
Eucommia ulmoides Leaves Alleviate Cognitive Dysfunction in Dextran Sulfate Sodium (DSS)-Induced Colitis Mice through Regulating JNK/TLR4 Signaling Pathway
 , ,
, ,  ,
,
Abstract
1. Introduction
2. Results
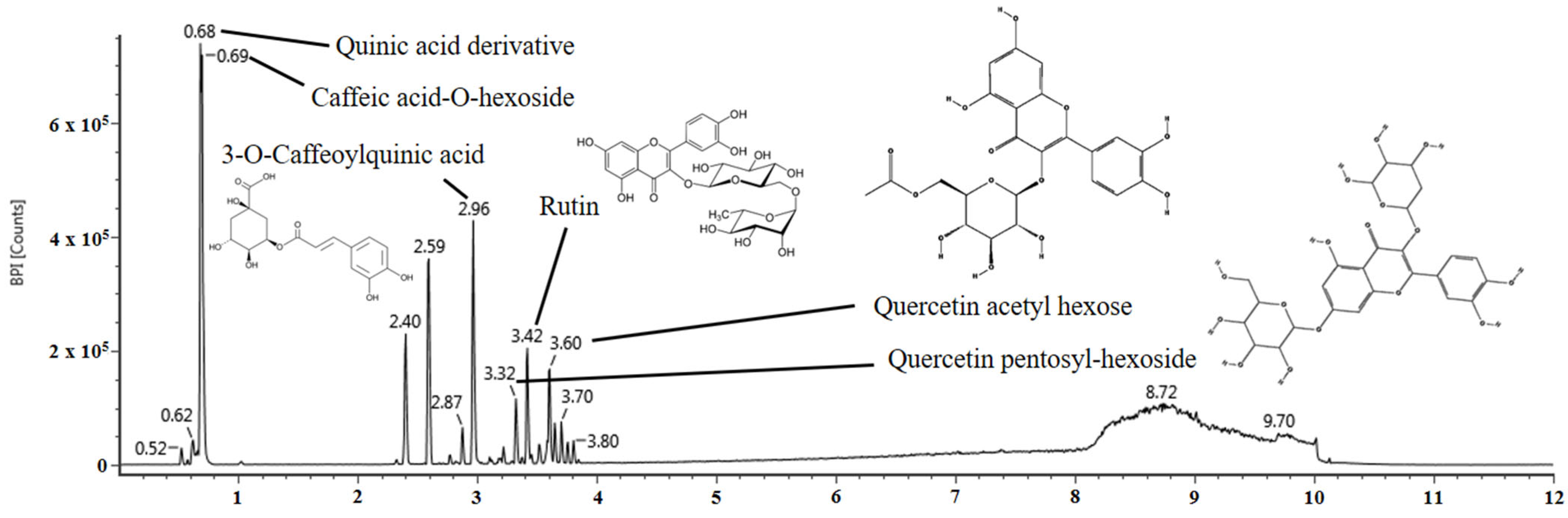
2.1. Identification of the Physiological Compound
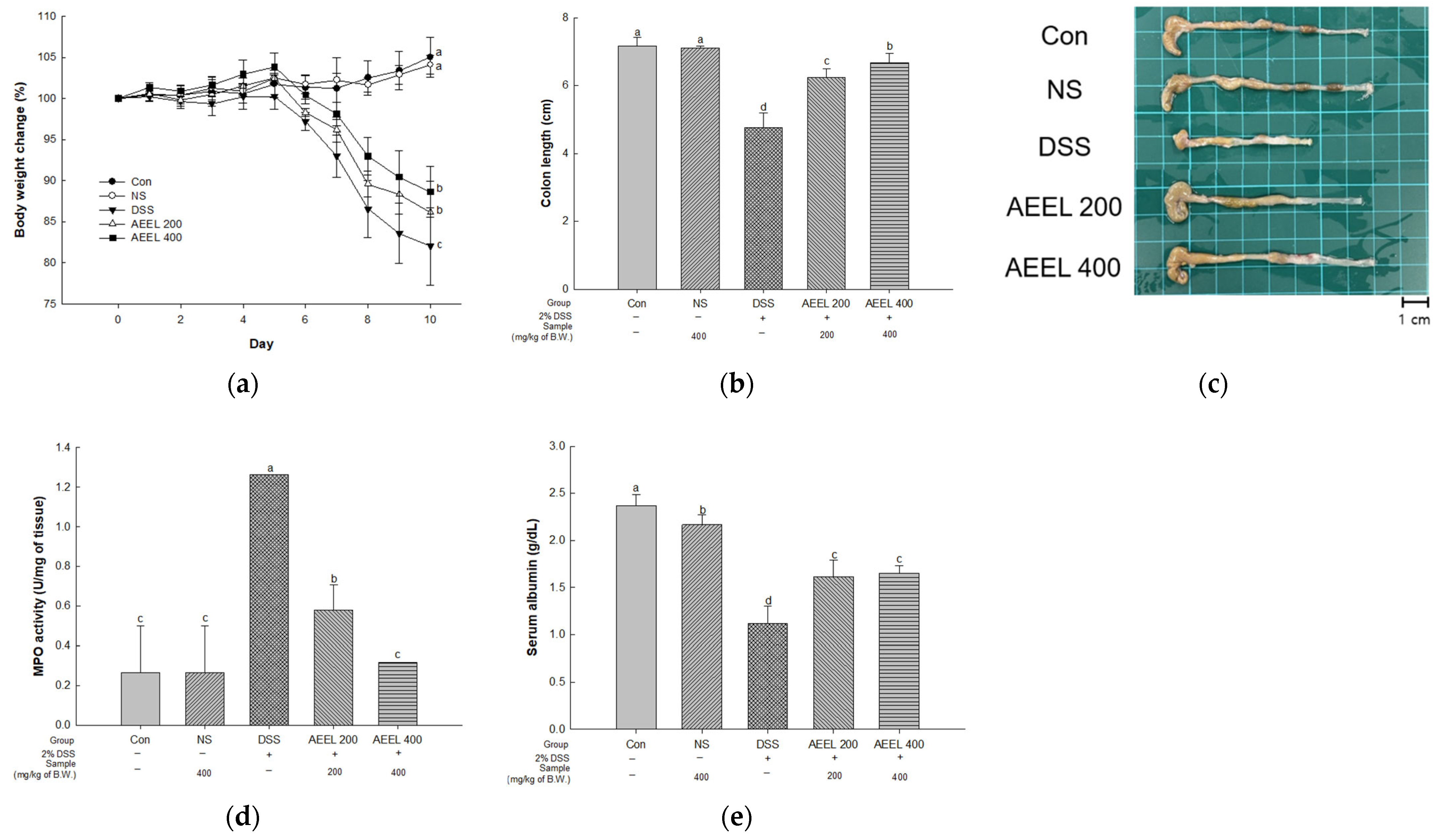
2.2. Alleviative Effect of AEEL on DSS-Induced Symptoms
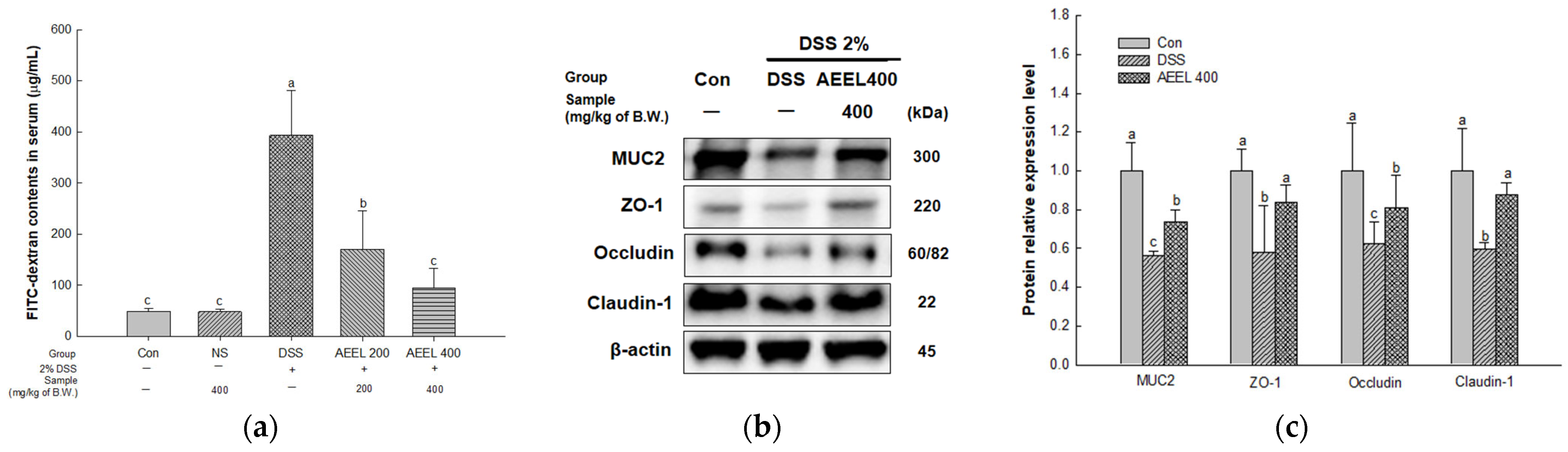
2.3. Alleviative Effect of AEEL on Intestinal Permeability Dysfunction
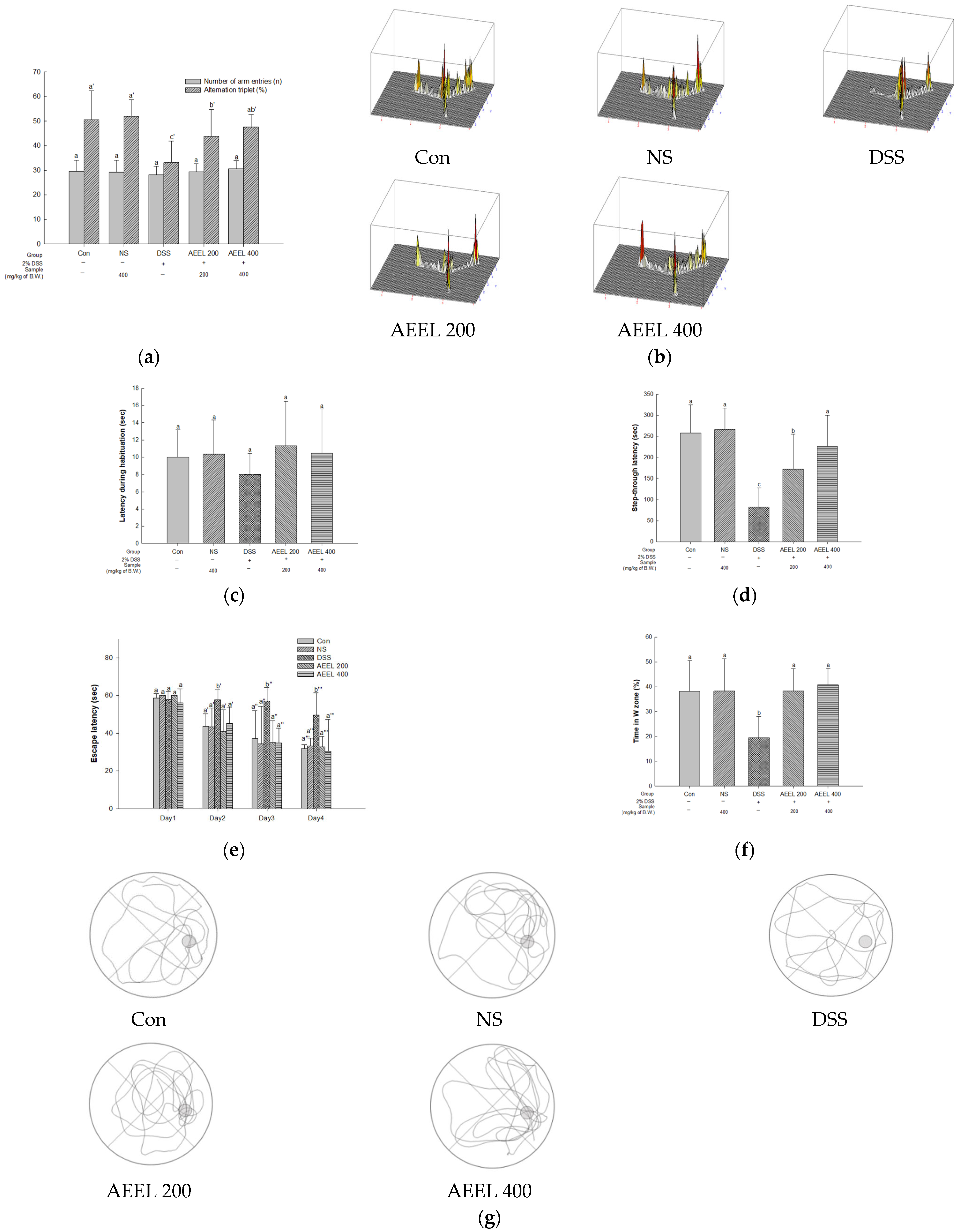
2.4. Animal Behavioral Tests
2.5. SCFAs in Feces
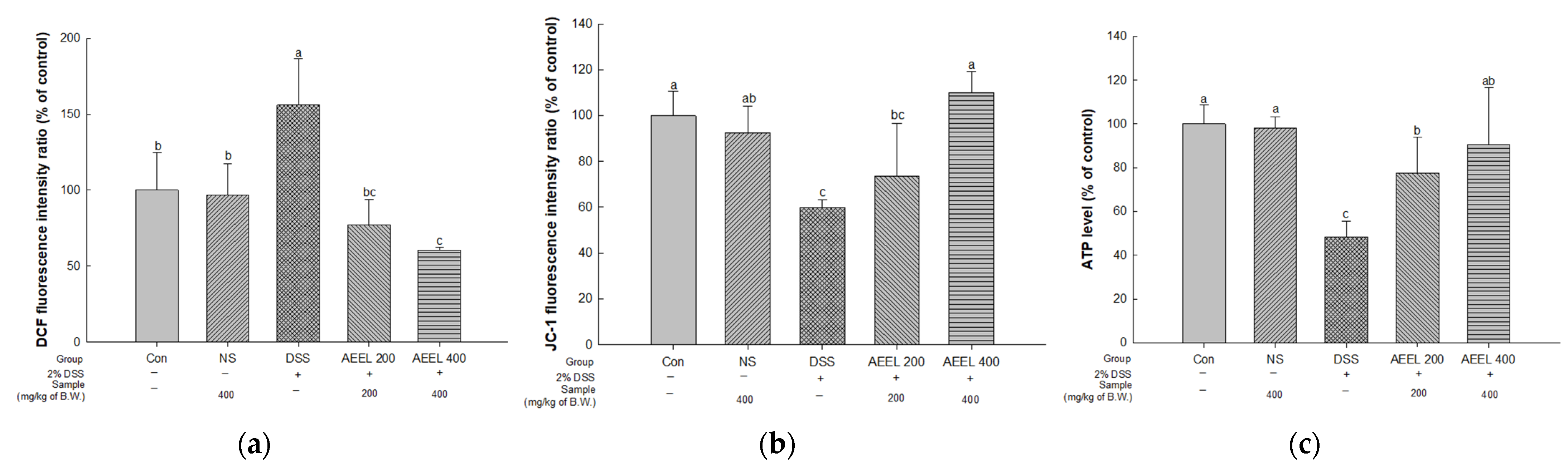
2.6. Alleviative Effect of AEEL on Mitochondrial Dysfunction in the Brain
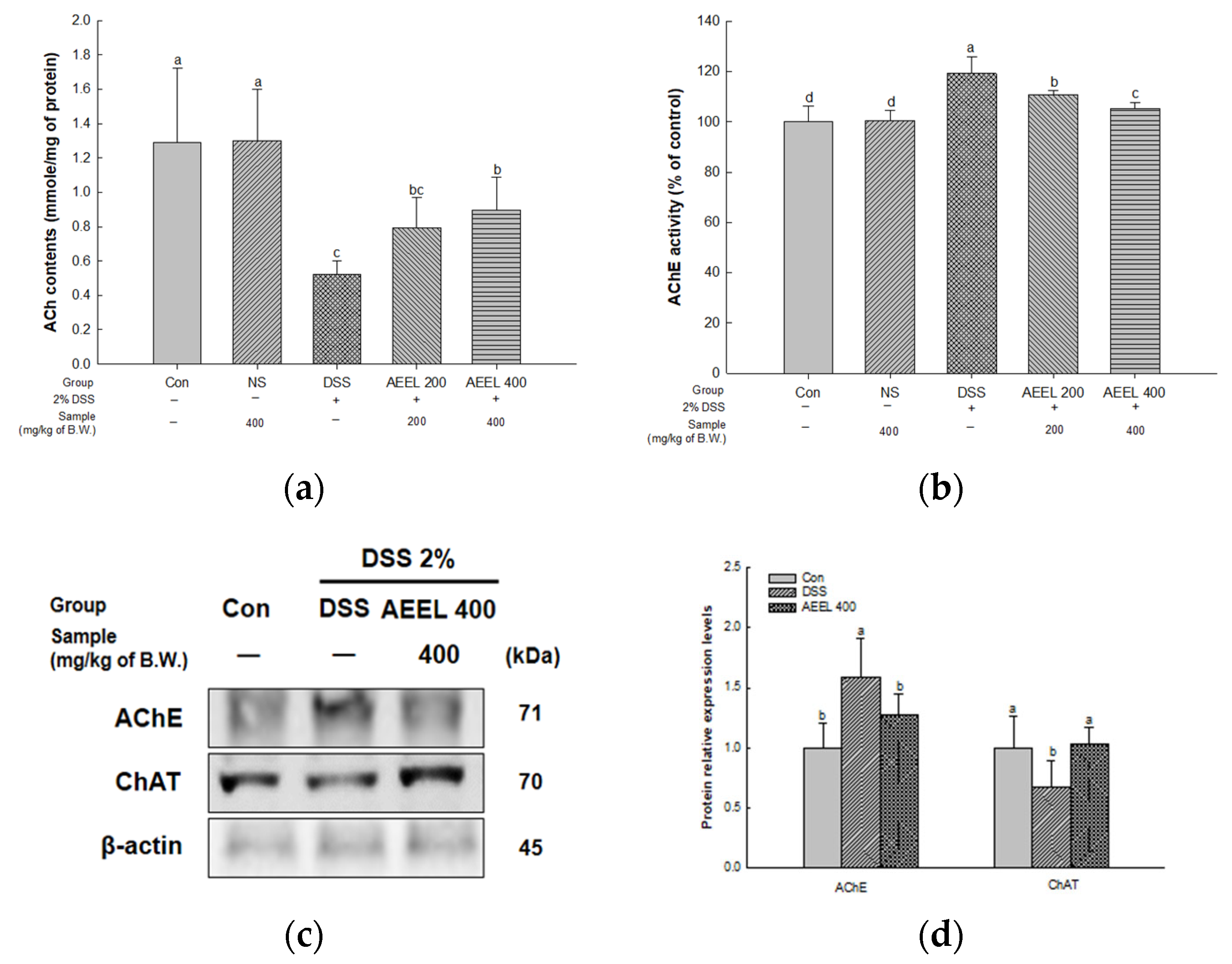
2.7. Alleviative Effect of AEEL on Cholinergic Dysfunction in Brain
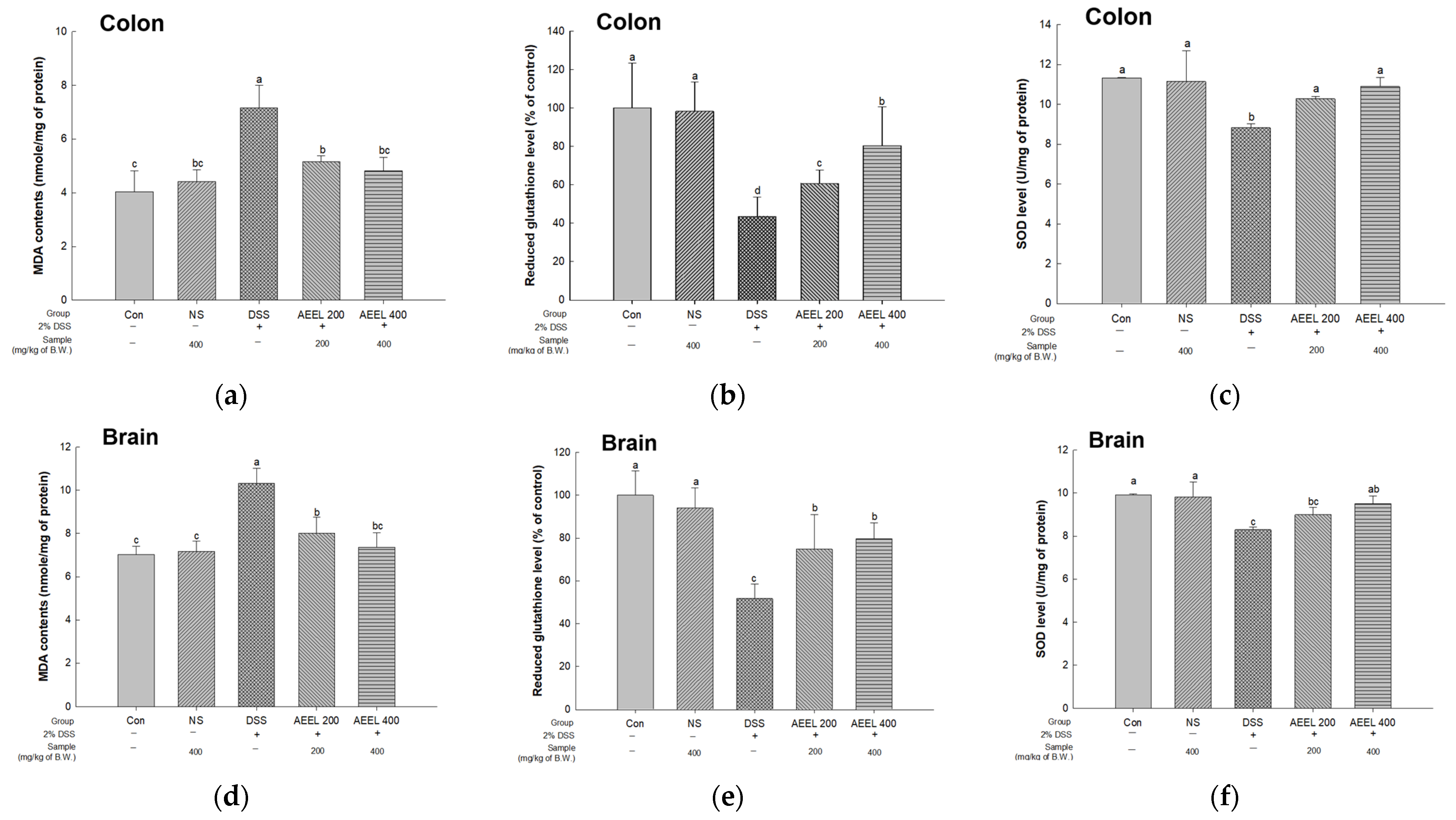
2.8. Alleviative Effect of AEEL on Oxidative Stress
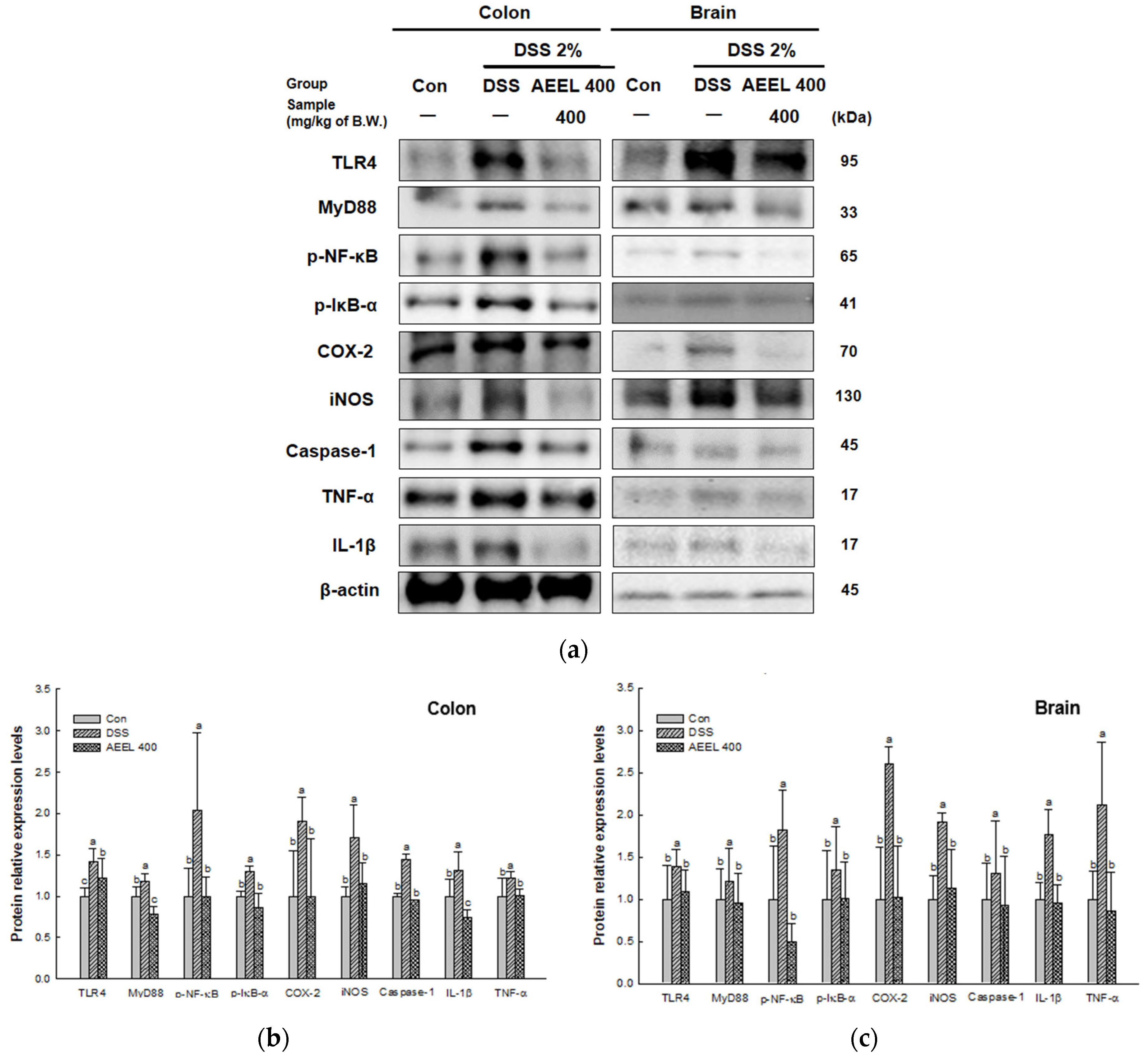
2.9. Alleviative Effect of AEEL on Inflammation
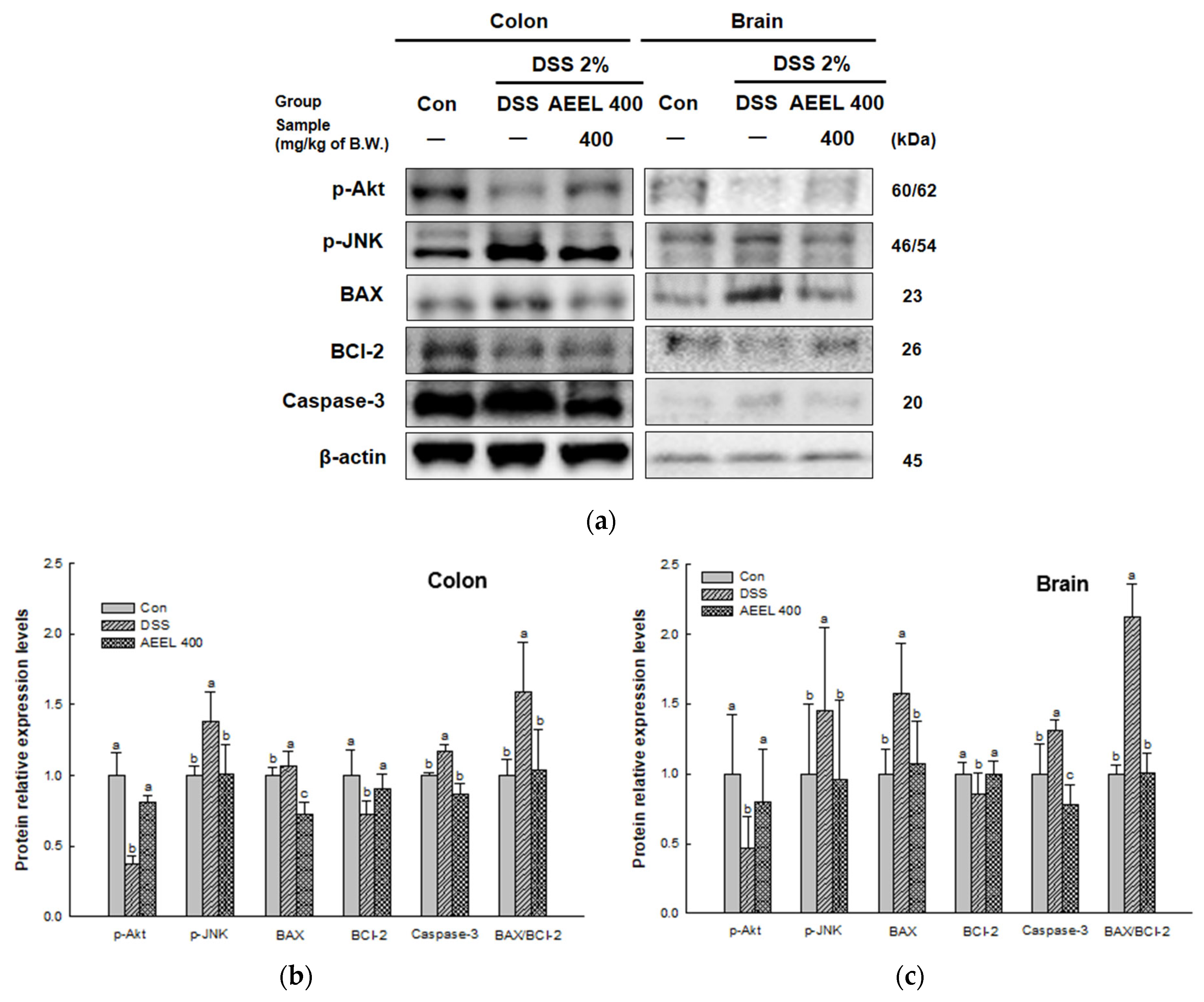
2.10. Alleviative Effect of AEEL on Apoptosis
3. Discussion
4. Materials and Methods
4.1. Chemicals
4.2. Preparation of Aqueous Extracts of E. ulmoides Leaves (AEEL)
4.3. UPLC Q-TOF/MSE
4.4. Animal Experiment Designs
4.5. MPO Activity in the Colon
4.6. Albumin Measurement in Serum
4.7. Fluorescein Isothiocyanate (FITC)-Dextran Contents in Serum
4.8. Animal Behavioral Tests
4.8.1. Y-maze Test
4.8.2. Passive Avoidance Test
4.8.3. Morris Water Maze Test
4.9. Determination of SCFAs in Feces
4.10. Assessment of Mitochondrial Function in the Brain
4.10.1. Extraction of Mitochondria
4.10.2. Mitochondrial ROS Level
4.10.3. MMP
4.10.4. Mitochondrial ATP Level
4.11. Cholinergic System
4.11.1. ACh Contents
4.11.2. AChE Activity
4.12. Antioxidant Activity
4.12.1. MDA Contents
4.12.2. Reduced GSH Level
4.12.3. SOD Level
4.13. Western Blot Assay
4.14. Statistical Analysis
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Tatiya-Aphiradee, N.; Chatuphonprasert, W.; Jarukamjorn, K. Immune response and inflammatory pathway of ulcerative colitis. J. Basic Clin. Physiol. Pharmacol. 2018, 30, 1–10. [Google Scholar] [CrossRef] [PubMed]
- Guan, Q. A comprehensive review and update on the pathogenesis of inflammatory bowel disease. J. Immunol. Res. 2019, 2019, 7247238. [Google Scholar] [CrossRef] [PubMed]
- Paone, P.; Cani, P.D. Mucus barrier, mucins and gut microbiota: The expected slimy partners? Gut 2020, 69, 2232–2243. [Google Scholar] [CrossRef] [PubMed]
- Lee, S.H. Intestinal permeability regulation by tight junction: Implication on inflammatory bowel diseases. Intest. Res. 2015, 13, 11–18. [Google Scholar] [CrossRef]
- Sameer, A.S.; Nissar, S. Toll-like receptors (TLRs): Structure, functions, signaling, and role of their polymorphisms in colorectal cancer susceptibility. BioMed Res. Int. 2021, 2022, 1157023. [Google Scholar] [CrossRef]
- Rivera, E.D.; Coffey, J.C.; Walsh, D.; Ehrenpreis, E.D. The mesentery, systemic inflammation, and Crohn’s disease. Inflamm. Bowel Dis. 2019, 25, 226–234. [Google Scholar] [CrossRef] [PubMed]
- Van Dyken, P.; Lacoste, B. Impact of metabolic syndrome on neuroinflammation and the blood–brain barrier. Front. Neurosci. 2018, 12, 930. [Google Scholar] [CrossRef]
- Bernhart, E.; Kogelnik, N.; Prasch, J.; Gottschalk, B.; Goeritzer, M.; Depaoli, M.R.; Sattler, W. 2-Chlorohexadecanoic acid induces ER stress and mitochondrial dysfunction in brain microvascular endothelial cells. Redox Biol. 2018, 15, 441–451. [Google Scholar] [CrossRef]
- Huang, X.; Hussain, B.; Chang, J. Peripheral inflammation and blood–brain barrier disruption: Effects and mechanisms. CNS Neurosci. Ther. 2021, 27, 36–47. [Google Scholar] [CrossRef]
- O’Riordan, K.J.; Collins, M.K.; Moloney, G.M.; Knox, E.G.; Aburto, M.R.; Fülling, C.; Cryan, J.F. Short chain fatty acids: Microbial metabolites for gut-brain axis signalling. Mol. Cell. Endocrinol. 2022, 546, 111572. [Google Scholar] [CrossRef]
- Parada Venegas, D.; De la Fuente, M.K.; Landskron, G.; González, M.J.; Quera, R.; Dijkstra, G.; Hermoso, M.A. Short chain fatty acids (SCFAs)-mediated gut epithelial and immune regulation and its relevance for inflammatory bowel diseases. Front. Immunol. 2019, 10, 277. [Google Scholar] [CrossRef] [PubMed]
- Feng, H.; Fan, J.; Song, Z.; Du, X.; Chen, Y.; Wang, J.; Song, G. Characterization and immunoenhancement activities of Eucommia ulmoides polysaccharides. Carbohydr. Polym. 2016, 136, 803–811. [Google Scholar] [CrossRef]
- Huang, L.; Lyu, Q.; Zheng, W.; Yang, Q.; Cao, G. Traditional application and modern pharmacological research of Eucommia ulmoides Oliv. Chin. Med. 2021, 16, 73. [Google Scholar] [CrossRef] [PubMed]
- Xing, Y.F.; He, D.; Wang, Y.; Zeng, W.; Zhang, C.; Lu, Y.; Xing, X.H. Chemical constituents, biological functions and pharmacological effects for comprehensive utilization of Eucommia ulmoides Oliver. Food Sci. Hum. Wellness 2019, 8, 177–188. [Google Scholar] [CrossRef]
- Kwon, S.H.; Kim, M.J.; Ma, S.X.; You, I.J.; Hwang, J.Y.; Oh, J.H.; Jang, C.G. Eucommia ulmoides Oliv. Bark. protects against hydrogen peroxide-induced neuronal cell death in SH-SY5Y cells. J. Ethnopharmacol. 2012, 142, 337–345. [Google Scholar] [CrossRef] [PubMed]
- Zhai, Z.; Niu, K.M.; Liu, Y.; Lin, C.; Wu, X. The gut microbiota-bile acids-TGR5 axis mediates Eucommia ulmoides leaf extract alleviation of injury to colonic epithelium integrity. Front. Microbiol. 2021, 12, 727681. [Google Scholar] [CrossRef]
- Aggarwal, M.; Alkhayyat, M.; Abou Saleh, M.; Sarmini, M.T.; Singh, A.; Garg, R.; Cohen, B.L. Alzheimer disease occurs more frequently in patients with inflammatory bowel disease: Insight from a nationwide study. J. Clin. Gastroenterol. 2023, 57, 501–507. [Google Scholar] [CrossRef]
- Wehkamp, J.; Götz, M.; Herrlinger, K.; Steurer, W.; Stange, E.F. Inflammatory bowel disease: Crohn’s disease and ulcerative colitis. Dtsch. Ärztebl. Int. 2016, 113, 72. [Google Scholar] [CrossRef]
- Hussain, T.; Tan, B.E.; Liu, G.; Oladele, O.A.; Rahu, N.; Tossou, M.C.; Yin, Y. Health-promoting properties of Eucommia ulmoides: A review. Evid. Based Complement. Altern. Med. 2016, 2016, 5202908. [Google Scholar] [CrossRef]
- Wan, F.; Cai, X.; Wang, M.; Chen, L.; Zhong, R.; Liu, L.; Zhang, H. Chlorogenic acid supplementation alleviates dextran sulfate sodium (DSS)-induced colitis via inhibiting inflammatory responses and oxidative stress, improving gut barrier integrity and Nrf-2/HO-1 pathway. J. Funct. Foods 2021, 87, 104808. [Google Scholar] [CrossRef]
- Moshahid Khan, M.; Raza, S.S.; Javed, H.; Ahmad, A.; Khan, A.; Islam, F.; Islam, F. Rutin protects dopaminergic neurons from oxidative stress in an animal model of Parkinson’s disease. Neurotox. Res. 2012, 22, 1–15. [Google Scholar] [CrossRef] [PubMed]
- Dong, J.; Chen, Y.; Tang, Y.; Xu, F.; Yu, C.; Li, Y.; Dai, N. Body mass index is associated with inflammatory bowel disease: A systematic review and meta-analysis. PLoS ONE 2015, 10, e0144872. [Google Scholar] [CrossRef] [PubMed]
- Khan, N.; Patel, D.; Shah, Y.; Trivedi, C.; Yang, Y.X. Albumin as a prognostic marker for ulcerative colitis. World J. Gastroenterol. 2017, 23, 8008. [Google Scholar] [CrossRef] [PubMed]
- Murakami, S.; Tasaka, Y.; Takatori, S.; Tanaka, A.; Kawasaki, H.; Araki, H. Effect of Eucommia ulmoides leaf extract on chronic dextran sodium sulfate-induced colitis in mice. Biol. Pharm. Bull. 2018, 41, 864–868. [Google Scholar] [CrossRef] [PubMed]
- Fang, J.; Wang, H.; Zhou, Y.; Zhang, H.; Zhou, H.; Zhang, X. Slimy partners: The mucus barrier and gut microbiome in ulcerative colitis. Exp. Mol. Med. 2021, 53, 772–787. [Google Scholar] [CrossRef] [PubMed]
- Ramos, G.P.; Papadakis, K.A. Mechanisms of disease: Inflammatory bowel diseases. Mayo Clin. Proc. 2019, 94, 155–165. [Google Scholar] [CrossRef] [PubMed]
- Wardill, H.R.; Mander, K.A.; Van Sebille, Y.Z.; Gibson, R.J.; Logan, R.M.; Bowen, J.M.; Sonis, S.T. Cytokine-mediated blood brain barrier disruption as a conduit for cancer/chemotherapy-associated neurotoxicity and cognitive dysfunction. Int. J. Cancer 2016, 139, 2635–2645. [Google Scholar] [CrossRef] [PubMed]
- He, X.F.; Li, L.L.; Xian, W.B.; Li, M.Y.; Zhang, L.Y.; Xu, J.H.; Hu, X.Q. Chronic colitis exacerbates NLRP3-dependent neuroinflammation and cognitive impairment in middle-aged brain. J. Neuroinflamm. 2021, 18, 153. [Google Scholar] [CrossRef]
- van Langenberg, D.R.; Yelland, G.W.; Robinson, S.R.; Gibson, P.R. Cognitive impairment in Crohn’s disease is associated with systemic inflammation, symptom burden and sleep disturbance. United Eur. Gastroenterol. J. 2017, 5, 579–587. [Google Scholar] [CrossRef]
- Heitman, E.; Ingram, D.K. Cognitive and neuroprotective effects of chlorogenic acid. Nutr. Neurosci. 2017, 20, 32–39. [Google Scholar] [CrossRef]
- Deleu, S.; Machiels, K.; Raes, J.; Verbeke, K.; Vermeire, S. Short chain fatty acids and its producing organisms: An overlooked therapy for IBD? EBioMedicine 2021, 66, 103293. [Google Scholar] [CrossRef]
- Mirzaei, R.; Bouzari, B.; Hosseini-Fard, S.R.; Mazaheri, M.; Ahmadyousefi, Y.; Abdi, M.; Karampoor, S. Role of microbiota-derived short-chain fatty acids in nervous system disorders. Biomed. Pharmacother. 2021, 139, 111661. [Google Scholar] [CrossRef] [PubMed]
- Liu, J.; Jin, Y.; Ye, Y.; Tang, Y.; Dai, S.; Li, M.; Lu, Z.Q. The neuroprotective effect of short chain fatty acids against sepsis-associated encephalopathy in mice. Front. Immunol. 2021, 12, 626894. [Google Scholar] [CrossRef] [PubMed]
- Zhao, X.; Wang, Y.; Nie, Z.; Han, L.; Zhong, X.; Yan, X.; Gao, X. Eucommia ulmoides leaf extract alters gut microbiota composition, enhances short-chain fatty acids production, and ameliorates osteoporosis in the senescence-accelerated mouse P6 (SAMP6) model. Food Sci. Nutr. 2020, 8, 4897–4906. [Google Scholar] [CrossRef] [PubMed]
- Wong, K.Y.; Roy, J.; Fung, M.L.; Heng, B.C.; Zhang, C.; Lim, L.W. Relationships between mitochondrial dysfunction and neurotransmission failure in Alzheimer’s disease. Aging Dis. 2020, 11, 1291. [Google Scholar] [CrossRef]
- Annesley, S.J.; Fisher, P.R. Mitochondria in health and disease. Cells 2019, 8, 680. [Google Scholar] [CrossRef]
- Cobley, J.N.; Fiorello, M.L.; Bailey, D.M. 13 reasons why the brain is susceptible to oxidative stress. Redox Biol. 2018, 15, 490–503. [Google Scholar] [CrossRef] [PubMed]
- Jankowska-Kulawy, A.; Klimaszewska-Łata, J.; Gul-Hinc, S.; Ronowska, A.; Szutowicz, A. Metabolic and cellular compartments of acetyl-CoA in the healthy and diseased brain. Int. J. Mol. Sci. 2022, 23, 10073. [Google Scholar] [CrossRef] [PubMed]
- Kong, D.; Ding, Y.; Liu, J.; Liu, R.; Zhang, J.; Zhou, Q.; Hai, C. Chlorogenic acid prevents paraquat-induced apoptosis via Sirt1-mediated regulation of redox and mitochondrial function. Free Radic. Res. 2019, 53, 680–693. [Google Scholar] [CrossRef]
- Kim, M.J.; Kang, J.Y.; Kim, J.M.; Moon, J.H.; Lee, H.L.; Jeong, H.R.; Heo, H.J. Effect of Ethyl Acetate Fraction from Eucommia ulmoides Leaves on PM 2.5-Induced Inflammation and Cognitive Dysfunction. Oxidative Med. Cell. Longev. 2022, 2022, 7157444. [Google Scholar] [CrossRef]
- Mogilevski, T.; Burgell, R.; Aziz, Q.; Gibson, P.R. the role of the autonomic nervous system in the pathogenesis and therapy of IBD. Aliment. Pharmacol. Ther. 2019, 50, 720–737. [Google Scholar] [CrossRef]
- Hampel, H.; Mesulam, M.M.; Cuello, A.C.; Farlow, M.R.; Giacobini, E.; Grossberg, G.T.; Khachaturian, Z.S. The cholinergic system in the pathophysiology and treatment of Alzheimer’s disease. Brain 2018, 141, 1917–1933. [Google Scholar] [CrossRef]
- Wong-Guerra, M.; Jiménez-Martin, J.; Pardo-Andreu, G.L.; Fonseca-Fonseca, L.A.; Souza, D.O.; de Assis, A.M.; Nuñez-Figueredo, Y. Mitochondrial involvement in memory impairment induced by scopolamine in rats. Neurol. Res. 2017, 39, 649–659. [Google Scholar] [CrossRef] [PubMed]
- Balmus, I.M.; Ciobica, A.; Trifan, A.; Stanciu, C. The implications of oxidative stress and antioxidant therapies in Inflammatory Bowel Disease: Clinical aspects and animal models. Saudi J. Gastroenterol. 2016, 22, 3. [Google Scholar] [PubMed]
- Bhatti, J.S.; Bhatti, G.K.; Reddy, P.H. Mitochondrial dysfunction and oxidative stress in metabolic disorders—A step towards mitochondria based therapeutic strategies. Biochim. Biophys. Acta (BBA) Mol. Basis Dis. 2017, 1863, 1066–1077. [Google Scholar] [CrossRef] [PubMed]
- Dunnill, C.; Patton, T.; Brennan, J.; Barrett, J.; Dryden, M.; Cooke, J.; Georgopoulos, N.T. Reactive oxygen species (ROS) and wound healing: The functional role of ROS and emerging ROS-modulating technologies for augmentation of the healing process. Int. Wound J. 2017, 14, 89–96. [Google Scholar] [CrossRef]
- Angelova, P.R.; Abramov, A.Y. Role of mitochondrial ROS in the brain: From physiology to neurodegeneration. FEBS Lett. 2018, 592, 692–702. [Google Scholar] [CrossRef]
- Snezhkina, A.V.; Kudryavtseva, A.V.; Kardymon, O.L.; Savvateeva, M.V.; Melnikova, N.V.; Krasnov, G.S.; Dmitriev, A.A. ROS generation and antioxidant defense systems in normal and malignant cells. Oxidative Med. Cell. Longev. 2019, 2019, 6175804. [Google Scholar] [CrossRef]
- Gong, M.; Li, Q.; Guo, H.; Cui, B.; Liu, Y.; Wang, P.; Wang, Z. Protective effect of active components of Eucommia ulmoides leaves on gastric ulcers in rats: Involvement of the PI3K/Akt/NF-κB pathway. J. Food Sci. 2022, 87, 3207–3222. [Google Scholar] [CrossRef]
- Shi, H.; Shi, A.; Dong, L.; Lu, X.; Wang, Y.; Zhao, J.; Guo, X. Chlorogenic acid protects against liver fibrosis in vivo and in vitro through inhibition of oxidative stress. Clin. Nutr. 2016, 35, 1366–1373. [Google Scholar] [CrossRef]
- De Nardo, D. Toll-like receptors: Activation, signalling and transcriptional modulation. Cytokine 2015, 74, 181–189. [Google Scholar] [CrossRef] [PubMed]
- Ye, S.; Zheng, Q.; Zhou, Y.; Bai, B.; Yang, D.; Zhao, Z. Chlojaponilactone B attenuates lipopolysaccharide-induced inflammatory responses by suppressing TLR4-mediated ROS generation and NF-κB signaling pathway. Molecules 2019, 24, 3731. [Google Scholar] [CrossRef]
- He, X.; Qian, Y.; Li, Z.; Fan, E.K.; Li, Y.; Wu, L.; Fan, J. TLR4-upregulated IL-1β and IL-1RI promote alveolar macrophage pyroptosis and lung inflammation through an autocrine mechanism. Sci. Rep. 2016, 6, 31663. [Google Scholar] [CrossRef] [PubMed]
- Hanscom, M.; Loane, D.J.; Aubretch, T.; Leser, J.; Molesworth, K.; Hedgekar, N.; Faden, A.I. Acute colitis during chronic experimental traumatic brain injury in mice induces dysautonomia and persistent extraintestinal, systemic, and CNS inflammation with exacerbated neurological deficits. J. Neuroinflamm. 2021, 18, 24. [Google Scholar] [CrossRef] [PubMed]
- Zheng, Y.; Li, L.; Chen, B.; Fang, Y.; Lin, W.; Zhang, T.; Lin, Z. Chlorogenic acid exerts neuroprotective effect against hypoxia-ischemia brain injury in neonatal rats by activating Sirt1 to regulate the Nrf2-NF-κB signaling pathway. Cell Commun. Signal. 2022, 20, 84. [Google Scholar] [CrossRef] [PubMed]
- Ghezzi, P.; Floridi, L.; Boraschi, D.; Cuadrado, A.; Manda, G.; Levic, S.; Selley, L. Oxidative stress and inflammation induced by environmental and psychological stressors: A biomarker perspective. Antioxid. Redox Signal. 2018, 28, 852–872. [Google Scholar] [CrossRef] [PubMed]
- Li, Y.; Sun, J.; Wu, R.; Bai, J.; Hou, Y.; Zeng, Y.; Meng, X. Mitochondrial MPTP: A novel target of ethnomedicine for stroke treatment by apoptosis inhibition. Front. Pharmacol. 2020, 11, 352. [Google Scholar] [CrossRef]
- Changizi, Z.; Moslehi, A.; Rohani, A.H.; Eidi, A. Chlorogenic acid induces 4T1 breast cancer tumor’s apoptosis via p53, Bax, Bcl-2, and caspase-3 signaling pathways in BALB/c mice. J. Biochem. Mol. Toxicol. 2021, 35, 22642. [Google Scholar] [CrossRef] [PubMed]
- Wang, W.; Zhu, M.; Xu, Z.; Li, W.; Dong, X.; Chen, Y.; Li, M. Ropivacaine promotes apoptosis of hepatocellular carcinoma cells through damaging mitochondria and activating caspase-3 activity. Biol. Res. 2019, 52, 36. [Google Scholar] [CrossRef]
- Setyaningsih, M.; Meliala, A.; Cempaka, R.; Arfian, N. Chlorogenic Acid Ameliorates Liver Function in Association with Bax Downregulation, P53 Downregulation and Bcl-2 Upregulation in Diabetic Wistar Rat. BIO Web Conf. 2022, 49, 03004. [Google Scholar] [CrossRef]
- Cheng, H.; Dong, H.; Wei, J.; You, G. Eucommia ulmoides extract alleviated spinal cord injury in rats by inhibiting endoplasmic reticulum stress and oxidative stress. Qual. Assur. Saf. Crops Foods 2022, 14, 84–90. [Google Scholar] [CrossRef]
- Lee, H.L.; Kim, J.M.; Moon, J.H.; Kim, M.J.; Jeong, H.R.; Go, M.J.; Heo, H.J. Anti-Amnesic Effect of Synbiotic Supplementation Containing Corni fructus and Limosilactobacillus reuteri in DSS-Induced Colitis Mice. Int. J. Mol. Sci. 2022, 24, 90. [Google Scholar] [CrossRef] [PubMed]
- Kim, J.M.; Lee, U.; Kang, J.Y.; Park, S.K.; Shin, E.J.; Kim, H.J.; Heo, H.J. Anti-amnesic effect of walnut via the regulation of BBB function and neuro-inflammation in Aβ1-42-induced mice. Antioxidants 2020, 9, 976. [Google Scholar] [CrossRef] [PubMed]
- Park, S.K.; Kang, J.Y.; Kim, J.M.; Park, S.H.; Kwon, B.S.; Kim, G.H.; Heo, H.J. Protective effect of fucoidan extract from Ecklonia cava on hydrogen peroxide-induced neurotoxicity. J. Microbiol. Biotechnol. 2018, 28, 40–49. [Google Scholar] [CrossRef]
- Kim, J.M.; Park, S.K.; Kang, J.Y.; Park, S.B.; Yoo, S.K.; Han, H.J.; Heo, H.J. Ethyl acetate fraction from persimmon (Diospyros kaki) ameliorates cerebral neuronal loss and cognitive deficit via the JNK/Akt pathway in TMT-induced mice. Int. J. Mol. Sci. 2018, 19, 1499. [Google Scholar] [CrossRef]











{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| No. | Retention Time (min) | Parent Ion (m/z) | Fragment Ion (m/z) | Compound |
|---|---|---|---|---|
| 1 | 0.68 | 533 | 191, 109, 85 | Quinic acid derivative |
| 2 | 0.69 | 683 | 341,179, 161, 113 | Caffeic acid-O-hexoside |
| 3 | 2.96 | 353 | 191, 173, 161, 85 | 3-O-Caffeoylquinic acid |
| 4 | 3.32 | 595 | 301, 300, 271, 255 | Quercetin pentosyl-hexoside |
| 5 | 3.42 | 609 | 301, 300, 271, 255, 151 | Rutin |
| 6 | 3.60 | 505 | 301, 300, 271, 255, 178 | Quercetin acetyl hexose |
| Acetic Acid | Propionic Acid | Butyric Acid | |
|---|---|---|---|
| Con | 8.04 ± 0.09 a | 4.79 ± 0.10 a | 2.08 ± 0.09 a |
| DSS | 4.42 ± 0.42 c | 4.65 ± 0.08 b | 1.69 ± 0.18 c |
| AEEL 400 | 6.23 ± 0.65 b | 4.65 ± 0.01 b | 1.93 ± 0.12 b |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Lee, H.S.; Kim, J.M.; Lee, H.L.; Go, M.J.; Lee, D.Y.; Kim, C.-W.; Kim, H.-J.; Heo, H.J. Eucommia ulmoides Leaves Alleviate Cognitive Dysfunction in Dextran Sulfate Sodium (DSS)-Induced Colitis Mice through Regulating JNK/TLR4 Signaling Pathway. Int. J. Mol. Sci. 2024, 25, 4063. https://doi.org/10.3390/ijms25074063
Lee HS, Kim JM, Lee HL, Go MJ, Lee DY, Kim C-W, Kim H-J, Heo HJ. Eucommia ulmoides Leaves Alleviate Cognitive Dysfunction in Dextran Sulfate Sodium (DSS)-Induced Colitis Mice through Regulating JNK/TLR4 Signaling Pathway. International Journal of Molecular Sciences. 2024; 25(7):4063. https://doi.org/10.3390/ijms25074063
Chicago/Turabian StyleLee, Han Su, Jong Min Kim, Hyo Lim Lee, Min Ji Go, Dong Yeol Lee, Chul-Woo Kim, Hyun-Jin Kim, and Ho Jin Heo. 2024. "Eucommia ulmoides Leaves Alleviate Cognitive Dysfunction in Dextran Sulfate Sodium (DSS)-Induced Colitis Mice through Regulating JNK/TLR4 Signaling Pathway" International Journal of Molecular Sciences 25, no. 7: 4063. https://doi.org/10.3390/ijms25074063
APA StyleLee, H. S., Kim, J. M., Lee, H. L., Go, M. J., Lee, D. Y., Kim, C.-W., Kim, H.-J., & Heo, H. J. (2024). Eucommia ulmoides Leaves Alleviate Cognitive Dysfunction in Dextran Sulfate Sodium (DSS)-Induced Colitis Mice through Regulating JNK/TLR4 Signaling Pathway. International Journal of Molecular Sciences, 25(7), 4063. https://doi.org/10.3390/ijms25074063