
Vasorelaxant and Blood Pressure-Lowering Effects of Cnidium monnieri Fruit Ethanol Extract in Sprague Dawley and Spontaneously Hypertensive Rats

Abstract
1. Introduction
2. Results
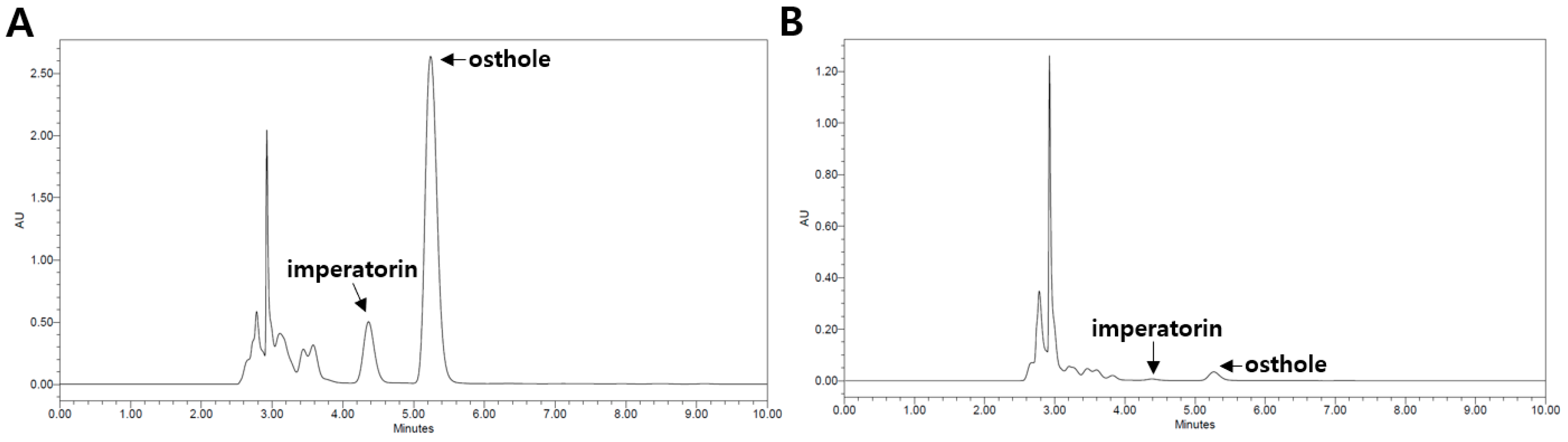
2.1. Qualitative and Quantitative High-Performance Liquid Chromatography (HPLC) Analysis of CM Extracts
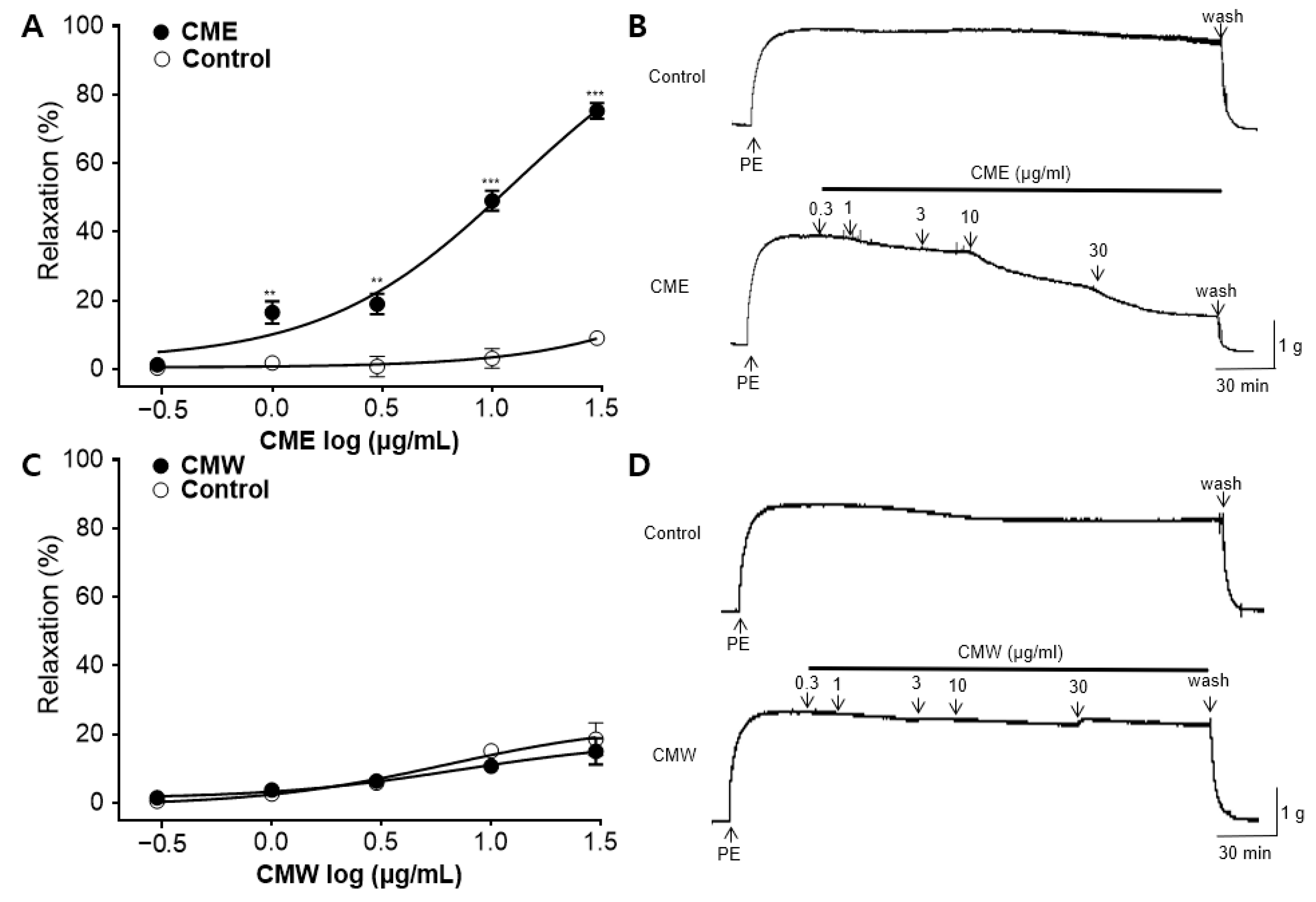
2.2. Vasorelaxant Effect of CME and CMW on Contractile Responses Induced by PE
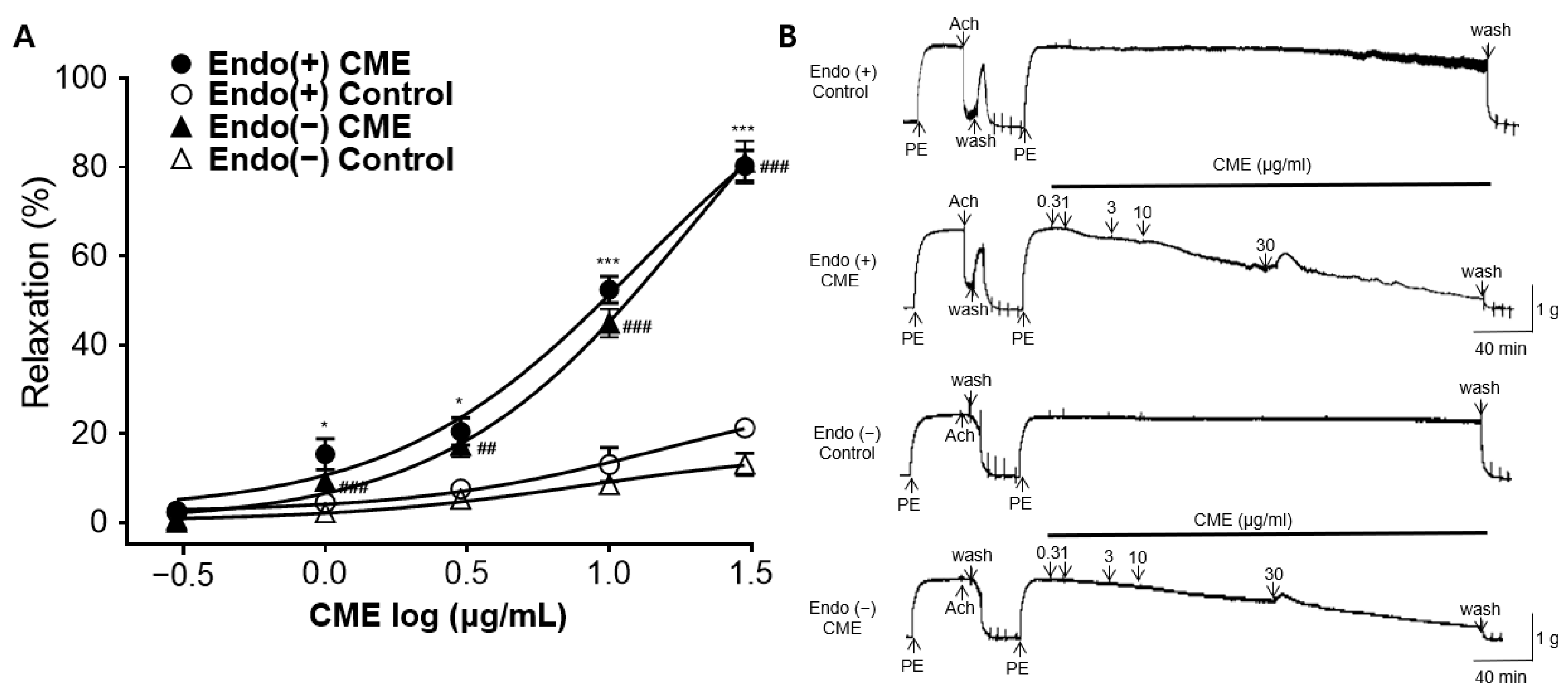
2.3. Vasorelaxant Effect of CME in Endothelium-Intact or Endothelium-Denuded Rat Aortic Rings
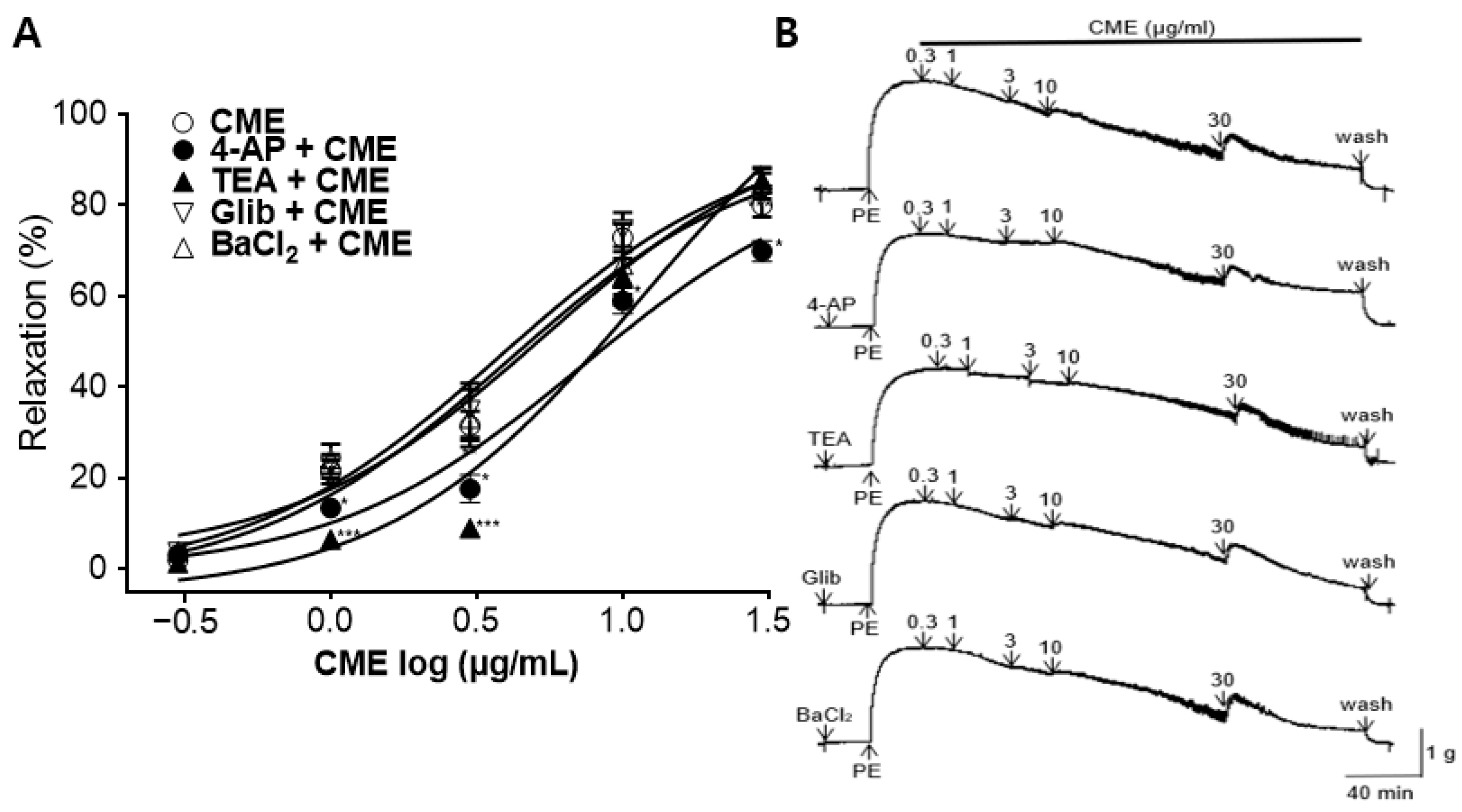
2.4. Vasorelaxant Effect of CME on Aortic Rings Pre-Incubated with K+ Channel Blockers
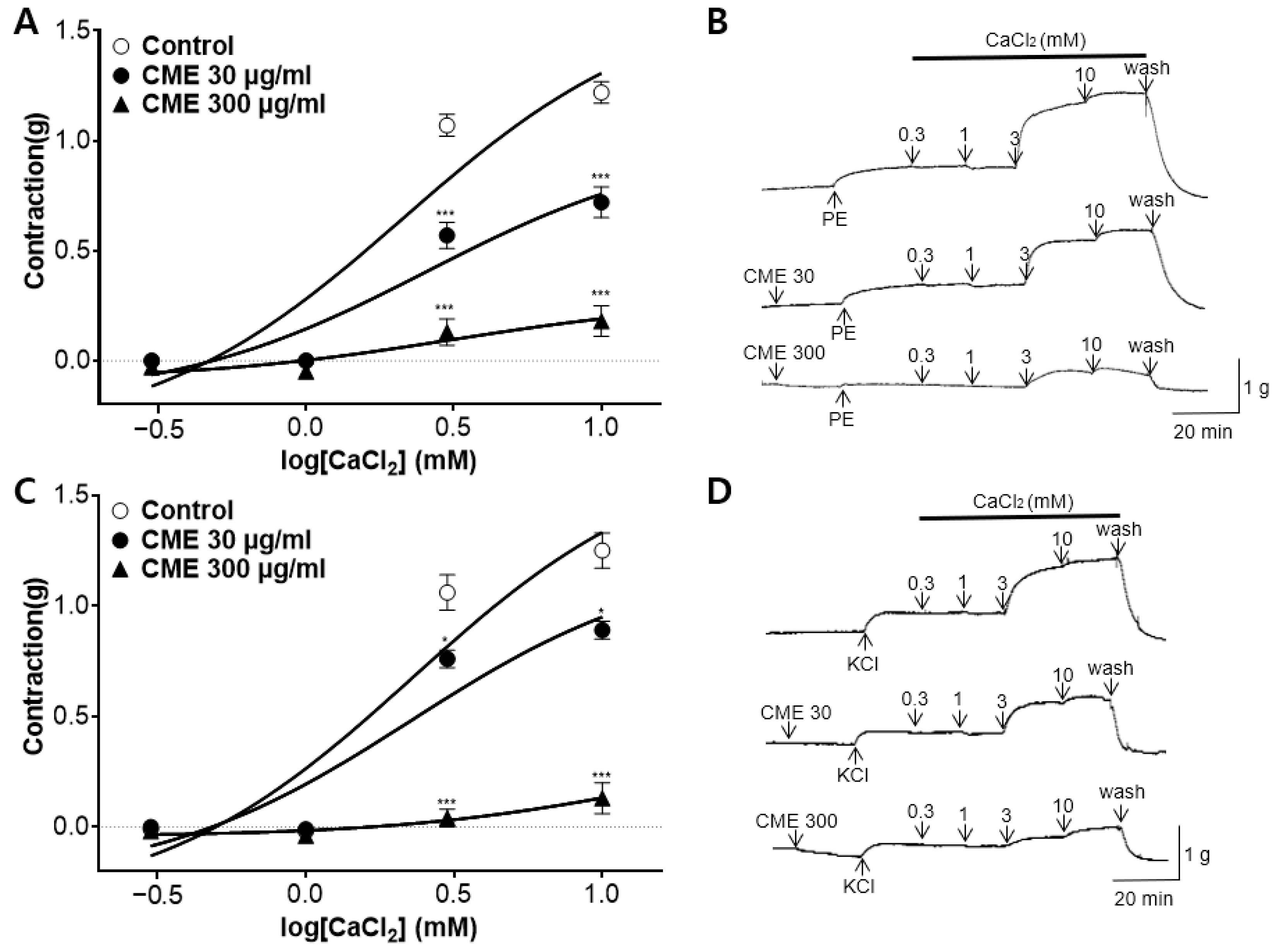
2.5. Effect of CME on Extracellular Ca2+-Induced Contraction Pre-Treated with PE and KCl
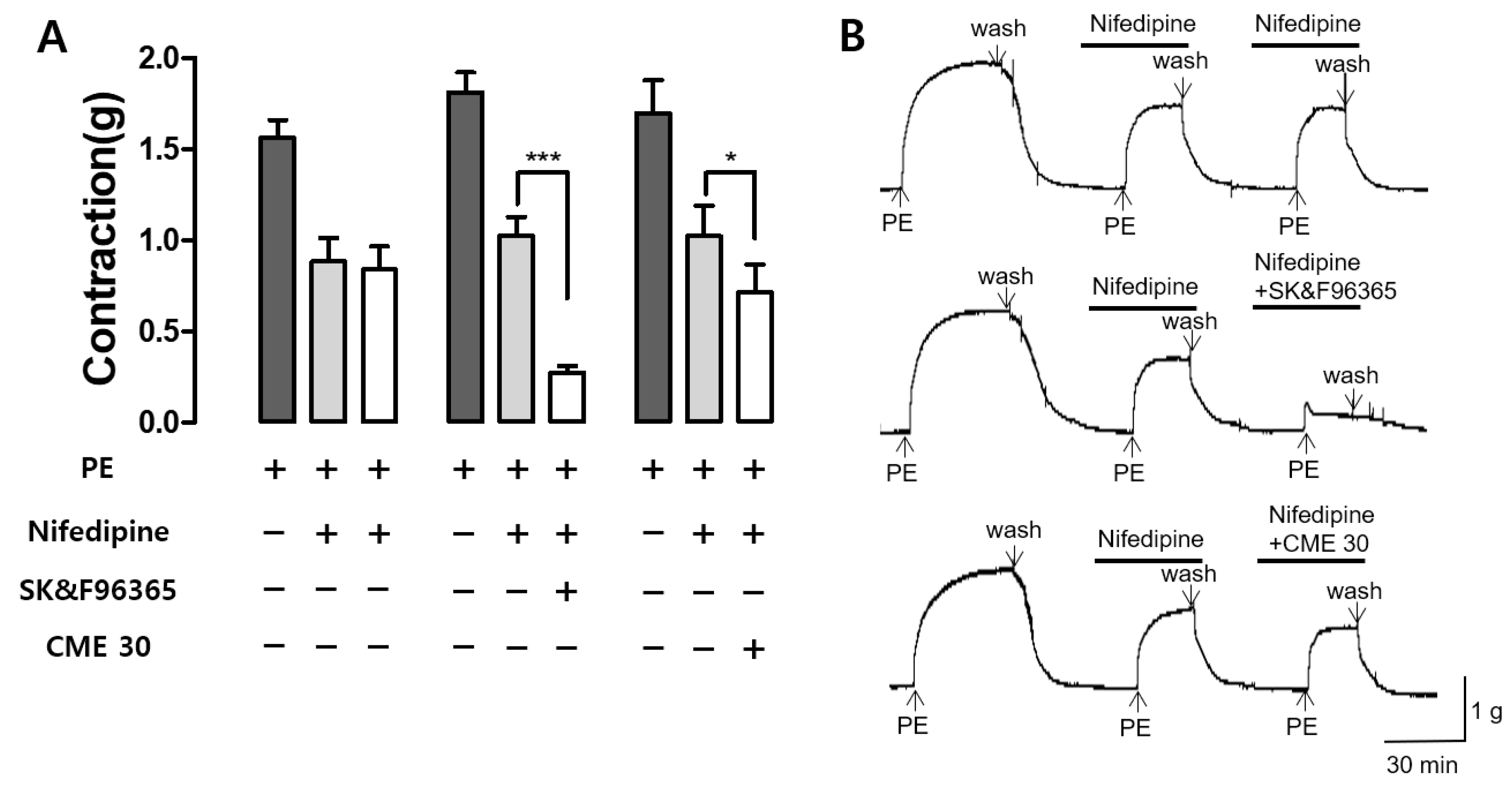
2.6. Effect of CME and SK&F96365 on PE-Induced Contraction in the Presence of Nifedipine
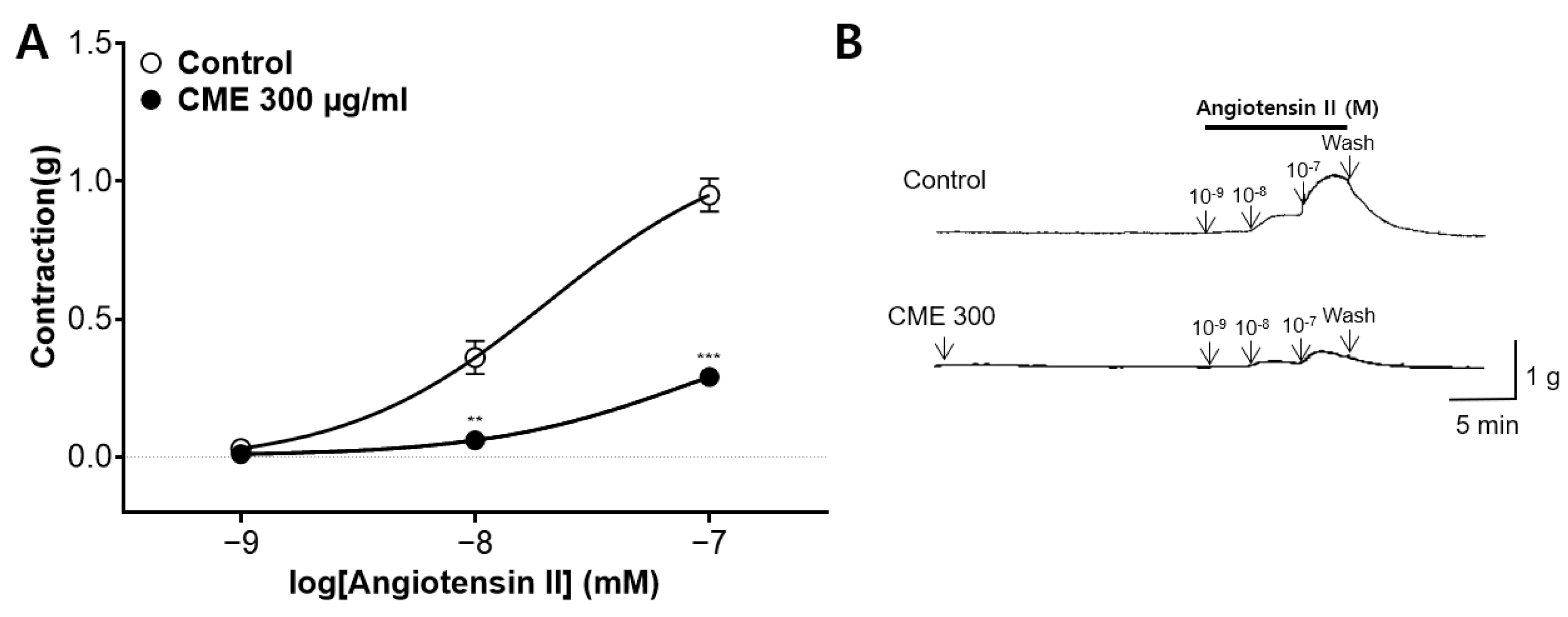
2.7. Inhibitory Effect of CME Pre-Treatment on Ang II-Induced Contraction
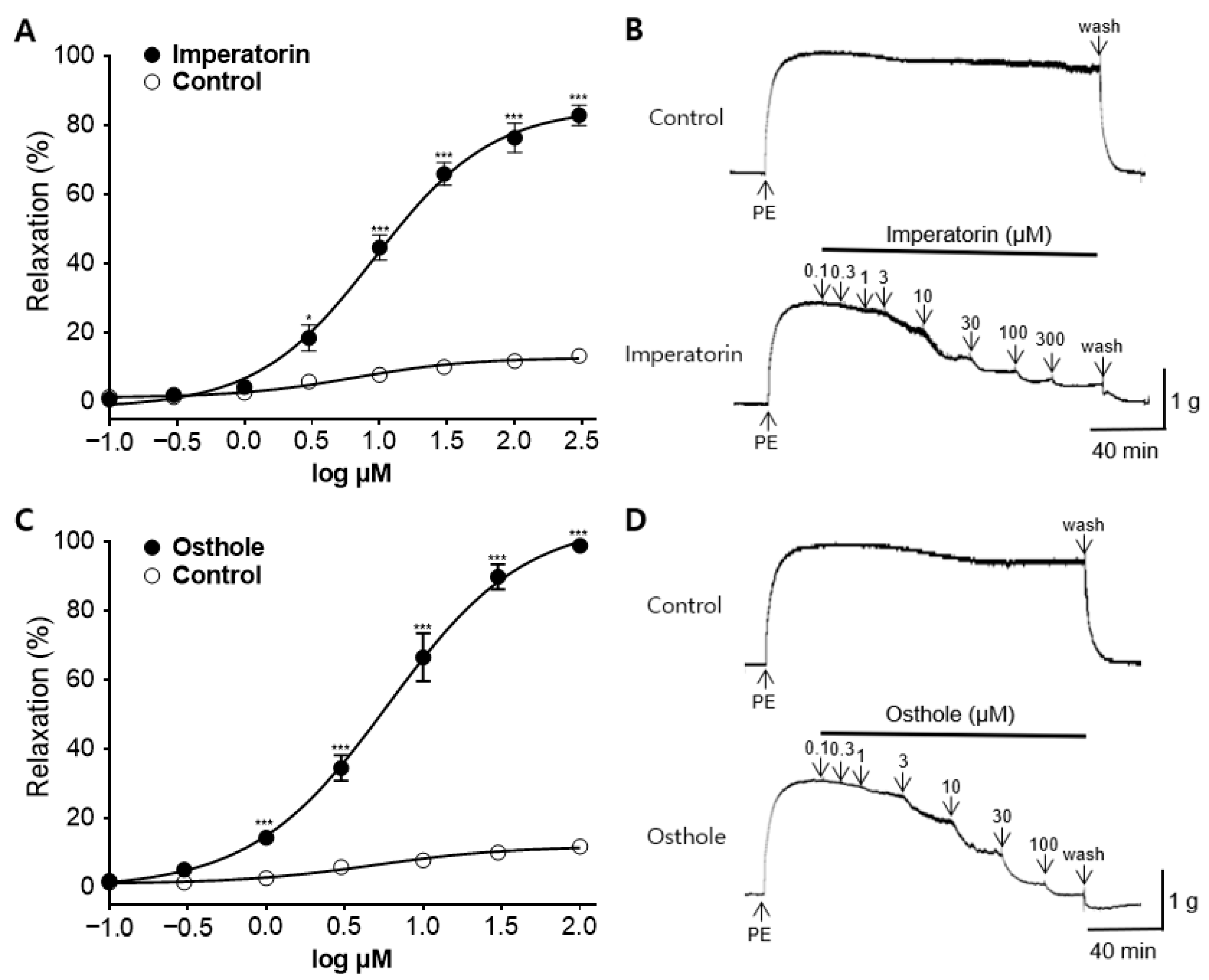
2.8. Vasorelaxant Effect of the Main Active Components in CME, Osthole, and Imperatorin
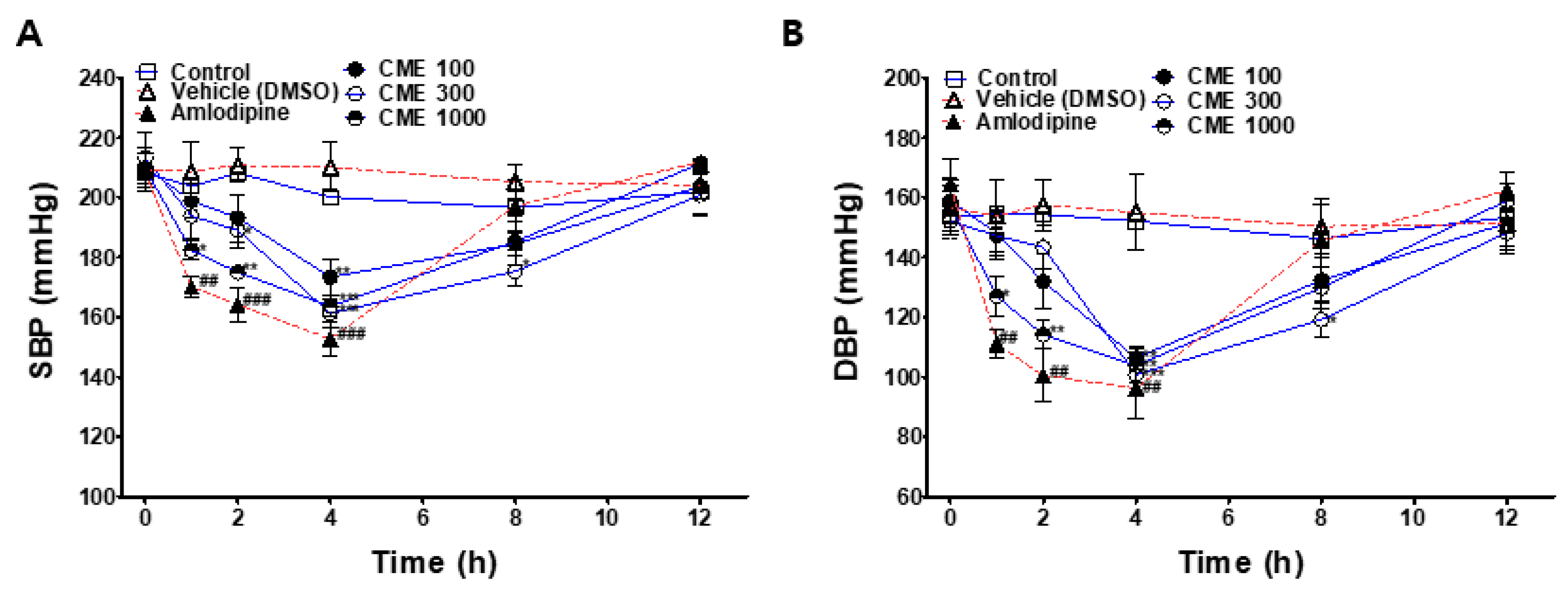
2.9. Hypotensive Effect of CME in SHR
3. Discussion
4. Materials and Methods
4.1. Materials and Chemicals
4.2. Sample Preparation
4.3. HPLC Analysis of CME and CMW
4.4. Animals
4.5. Confirmation of the Mechanism Associated with Vasodilatory Effect
4.5.1. Preparation of Rat Aortic Rings
4.5.2. Measurement of Tension in Rat Aortic Rings during PE-Induced Constriction
4.5.3. Role of CME on Endothelium-Intact and Endothelium-Denuded Rat Aortic Rings
4.5.4. Role of K+ Channel on the Effect of CME
4.5.5. Role of CME on Extracellular Ca2+-Induced Contraction
4.5.6. Role of CME in PE-Induced Contraction in the Presence of Nifedipine
4.5.7. Role of CME Pre-Treatment on Ang II-Induced Contraction
4.6. Measurement of Blood Pressure
4.7. Data Analysis
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Zhou, B.; Carrillo-Larco, R.M.; Danaei, G.; Riley, L.M.; Paciorek, C.J.; Stevens, G.A.; Gregg, E.W.; Bennett, J.E.; Solomon, B.; Singleton, R.K.; et al. (NCD-RisC), NCD Risk Factor Collaboration. Worldwide Trends in Hypertension Prevalence and Progress in Treatment and Control from 1990 to 2019: A Pooled Analysis of 1201 Population-Representative Studies with 104 million Participants. Lancet 2021, 398, 957–980. [Google Scholar] [CrossRef] [PubMed]
- Zhou, B.; Perel, P.; Mensah, G.A.; Ezzati, M. Global Epidemiology, Health Burden and Effective Interventions for Elevated Blood Pressure and Hypertension. Nat. Rev. Cardiol. 2021, 18, 785–802. [Google Scholar] [CrossRef]
- Przezak, A.; Bielka, W.; Pawlik, A. Hypertension and Type 2 Diabetes—The Novel Treatment Possibilities. Int. J. Mol. Sci. 2022, 23, 6500. [Google Scholar] [CrossRef] [PubMed]
- Bhatt, S.P.; Luqman-Arafath, T.K.; Guleria, R. Non-Pharmacological Management of Hypertension. Indian J. Med. Sci. 2007, 61, 616–624. [Google Scholar]
- Doroszko, A.; Janus, A.; Szahidewicz-Krupska, E.; Mazur, G.; Derkacz, A. Resistant Hypertension. Adv. Clin. Exp. Med. 2016, 25, 173–183. [Google Scholar] [CrossRef] [PubMed]
- Chrysant, S.G.; Chrysant, G.S. Herbs Used for the Treatment of Hypertension and Their Mechanism of Action. Curr. Hypertens. Rep. 2017, 19, 77. [Google Scholar] [CrossRef]
- Lee, S.; Jo, C.; Choi, H.Y.; Lee, K. Effect of Co-Administration of Curcumin with Amlodipine in Hypertension. Nutrients 2021, 13, 2797. [Google Scholar] [CrossRef] [PubMed]
- Jung, J.; Shin, S.; Park, J.; Lee, K.; Choi, H.Y. Hypotensive and Vasorelaxant Effects of Sanguisorbae Radix Ethanol Extract in Spontaneously Hypertensive and Sprague Dawley Rats. Nutrients 2023, 15, 4510. [Google Scholar] [CrossRef] [PubMed]
- Shin, S.; Park, J.; Choi, H.Y.; Bu, Y.; Lee, K. Sakuranetin as a Potential Regulator of Blood Pressure in Spontaneously Hypertensive Rats by Promoting Vasorelaxation through Calcium Channel Blockade. Biomedicines 2024, 12, 346. [Google Scholar] [CrossRef]
- Shaito, A.; Thuan, D.T.B.; Phu, H.T.; Nguyen, T.H.D.; Hasan, H.; Halabi, S.; Abdelhady, S.; Nasrallah, G.K.; Eid, A.H.; Pintus, G. Herbal Medicine for Cardiovascular Diseases: Efficacy, Mechanisms, and Safety. Front. Pharmacol. 2020, 11, 422. [Google Scholar] [CrossRef] [PubMed]
- Sameiyan, E.; Hayes, A.W.; Karimi, G. The Effect of Medicinal Plants on Multiple Drug Resistance through Autophagy: A Review of in Vitro Studies. Eur. J. Pharmacol. 2019, 852, 244–253. [Google Scholar] [CrossRef] [PubMed]
- Baharvand-Ahmadi, B.; Bahmani, M.; Tajeddini, P.; Rafieian-Kopaei, M.; Naghdi, N. An Ethnobotanical Study of Medicinal Plants Administered for the Treatment of Hypertension. J. Renal. Inj. Prev. 2016, 5, 123–128. [Google Scholar] [CrossRef] [PubMed]
- Kim, B.; Kwon, Y.; Lee, S.; Lee, K.; Ham, I.; Choi, H.Y. Vasorelaxant Effects of Angelica Decursiva Root on Isolated Rat Aortic Rings. BMC Complement. Altern. Med. 2017, 17, 474. [Google Scholar] [CrossRef] [PubMed]
- Lee, K.; Park, G.; Ham, I.; Yang, G.; Lee, M.; Bu, Y.; Kim, H.; Choi, H.Y. Vasorelaxant Effect of Osterici Radix Ethanol Extract on Rat Aortic Rings. Evid. Based Complement. Alternat. Med. 2013, 2013, 350964. [Google Scholar] [CrossRef] [PubMed]
- Kim, B.; Lee, K.; Chinannai, K.S.; Ham, I.; Bu, Y.; Kim, H.; Choi, H.Y. Endothelium-Independent Vasorelaxant Effect of Ligusticum jeholense Root and Rhizoma on Rat Thoracic Aorta. Molecules 2015, 20, 10721–10733. [Google Scholar] [CrossRef] [PubMed]
- Lee, K.; Shin, M.S.; Ham, I.; Choi, H.Y. Investigation of the Mechanisms of Angelica dahurica Root Extract-Induced Vasorelaxation in Isolated Rat Aortic Rings. BMC Complement. Altern. Med. 2015, 15, 395. [Google Scholar] [CrossRef] [PubMed]
- Kim, E.Y.; Rhyu, M.R. Synergistic Vasorelaxant and Antihypertensive Effects of Ligusticum wallichii and Angelica Gigas. J. Ethnopharmacol. 2010, 130, 545–551. [Google Scholar] [CrossRef] [PubMed]
- Testai, L.; Silvio, C.; Ammar, B.; Luisa, P.; Vincenzo, C.; Martinotti, E. Vasorelaxant Effects of the Chloroformic Crude Extract of Bupleurum fruticosum L. (Umbelliferae) Roots on Rat Thoracic Aorta. J. Ethnopharmacol. 2005, 96, 93–97. [Google Scholar] [CrossRef] [PubMed]
- Lee, J.W.; Roh, T.C.; Rho, M.C.; Kim, Y.K.; Lee, H.S. Mechanisms of Relaxant Action of a Pyranocoumarin from Peucedanum japonicum in Isolated Rat Thoracic Aorta. Planta Med. 2002, 68, 891–895. [Google Scholar] [CrossRef] [PubMed]
- Esmaeili, H.; Esmailidehaj, M.; Zarch, S.E.; Azizian, H. Role of the Potassium Channels in Vasorelaxant Effect of Asafoetida Essential Oil. Avicenna J. Phytomed. 2020, 10, 407–416. [Google Scholar] [PubMed]
- Sargazi Zadeh, G.; Panahi, N. Endothelium-Independent Vasorelaxant Activity of Trachyspermum Ammi Essential Oil on Rat Aorta. Clin. Exp. Hypertens. 2017, 39, 133–138. [Google Scholar] [CrossRef] [PubMed]
- Senejoux, F.; Demougeot, C.; Cuciureanu, M.; Miron, A.; Cuciureanu, R.; Berthelot, A.; Girard-Thernier, C. Vasorelaxant Effects and Mechanisms of Action of Heracleum sphondylium L. (Apiaceae) in Rat Thoracic Aorta. J. Ethnopharmacol. 2013, 147, 536–539. [Google Scholar] [CrossRef] [PubMed]
- Sun, Y.; Yang, A.W.H.; Lenon, G.B. Phytochemistry, Ethnopharmacology, Pharmacokinetics and Toxicology of Cnidium monnieri (L.) Cusson. Int. J. Mol. Sci. 2020, 21, 1006. [Google Scholar] [CrossRef] [PubMed]
- Chiou, W.F.; Huang, Y.L.; Chen, C.F.; Chen, C.C. Vasorelaxing Effect of Coumarins from Cnidium monnieri on Rabbit Corpus Cavernosum. Planta Med 2001, 67, 282–284. [Google Scholar] [CrossRef] [PubMed]
- Delong, C.; Sharma, S. Physiology, Peripheral Vascular Resistance. In StatPearls; StatPearls: Treasure Island, FL, USA. Available online: https://www.ncbi.nlm.nih.gov/books/NBK538308/ (accessed on 10 February 2024).
- He, Y.; Qu, S.; Wang, J.; He, X.; Lin, W.; Zhen, H.; Zhang, X. Neuroprotective Effects of Osthole Pretreatment against Traumatic Brain Injury in Rats. Brain Res. 2012, 1433, 127–136. [Google Scholar] [CrossRef] [PubMed]
- Chen, Y.H.; Guo, D.S.; Lu, M.H.; Yue, J.Y.; Liu, Y.; Shang, C.M.; An, D.R.; Zhao, M.M. Inhibitory Effect of Osthole from Cnidium monnieri on Tobacco Mosaic Virus (Tmv) Infection in Nicotiana glutinosa. Molecules 2019, 25, 65. [Google Scholar] [CrossRef] [PubMed]
- Li, R.; Song, P.; Tang, G.; Wei, J.; Rao, L.; Ma, L.; Jiang, M.; Huang, J.; Xu, Q.; Wu, J.; et al. Osthole Attenuates Macrophage Activation in Experimental Asthma by Inhibitingthe Nf-ĸb/Mif Signaling Pathway. Front. Pharmacol. 2021, 12, 572463. [Google Scholar] [CrossRef] [PubMed]
- Khan, S.A.; Lee, T.K.W. Network-Pharmacology-Based Study on Active Phytochemicals and Molecular Mechanism of Cnidium monnieri in Treating Hepatocellular Carcinoma. Int. J. Mol. Sci. 2022, 23, 5400. [Google Scholar] [CrossRef]
- Li, X.; Zhang, C.; Wu, E.; Han, L.; Deng, X.; Shi, Z. Uplc-Q-Tof/Ms-Based Metabolomics Approach Reveals Osthole Intervention in Breast Cancer 4t1 Cells. Int. J. Mol. Sci. 2023, 24, 1168. [Google Scholar] [CrossRef] [PubMed]
- Tsai, C.F.; Yeh, W.L.; Chen, J.H.; Lin, C.; Huang, S.S.; Lu, D.Y. Osthole Suppresses the Migratory Ability of Human Glioblastoma Multiforme Cells Via Inhibition of Focal Adhesion Kinase-Mediated Matrix Metalloproteinase-13 Expression. Int. J. Mol. Sci. 2014, 15, 3889–3903. [Google Scholar] [CrossRef] [PubMed]
- Chang, C.I.; Hu, W.C.; Shen, C.P.; Hsu, B.D.; Lin, W.Y.; Sung, P.J.; Wang, W.H.; Wu, J.B.; Kuo, Y.H. 8-Alkylcoumarins from the Fruits of Cnidium monnieri Protect against Hydrogen Peroxide Induced Oxidative Stress Damage. Int. J. Mol. Sci. 2014, 15, 4608–4618. [Google Scholar] [CrossRef]
- Lee, T.H.; Chen, Y.C.; Hwang, T.L.; Shu, C.W.; Sung, P.J.; Lim, Y.P.; Kuo, W.L.; Chen, J.J. New Coumarins and Anti-Inflammatory Constituents from the Fruits of Cnidium monnieri. Int. J. Mol. Sci. 2014, 15, 9566–9578. [Google Scholar] [CrossRef] [PubMed]
- Liang, H.J.; Suk, F.M.; Wang, C.K.; Hung, L.F.; Liu, D.Z.; Chen, N.Q.; Chen, Y.C.; Chang, C.C.; Liang, Y.C. Osthole, a Potential Antidiabetic Agent, Alleviates Hyperglycemia in Db/Db Mice. Chem. Biol. Interact. 2009, 181, 309–315. [Google Scholar] [CrossRef] [PubMed]
- Qin, L.P.; Zhang, J.Q.; Shi, H.P. Effects of Coumarins from Cnidium monnieri on the Function of Pituitary-Adrenocortical Axis in Kidney Yang Deficiency Rats. Zhongguo Zhong Xi Yi Jie He Za Zhi 1997, 17, 227–229. [Google Scholar]
- Dau, T.D.; Le, M.H.; Nguyen, T.T.G.H.; Dang, T.N.M.; Do, T.N.Q.; Tran, Q.T. Effects of Cnidium monnieri (L.) Cuss. Fruit Extract on Sexual Behaviors in Male Rats. Clin. Phytosci. 2020, 6, 80. [Google Scholar] [CrossRef]
- Flores-Morales, V.; Villasana-Ruíz, A.P.; Garza-Veloz, I.; González-Delgado, S.; Martinez-Fierro, M.L. Martinez-Fierro. Therapeutic Effects of Coumarins with Different Substitution Patterns. Molecules 2023, 28, 2413. [Google Scholar] [CrossRef]
- Huang, Y.; Sun, M.; Cui, H.; Kong, L.; Zhai, H.; Wang, Y.; Lü, C.; Fan, D. Vasorelaxant Effect of Osthole on Isolated Thoracic Aortic Rings in Rats. J. Tradit. Chin. Med. 2019, 39, 492–501. [Google Scholar]
- Zhang, Y.; Cao, Y.; Wang, Q.; Zheng, L.; Zhang, J.; He, L. A Potential Calcium Antagonist and Its Antihypertensive Effects. Fitoterapia 2011, 82, 988–996. [Google Scholar] [CrossRef] [PubMed]
- Zhang, Y.; Cao, Y.; Zhan, Y.; Duan, H.; He, L. Furanocoumarins-Imperatorin Inhibits Myocardial Hypertrophy Both in Vitro and in Vivo. Fitoterapia 2010, 81, 1188–1195. [Google Scholar] [CrossRef] [PubMed]
- Mónica, F.Z.; Bian, K.; Murad, F. The Endothelium-Dependent Nitric Oxide-Cgmp Pathway. Adv. Pharmacol. 2016, 77, 1–27. [Google Scholar]
- Moncada, S.; Vane, J.R. The Role of Prostacyclin in Vascular Tissue. Fed. Proc. 1979, 38, 66–71. [Google Scholar] [PubMed]
- Buchwalow, I.B.; Cacanyiova, S.; Neumann, J.; Samoilova, V.E.; Boecker, W.; Kristek, F. The Role of Arterial Smooth Muscle in Vasorelaxation. Biochem. Biophys. Res. Commun. 2008, 377, 504–507. [Google Scholar] [CrossRef] [PubMed]
- Sandoo, A.; van Zanten, J.J.; Metsios, G.S.; Carroll, D.; Kitas, G.D. The Endothelium and Its Role in Regulating Vascular Tone. Open Cardiovasc. Med. J. 2010, 4, 302–312. [Google Scholar] [CrossRef]
- Jackson, W.F. Potassium Channels in Regulation of Vascular Smooth Muscle Contraction and Growth. Adv. Pharmacol. 2017, 78, 89–144. [Google Scholar] [PubMed]
- Gurney, A.M.; Clapp, L.H. Calcium Channels and Vasodilation. In Advances in Molecular and Cell Biology; Bittar, E.E., Chen, L.B., Eds.; Elsevier: Amsterdam, The Netherlands, 1994; pp. 21–41. [Google Scholar]
- Huang, Y.; Ho, I.H. Separate Activation of Intracellular Ca2+ Release, Voltage-Dependent and Receptor-Operated Ca2+ Channels in the Rat Aorta. Chin. J. Physiol. 1996, 39, 1–8. [Google Scholar] [PubMed]
- Kim, J.E.; Choi, B.K.; Choi, J.Y.; Ryu, T.; Roh, W.S.; Song, S.Y. Role of Calcium Channels Responsible for Phenylephrine-Induced Contraction in Rat Aorta 3 Days after Acute Myocardial Infarction. Korean J. Anesthesiol. 2014, 66, 143–152. [Google Scholar] [CrossRef] [PubMed]
- Ko, F.N.; Wu, T.S.; Liou, M.J.; Huang, T.F.; Teng, C.M. Vasorelaxation of Rat Thoracic Aorta Caused by Osthole Isolated from Angelica pubescens. Eur. J. Pharmacol. 1992, 219, 29–34. [Google Scholar] [CrossRef] [PubMed]
- Vukelic, S.; Griendling, K.K. Angiotensin Ii, from Vasoconstrictor to Growth Factor: A Paradigm Shift. Circ. Res. 2014, 114, 754–757. [Google Scholar] [CrossRef] [PubMed]
- Yim, H.E.; Yoo, K.H. Renin-Angiotensin System—Considerations for Hypertension and Kidney. Electrolyte Blood Press 2008, 6, 42–50. [Google Scholar] [CrossRef] [PubMed]
- Tao, L.; Gu, X.; Xu, E.; Ren, S.; Zhang, L.; Liu, W.; Lin, X.; Yang, J.; Chen, C. Osthole Protects against Ang Ii-Induced Endotheliocyte Death by Targeting Nf-Κb Pathway and Keap-1/Nrf2 Pathway. Am. J. Transl. Res. 2019, 11, 142–159. [Google Scholar] [PubMed]
- He, J.-Y.; Zhang, W.; He, L.-C.; Cao, Y.-X. Imperatorin Induces Vasodilatation Possibly Via Inhibiting Voltage Dependent Calcium Channel and Receptor-Mediated Ca2+ Influx and Release. Eur. J. Pharmacol. 2007, 573, 170–175. [Google Scholar] [CrossRef] [PubMed]
- Wu, S.N.; Lo, Y.K.; Chen, C.C.; Li, H.F.; Chiang, H.T. Inhibitory Effect of the Plant-Extract Osthole on L-Type Calcium Current in Ng108-15 Neuronal Cells. Biochem. Pharmacol. 2002, 63, 199–206. [Google Scholar] [CrossRef] [PubMed]











{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Plant and Part | Solvent | Yield (%) | Abbreviation | Region |
|---|---|---|---|---|
| Cnidium monnieri (L.) Cusson Fruit | Water | 8.05 | CMW | China |
| Cnidium monnieri (L.) Cusson Fruit | 50% Ethanol | 8.10 | CME | China |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Park, J.; Shin, S.; Bu, Y.; Choi, H.-y.; Lee, K. Vasorelaxant and Blood Pressure-Lowering Effects of Cnidium monnieri Fruit Ethanol Extract in Sprague Dawley and Spontaneously Hypertensive Rats. Int. J. Mol. Sci. 2024, 25, 4223. https://doi.org/10.3390/ijms25084223
Park J, Shin S, Bu Y, Choi H-y, Lee K. Vasorelaxant and Blood Pressure-Lowering Effects of Cnidium monnieri Fruit Ethanol Extract in Sprague Dawley and Spontaneously Hypertensive Rats. International Journal of Molecular Sciences. 2024; 25(8):4223. https://doi.org/10.3390/ijms25084223
Chicago/Turabian StylePark, Junkyu, Sujin Shin, Youngmin Bu, Ho-young Choi, and Kyungjin Lee. 2024. "Vasorelaxant and Blood Pressure-Lowering Effects of Cnidium monnieri Fruit Ethanol Extract in Sprague Dawley and Spontaneously Hypertensive Rats" International Journal of Molecular Sciences 25, no. 8: 4223. https://doi.org/10.3390/ijms25084223
APA StylePark, J., Shin, S., Bu, Y., Choi, H.-y., & Lee, K. (2024). Vasorelaxant and Blood Pressure-Lowering Effects of Cnidium monnieri Fruit Ethanol Extract in Sprague Dawley and Spontaneously Hypertensive Rats. International Journal of Molecular Sciences, 25(8), 4223. https://doi.org/10.3390/ijms25084223