

Gene–Environment Interactions in Irrational Beliefs: The Roles of Childhood Adversity and Multiple Candidate Genes
 ,
,
Abstract
:1. Introduction
2. Results
2.1. Preliminary Analyses
2.1.1. Genotype Frequencies
2.1.2. Correlational Analysis
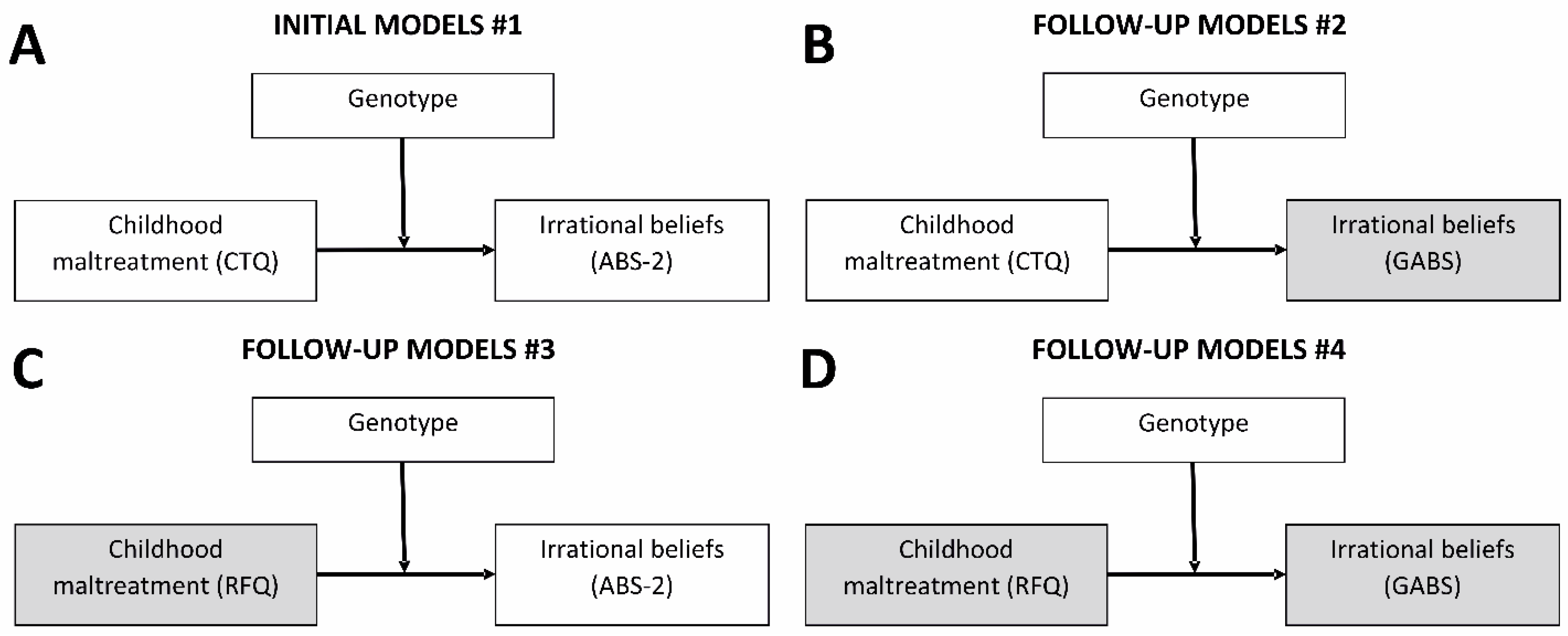
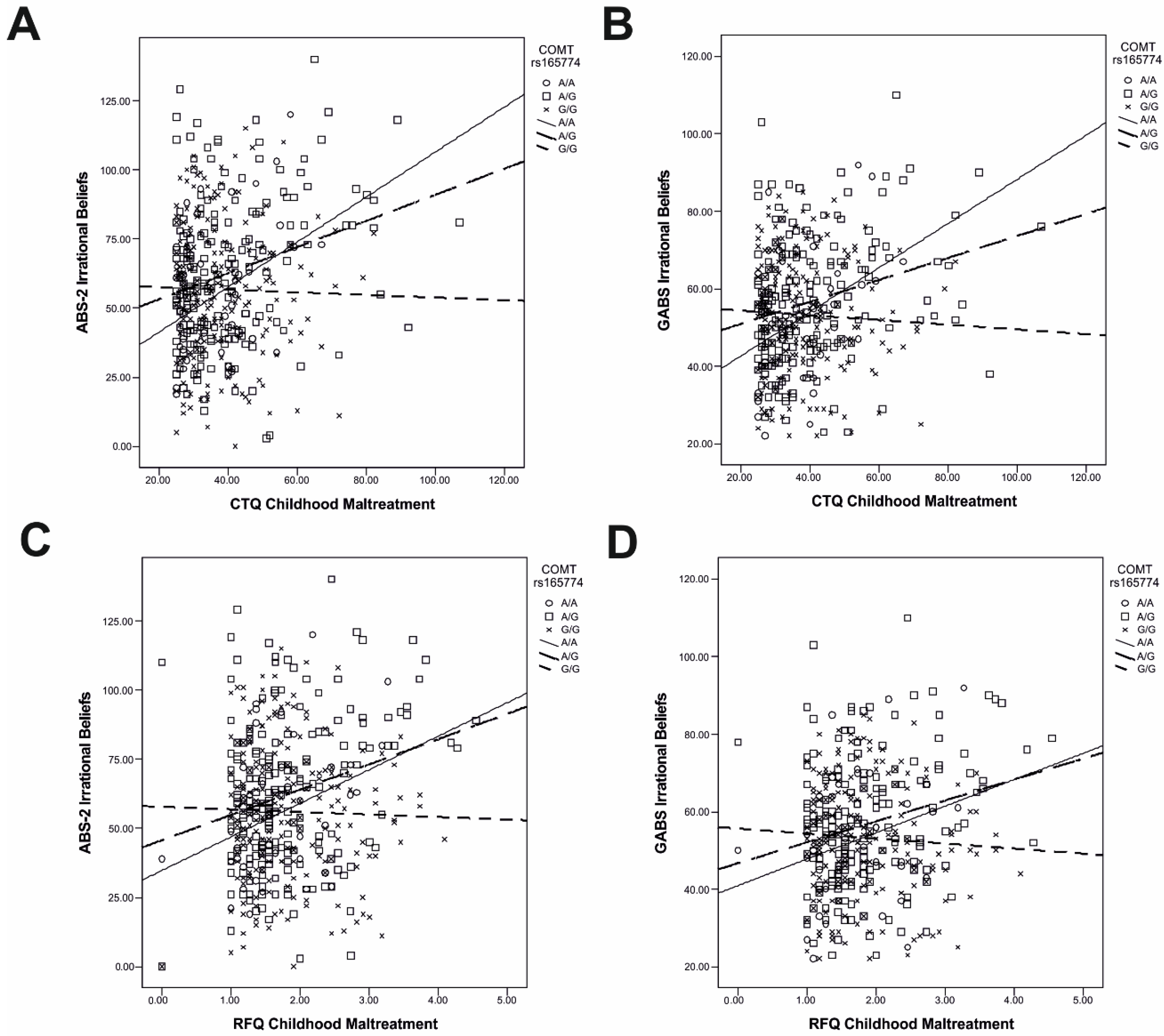
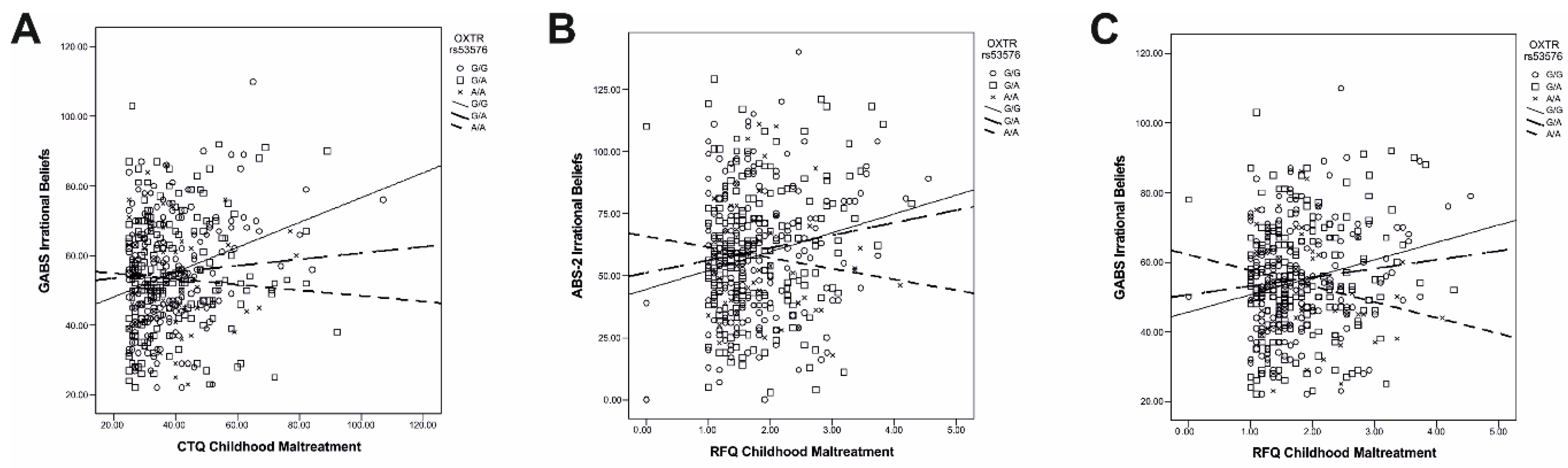
2.2. Gene–Environment Interactions
3. Discussion
4. Material and Methods
4.1. Participants
4.2. Measures
4.2.1. Genotyping
4.2.2. Childhood Adversity
4.2.3. Irrational Beliefs
4.3. Statistical Analysis
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- David, D.; Cristea, I.; Hofmann, S.G. Why cognitive behavioral therapy is the current gold standard of psychotherapy. Front. Psychiatry 2018, 9, 4. [Google Scholar] [CrossRef] [PubMed]
- Miller, G.A. The cognitive revolution: A historical perspective. Trends Cogn. Sci. 2003, 7, 141–144. [Google Scholar] [CrossRef]
- Beck, A.T. Depression. Clinical, Experimental and Theoretical Aspects; Hoeber: New York, NY, USA, 1967. [Google Scholar]
- Ellis, A. Reason and Emotion in Psychotherapy; Citadel: Secaucus, NJ, USA, 1962. [Google Scholar]
- David, D.; Cardoș, R.; Cândea, D.; Oltean, H.; Ştefan, S. REBT and Depressive Disorders. In REBT with Diverse Client Problems and Populations; Dryden, W., Bernard, M.E., Eds.; Springer International Publishing: Cham, Switzerland, 2019; pp. 23–44. [Google Scholar] [CrossRef]
- Ellis, A. Thinking processes involved in irrational beliefs and their disturbed consequences. J. Cogn. Psychother. 1995, 9, 105–116. [Google Scholar] [CrossRef]
- Soflau, R.; David, D. A meta-analytical approach of the relationships between the irrationality of beliefs and the functionality of automatic thoughts. Cogn. Ther. Res. 2017, 41, 178–192. [Google Scholar] [CrossRef]
- Soflau, R.; David, D. The impact of irrational beliefs on paranoid thoughts. Behav. Cogn. Psychother. 2019, 47, 270–286. [Google Scholar] [CrossRef]
- Vukosavljevic-Gvozden, T.; Filipovic, S.; Opacic, G. The mediating role of symptoms of psychopathology between irrational beliefs and Internet gaming addiction. J. Ration.-Emotive Cogn.-Behav. Ther. 2015, 33, 387–405. [Google Scholar] [CrossRef]
- Balkis, M.; Duru, E. Procrastination and rational/irrational beliefs: A moderated mediation model. J. Ration.-Emotive Cogn.-Behav. Ther. 2019, 37, 299–315. [Google Scholar] [CrossRef]
- Szentagotai-Tatar, A.; Freeman, A. An analysis of the relationship between irrational beliefs and automatic thoughts in predicting distress. J. Cogn. Behav. Psychother. 2007, 7, 1–9. [Google Scholar]
- Vîslă, A.; Flückiger, C.; Grosse Holtforth, M.; David, D. Irrational beliefs and psychological distress: A meta-analysis. Psychother. Psychosom. 2016, 85, 8–15. [Google Scholar] [CrossRef]
- David, D.; Lynn, S.J.; Ellis, A. Rational and Irrational Beliefs: Research, Theory, and Clinical Practice; Oxford University Press: New York, NY, USA, 2009. [Google Scholar]
- David, D.; Szentagotai, A.; Lupu, V.; Cosman, D. Rational emotive behavior therapy, cognitive therapy, and medication in the treatment of major depressive disorder: A randomized clinical trial, posttreatment outcomes, and six-month follow-up. J. Clin. Psychol. 2008, 64, 728–746. [Google Scholar] [CrossRef]
- Ellis, A. The biological basis of human irrationality. J. Individ. Psychol. Health 1976, 32, 145–168. [Google Scholar]
- Ellis, A. Is rational-emotive therapy (RET)“rationalist” or “constructivist”? J. Ration.-Emotive Cogn.-Behav. Ther. 1990, 8, 169–193. [Google Scholar] [CrossRef]
- Beck, A.T.; Haigh, E.A. Advances in cognitive theory and therapy: The generic cognitive model. Annu. Rev. Clin. Psychol. 2014, 10, 1–24. [Google Scholar] [CrossRef] [PubMed]
- Monroe, S.M.; Simons, A.D. Diathesis-stress theories in the context of life stress research: Implications for the depressive disorders. Psychol. Bull. 1991, 110, 406–425. [Google Scholar] [CrossRef]
- Chen, J.; Li, X. Genetic and environmental etiologies of adolescent dysfunctional attitudes: A twin study. Twin Res. Hum. Genet. 2014, 17, 16–22. [Google Scholar] [CrossRef] [PubMed]
- Podina, I.; Popp, R.; Pop, I.; David, D. Genetic correlates of maladaptive beliefs: COMT Val158Met and irrational cognitions linked depending on distress. Behav. Ther. 2015, 46, 797–808. [Google Scholar] [CrossRef]
- Duru, E.; Balkis, M. Childhood trauma, depressive symptoms and rational/irrational beliefs: A moderated mediation model. Curr. Psychol. 2022, 41, 7258–7270. [Google Scholar] [CrossRef]
- Soflau, R.; Szentagotai-Tatar, A.; Oltean, L.E. Childhood adversity, resilience, and paranoia during the COVID-19 outbreak. The mediating role of irrational beliefs and affective disturbance. J. Ration.-Emotive Cogn.-Behav. Ther. 2023, 1–16. [Google Scholar] [CrossRef]
- Estévez, A.; Ozerinjauregi, N.; Herrero-Fernández, D.; Jauregui, P. The mediator role of early maladaptive schemas between childhood sexual abuse and impulsive symptoms in female survivors of CSA. J. Interpers. Violence 2016, 34, 763–784. [Google Scholar] [CrossRef]
- Feyzioğlu, A.; Taşlıoğlu Sayıner, A.C.; Özçelik, D.; Tarımtay Altun, F.; Budak, E.N. The mediating role of early maladaptive schemas in the relationship between early childhood trauma and alexithymia. Curr. Psychol. 2023, 42, 17851–17861. [Google Scholar] [CrossRef]
- Wright, M.O.D.; Crawford, E.; Del Castillo, D. Childhood emotional maltreatment and later psychological distress among college students: The mediating role of maladaptive schemas. Child Abus. Negl. 2009, 33, 59–68. [Google Scholar] [CrossRef]
- Berman, I.S.; Petretric, P.; Bridges, A.J. Beyond child maltreatment: The incremental value of household dysfunction in the prediction of negative beliefs and internalizing symptoms in women. J. Am. Coll. Health 2021, 69, 537–545. [Google Scholar] [CrossRef]
- Gibb, B.E.; Alloy, L.B.; Abramson, L.Y.; Rose, D.T.; Whitehouse, W.G.; Donovan, P.; Hogan, M.E.; Cronholm, J.; Tierney, S. History of childhood maltreatment, negative cognitive styles, and episodes of depression in adulthood. Cogn. Ther. Res. 2001, 25, 425–446. [Google Scholar] [CrossRef]
- Kaysen, D.; Scher, C.D.; Mastnak, J.; Resick, P. Cognitive mediation of childhood maltreatment and adult depression in recent crime victims. Behav. Ther. 2005, 36, 235–244. [Google Scholar] [CrossRef] [PubMed]
- Lesch, K.P.; Bengel, D.; Heils, A.; Sabol, S.Z.; Greenberg, B.D.; Petri, S.; Benjamin, J.; Muller, C.R.; Hamer, D.H.; Murphy, D.L. Association of anxiety-related traits with a polymorphism in the serotonin transporter gene regulatory region. Science 1996, 274, 1527–1531. [Google Scholar] [CrossRef] [PubMed]
- Gaysina, D.; Xu, M.K.; Barnett, J.H.; Croudace, T.J.; Wong, A.; Richards, M.; Jones, P.B.; The LHA Genetics Group. The catechol-O-methyltransferase gene (COMT) and cognitive function from childhood through adolescence. Biol. Psychol. 2013, 92, 359–364. [Google Scholar] [CrossRef]
- Meloto, C.B.; Segall, S.K.; Smith, S.; Parisien, M.; Shabalina, S.A.; Rizzatti-Barbosa, C.M.; Gauthier, J.; Tsao, D.; Convertino, M.; Piltonen, M.H.; et al. COMT gene locus: New functional variants. Pain 2015, 156, 2072–2083. [Google Scholar] [CrossRef] [PubMed]
- Chen, J.; Lipska, B.K.; Halim, N.; Ma, Q.D.; Matsumoto, M.; Melhem, S.; Kolachana, B.S.; Hyde, T.M.; Herman, M.M.; Apud, J.; et al. Functional analysis of genetic variation in catechol-O-methyltransferase (COMT): Effects on mRNA, protein, and enzyme activity in postmortem human brain. Am. J. Hum. Genet. 2004, 75, 807–821. [Google Scholar] [CrossRef]
- Roussos, P.; Giakoumaki, S.G.; Pavlakis, S.; Bitsios, P. Planning, decision-making and the COMT rs4818 polymorphism in healthy males. Neuropsychologia 2008, 46, 757–763. [Google Scholar] [CrossRef]
- Galecka, E.; Szemraj, J.; Bienkiewicz, M.; Majsterek, I.; Przybylowska-Sygut, K.; Galecki, P.; Lewinski, A. Single nucleotide polymorphisms of NR3C1 gene and recurrent depressive disorder in population of Poland. Mol. Biol. Rep. 2013, 40, 1693–1699. [Google Scholar] [CrossRef]
- Tops, S.; Habel, U.; Radke, S. Genetic and epigenetic regulatory mechanisms of the oxytocin receptor gene (OXTR) and the (clinical) implications for social behavior. Horm. Behav. 2019, 108, 84–93. [Google Scholar] [CrossRef]
- Camerini, L.; Zurchimitten, G.; Bock, B.; Xavier, J.; Bastos, C.R.; Martins, E.; Ardais, A.P.; Dos Santos Motta, J.V.; Pires, A.J.; de Matos, M.B.; et al. Genetic variations in elements of the oxytocinergic pathway are associated with attention/hyperactivity problems and anxiety problems in childhood. Child Psychiatry Hum. Dev. 2022, 55, 552–563. [Google Scholar] [CrossRef] [PubMed]
- Mannisto, P.T.; Kaakkola, S. Catechol-O-methyltransferase (COMT): Biochemistry, molecular biology, pharmacology, and clinical efficacy of the new selective COMT inhibitors. Pharmacol. Rev. 1999, 51, 593–628. [Google Scholar]
- Kaenmaki, M.; Tammimaki, A.; Myohanen, T.; Pakarinen, K.; Amberg, C.; Karayiorgou, M.; Gogos, J.A.; Mannisto, P.T. Quantitative role of COMT in dopamine clearance in the prefrontal cortex of freely moving mice. J. Neurochem. 2010, 114, 1745–1755. [Google Scholar] [CrossRef]
- Desbonnet, L.; Tighe, O.; Karayiorgou, M.; Gogos, J.A.; Waddington, J.L.; O’Tuathaigh, C.M. Physiological and behavioural responsivity to stress and anxiogenic stimuli in COMT-deficient mice. Behav. Brain Res. 2012, 228, 351–358. [Google Scholar] [CrossRef] [PubMed]
- Quintana, D.S.; Rokicki, J.; van der Meer, D.; Alnaes, D.; Kaufmann, T.; Cordova-Palomera, A.; Dieset, I.; Andreassen, O.A.; Westlye, L.T. Oxytocin pathway gene networks in the human brain. Nat. Commun. 2019, 10, 668. [Google Scholar] [CrossRef]
- Lee, H.J.; Caldwell, H.K.; Macbeth, A.H.; Tolu, S.G.; Young, W.S., III. A conditional knockout mouse line of the oxytocin receptor. Endocrinology 2008, 149, 3256–3263. [Google Scholar] [CrossRef]
- Takayanagi, Y.; Yoshida, M.; Bielsky, I.F.; Ross, H.E.; Kawamata, M.; Onaka, T.; Yanagisawa, T.; Kimura, T.; Matzuk, M.M.; Young, L.J.; et al. Pervasive social deficits, but normal parturition, in oxytocin receptor-deficient mice. Proc. Natl. Acad. Sci. USA 2005, 102, 16096–16101. [Google Scholar] [CrossRef] [PubMed]
- Seib, C.; Whiteside, E.; Voisey, J.; Lee, K.; Alexander, K.; Humphreys, J.; Chopin, L.; Anderson, D. Stress, COMT polymorphisms, and depressive symptoms in older Australian women: An exploratory study. Genet. Test. Mol. Biomark. 2016, 20, 478–481. [Google Scholar] [CrossRef]
- McQuaid, R.J.; McInnis, O.A.; Stead, J.D.; Matheson, K.; Anisman, H. A paradoxical association of an oxytocin receptor gene polymorphism: Early-life adversity and vulnerability to depression. Front. Neurosci. 2013, 7, 128. [Google Scholar] [CrossRef]
- Duncan, L.E.; Keller, M.C. A critical review of the first 10 years of candidate gene-by-environment interaction research in psychiatry. Am. J. Psychiatry 2011, 168, 1041–1049. [Google Scholar] [CrossRef] [PubMed]
- Duncan, L.E.; Ostacher, M.; Ballon, J. How genome-wide association studies (GWAS) made traditional candidate gene studies obsolete. Neuropsychopharmacology 2019, 44, 1518–1523. [Google Scholar] [CrossRef] [PubMed]
- Border, R.; Johnson, E.C.; Evans, L.M.; Smolen, A.; Berley, N.; Sullivan, P.F.; Keller, M.C. No support for historic candidate gene or candidate gene-by-interaction hypotheses for major depression across multiple large samples. Am. J. Psychiatry 2019, 176, 376–387. [Google Scholar] [CrossRef] [PubMed]
- van de Weijer, M.P.; Pelt, D.H.M.; de Vries, L.P.; Baselmans, B.M.L.; Bartels, M. A re-evaluation of candidate gene studies for well-being in light of genome-wide evidence. J. Happiness Stud. 2022, 23, 3031–3053. [Google Scholar] [CrossRef] [PubMed]
- Wu, S.; Jia, M.; Ruan, Y.; Liu, J.; Guo, Y.; Shuang, M.; Gong, X.; Zhang, Y.; Yang, X.; Zhang, D. Positive association of the oxytocin receptor gene (OXTR) with autism in the Chinese Han population. Biol. Psychiatry 2005, 58, 74–77. [Google Scholar] [CrossRef] [PubMed]
- Blomeyer, D.; Treutlein, J.; Esser, G.; Schmidt, M.H.; Schumann, G.; Laucht, M. Interaction between CRHR1 gene and stressful life events predicts adolescent heavy alcohol use. Biol. Psychiatry 2008, 63, 146–151. [Google Scholar] [CrossRef] [PubMed]
- Miu, A.C.; Crisan, L.G.; Chis, A.; Ungureanu, L.; Druga, B.; Vulturar, R. Somatic markers mediate the effect of serotonin transporter gene polymorphisms on Iowa Gambling Task. Genes Brain Behav. 2012, 11, 398–403. [Google Scholar] [CrossRef] [PubMed]
- Wang, C.K.; Aleksic, A.; Xu, M.S.; Procyshyn, R.M.; Ross, C.J.; Vila-Rodriguez, F.; Ramos-Miguel, A.; Yan, R.; Honer, W.G.; Barr, A.M. A tetra-primer amplification refractory system technique for the cost-effective and novel genotyping of eight single-nucleotide polymorphisms of the catechol-O-methyltransferase gene. Genet. Test. Mol. Biomark. 2016, 20, 465–470. [Google Scholar] [CrossRef] [PubMed]
- Wang, C.K.; Xu, M.S.; Ross, C.J.; Lo, R.; Procyshyn, R.M.; Vila-Rodriguez, F.; White, R.F.; Honer, W.G.; Barr, A.M. Development of a cost-efficient novel method for rapid, concurrent genotyping of five common single nucleotide polymorphisms of the brain derived neurotrophic factor (BDNF) gene by tetra-primer amplification refractory mutation system. Int. J. Methods Psychiatr. Res. 2015, 24, 235–244. [Google Scholar] [CrossRef]
- Bernstein, D.P.; Fink, L. Childhood Trauma Questionnaire: A retrospective Self-Report Manual; The Psychological Corporation: San Antonio, TX, USA, 1998. [Google Scholar]
- Taylor, S.E.; Lerner, J.S.; Sage, R.M.; Lehman, B.J.; Seeman, T.E. Early environment, emotions, responses to stress, and health. J. Personal. 2004, 72, 1365–1393. [Google Scholar] [CrossRef]
- Di Giuseppe, R.; Leaf, R.; Gorman, B.; Robin, M.W. The development of a measure of irrational/rational beliefs. J. Ration.-Emotive Cogn.-Behav. Ther. 2018, 36, 47–79. [Google Scholar]
- Lindner, H.; Kirkby, R.; Wertheim, E.; Birch, P. A brief assessment of irrational thinking: The shortened general attitude and belief scale. Cogn. Ther. Res. 1999, 23, 651–663. [Google Scholar] [CrossRef]
- Caspi, A.; Hariri, A.R.; Holmes, A.; Uher, R.; Moffitt, T.E. Genetic sensitivity to the environment: The case of the serotonin transporter gene and its implications for studying complex diseases and traits. Am. J. Psychiatry 2010, 167, 509–527. [Google Scholar] [CrossRef] [PubMed]
- Caspi, A.; McClay, J.; Moffitt, T.E.; Mill, J.; Martin, J.; Craig, I.W.; Taylor, A.; Poulton, R. Role of genotype in the cycle of violence in maltreated children. Science 2002, 297, 851–854. [Google Scholar] [CrossRef] [PubMed]
- Caspi, A.; Sugden, K.; Moffitt, T.E.; Taylor, A.; Craig, I.W.; Harrington, H.; McClay, J.; Mill, J.; Martin, J.; Braithwaite, A.; et al. Influence of life stress on depression: Moderation by a polymorphism in the 5-HTT gene. Science 2003, 301, 386–389. [Google Scholar] [CrossRef] [PubMed]
- Miu, A.C.; Homberg, J.R.; Lesch, K.P. Genes, Brain, and Emotions: Interdisciplinary and Translational Perspectives, 1st ed.; Oxford University Press: Oxford, UK, 2019. [Google Scholar]
- Rodriguez, S.; Gaunt, T.R.; Day, I.N. Hardy-Weinberg equilibrium testing of biological ascertainment for Mendelian randomization studies. Am. J. Epidemiol. 2009, 169, 505–514. [Google Scholar] [CrossRef]
- Keller, M.C. Gene x environment interaction studies have not properly controlled for potential confounders: The problem and the (simple) solution. Biol. Psychiatry 2014, 75, 18–24. [Google Scholar] [CrossRef]
- Hayes, A.F. Introduction to Mediation, Moderation, and Conditional Process Analysis: A Regression-Based Approach, 3rd ed.; Guilford Press: New York, NY, USA, 2022. [Google Scholar]




{kind=link}
{kind=link}
{kind=link}
| Gene | Position on Chromosome, RefSeq | RefSNPID | Ancestral Allele (A1) | Minor Allele (A2) | Allele Functional Characterization | References |
|---|---|---|---|---|---|---|
| SLC6A4 | chr17, NC_000017.11:g.30237328T>C (promoter region) | rs25531 (5-HTTLPR) | T (or L) | C (or S) | The short (C or S) allele is associated with reduced serotonin transporter expression and function compared with the long allele (T or L). | [29] |
| COMT | chr22, NC_000022.11:g.19962429A>G (promoter region) | rs6269 | A | G | The G allele is associated with lower expression of COMT mRNA in the human brain. | [30] |
| chr22, NC_000022.11:g.19942598A>G, (intron region) | rs737865 | A | G | The functionality is unknown. | - | |
| chr22, NC_000022.11:g.19965038G>A (3′-untranslated region) | rs165774 | G | A | The minor A allele is associated with a increased mRNA stability and with substantially decreased activity of the product molecule, most likely through modulation of translational efficiency. | [31] | |
| chr22, NC_000022.11:g.19940569G>A (promoter region) | rs2075507 | G | A | The G allele may be associated with reduced COMT activity. | [32] | |
| chr22, NC_000022.11:g.19963684C>G (coding region) | rs4818 | C | G | It is a synonymous polymorphism, but evidence suggests that the C allele is associated with a lower enzyme activity. | [33] | |
| BDNF | chr11, NC_000011.10:g.27658369C>T (coding region) | rs6265 | G | A | The A allele is associated with a lower expression of BDNF mRNA. | [32] |
| chr11, NC_000011.10:g.27703198C>G (intron region) | rs988748 | C | G | The functionality is unknown. | - | |
| chr11, NC_000011.10:g.27678578C>T (intron region) | rs7103411 | C | T | The functionality is unknown. | - | |
| chr11, NC_000011.10:g.27662970A>G (intron region) | rs11030104 | T | C | The functionality is unknown. | - | |
| chr11, NC_000011.10:g.27646808G>C (intron region) | rs16917204 (rs11757) | G | C | The functionality is unknown. | - | |
| CRHR1 | chr17, NC_000017.11:g.45818570A>C (intron region) | rs242938 | A | G | The functionality is unknown. | - |
| NR3C1 | chr5, NC_000005.10:g.143399010G>C (intron 2) | rs41423247 (or BclI) | C | G | The sensitivity to glucocorticoids appears to be different and tissue-specific. | [34] |
| chr5, NC_000005.10:g.143400774C>A/ NC_000005.10:g.143400772C>G (coding region) | rs6189/rs6190 (ER22/23EK) | G | A | The first variant is silent. The second variant results in arginine (R) to lysine (K) change. The presence of the ER22/23EK A allele is related to resistance to glucocorticoids. | ||
| chr5, NC_000005.10:g.143399752T>A (coding region) | rs56149945 (N363S) | A | G | The G allele is associated with increased sensitivity to glucocorticoids. | ||
| OXTR | chr3, NC_000003.12:g.8762685A>G (intron region) | rs53576 | A | G | The functionality is unknown. | [35] |
| chr3, NC_000003.12:g.8760542G>A (intron region) | rs2254298 | G | A | The functionality is unknown. | [36] |
| Gene Polymorphism (N) | Genotype (Frequency %) | Allele (Frequency %) | Hardy–Weinberg Equilibrium | |||
|---|---|---|---|---|---|---|
| COMT rs6269 (N = 309) | AA (37.54) | AG (44.66) | GG (17.8) | A (59.87) | G (40.13) | χ2 = 1.53, p = 0.214 |
| COMT rs737865 (N = 438) | AA (47.72) | AG (42.01) | GG (10.27) | A (68.73) | G (31.27) | χ2 = 0.22, p = 0.633 |
| COMT rs165774 (N = 438) | AA (9.36) | AG (44.52) | GG (46.12) | A (31.62) | G (68.38) | χ2 = 0.38, p = 0.536 |
| COMT rs2075507 (N = 309) | AA (18.77) | AG (43.69) | GG (37.54) | A (40.62) | G (59.38) | χ2 = 2.74, p = 0.097 |
| COMT rs4818 (N = 309) | CC (39.48) | CG (42.72) | GG (17.8) | C (60.84) | G (39.16) | χ2 = 3.30, p = 0.068 |
| NR3C1 BclI (N = 309) | CC (46.6) | CG (38.83) | GG (14.56) | C (66.10) | G (33.90) | χ2 = 5.58, p = 0.018 |
| NR3C1 N363S (N = 308) | AA (94.16) | AG (5.84) | GG (0) | A (97.08) | G (2.92) | χ2 = 0.27, p = 0.597 |
| NR3C1 ER22/23EK (N = 309) | GG (95.15) | GA (4.85) | AA (0) | G (97.58) | A (2.42) | χ2 = 0.19, p = 0.661 |
| OXTR rs2254298 (N = 309) | GG (85.52) | AG (16.5) | AA (0.97) | G (93.77) | A (9.23) | χ2 = 0.06, p = 0.661 |
| OXTR rs53576 (N = 438) | GG (45.89) | AG (43.15) | AA (10.96) | G (67.47) | A (32.54) | χ2 = 0.12, p = 0.721 |
| CRHR1 rs242938 (N = 309) | GG (84.14) | GA (15.53) | AA (0.32) | G (91.91) | A (8.09) | χ2 = 0.61, p = 0.434 |
| SLC6A4 5HTTLPR/rs25531 (N = 309) | L′/L′ (24.27) | L′/S′ (51.46) | S′/S′ (24.27) | L′ (50) | S′ (50) | χ2 = 0.09, p = 0.763 |
| BDNF rs6265 (N = 309) | GG (60.84) | GA (33.66) | AA (5.50) | G (77.67) | A (22.33) | χ2 = 0.27, p = 0.763 |
| BDNF rs988748 (N = 309) | GG (55.99) | CG (37.22) | CC (6.80) | G (74.60) | C (25.40) | χ2 = 0.10, p = 0.750 |
| BDNF rs7103411 (N = 309) | TT (58.90) | TC (37.54) | CC (3.56) | T (77.67) | C (22.33) | χ2 = 2.09, p = 0.148 |
| BDNF rs1103014 (N = 309) | TT (55.99) | TC (35.92) | CC (8.09) | T (73.95) | C (26.05) | χ2 = 1.41, p = 0.234 |
| BDNF rs11757 (N = 309) | GG (55.02) | CG (37.54) | CC (7.44) | G (73.79) | C (26.21) | χ2 = 0.27, p = 0.603 |
| CTQ-SF Total Score | RFQ Total Score | ABS-2 Total Irrationality Score | GABS Total Irrationality Score | PHQ-9 Score | GAD-7 Score | |
|---|---|---|---|---|---|---|
| CTQ-SF total score | - | 0.80 ** | 0.15 ** | 0.15 ** | 0.35 ** | 039 ** |
| RFQ total score | - | 0.13 ** | 0.12 ** | 0.23 ** | 0.26 ** | |
| ABS-2 irrationality score | - | 0.87 ** | 0.42 ** | 0.47 ** | ||
| GABS irrationality score | - | 0.38 ** | 0.47 ** | |||
| PHQ-9 | - | 0.80 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Chiș, A.; Oltean, L.-E.; Bîlc, M.; Vulturar, R.; Șoflău, R.; David, D.; Szentágotai-Tătar, A.; Miu, A.C. Gene–Environment Interactions in Irrational Beliefs: The Roles of Childhood Adversity and Multiple Candidate Genes. Int. J. Mol. Sci. 2024, 25, 4206. https://doi.org/10.3390/ijms25084206
Chiș A, Oltean L-E, Bîlc M, Vulturar R, Șoflău R, David D, Szentágotai-Tătar A, Miu AC. Gene–Environment Interactions in Irrational Beliefs: The Roles of Childhood Adversity and Multiple Candidate Genes. International Journal of Molecular Sciences. 2024; 25(8):4206. https://doi.org/10.3390/ijms25084206
Chicago/Turabian StyleChiș, Adina, Lia-Ecaterina Oltean, Mirela Bîlc, Romana Vulturar, Radu Șoflău, Daniel David, Aurora Szentágotai-Tătar, and Andrei C. Miu. 2024. "Gene–Environment Interactions in Irrational Beliefs: The Roles of Childhood Adversity and Multiple Candidate Genes" International Journal of Molecular Sciences 25, no. 8: 4206. https://doi.org/10.3390/ijms25084206