus umbellata (Thunb.) in streptozotocin-induced diabetic rats: Pharmacological and computational approach. BMC Complement. Altern. Med. 2018, 18, 332. [Google Scholar] [CrossRef]
- Kim, J.-I.; Baek, H.-J.; Han, D.-W.; Yun, J.-A. Autumn olive (Elaeagnus umbellata Thunb.) berry reduces fasting and postprandial glucose levels in mice. Nutr. Res. Pract. 2019, 13, 11–16. [Google Scholar] [CrossRef]
- Nazir, N.; Zahoor, M.; Ullah, L.; Ezzeldin, E.; Mostafa, G.A.E. Curative effect of catechin isolated from Elaeagnus umbellata thunb. berries for diabetes and related complications in streptozotocin-induced diabetic rats model. Molecules 2020, 26, 137. [Google Scholar] [CrossRef]
- Kim, Y.; Lee, S.-b.; Cho, M.; Choe, S.; Jang, M. Indian almond (Terminalia catappa linn.) leaf extract extends lifespan by improving lipid metabolism and antioxidant activity dependent on AMPK signaling pathway in Caenorhabditis elegans under high-glucose-diet conditions. Antioxidants 2023, 13, 14. [Google Scholar] [CrossRef]
- Cho, M.; Kim, Y.; You, S.; Hwang, D.Y.; Jang, M. Chlorogenic acid of Cirsium japonicum resists oxidative stress caused by aging and prolongs healthspan via SKN-1/Nrf2 and DAF-16/FOXO in Caenorhabditis elegans. Metabolites 2023, 13, 224. [Google Scholar] [CrossRef]
- Wu, M.-C.; Hu, H.-T.; Yang, L.; Yang, L. Proteomic analysis of up-accumulated proteins associated with fruit quality during autumn olive (Elaeagnus umbellata) fruit ripening. J. Agric. Food Chem. 2011, 59, 577–583. [Google Scholar] [CrossRef]
- Sahan, Y.; Gocmen, D.; Cansev, A.; Celik, G.; Aydin, E.; Dundar, A.N.; Dulger, D. Chemical and techno-functional properties of flours from peeled and unpeeled oleaster (Elaeagnus angustifolia L.). J. Appl. Bot. Food Qual. 2015, 88, 34–41. [Google Scholar]
- Jang, M.; Kim, K.-H.; Kim, G.-H. Antioxidant capacity of thistle (Cirsium japonicum) in various drying methods and their protection effect on neuronal PC12 cells and Caenorhabditis elegans. Antioxidants 2020, 9, 200. [Google Scholar] [CrossRef]
- Jang, M.; Choi, H.-Y.; Kim, G.-H. Inhibitory effects of Orostachys malacophyllus var. iwarenge extracts on reactive oxygen species production and lipid accumulation during 3T3-L1 adipocyte differentiation. Food Sci. Biotechnol. 2019, 28, 227–236. [Google Scholar] [CrossRef]
- Cyboran, S.; Strugała, P.; Włoch, A.; Oszmiański, J.; Kleszczyńska, Y. Concentrated green tea supplement: Biological activity and molecular mechanisms. Life Sci. 2015, 126, 1–9. [Google Scholar] [CrossRef]
- Murase, T.; Nagasawa, A.; Suzuki, J.; Hase, T.; Tokimitsu, I. Beneficial effects of tea catechins on diet-induced obesity: Stimulation of lipid catabolism in the liver. Int. J. Obes. 2002, 26, 1459–1464. [Google Scholar] [CrossRef] [PubMed]
- Isemura, M. Catechin in human health and disease. Molecules 2019, 24, 528. [Google Scholar] [CrossRef] [PubMed]
- Jiang, Y.; Ding, S.; Li, F.; Zhang, C.; Sun-Waterhouse, D.; Chen, Y.; Li, D. Effects of (+)-catechin on the differentiation and lipid metabolism of 3T3-L1 adipocytes. J. Funct. Foods 2019, 62, 103558. [Google Scholar] [CrossRef]
- Chung, K.W. Advances in understanding of the role of lipid metabolism in aging. Cells 2021, 10, 880. [Google Scholar] [CrossRef] [PubMed]
- Kao, Y.-C.; Ho, P.-C.; Tu, Y.-K.; Jou, I.-M.; Tsai, K.-J. Lipids and Alzheimer’s disease. Int. J. Mol. Sci. 2020, 21, 1505. [Google Scholar] [CrossRef] [PubMed]
- Sato, N.; Morishita, R. The roles of lipid and glucose metabolism in modulation of β-amyloid, tau, and neurodegeneration in the pathogenesis of Alzheimer disease. Front. Aging Neurosci. 2015, 7, 199. [Google Scholar] [CrossRef] [PubMed]
- Luo, X.; Cheng, C.; Tan, Z.; Li, N.; Tang, M.; Yang, L.; Cao, Y. Emerging roles of lipid metabolism in cancer metastasis. Mol. Cancer 2017, 16, 76. [Google Scholar] [CrossRef]
- McGrath, C.M.; Young, S.P. Lipid and metabolic changes in rheumatoid arthritis. Curr. Rheumatol. Rep. 2015, 17, 57. [Google Scholar] [CrossRef]
- Mennes, E.; Dungan, C.M.; Frendo-Cumbo, S.; Williamson, D.L.; Wright, D.C. Aging-associated reductions in lipolytic and mitochondrial proteins in mouse adipose tissue are not rescued by metformin treatment. J. Gerontol. Ser. A Biomed. Sci. Med. Sci. 2014, 69, 1060–1068. [Google Scholar] [CrossRef]
- Camell, C.D.; Sander, J.; Spadaro, O.; Lee, A.; Nguyen, K.Y.; Wing, A.; Goldberg, E.L.; Youm, Y.-H.; Brown, C.W.; Elsworth, J. Inflammasome-driven catecholamine catabolism in macrophages blunts lipolysis during ageing. Nature 2017, 550, 119–123. [Google Scholar] [CrossRef]
- Szlejf, C.; Parra-Rodríguez, L.; Rosas-Carrasco, O. Osteosarcopenic obesity: Prevalence and relation with frailty and physical performance in middle-aged and older women. J. Am. Med. Dir. Assoc. 2017, 18, 733.e731–733.e735. [Google Scholar] [CrossRef]
- Gross, D.; Van Den Heuvel, A.; Birnbaum, M. The role of FoxO in the regulation of metabolism. Oncogene 2008, 27, 2320–2336. [Google Scholar] [CrossRef]
- Schreiber, R.; Xie, H.; Schweiger, M. Of mice and men: The physiological role of adipose triglyceride lipase (ATGL). Biochim. Et Biophys. Acta (BBA)—Mol. Cell Biol. Lipids 2019, 1864, 880–899. [Google Scholar] [CrossRef] [PubMed]
- Chakrabarti, P.; Kandror, K.V. FoxO1 controls insulin-dependent adipose triglyceride lipase (ATGL) expression and lipolysis in adipocytes. J. Biol. Chem. 2009, 284, 13296–13300. [Google Scholar] [CrossRef]
- Han, Y.; Hu, Z.; Cui, A.; Liu, Z.; Ma, F.; Xue, Y.; Liu, Y.; Zhang, F.; Zhao, Z.; Yu, Y. Post-translational regulation of lipogenesis via AMPK-dependent phosphorylation of insulin-induced gene. Nat. Commun. 2019, 10, 623. [Google Scholar] [CrossRef]
- Jang, M.; Choi, S.I. Schisandrin C isolated from Schisandra chinensis fruits inhibits lipid accumulation by regulating adipogenesis and lipolysis through AMPK signaling in 3T3-L1 adipocytes. J. Food Biochem. 2022, 46, e14454. [Google Scholar] [CrossRef]
- Kwon, J.Y.; Kershaw, J.; Chen, C.-Y.; Komanetsky, S.M.; Zhu, Y.; Guo, X.; Myer, P.R.; Applegate, B.; Kim, K.-H. Piceatannol antagonizes lipolysis by promoting autophagy-lysosome-dependent degradation of lipolytic protein clusters in adipocytes. J. Nutr. Biochem. 2022, 105, 108998. [Google Scholar] [CrossRef]
- Lee, J.H.; Kong, J.; Jang, J.Y.; Han, J.S.; Ji, Y.; Lee, J.; Kim, J.B. Lipid droplet protein LID-1 mediates ATGL-1-dependent lipolysis during fasting in Caenorhabditis elegans. Mol. Cell Biol. 2014, 34, 4165–4176. [Google Scholar] [CrossRef]
- Stiernagle, T. Maintenance of C. elegans. In Community Worm Book: Worm Book; TCeR, Ed.; Beijing Institute of Technology Press: Beijing, China, 2006. [Google Scholar]
- OECD. Report of the OECD Seminar on Risk Reduction through Prevention, Detection and Control of the Illegal International Trade in Agricultural Pesticides; OECD: Paris, France, 2014. [Google Scholar] [CrossRef]
- Gomes, W.F.; França, F.R.M.; Denadai, M.; Andrade, J.K.S.; da Silva Oliveira, E.M.; de Brito, E.S.; Rodrigues, S.; Narain, N. Effect of freeze-and spray-drying on physico-chemical characteristics, phenolic compounds and antioxidant activity of papaya pulp. J. Food Sci. Technol. 2018, 55, 2095–2102. [Google Scholar] [CrossRef]




{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Samples | TPC (mg GAE/100 g) | DPPH (mg AA/100 g) |
|---|---|---|
| FAOB | 258.23 1.00 | 3842.23 294.91 |
| SAOB | 63.02 2.00 | 3151.78 395.08 |
| p-value * | 0.05 | 0.03 |
| Samples | Catechin (mg/100 g) |
|---|---|
| FAOB | 154.02 5.29 |
| SAOB | 27.53 3.05 |
| p-value * | 0.002 |
| HPLC (UltiMate 3000, Thermo Scientific, Sunnyvale, CA, USA) | |
|---|---|
| Column | Waters symmetry C18 column (4.6 × 150 mm, 5 µm) |
| Mobile phase | (A) 0.1% (v/v) aqueous phosphoric acid (B) Acetonitrile (Duksan, Ansan-si, Gyeonggi-do, Republic of Korea) Gradient method: 50–90% solvent B for 1 min, 90–85% solvent B for 2 min, 85–80% solvent B for 2 min, 80–70% solvent B for 3 min, 70–30% solvent B for 3 min, and a linear step from 30 to 10% solvent B for 9 min. |
| Flow rate | 0.8 mL/min |
| Inject volume | 20 μL |
| Detector | Diode array detector (DAD) |
| Wavelength | 280 nm |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Kim, Y.; Nam, S.; Lim, J.; Jang, M. Autumn Olive (Elaeagnus umbellata Thunb.) Berries Improve Lipid Metabolism and Delay Aging in Middle-Aged Caenorhabditis elegans. Int. J. Mol. Sci. 2024, 25, 3418. https://doi.org/10.3390/ijms25063418
Kim Y, Nam S, Lim J, Jang M. Autumn Olive (Elaeagnus umbellata Thunb.) Berries Improve Lipid Metabolism and Delay Aging in Middle-Aged Caenorhabditis elegans. International Journal of Molecular Sciences. 2024; 25(6):3418. https://doi.org/10.3390/ijms25063418
Chicago/Turabian StyleKim, Yebin, Seonghyeon Nam, Jongbin Lim, and Miran Jang. 2024. "Autumn Olive (Elaeagnus umbellata Thunb.) Berries Improve Lipid Metabolism and Delay Aging in Middle-Aged Caenorhabditis elegans" International Journal of Molecular Sciences 25, no. 6: 3418. https://doi.org/10.3390/ijms25063418
APA StyleKim, Y., Nam, S., Lim, J., & Jang, M. (2024). Autumn Olive (Elaeagnus umbellata Thunb.) Berries Improve Lipid Metabolism and Delay Aging in Middle-Aged Caenorhabditis elegans. International Journal of Molecular Sciences, 25(6), 3418. https://doi.org/10.3390/ijms25063418