
Transcriptional Changes in Radiation-Induced Lung Injury: A Comparative Analysis of Two Radiation Doses for Preclinical Research

 , , , ,
, , , ,
Abstract
1. Introduction
2. Results
2.1. Transcriptomic Analysis of 75 Gy-Based Small-Animal Model
2.2. Lowest Single Irradiation Dose to Induce RILF within 2 Months
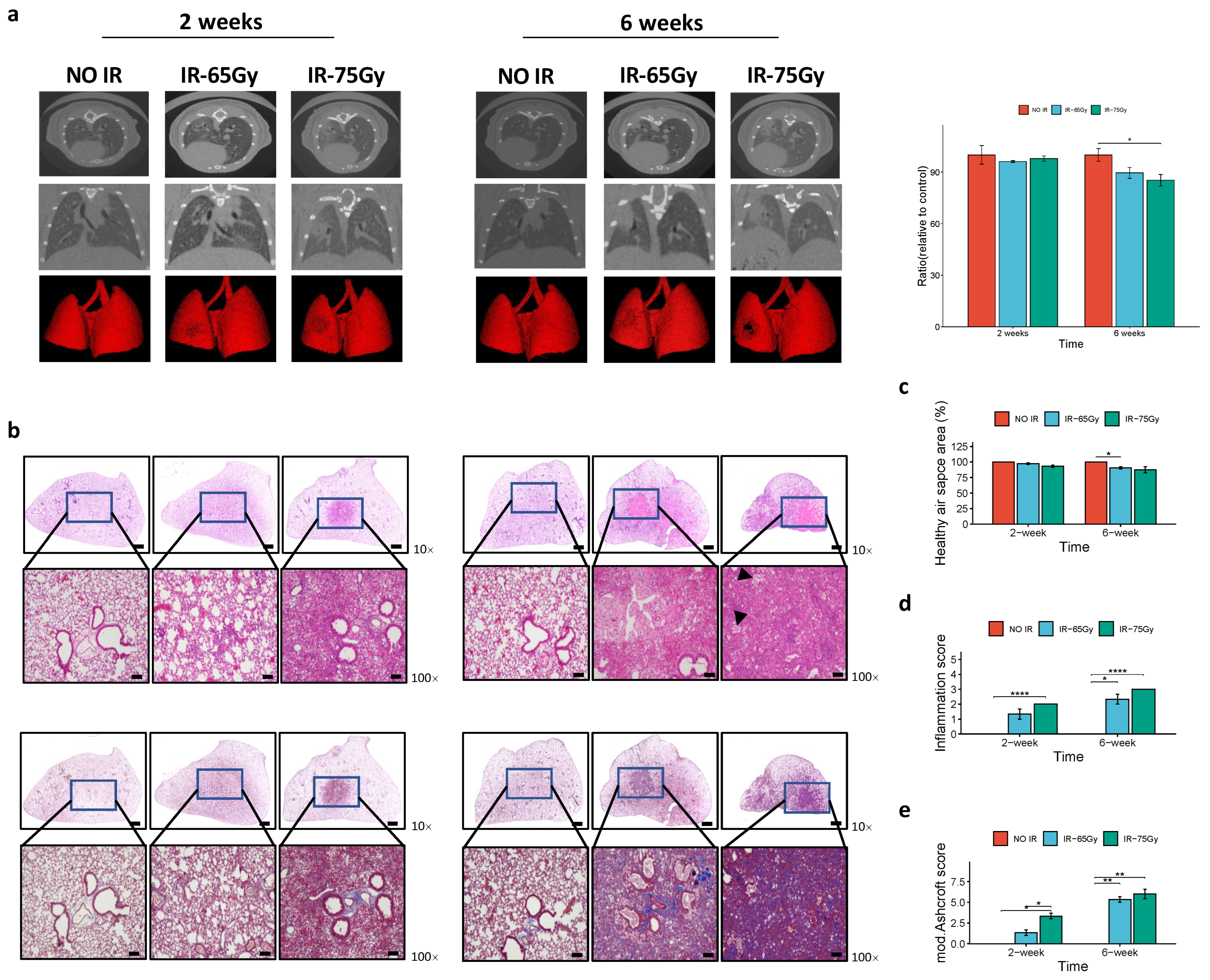
2.3. Comparative Imaging and Histopathological Analysis of 65 Gy and 75 Gy Models
2.4. Comparative Transcriptomic Analysis between 65 Gy and 75 Gy Models
2.5. Validation of Similarities and Differences between 65 Gy and 75 Gy Models by Reverse Transcription Quantitative PCR (RT-qPCR)
3. Discussion
4. Materials and Methods
4.1. Animal Experiments
4.2. Micro-CT Analysis
4.3. Histopathological Analysis
4.4. Extraction, Purification, and Sequencing of RNA
4.5. Whole-Genome Transcriptomic Analysis
4.6. Identification of DEGs
4.7. Expression and DEG Data Interpretation and Analysis
4.8. Functional Enrichment, Network Analysis, and PPI Module Construction
4.9. RT-qPCR
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Kong, F.-M.; Hayman, J.A.; Griffith, K.A.; Kalemkerian, G.P.; Arenberg, D.; Lyons, S.; Turrisi, A.; Lichter, A.; Fraass, B.; Eisbruch, A.; et al. Final Toxicity Results of a Radiation-Dose Escalation Study in Patients with Non–Small-Cell Lung Cancer (NSCLC): Predictors for Radiation Pneumonitis and Fibrosis. Int. J. Radiat. Oncol. Biol. Phys. 2006, 65, 1075–1086. [Google Scholar] [CrossRef] [PubMed]
- Vallée, A.; Lecarpentier, Y.; Guillevin, R.; Vallée, J.N. Interactions between TGF-Β1, Canonical WNT/β-Catenin Pathway and PPAR γ in Radiation-Induced Fibrosis. Oncotarget 2017, 8, 90579–90604. [Google Scholar] [CrossRef] [PubMed]
- Straub, J.M.; New, J.; Hamilton, C.D.; Lominska, C.; Shnayder, Y.; Thomas, S.M. Radiation-Induced Fibrosis: Mechanisms and Implications for Therapy. J. Cancer Res. Clin. Oncol. 2015, 141, 1985–1994. [Google Scholar] [CrossRef] [PubMed]
- Garcia, A.N.; Casanova, N.G.; Valera, D.G.; Sun, X.; Song, J.H.; Kempf, C.L.; Moreno-Vinasco, L.; Burns, K.; Bermudez, T.; Valdez, M.; et al. Involvement of ENAMPT/TLR4 Signaling in Murine Radiation Pneumonitis: Protection by ENAMPT Neutralization. Transl. Res. 2022, 239, 44. [Google Scholar] [CrossRef] [PubMed]
- Beach, T.A.; Johnston, C.J.; Groves, A.M.; Williams, J.P.; Finkelstein, J.N. Radiation Induced Pulmonary Fibrosis as a Model of Progressive Fibrosis: Contributions of DNA Damage, Inflammatory Response and Cellular Senescence Genes. Exp. Lung Res. 2017, 43, 134–149. [Google Scholar] [CrossRef] [PubMed]
- Jin, H.; Kang, G.Y.; Jeon, S.; Kim, J.M.; Park, Y.N.; Cho, J.; Lee, Y.S. Identification of Molecular Signatures Involved in Radiation-Induced Lung Fibrosis. J. Mol. Med. 2019, 97, 37–47. [Google Scholar] [CrossRef]
- Kim, H.; Park, S.H.; Han, S.Y.; Lee, Y.S.; Cho, J.; Kim, J.M. LXA4-FPR2 Signaling Regulates Radiation-Induced Pulmonary Fibrosis via Crosstalk with TGF-β/Smad Signaling. Cell Death Dis. 2020, 11, 653. [Google Scholar] [CrossRef]
- Shin, D.; Lee, G.; Lee, S.; Park, S.; Jung, K.H.; Lee, J.H.; Lee, J.M.; Kim, J.Y.; Cho, J.; Bae, H. Adenovirus-Mediated Foxp3 Expression in Lung Epithelial Cells Ameliorates Acute Radiation-Induced Pneumonitis in Mice. Gene Ther. 2017, 24, 104–112. [Google Scholar] [CrossRef] [PubMed]
- Li, Y.; Zou, L.; Yang, X.; Chu, L.; Ni, J.; Chu, X.; Guo, T.; Zhu, Z. Identification of LncRNA, MicroRNA, and MRNA-Associated CeRNA Network of Radiation-Induced Lung Injury in a Mice Model. Dose Response 2019, 17, 1–7. [Google Scholar] [CrossRef]
- Zhang, T.; Cheng, G.; Sun, L.; Deng, L.; Wang, X.; Bi, N. Transcriptome Alteration Spectrum in Rat Lung Induced by Radiotherapy. Sci. Rep. 2019, 9, 19701. [Google Scholar] [CrossRef]
- Thakur, P.; DeBo, R.; Dugan, G.O.; Bourland, J.D.; Michalson, K.T.; Olson, J.D.; Register, T.C.; Kock, N.D.; Cline, J.M. Clinicopathologic and Transcriptomic Analysis of Radiation-Induced Lung Injury in Nonhuman Primates. Int. J. Radiat. Oncol. Biol. Phys. 2021, 111, 249–259. [Google Scholar] [CrossRef]
- Liu, X.; Shao, C.; Fu, J. Promising Biomarkers of Radiation-Induced Lung Injury: A Review. Biomedicines 2021, 9, 1181. [Google Scholar] [CrossRef]
- Arakawa, S.; Suzukawa, M.; Watanabe, K.; Kobayashi, K.; Matsui, H.; Nagai, H.; Nagase, T.; Ohta, K. Secretory Immunoglobulin A Induces Human Lung Fibroblasts to Produce Inflammatory Cytokines and Undergo Activation. Clin. Exp. Immunol. 2019, 195, 287–301. [Google Scholar] [CrossRef] [PubMed]
- Kim, J.Y.; Shin, D.; Lee, G.; Kim, J.M.; Kim, D.; An, Y.M.; Yoo, B.R.; Chang, H.; Kim, M.; Cho, J.; et al. Standardized Herbal Formula PM014 Inhibits Radiation-Induced Pulmonary Inflammation in Mice. Sci. Rep. 2017, 7, 45001. [Google Scholar] [CrossRef] [PubMed]
- Hong, Z.Y.; Lee, C.G.; Shim, H.S.; Lee, E.J.; Song, K.H.; Choi, B.W.; Cho, J.; Story, M.D. Time, Dose, and Volume Responses in a Mouse Pulmonary Injury Model Following Ablative Irradiation. Lung 2016, 194, 81–90. [Google Scholar] [CrossRef] [PubMed]
- Romero, F.; Shah, D.; Duong, M.; Penn, R.B.; Fessler, M.B.; Madenspacher, J.; Stafstrom, W.; Kavuru, M.; Lu, B.; Kallen, C.B.; et al. A Pneumocyte-Macrophage Paracrine Lipid Axis Drives the Lung toward Fibrosis. Am. J. Respir. Cell Mol. Biol. 2015, 53, 74–86. [Google Scholar] [CrossRef] [PubMed]
- Huang, S.K.; Peters-Golden, M. Eicosanoid Lipid Mediators in Fibrotic Lung Diseases: Ready for Prime Time? Chest 2008, 133, 1442. [Google Scholar] [CrossRef] [PubMed]
- Bärnthaler, T.; Theiler, A.; Zabini, D.; Trautmann, S.; Stacher-Priehse, E.; Lanz, I.; Klepetko, W.; Sinn, K.; Flick, H.; Scheidl, S.; et al. Inhibiting Eicosanoid Degradation Exerts Antifibrotic Effects in a Pulmonary Fibrosis Mouse Model and Human Tissue. J. Allergy Clin. Immunol. Pract. 2020, 145, 818–833.e11. [Google Scholar] [CrossRef] [PubMed]
- Geng, J.; Liu, Y.; Dai, H.; Wang, C. Fatty Acid Metabolism and Idiopathic Pulmonary Fibrosis. Front. Physiol. 2022, 12, 794629. [Google Scholar] [CrossRef]
- Huang, Y.; Zhang, W.; Yu, F.; Gao, F. The Cellular and Molecular Mechanism of Radiation-Induced Lung Injury. Med. Sci. Monit. 2017, 23, 3446–3450. [Google Scholar] [CrossRef]
- Nguyen, T.M.D. Adiponectin: Role in Physiology and Pathophysiology. Int. J. Prev. Med. 2020, 11, 136. [Google Scholar] [CrossRef] [PubMed]
- Wright, J.M.; Merlo, C.A.; Reynolds, J.B.; Zeitlin, P.L.; Garcia, J.G.N.; Guggino, W.B.; Boyle, M.P. Respiratory Epithelial Gene Expression in Patients with Mild and Severe Cystic Fibrosis Lung Disease. Am. J. Respir. Cell Mol. Biol. 2006, 35, 327–336. [Google Scholar] [CrossRef] [PubMed]
- Palanisamy, K.; Raman, R.; Sulochana, K.N.; Chidambaram, S. Adiponectin: A Potential Candidate for Treating Fibrosis in Posterior Segment of the Eye. Med. Hypotheses 2019, 123, 9–12. [Google Scholar] [CrossRef] [PubMed]
- Yao, R.; Cao, Y.; He, Y.R.; Lau, W.B.; Zeng, Z.; Liang, Z.A. Adiponectin Attenuates Lung Fibroblasts Activation and Pulmonary Fibrosis Induced by Paraquat. PLoS ONE 2015, 10, e0125169. [Google Scholar] [CrossRef] [PubMed]
- Surendar, J.; Frohberger, S.J.; Karunakaran, I.; Schmitt, V.; Stamminger, W.; Neumann, A.L.; Wilhelm, C.; Hoerauf, A.; Hübner, M.P. Adiponectin Limits IFN-γ and Il-17 Producing CD4 T Cells in Obesity by Restraining Cell Intrinsic Glycolysis. Front. Immunol. 2019, 10, 478580. [Google Scholar] [CrossRef] [PubMed]
- Kursawe, R.; Santoro, N. Metabolic Syndrome in Pediatrics. Adv. Clin. Chem. 2014, 65, 91–142. [Google Scholar] [CrossRef] [PubMed]
- Chabowski, A.; Górski, J. Muscle Lipid Metabolism. In Muscle and Exercise Physiology; Academic Press: Cambridge, MA, USA, 2018; pp. 271–284. ISBN 9780128145944. [Google Scholar]
- McManus, B.; Knight, D. Long-Chain Fatty Acids Are Bad in IPF, or Are They? Respirology 2021, 26, 220–221. [Google Scholar] [CrossRef] [PubMed]
- Zhang, Z.; Kruglikov, I.; Zhao, S.; Zi, Z.; Gliniak, C.M.; Li, N.; Wang, M.-y.; Zhu, Q.; Kusminski, C.M.; Scherer, P.E. Dermal Adipocytes Contribute to the Metabolic Regulation of Dermal Fibroblasts. Exp. Dermatol. 2021, 30, 102–111. [Google Scholar] [CrossRef]
- Sohn, J.H.; Lee, Y.K.; Han, J.S.; Jeon, Y.G.; Kim, J.I.; Choe, S.S.; Kim, S.J.; Yoo, H.J.; Kim, J.B. Perilipin 1 (Plin1) Deficiency Promotes Inflammatory Responses in Lean Adipose Tissue through Lipid Dysregulation. J. Biol. Chem. 2018, 293, 13974–13988. [Google Scholar] [CrossRef]
- Truong, N.; Love-Rutledge, S.; Lydic, T.; Olson, L.K. Effect of Interferon Gamma on Neutral Lipid Levels, Lipid Droplet Formation, and Antiviral Responses in Pancreatic Islets and INS-1 β Cells. FASEB J. 2019, 33, 654.11. [Google Scholar] [CrossRef]
- Park, S.H.; Kim, J.Y.; Kim, J.M.; Yoo, B.R.; Han, S.Y.; Jung, Y.J.; Bae, H.; Cho, J. PM014 Attenuates Radiation-Induced Pulmonary Fibrosis via Regulating NF-KB and TGF-B1/NOX4 Pathways. Sci. Rep. 2020, 10, 16112. [Google Scholar] [CrossRef] [PubMed]
- Zhang, J.; Cui, R.; Feng, Y.; Gao, W.; Bi, J.; Li, Z.; Liu, C. Serotonin Exhibits Accelerated Bleomycin-Induced Pulmonary Fibrosis through TPH1 Knockout Mouse Experiments. Mediators Inflamm. 2018, 2018, 7967868. [Google Scholar] [CrossRef] [PubMed]
- Ashcroft, T.; Simpson, J.M.; Timbrell, V. Simple Method of Estimating Severity of Pulmonary Fibrosis on a Numerical Scale. J. Clin. Pathol. 1988, 41, 467–470. [Google Scholar] [CrossRef] [PubMed]
- Hübner, R.H.; Gitter, W.; el Mokhtari, N.E.; Mathiak, M.; Both, M.; Bolte, H.; Freitag-Wolf, S.; Bewig, B. Standardized Quantification of Pulmonary Fibrosis in Histological Samples. Biotechniques 2008, 44, 507–517. [Google Scholar] [CrossRef] [PubMed]
- Chen, Y.; Yu, Q.; Xu, C.B. A Convenient Method for Quantifying Collagen Fibers in Atherosclerotic Lesions by Imagej Software. Int. J. Clin. Exp. Med. 2017, 10, 14904–14910. [Google Scholar]
- Robinson, M.D.; McCarthy, D.J.; Smyth, G.K. EdgeR: A Bioconductor Package for Differential Expression Analysis of Digital Gene Expression Data. Bioinformatics 2010, 26, 139. [Google Scholar] [CrossRef]
- Benjamini, Y.; Hochberg, Y. Controlling the False Discovery Rate: A Practical and Powerful Approach to Multiple Testing. J. R Stat. Soc. Ser. B Methodol. 1995, 57, 289–300. [Google Scholar] [CrossRef]
- Rohart, F.; Gautier, B.; Singh, A.; Lê Cao, K.A. MixOmics: An R Package for ‘omics Feature Selection and Multiple Data Integration. PLoS Comput. Biol. 2017, 13, e1005752. [Google Scholar] [CrossRef] [PubMed]
- Warnes, G.R.; Bolker, B.; Bonebakker, L.; Gentleman, R.; Liaw, W.H.A.; Lumley, T.; Maechler, M.; Magnusson, A.; Moeller, S.; Schwartz, M.; et al. Gplots: Various R Programming Tools for Plotting Data. R Package Version 3.0.1.1. 2019. Available online: http://CRAN.R-project.org/package=gplots (accessed on 1 February 2023).
- Chen, H.; Boutros, P.C. VennDiagram: A Package for the Generation of Highly-Customizable Venn and Euler Diagrams in R. BMC Bioinform. 2011, 12, 35. [Google Scholar] [CrossRef]
- Tripathi, S.; Pohl, M.O.; Zhou, Y.; Rodriguez-Frandsen, A.; Wang, G.; Stein, D.A.; Moulton, H.M.; Dejesus, P.; Che, J.; Mulder, L.C.F.; et al. Meta- and Orthogonal Integration of Influenza “OMICs” Data Defines a Role for UBR4 in Virus Budding. Cell Host Microbe 2015, 18, 723–735. [Google Scholar] [CrossRef]
- Giurgiu, M.; Reinhard, J.; Brauner, B.; Dunger-Kaltenbach, I.; Fobo, G.; Frishman, G.; Montrone, C.; Ruepp, A. CORUM: The Comprehensive Resource of Mammalian Protein Complexes—2019. Nucleic Acids Res. 2019, 47, D559–D563. [Google Scholar] [CrossRef]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Time | Group | Replicate | RNA-Seq Data | Alignment Data | ||||
|---|---|---|---|---|---|---|---|---|
| Raw Reads | Trimmed Reads | Raw Reads vs. Trimmed Reads | GC Content (%) | Total Mapped | Total Unmapped | |||
| 2 weeks | Control | 1 | 66,847,010 | 66,073,590 | 98.84 | 48.79 | 65,182,623 (98.65%) | 890,967 (1.35%) |
| 2 | 63,021,390 | 62,284,648 | 98.83 | 49.34 | 61,617,863 (98.93%) | 666,785 (1.07%) | ||
| 3 | 72,993,584 | 72,081,246 | 98.75 | 49.53 | 71,293,386 (98.91%) | 787,860 (1.09%) | ||
| 65 Gy | 1 | 79,873,572 | 78,875,404 | 98.75 | 49.60 | 78,075,192 (98.99%) | 800,212 (1.01%) | |
| 2 | 82,604,278 | 81,604,686 | 98.79 | 49.40 | 80,581,993 (98.75%) | 1,022,693 (1.25%) | ||
| 3 | 73,278,816 | 72,414,130 | 98.82 | 49.56 | 71,653,924 (98.95%) | 760,206 (1.05%) | ||
| 75 Gy | 1 | 77,172,286 | 76,155,556 | 98.68 | 49.17 | 75,122,624 (98.64%) | 1,032,932 (1.36%) | |
| 2 | 89,558,428 | 88,540,584 | 98.86 | 49.21 | 87,557,242 (98.89%) | 983,342 (1.11%) | ||
| 3 | 76,282,682 | 75,317,680 | 98.73 | 49.49 | 74,579,006 (99.02%) | 738,674 (0.98%) | ||
| 6 weeks | Control | 1 | 85,460,960 | 84,448,858 | 98.82 | 49.27 | 83,548,257 (98.93%) | 900,601 (1.07%) |
| 2 | 77,140,466 | 76,145,872 | 98.71 | 49.44 | 75,236,435 (98.81%) | 909,437 (1.19%) | ||
| 3 | 78,125,372 | 76,988,642 | 98.54 | 49.36 | 76,054,270 (98.79%) | 934,372 (1.21%) | ||
| 65 Gy | 1 | 80,695,798 | 79,595,842 | 98.64 | 49.56 | 78,625,912 (98.78%) | 969,930 (1.22%) | |
| 2 | 60,530,124 | 59,600,646 | 98.46 | 50.35 | 58,558,124 (98.25%) | 1,042,522 (1.75%) | ||
| 3 | 77,818,676 | 76,762,422 | 98.64 | 49.33 | 75,816,780 (98.77%) | 945,642 (1.23%) | ||
| 75 Gy | 1 | 82,035,406 | 80,945,774 | 98.67 | 49.26 | 79,905,992 (98.72%) | 1,039,782 (1.28%) | |
| 2 | 72,804,658 | 72,064,966 | 98.98 | 48.55 | 71,335,395 (98.99%) | 729,571 (1.01%) | ||
| 3 | 73,353,186 | 72,494,140 | 98.83 | 48.75 | 71,765,561 (98.99%) | 728,579 (1.01%) | ||
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Farh, M.E.-A.; Kim, H.-J.; Kim, S.-Y.; Lee, J.-H.; Lee, H.; Cui, R.; Han, S.; Kim, D.W.; Park, S.; Lee, Y.-J.; et al. Transcriptional Changes in Radiation-Induced Lung Injury: A Comparative Analysis of Two Radiation Doses for Preclinical Research. Int. J. Mol. Sci. 2024, 25, 3766. https://doi.org/10.3390/ijms25073766
Farh ME-A, Kim H-J, Kim S-Y, Lee J-H, Lee H, Cui R, Han S, Kim DW, Park S, Lee Y-J, et al. Transcriptional Changes in Radiation-Induced Lung Injury: A Comparative Analysis of Two Radiation Doses for Preclinical Research. International Journal of Molecular Sciences. 2024; 25(7):3766. https://doi.org/10.3390/ijms25073766
Chicago/Turabian StyleFarh, Mohamed El-Agamy, Hyun-Jin Kim, Sang-Yeon Kim, Jae-Hee Lee, Hajeong Lee, Ronglan Cui, Soorim Han, Dong Wook Kim, Sunjoo Park, Yoon-Jin Lee, and et al. 2024. "Transcriptional Changes in Radiation-Induced Lung Injury: A Comparative Analysis of Two Radiation Doses for Preclinical Research" International Journal of Molecular Sciences 25, no. 7: 3766. https://doi.org/10.3390/ijms25073766
APA StyleFarh, M. E.-A., Kim, H.-J., Kim, S.-Y., Lee, J.-H., Lee, H., Cui, R., Han, S., Kim, D. W., Park, S., Lee, Y.-J., Lee, Y.-S., Sohn, I., & Cho, J. (2024). Transcriptional Changes in Radiation-Induced Lung Injury: A Comparative Analysis of Two Radiation Doses for Preclinical Research. International Journal of Molecular Sciences, 25(7), 3766. https://doi.org/10.3390/ijms25073766