
Perineuronal Net Microscopy: From Brain Pathology to Artificial Intelligence

 , , ,
, , ,
Abstract
1. Introduction
2. Low-Resolution versus High-Resolution Microscopy in PNN Research
- (1)
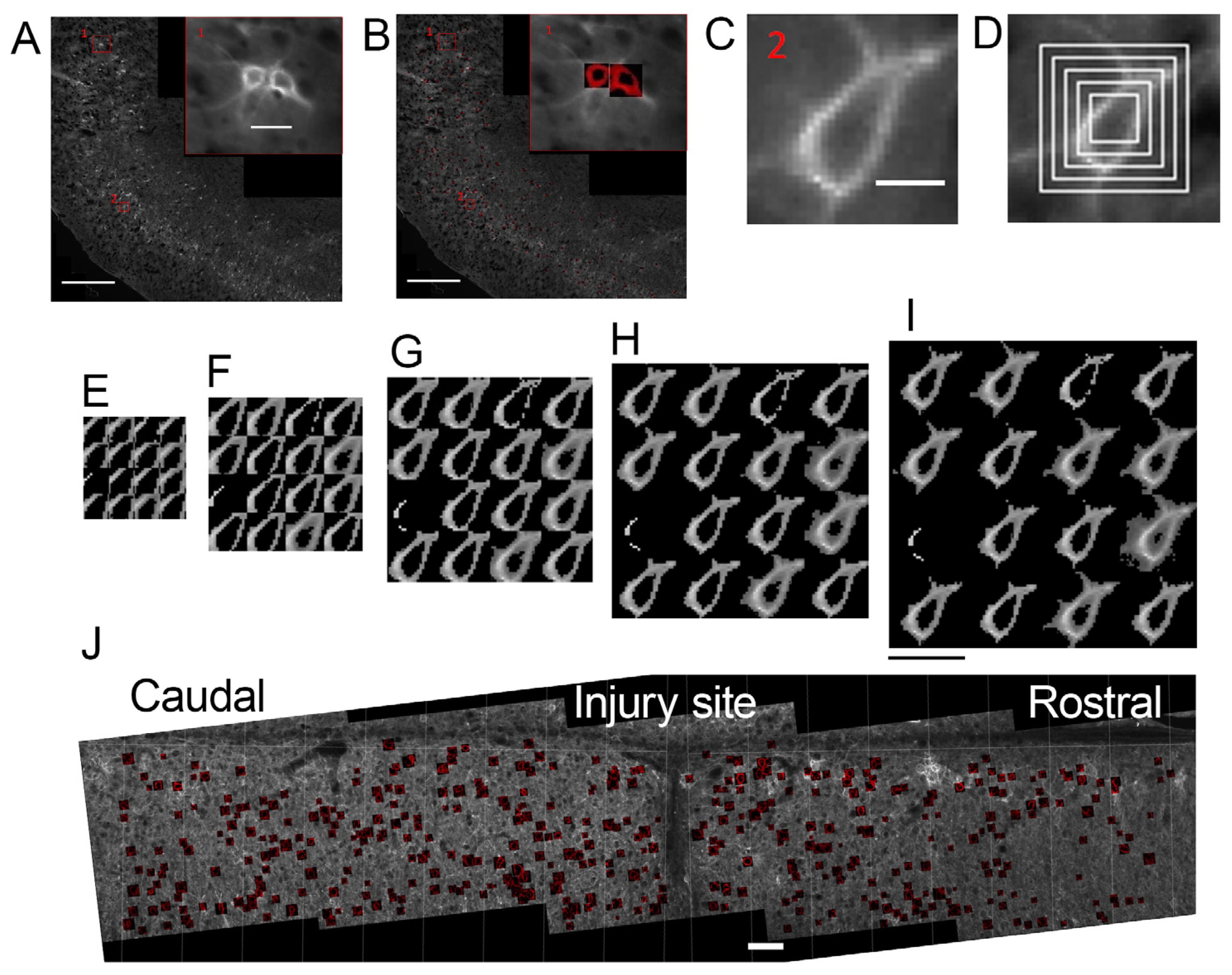
- The majority of experimental reports use low optical resolution light (mostly fluorescent) microscopy (10×, 20× objectives, NA within 0.25–0.8 range) to quantify the cell density of PNN+ neurons in tissue sections and to compare the staining intensity of the PNN-associated markers between experimental conditions (Table 1). This is a very important type of methodology and much of our knowledge about PNN structure and function was gained with the help of these techniques (reviewed in [4,10,22]). A number of image analysis tools were developed for the quantification of PNN parameters in low-resolution images [32,33,34,35] (Figure 2). Recently, Lupori and co-authors published “A comprehensive atlas of perineuronal net distribution and colocalization with parvalbumin in the adult mouse brain” [36], raising PNN microscopy studies to a remarkable new level and suggesting new opportunities for high-content structural and functional studies of the brain ECM, as discussed below (review Section 10). Essentially, in this case, machine learning generated a large amount of PNN+ cell annotation data that was spatially resolved and could be transferred to standardized brain atlas coordinates. Thus, among other interesting options, the approach allows for a systematic comparison of the PNN distribution to brain connectomics and spatial transcriptomics data [37].
- (2)
- A smaller number of reports addressed high-resolution structure of single PNN meshes (or single PNN units)—polygonal or round barriers consisting of ECM molecules and surrounding individual synapses [14,20,38,39,40,41,42,43,44] (Table 1). These studies revealed another level of the PNN microstructure, shedding light on the delicate architecture of single synapses and their ECM coat at the sub-micrometer scale. Confocal microscopy was performed with NA = 1.4, allowing for a higher optical resolution [14,20,42]; higher resolutions were obtained with Superresolution Structured Illumination Microscopy (SR-SIM) (Zeiss, Oberkochen, Germany), stimulated emission depletion (STED), stochastic optical reconstruction microscopy (STORM) (Nikon, Tokyo, Japan), AiryScan (Zeiss, Oberkochen, Germany) super-resolution [39,40,44] and electron microscopy (Zeiss, Oberkochen, Germany) [38,43].

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Method | Disease (Model) or Manipulation | References | Markers |
|---|---|---|---|
| Non-fluorescent light microscopy | Normal brain and spinal cord | [9,45,46,47,48]; [49] (×40); [50,51] | Neurocan, Cat-301, versican, phosphacan, WFA, PV, HABP, TN-R, aggrecan, Sema3A, Sema3B, neurocan, brevican, Crtl1, NG2, APC, GFAP, NeuN, HAPLN1, CD44, BRAL2 |
| Alzheimer’s disease (AD) | [52] (×10; ×20) | Wisteria floribunda agglutinin (WFA) | |
| Schizophrenia | [53]; [54] (×1.6; ×40); [55] (×2.5/0.12; ×20/0.5); [56] | WFA, Aggrecan (Cat 301), CS56, 3B3, GFAP, ACAN | |
| Crtl1/Hapln1 deficiency | [57] | WFA | |
| TauP301L—Acan mouse model | [58] | Aggrecan, ChAT | |
| Dementia | [59,60] | WFA, Cat-316, Sema3A, NeuN | |
| Sleep deprivation | [61] (×40) | WFA | |
| Substance use disorder | [62] (×20; ×40) | WFA | |
| Monocular deprivation | [63] | Cat-315, Crtl-1 | |
| Spinal cord injury | [64] | WFA, 2B6 | |
| Epifluorescent microsopy | Normal brain | [65]; [33] (×10/0.6; ×20/0.8); [49] (×40); [36] (×10) | WFA, Kv3.1b, Cat-301, Neurocan, brevican, versican, phosphacan, TN-R, HABP, aggrecan, PV, GFAP |
| In vitro modeling of PNNs | [66] | WFA, Has-3, aggrecan, Crtl1 | |
| Sema3A binding to the PNNs | [67] | WFA | |
| Spinal cord injury | [68]; [33] (×10/0.6; ×20/0.8); [69] | WFA, PV, NeuN, aggrecan, Crtl1, ChAT, HABP | |
| tPA deficiency in FS-PV interneurons | [70] | WFA, PV, GABA, NeuN, Iba1 | |
| Perinatal penicillin exposure | [71] (×10/0.45) | WFA, PV | |
| Substance use disorder | [72] (×10; ×20; ×40); [73] | WFA | |
| Hibernation | [74] | WFA | |
| Epilepsy | [75] (×10) | WFA, PV, Cat-315 | |
| Schizophrenia | [76] (×20/0.75; ×60/1.4); [77] (×40) | WFA, PV, aggrecan, NeuN, 8-oxo-DG | |
| AD | [78] | WFA, PV, PCP4 | |
| Neuropilin1-Fc injection to visual cortex | [79] (×20/0.5) | WFA, PV, Sema-3A | |
| Ptprz1 deficiency | [11] (×10; ×20; ×63) | WFA, aggrecan, HAPLN1, neurocan, brevican, tenascin-R 619, phosphacan | |
| 4-methylumbelliferone treatment | [80] (×20) | WFA | |
| Purkinje Cell Degeneration | [81] (×10; ×63/1.4) | Aggrecan, GAD 65/67, vGlut1, vGlut2, brevican, Haplnq, Hapln4, HABP, TN-R, GFAP | |
| Ventral hippocampal PNN depletion | [82] | WFA, PV | |
| Monocular deprivation | [21] (×20; ×40), [83] (×20); [84,85] | WFA, neurocan, PV | |
| Confocal microsopy | Normal adult brain | [38]; [14] (×5/0.16; ×63/1.40); [32] (×20/0.7); [45] (×20), [86] (×20), [87]; [48] (×40), [88] (×63); [50] (×20), [51]; [89] (×40/1.1); [90] (×40) | WFA, Sema3A, SV2, GAD67, aggrecan, versican, phosphacan, TN-R, PV, NeuN, ChAT, neurocan, brevican, calbindin, C6S, GlyT2, vGlut1, Hapln1, GlycR, GABAaR, substance P, PSD95, Ankyrin G, Cat-315 |
| Enriched environment | [91] (×100/1.4); [92,93] | WFA, PV, GAD67, Aggrecan, Neurocan, VGlut1, Sema3A, calbindin, VGlut2, SMI32 | |
| Co-culture of hippocampal neurons and cortical astrocytes | [94] | Aggrecan, vGlut, PSD-95, VGAT, gephyrin | |
| lenti-cmv-Nptx2-myc injection to somatosensory cortex | [95] (×40) | WFA, NeuN, PV | |
| Eyeblink conditioning | [25] (×63) | WFA, VGAT, gephyrine, NeuN, aggrecan | |
| AD | [52] (×20); [96] (×20, ×63); [97] (×10/0.3; ×63/1.4); [98] | WFA, Aβ (Amylo-Glo), CD68, Iba1, Thioflavin-S, PV, Aggrecan, Crtl1, GAD65/67, vGlut1, Cat-301, calretinin, MAP2, VGAT, brevican | |
| Schizophrenia, bipolar disorder | [42] (×20/0.5; ×63/1.4); [99]; [100] (×20/0.5); [101] (×20; ×40) | WFA, PV, HNK-1, S100-β, CS56, MMP9, 8-oxo-dG, CD68, Iba1 | |
| Substance use disorder | [102] (×20/0.7; ×63/1.4); [103] (×20/0.75; ×20/0.7); [104] (×40); [105] (×63/1.4); [106] (×40), [107]; [32] (×20/0.7); [62] | WFA, PV, GAD65/67, VGlut1, c-Fos, Calretinin, mGluR2, SMI32, SYN1 | |
| Amyotrophic lateral sclerosis (ALS) | [108] | WFA, aggrecan, NeuN | |
| Dementia | [60] | WFA, HAPLN1, 6B4, 7B7 Cat-316, Sema3A | |
| Epilepsy | [109]; [75] (×10; ×100) | WFA, PV, Cat-315, GFAP | |
| Huntington’s disease | [96] (×20) | WFA, Iba1, PV | |
| Cartilage matrix deficiency | [110] (×63) | WFA, aggrecan, GABA, PV, Hapln1, brevican, tenascin R, versican, phosphacan, HABP | |
| tPA deficiency in FS-PV interneurons | [70] (×40/1.44) | WFA, PV, aggrecan | |
| Deletion Npy1r in forebrain excitatory neurons | [111] (×40/1) | WFA, aggrecan, PV, c-Fos, NeuN | |
| Acan gene deletion | [112] (×10; ×63); [113] (×10/0.45; ×63/1.4) | Aggrecan, WFA, Tn-R, versican, neurocan, Ctrl-1, brevican, phosphacan, Bral2, PV | |
| Brevican gene deletion | [114] (×63/1.2) | Brevican, aggrecan, neurocan, HAPLN1, calbindin, CtBP2, HAPLN4, vGlut3, Cav1.3, CtBP2, GluR2/3, GluR4, MBP, SMI32 | |
| Monocular deprivation | [115] (×10/0.45) | WFA, PV | |
| Fear conditioning | [116] (×40); [117] (×40/1.4); [118] | WFA, Hapln1, PV, Zif268 | |
| Oxidative stress | [119] (×20; ×40; ×63); [120] | WFA, PV, 8-oxo-dG, calbindin, calretinin, Lipofuscin, SMI 311, CSPG | |
| Fluoxetine treatment | [121]; [122] (×10/0.45); [123] | WFA, PV | |
| Anxiety (maternal separation with early weaning) | [124] (×20; ×63/1.4) | WFA, PV, OTX2, SST, CR | |
| Tetrodoxin, NBQX, diltiazem treatment | [125] | WFA, PV, tenascin-R (monoclonal a/b 596), Aggrecan, HABP, NeuN, Synbrev, GFAP, VGAT | |
| PLX3397 treatment | [126] (×10/0.3; ×63/1.4) | WFA, PV, versican | |
| Somatosensory deprivation (whisker shaving model) | [20] (×63/1.4) | WFA, VGAT | |
| Enriched environment, cartilage LP1 deficiency | [127] (×63) | WFA, SMI32, HABP, calbindin | |
| Deletion of chondroitin 6-sulfotransferase (chst3) | [128] (×63) | WFA, PV | |
| Poly I:C injection during gestation | [129] | Aggrecan, vGlut, PSD-95 | |
| Tenascin-C, tenascin-R, brevican, neurocan deficiency | [130] (×63) | Aggrecan, PSD95, vGlut1, VGAT, gephyrin, NF200, WFA | |
| tenascin-R deletion | [131] | WFA, TN-R, PV, ChAT, aggrecan, NeuN, TN-C | |
| Early social isolation | [132] (×10) | WFA, PV | |
| Social disfunction model | [133] (×20; ×63) | WFA, PCP4, OTX2, PV, RGS14 | |
| Unilateral labyrinthectomy | [134] (×63) | WFA, SMI32, NeuN, brevican, | |
| PNN removal | [135] (×4/0.2; ×60/1.4); [136] (×10/0.4); [137]; [138] | WFA, vGlut1, vGlut2, VGAT, PV, aggrecan, versican, brevican, neurocan, phosphacan, proteoglycan Di-4S (2B6) | |
| Spinal cord injury | [139] (×40, ×63); [140] | WFA, ChAT, NeuN, β-III Tubulin, 5-HT, Iba1, GFAP, Cat-301 | |
| Multiphoton microscopy | Normal brain | [89] (×10/0.6; ×25/0.95) | WFA |
| Super-resolution microscopy | Ischemia | [39] (×20/0.8; ×100/1.46); [40] (×10/0.45; ×20/0.8; ×100/1.46) | WFA, Iba1, GFAP, PV, Kv3.1, VGAT, VGluT1, aggrecan |
| Rett syndrome | [41] (×60/1.4) | WFA, synaptotagmin-2, PV, VGLUT2 | |
| Pain | [44] (×63/1.4) | Aggrecan, Pax2, NeuN, VGAT, VGLUT2, Gephyrin, c-Fos, WFA, CD68, Iba1 | |
| Electron microscopy | Normal brain | [38,43] | WFA |
| Enriched environment | [91] | WFA | |
| AD | [98] | Brevican, aggrecan (HAG7D4) | |
| Hibernation-like state | [113] | ||
| AI-assisted | Normal brain | [34,36] | WFA, parvalbumin |
3. Normal Brain Functions Addressed with PNN Microscopy
3.1. PNN in Fear and Memory
3.2. Metal Binding
4. PNN Structural Studies in Brain Pathology
4.1. PNN Structural Studies in Schizophrenia
4.2. Epilepsy
4.3. Alzheimer’s Disease
4.4. Drug Abuse
4.5. Spinal Cord Injury
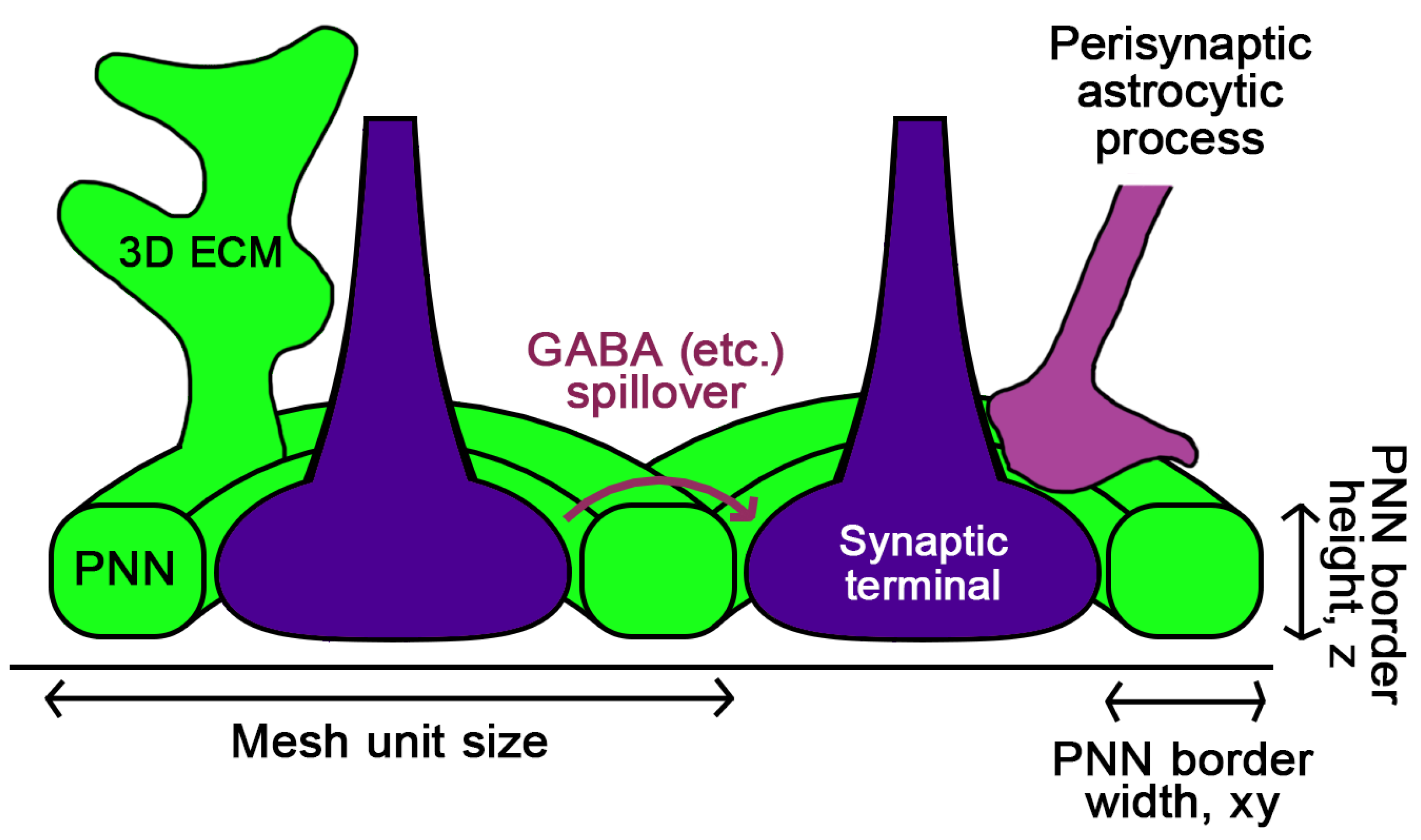
5. How Could the PNN Mesh Geometry Affect the Synapse?
5.1. The Mesh Area
5.2. The Mesh 3D Thickness
5.3. The Intersynaptic Layer Width
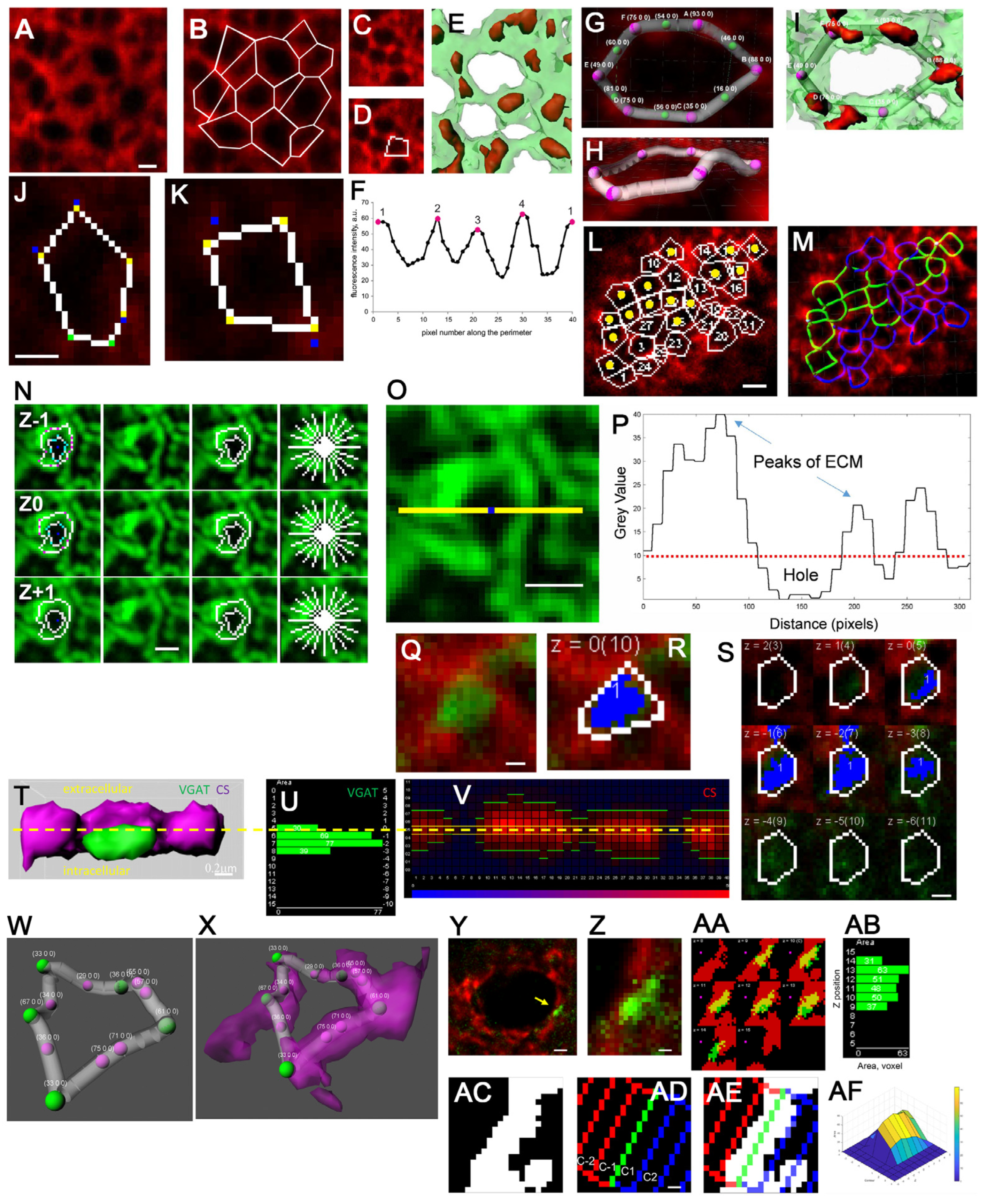
6. PNN Single-Mesh Studies
7. Perineuronal Net as a Potential Drug Target
8. Future Methodological Perspective for PNN Microscopy
8.1. Multiphoton Microscopy
8.2. Super-Resolution Microscopy
8.3. Electron Microscopy
8.4. Technical Aspects of Introducing AI Tools in Biomedical Research
9. AI Tools in Brain Pathology Studies
10. AI Tools for PNN Studies
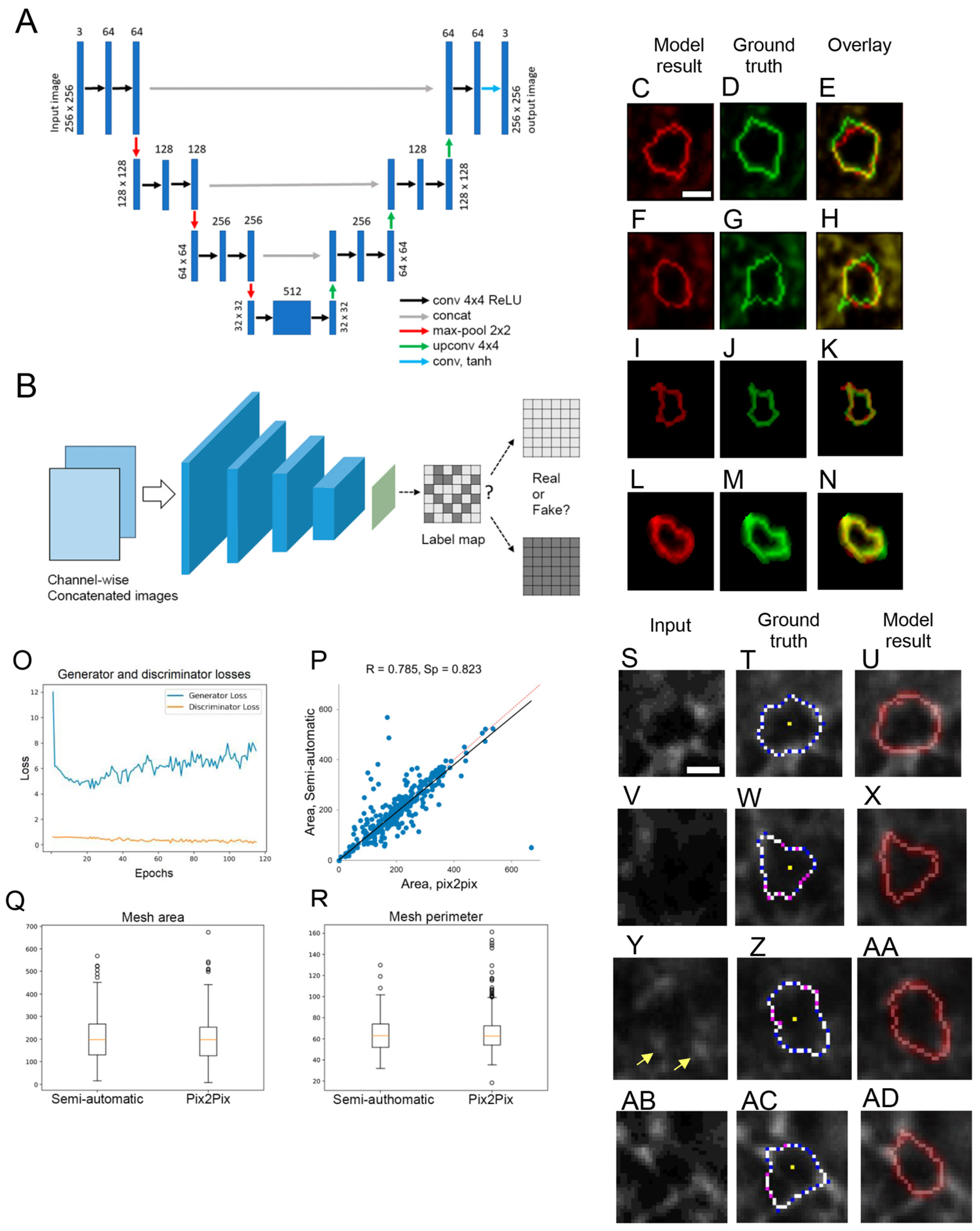
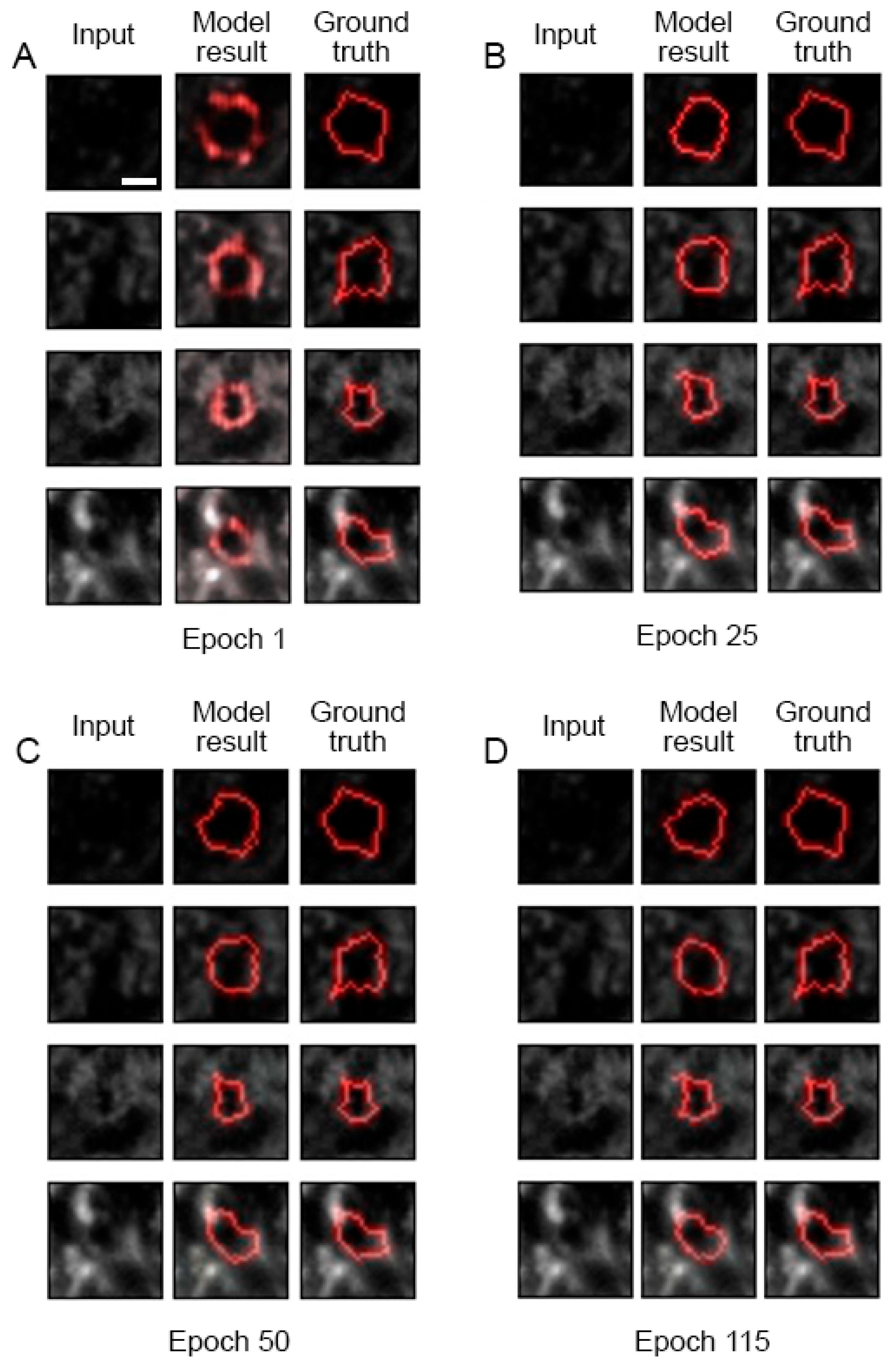
11. AI-Assisted PNN Mesh Tracing
Results and Discussion
12. Conclusions
- -
- Implementation of high-throughput instrumental upgrades both in low and high-resolution microscopy to speed up the pipeline for the data collection;
- -
- Transition from low-resolution microscopy meant for counting PNN numbers to high-resolution imaging aiming at insights into synaptic structure and function. Superresolution microscopy, multiphoton microscopy, correlative light–electron microscopy (CLEM) and electron tomography are instrumental for efficient progress along these lines;
- -
- Expanding the range of quantitative image analysis methods in order to increase collected structural information and build high-resolution 3D models elucidating the structural basis of physiological functions and brain pathologies. Integrating the PNN data into connectomics research may be particularly fruitful;
- -
- Implementation of AI instruments aimed at high-content unbiased quantitative microscopy data analysis and achieving new unprecedented levels of insight into PNN structure and function.
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Fawcett, J.W.; Oohashi, T.; Pizzorusso, T. The roles of perineuronal nets and the perinodal extracellular matrix in neuronal function. Nat. Rev. Neurosci. 2019, 20, 451–465. [Google Scholar] [CrossRef] [PubMed]
- Dong, Y.; Zhao, K.; Qin, X.; Du, G.; Gao, L. The mechanisms of perineuronal net abnormalities in contributing aging and neurological diseases. Ageing Res. Rev. 2023, 92, 102092. [Google Scholar] [CrossRef] [PubMed]
- Pantazopoulos, H.; Katsel, P.; Haroutunian, V.; Chelini, G.; Klengel, T.; Berretta, S. Molecular signature of extracellular matrix pathology in schizophrenia. Eur. J. Neurosci. 2021, 53, 3960–3987. [Google Scholar] [CrossRef]
- Carceller, H.; Gramuntell, Y.; Klimczak, P.; Nacher, J. Perineuronal Nets: Subtle Structures with Large Implications. Neuroscientist 2023, 29, 569–590. [Google Scholar] [CrossRef] [PubMed]
- Zeug, A.; Stawarski, M.; Bieganska, K.; Korotchenko, S.; Wlodarczyk, J.; Dityatev, A.; Ponimaskin, E. Current microscopic methods for the neural ECM analysis. In Progress in Brain Research; Elsevier: Amsterdam, The Netherlands, 2014; Volume 214, pp. 287–312. [Google Scholar] [CrossRef]
- Celio, M.R.; Spreafico, R.; De Biasi, S.; Vitellaro-Zuccarello, L. Perineuronal nets: Past and present. Trends Neurosci. 1998, 21, 510–515. [Google Scholar] [CrossRef] [PubMed]
- Ruoslahti, E. Brain extracellular matrix. Glycobiology 1996, 6, 489–492. [Google Scholar] [CrossRef]
- Yamaguchi, Y. Lecticans: Organizers of the brain extracellular matrix. Cell. Mol. Life Sci. 2000, 57, 276–289. [Google Scholar] [CrossRef]
- Deepa, S.S.; Carulli, D.; Galtrey, C.; Rhodes, K.; Fukuda, J.; Mikami, T.; Sugahara, K.; Fawcett, J.W. Composition of Perineuronal Net Extracellular Matrix in Rat Brain. J. Biol. Chem. 2006, 281, 17789–17800. [Google Scholar] [CrossRef] [PubMed]
- Kwok, J.C.F.; Dick, G.; Wang, D.; Fawcett, J.W. Extracellular matrix and perineuronal nets in CNS repair. Dev. Neurobiol. 2011, 71, 1073–1089. [Google Scholar] [CrossRef]
- Eill, G.J.; Sinha, A.; Morawski, M.; Viapiano, M.S.; Matthews, R.T. The protein tyrosine phosphatase RPTPζ/phosphacan is critical for perineuronal net structure. J. Biol. Chem. 2020, 295, 955–968. [Google Scholar] [CrossRef]
- Carulli, D.; De Winter, F.; Verhaagen, J. Semaphorins in Adult Nervous System Plasticity and Disease. Front. Synaptic Neurosci. 2021, 13, 672891. [Google Scholar] [CrossRef]
- Ferrer-Ferrer, M.; Dityatev, A. Shaping Synapses by the Neural Extracellular Matrix. Front. Neuroanat. 2018, 12, 40. [Google Scholar] [CrossRef]
- Arnst, N.; Kuznetsova, S.; Lipachev, N.; Shaikhutdinov, N.; Melnikova, A.; Mavlikeev, M.; Uvarov, P.; Baltina, T.V.; Rauvala, H.; Osin, Y.N.; et al. Spatial patterns and cell surface clusters in perineuronal nets. Brain Res. 2016, 1648, 214–223. [Google Scholar] [CrossRef] [PubMed]
- Bosiacki, M.; Gąssowska-Dobrowolska, M.; Kojder, K.; Fabiańska, M.; Jeżewski, D.; Gutowska, I.; Lubkowska, A. Perineuronal Nets and Their Role in Synaptic Homeostasis. Int. J. Mol. Sci. 2019, 20, 4108. [Google Scholar] [CrossRef]
- Lander, C.; Zhang, H.; Hockfield, S. Neurons Produce a Neuronal Cell Surface-Associated Chondroitin Sulfate Proteoglycan. J. Neurosci. 1998, 18, 174–183. [Google Scholar] [CrossRef]
- Brückner, G.; Kacza, J.; Grosche, J. Perineuronal Nets Characterized by Vital Labelling, Confocal and Electron Microscopy in Organotypic Slice Cultures of Rat Parietal Cortex and Hippocampus. J. Mol. Histol. 2003, 35, 115–122. [Google Scholar] [CrossRef] [PubMed]
- McRae, P.A.; Rocco, M.M.; Kelly, G.; Brumberg, J.C.; Matthews, R.T. Sensory Deprivation Alters Aggrecan and Perineuronal Net Expression in the Mouse Barrel Cortex. J. Neurosci. 2007, 27, 5405–5413. [Google Scholar] [CrossRef]
- Carulli, D.; Pizzorusso, T.; Kwok, J.C.F.; Putignano, E.; Poli, A.; Forostyak, S.; Andrews, M.R.; Deepa, S.S.; Glant, T.T.; Fawcett, J.W. Animals lacking link protein have attenuated perineuronal nets and persistent plasticity. Brain 2010, 133, 2331–2347. [Google Scholar] [CrossRef] [PubMed]
- Lipachev, N.; Melnikova, A.; Fedosimova, S.; Arnst, N.; Kochneva, A.; Shaikhutdinov, N.; Dvoeglazova, A.; Titova, A.; Mavlikeev, M.; Aganov, A.; et al. Postnatal development of the microstructure of cortical GABAergic synapses and perineuronal nets requires sensory input. Neurosci. Res. 2022, 182, 32–40. [Google Scholar] [CrossRef] [PubMed]
- Pizzorusso, T.; Medini, P.; Berardi, N.; Chierzi, S.; Fawcett, J.W.; Maffei, L. Reactivation of Ocular Dominance Plasticity in the Adult Visual Cortex. Science 2002, 298, 1248–1251. [Google Scholar] [CrossRef]
- Miyata, S.; Kitagawa, H. Mechanisms for modulation of neural plasticity and axon regeneration by chondroitin sulphate. J. Biochem. 2015, 157, 13–22. [Google Scholar] [CrossRef] [PubMed]
- Hou, X.; Yoshioka, N.; Tsukano, H.; Sakai, A.; Miyata, S.; Watanabe, Y.; Yanagawa, Y.; Sakimura, K.; Takeuchi, K.; Kitagawa, H.; et al. Chondroitin Sulfate Is Required for Onset and Offset of Critical Period Plasticity in Visual Cortex. Sci. Rep. 2017, 7, 12646. [Google Scholar] [CrossRef]
- Dityatev, A.; Rusakov, D.A. Molecular signals of plasticity at the tetrapartite synapse. Curr. Opin. Neurobiol. 2011, 21, 353–359. [Google Scholar] [CrossRef] [PubMed]
- Carulli, D.; Broersen, R.; De Winter, F.; Muir, E.M.; Mešković, M.; De Waal, M.; De Vries, S.; Boele, H.-J.; Canto, C.B.; De Zeeuw, C.I.; et al. Cerebellar plasticity and associative memories are controlled by perineuronal nets. Proc. Natl. Acad. Sci. USA 2020, 117, 6855–6865. [Google Scholar] [CrossRef] [PubMed]
- Frischknecht, R.; Heine, M.; Perrais, D.; Seidenbecher, C.I.; Choquet, D.; Gundelfinger, E.D. Brain extracellular matrix affects AMPA receptor lateral mobility and short-term synaptic plasticity. Nat. Neurosci. 2009, 12, 897–904. [Google Scholar] [CrossRef] [PubMed]
- Tewari, B.P.; Chaunsali, L.; Campbell, S.L.; Patel, D.C.; Goode, A.E.; Sontheimer, H. Perineuronal nets decrease membrane capacitance of peritumoral fast spiking interneurons in a model of epilepsy. Nat. Commun. 2018, 9, 4724. [Google Scholar] [CrossRef]
- Tewari, B.; Woo, A.; Prim, C.; Chaunsali, L.; Kimbrough, I.; Engel, K.; Browning, J.; Campbell, S.; Sontheimer, H. Perineuronal Nets Support Astrocytic Ion and Glutamate Homeostasis at Tripartite Synapses. Res. Sq. 2023. preprint. [Google Scholar] [CrossRef]
- Dityatev, A.; Wehrle-Haller, B.; Pitkänen, A. Preface. In Progress in Brain Research; Elsevier: Amsterdam, The Netherlands, 2014; Volume 214, pp. xiii–xvii. [Google Scholar] [CrossRef]
- Fawcett, J.W.; Fyhn, M.; Jendelova, P.; Kwok, J.C.F.; Ruzicka, J.; Sorg, B.A. The extracellular matrix and perineuronal nets in memory. Mol. Psychiatry 2022, 27, 3192–3203. [Google Scholar] [CrossRef]
- Mueller-Buehl, C.; Wegrzyn, D.; Bauch, J.; Faissner, A. Regulation of the E/I-balance by the neural matrisome. Front. Mol. Neurosci. 2023, 16, 1102334. [Google Scholar] [CrossRef] [PubMed]
- Slaker, M.; Barnes, J.; Sorg, B.A.; Grimm, J.W. Impact of Environmental Enrichment on Perineuronal Nets in the Prefrontal Cortex following Early and Late Abstinence from Sucrose Self-Administration in Rats. PLoS ONE 2016, 11, e0168256. [Google Scholar] [CrossRef]
- Lipachev, N.; Arnst, N.; Melnikova, A.; Jäälinoja, H.; Kochneva, A.; Zhigalov, A.; Kulesskaya, N.; Aganov, A.V.; Mavlikeev, M.; Rauvala, H.; et al. Quantitative changes in perineuronal nets in development and posttraumatic condition. J. Mol. Histol. 2019, 50, 203–216. [Google Scholar] [CrossRef] [PubMed]
- Ciampi, L.; Carrara, F.; Totaro, V.; Mazziotti, R.; Lupori, L.; Santiago, C.; Amato, G.; Pizzorusso, T.; Gennaro, C. Learning to count biological structures with raters’ uncertainty. Med. Image Anal. 2022, 80, 102500. [Google Scholar] [CrossRef]
- Slaker, M.L.; Harkness, J.H.; Sorg, B.A. A standardized and automated method of perineuronal net analysis using Wisteria floribunda agglutinin staining intensity. IBRO Rep. 2016, 1, 54–60. [Google Scholar] [CrossRef]
- Lupori, L.; Totaro, V.; Cornuti, S.; Ciampi, L.; Carrara, F.; Grilli, E.; Viglione, A.; Tozzi, F.; Putignano, E.; Mazziotti, R.; et al. A comprehensive atlas of perineuronal net distribution and colocalization with parvalbumin in the adult mouse brain. Cell Rep. 2023, 42, 112788. [Google Scholar] [CrossRef] [PubMed]
- Tippani, M.; Divecha, H.R.; Catallini, J.L.; Kwon, S.H.; Weber, L.M.; Spangler, A.; Jaffe, A.E.; Hyde, T.M.; Kleinman, J.E.; Hicks, S.C.; et al. VistoSeg: Processing utilities for high-resolution images for spatially resolved transcriptomics data. Biol. Imaging 2023, 3, e23. [Google Scholar] [CrossRef]
- Brückner, G.; Szeöke, S.; Pavlica, S.; Grosche, J.; Kacza, J. Axon initial segment ensheathed by extracellular matrix in perineuronal nets. Neuroscience 2006, 138, 365–375. [Google Scholar] [CrossRef] [PubMed]
- Dzyubenko, E.; Manrique-Castano, D.; Kleinschnitz, C.; Faissner, A.; Hermann, D.M. Topological remodeling of cortical perineuronal nets in focal cerebral ischemia and mild hypoperfusion. Matrix Biol. 2018, 74, 121–132. [Google Scholar] [CrossRef] [PubMed]
- Dzyubenko, E.; Willig, K.I.; Yin, D.; Sardari, M.; Tokmak, E.; Labus, P.; Schmermund, B.; Hermann, D.M. Structural changes in perineuronal nets and their perforating GABAergic synapses precede motor coordination recovery post stroke. J. Biomed. Sci. 2023, 30, 76. [Google Scholar] [CrossRef]
- Sigal, Y.M.; Bae, H.; Bogart, L.J.; Hensch, T.K.; Zhuang, X. Structural maturation of cortical perineuronal nets and their perforating synapses revealed by superresolution imaging. Proc. Natl. Acad. Sci. USA 2019, 116, 7071–7076. [Google Scholar] [CrossRef]
- Kaushik, R.; Lipachev, N.; Matuszko, G.; Kochneva, A.; Dvoeglazova, A.; Becker, A.; Paveliev, M.; Dityatev, A. Fine structure analysis of perineuronal nets in the ketamine model of schizophrenia. Eur. J. Neurosci. 2021, 53, 3988–4004. [Google Scholar] [CrossRef] [PubMed]
- Lev-Ram, V.; Lemieux, S.P.; Deerinck, T.J.; Bushong, E.A.; Toyama, B.H.; Perez, A.; Pritchard, D.R.; Park, S.K.R.; McClatchy, D.B.; Savas, J.N.; et al. Do perineuronal nets stabilize the engram of a Synaptic Circuit? Neuroscience 2023. preprint. [Google Scholar] [CrossRef]
- Tansley, S.; Gu, N.; Guzmán, A.U.; Cai, W.; Wong, C.; Lister, K.C.; Muñoz-Pino, E.; Yousefpour, N.; Roome, R.B.; Heal, J.; et al. Microglia-mediated degradation of perineuronal nets promotes pain. Science 2022, 377, 80–86. [Google Scholar] [CrossRef] [PubMed]
- Vo, T.; Carulli, D.; Ehlert, E.M.E.; Kwok, J.C.F.; Dick, G.; Mecollari, V.; Moloney, E.B.; Neufeld, G.; De Winter, F.; Fawcett, J.W.; et al. The chemorepulsive axon guidance protein semaphorin3A is a constituent of perineuronal nets in the adult rodent brain. Mol. Cell. Neurosci. 2013, 56, 186–200. [Google Scholar] [CrossRef]
- Carulli, D.; Rhodes, K.E.; Fawcett, J.W. Upregulation of aggrecan, link protein 1, and hyaluronan synthases during formation of perineuronal nets in the rat cerebellum. J. Comp. Neurol. 2007, 501, 83–94. [Google Scholar] [CrossRef] [PubMed]
- Galtrey, C.M.; Kwok, J.C.F.; Carulli, D.; Rhodes, K.E.; Fawcett, J.W. Distribution and synthesis of extracellular matrix proteoglycans, hyaluronan, link proteins and tenascin-R in the rat spinal cord. Eur. J. Neurosci. 2008, 27, 1373–1390. [Google Scholar] [CrossRef] [PubMed]
- Pantazopoulos, H.; Lange, N.; Hassinger, L.; Berretta, S. Subpopulations of neurons expressing parvalbumin in the human amygdala. J. Comp. Neurol. 2006, 496, 706–722. [Google Scholar] [CrossRef]
- Pantazopoulos, H.; Murray, E.A.; Berretta, S. Total number, distribution, and phenotype of cells expressing chondroitin sulfate proteoglycans in the normal human amygdala. Brain Res. 2008, 1207, 84–95. [Google Scholar] [CrossRef] [PubMed][Green Version]
- Weinrich, L.; Sonntag, M.; Arendt, T.; Morawski, M. Neuroanatomical characterization of perineuronal net components in the human cochlear nucleus and superior olivary complex. Hear. Res. 2018, 367, 32–47. [Google Scholar] [CrossRef]
- Jäger, C.; Lendvai, D.; Seeger, G.; Brückner, G.; Matthews, R.T.; Arendt, T.; Alpár, A.; Morawski, M. Perineuronal and perisynaptic extracellular matrix in the human spinal cord. Neuroscience 2013, 238, 168–184. [Google Scholar] [CrossRef] [PubMed]
- Rahmani, R.; Rambarack, N.; Singh, J.; Constanti, A.; Ali, A.B. Age-Dependent Sex Differences in Perineuronal Nets in an APP Mouse Model of Alzheimer’s Disease Are Brain Region-Specific. Int. J. Mol. Sci. 2023, 24, 14917. [Google Scholar] [CrossRef]
- Pantazopoulos, H.; Woo, T.-U.W.; Lim, M.P.; Lange, N.; Berretta, S. Extracellular Matrix-Glial Abnormalities in the Amygdala and Entorhinal Cortex of Subjects Diagnosed with Schizophrenia. Arch. Gen. Psychiatry 2010, 67, 155. [Google Scholar] [CrossRef] [PubMed]
- Pantazopoulos, H.; Markota, M.; Jaquet, F.; Ghosh, D.; Wallin, A.; Santos, A.; Caterson, B.; Berretta, S. Aggrecan and chondroitin-6-sulfate abnormalities in schizophrenia and bipolar disorder: A postmortem study on the amygdala. Transl. Psychiatry 2015, 5, e496. [Google Scholar] [CrossRef] [PubMed]
- Kilonzo, V.W.; Sweet, R.A.; Glausier, J.R.; Pitts, M.W. Deficits in Glutamic Acid Decarboxylase 67 Immunoreactivity, Parvalbumin Interneurons, and Perineuronal Nets in the Inferior Colliculus of Subjects with Schizophrenia. Schizophr. Bull. 2020, 46, 1053–1059. [Google Scholar] [CrossRef] [PubMed]
- Mauney, S.A.; Athanas, K.M.; Pantazopoulos, H.; Shaskan, N.; Passeri, E.; Berretta, S.; Woo, T.-U.W. Developmental Pattern of Perineuronal Nets in the Human Prefrontal Cortex and Their Deficit in Schizophrenia. Biol. Psychiatry 2013, 74, 427–435. [Google Scholar] [CrossRef] [PubMed]
- Romberg, C.; Yang, S.; Melani, R.; Andrews, M.R.; Horner, A.E.; Spillantini, M.G.; Bussey, T.J.; Fawcett, J.W.; Pizzorusso, T.; Saksida, L.M. Depletion of Perineuronal Nets Enhances Recognition Memory and Long-Term Depression in the Perirhinal Cortex. J. Neurosci. 2013, 33, 7057–7065. [Google Scholar] [CrossRef] [PubMed]
- Schmidt, S.; Stapf, C.; Schmutzler, S.; Lachmann, I.; Arendt, T.; Holzer, M.; Sonntag, M.; Morawski, M. Aggrecan modulates the expression and phosphorylation of tau in a novel bigenic TauP301L—Acan mouse model. Eur. J. Neurosci. 2021, 53, 3889–3904. [Google Scholar] [CrossRef] [PubMed]
- Yang, S.; Cacquevel, M.; Saksida, L.M.; Bussey, T.J.; Schneider, B.L.; Aebischer, P.; Melani, R.; Pizzorusso, T.; Fawcett, J.W.; Spillantini, M.G. Perineuronal net digestion with chondroitinase restores memory in mice with tau pathology. Exp. Neurol. 2015, 265, 48–58. [Google Scholar] [CrossRef] [PubMed]
- Yang, S.; Hilton, S.; Alves, J.N.; Saksida, L.M.; Bussey, T.; Matthews, R.T.; Kitagawa, H.; Spillantini, M.G.; Kwok, J.C.F.; Fawcett, J.W. Antibody recognizing 4-sulfated chondroitin sulfate proteoglycans restores memory in tauopathy-induced neurodegeneration. Neurobiol. Aging 2017, 59, 197–209. [Google Scholar] [CrossRef] [PubMed]
- Pantazopoulos, H.; Gisabella, B.; Rexrode, L.; Benefield, D.; Yildiz, E.; Seltzer, P.; Valeri, J.; Chelini, G.; Reich, A.; Ardelt, M.; et al. Circadian Rhythms of Perineuronal Net Composition. eNeuro 2020, 7, 1–21. [Google Scholar] [CrossRef]
- Valeri, J.; Stiplosek, C.; O’Donovan, S.M.; Sinclair, D.; Grant, K.; Bollavarapu, R.; Platt, D.M.; Stockmeier, C.A.; Gisabella, B.; Pantazopoulos, H. Extracellular Matrix Abnormalities in the Hippocampus of Subjects with Substance Use Disorder. Addict. Med. 2023. preprint. [Google Scholar] [CrossRef]
- Gáti, G.; Morawski, M.; Lendvai, D.; Matthews, R.T.; Jäger, C.; Zachar, G.; Arendt, T.; Alpár, A. Chondroitin sulphate proteoglycan-based perineuronal net establishment is largely activity-independent in chick visual system. J. Chem. Neuroanat. 2010, 40, 243–247. [Google Scholar] [CrossRef] [PubMed]
- Rosenzweig, E.S.; Salegio, E.A.; Liang, J.J.; Weber, J.L.; Weinholtz, C.A.; Brock, J.H.; Moseanko, R.; Hawbecker, S.; Pender, R.; Cruzen, C.L.; et al. Chondroitinase improves anatomical and functional outcomes after primate spinal cord injury. Nat. Neurosci. 2019, 22, 1269–1275. [Google Scholar] [CrossRef]
- Carulli, D.; Rhodes, K.E.; Brown, D.J.; Bonnert, T.P.; Pollack, S.J.; Oliver, K.; Strata, P.; Fawcett, J.W. Composition of perineuronal nets in the adult rat cerebellum and the cellular origin of their components. J. Comp. Neurol. 2006, 494, 559–577. [Google Scholar] [CrossRef] [PubMed]
- Kwok, J.C.F.; Carulli, D.; Fawcett, J.W. In vitro modeling of perineuronal nets: Hyaluronan synthase and link protein are necessary for their formation and integrity. J. Neurochem. 2010, 114, 1447–1459. [Google Scholar] [CrossRef] [PubMed]
- Dick, G.; Tan, C.L.; Alves, J.N.; Ehlert, E.M.E.; Miller, G.M.; Hsieh-Wilson, L.C.; Sugahara, K.; Oosterhof, A.; Van Kuppevelt, T.H.; Verhaagen, J.; et al. Semaphorin 3A Binds to the Perineuronal Nets via Chondroitin Sulfate Type E Motifs in Rodent Brains. J. Biol. Chem. 2013, 288, 27384–27395. [Google Scholar] [CrossRef] [PubMed]
- Wang, D.; Ichiyama, R.M.; Zhao, R.; Andrews, M.R.; Fawcett, J.W. Chondroitinase Combined with Rehabilitation Promotes Recovery of Forelimb Function in Rats with Chronic Spinal Cord Injury. J. Neurosci. 2011, 31, 9332–9344. [Google Scholar] [CrossRef]
- Štepánková, K.; Chudíčková, M.; Šimková, Z.; Martinez-Varea, N.; Kubinová, Š.; Urdzíková, L.M.; Jendelová, P.; Kwok, J.C.F. Low oral dose of 4-methylumbelliferone reduces glial scar but is insufficient to induce functional recovery after spinal cord injury. Sci. Rep. 2023, 13, 19183. [Google Scholar] [CrossRef]
- Lépine, M.; Douceau, S.; Devienne, G.; Prunotto, P.; Lenoir, S.; Regnauld, C.; Pouettre, E.; Piquet, J.; Lebouvier, L.; Hommet, Y.; et al. Parvalbumin interneuron-derived tissue-type plasminogen activator shapes perineuronal net structure. BMC Biol. 2022, 20, 218. [Google Scholar] [CrossRef] [PubMed]
- Perna, J.; Lu, J.; Mullen, B.; Liu, T.; Tjia, M.; Weiser, S.; Ackman, J.; Zuo, Y. Perinatal Penicillin Exposure Affects Cortical Development and Sensory Processing. Front. Mol. Neurosci. 2021, 14, 704219. [Google Scholar] [CrossRef] [PubMed]
- Roura-Martínez, D.; Díaz-Bejarano, P.; Ucha, M.; Paiva, R.R.; Ambrosio, E.; Higuera-Matas, A. Comparative analysis of the modulation of perineuronal nets in the prefrontal cortex of rats during protracted withdrawal from cocaine, heroin and sucrose self-administration. Neuropharmacology 2020, 180, 108290. [Google Scholar] [CrossRef]
- Xue, Y.-X.; Xue, L.-F.; Liu, J.-F.; He, J.; Deng, J.-H.; Sun, S.-C.; Han, H.-B.; Luo, Y.-X.; Xu, L.-Z.; Wu, P.; et al. Depletion of Perineuronal Nets in the Amygdala to Enhance the Erasure of Drug Memories. J. Neurosci. 2014, 34, 6647–6658. [Google Scholar] [CrossRef] [PubMed]
- Marchand, A.; Schwartz, C. Perineuronal net expression in the brain of a hibernating mammal. Brain Struct. Funct. 2020, 225, 45–56. [Google Scholar] [CrossRef]
- Ueno, H.; Suemitsu, S.; Murakami, S.; Kitamura, N.; Wani, K.; Takahashi, Y.; Matsumoto, Y.; Okamoto, M.; Ishihara, T. Alteration of Extracellular Matrix Molecules and Perineuronal Nets in the Hippocampus of Pentylenetetrazol-Kindled Mice. Neural Plast. 2019, 2019, 8924634. [Google Scholar] [CrossRef] [PubMed]
- Enwright, J.F.; Sanapala, S.; Foglio, A.; Berry, R.; Fish, K.N.; Lewis, D.A. Reduced Labeling of Parvalbumin Neurons and Perineuronal Nets in the Dorsolateral Prefrontal Cortex of Subjects with Schizophrenia. Neuropsychopharmacology 2016, 41, 2206–2214. [Google Scholar] [CrossRef] [PubMed]
- Steullet, P.; Cabungcal, J.-H.; Bukhari, S.A.; Ardelt, M.I.; Pantazopoulos, H.; Hamati, F.; Salt, T.E.; Cuenod, M.; Do, K.Q.; Berretta, S. The thalamic reticular nucleus in schizophrenia and bipolar disorder: Role of parvalbumin-expressing neuron networks and oxidative stress. Mol. Psychiatry 2018, 23, 2057–2065. [Google Scholar] [CrossRef] [PubMed]
- Rey, C.C.; Robert, V.; Bouisset, G.; Loisy, M.; Lopez, S.; Cattaud, V.; Lejards, C.; Piskorowski, R.A.; Rampon, C.; Chevaleyre, V.; et al. Altered inhibitory function in hippocampal CA2 contributes in social memory deficits in Alzheimer’s mouse model. iScience 2022, 25, 103895. [Google Scholar] [CrossRef]
- Boggio, E.M.; Ehlert, E.M.; Lupori, L.; Moloney, E.B.; De Winter, F.; Vander Kooi, C.W.; Baroncelli, L.; Mecollari, V.; Blits, B.; Fawcett, J.W.; et al. Inhibition of Semaphorin3A Promotes Ocular Dominance Plasticity in the Adult Rat Visual Cortex. Mol. Neurobiol. 2019, 56, 5987–5997. [Google Scholar] [CrossRef]
- Dubisova, J.; Burianova, J.S.; Svobodova, L.; Makovicky, P.; Martinez-Varea, N.; Cimpean, A.; Fawcett, J.W.; Kwok, J.C.F.; Kubinova, S. Oral treatment of 4-methylumbelliferone reduced perineuronal nets and improved recognition memory in mice. Brain Res. Bull. 2022, 181, 144–156. [Google Scholar] [CrossRef]
- Blosa, M.; Bursch, C.; Weigel, S.; Holzer, M.; Jäger, C.; Janke, C.; Matthews, R.T.; Arendt, T.; Morawski, M. Reorganization of Synaptic Connections and Perineuronal Nets in the Deep Cerebellar Nuclei of Purkinje Cell Degeneration Mutant Mice. Neural Plast. 2016, 2016, 2828536. [Google Scholar] [CrossRef] [PubMed]
- Shah, A.; Lodge, D.J. A loss of hippocampal perineuronal nets produces deficits in dopamine system function: Relevance to the positive symptoms of schizophrenia. Transl. Psychiatry 2013, 3, e215. [Google Scholar] [CrossRef]
- Napoli, D.; Lupori, L.; Mazziotti, R.; Sagona, G.; Bagnoli, S.; Samad, M.; Sacramento, E.K.; Kirkpartick, J.; Putignano, E.; Chen, S.; et al. MiR-29 coordinates age-dependent plasticity brakes in the adult visual cortex. EMBO Rep. 2020, 21, e50431. [Google Scholar] [CrossRef]
- Lensjø, K.K.; Lepperød, M.E.; Dick, G.; Hafting, T.; Fyhn, M. Removal of Perineuronal Nets Unlocks Juvenile Plasticity Through Network Mechanisms of Decreased Inhibition and Increased Gamma Activity. J. Neurosci. 2017, 37, 1269–1283. [Google Scholar] [CrossRef]
- Erchova, I.; Vasalauskaite, A.; Longo, V.; Sengpiel, F. Enhancement of visual cortex plasticity by dark exposure. Philos. Trans. R. Soc. B Biol. Sci. 2017, 372, 20160159. [Google Scholar] [CrossRef] [PubMed]
- Irvine, S.; Kwok, J. Perineuronal Nets in Spinal Motoneurones: Chondroitin Sulphate Proteoglycan around Alpha Motoneurones. Int. J. Mol. Sci. 2018, 19, 1172. [Google Scholar] [CrossRef]
- Dauth, S.; Grevesse, T.; Pantazopoulos, H.; Campbell, P.H.; Maoz, B.M.; Berretta, S.; Parker, K.K. Extracellular matrix protein expression is brain region dependent. J. Comp. Neurol. 2016, 524, 1309–1336. [Google Scholar] [CrossRef] [PubMed]
- Baidoe-Ansah, D.; Sakib, S.; Jia, S.; Mirzapourdelavar, H.; Strackeljan, L.; Fischer, A.; Aleshin, S.; Kaushik, R.; Dityatev, A. Aging-Associated Changes in Cognition, Expression and Epigenetic Regulation of Chondroitin 6-Sulfotransferase Chst3. Cells 2022, 11, 2033. [Google Scholar] [CrossRef]
- Benbenishty, A.; Peled-Hajaj, S.; Krishnaswamy, V.R.; Har-Gil, H.; Havusha-Laufer, S.; Ruggiero, A.; Slutsky, I.; Blinder, P.; Sagi, I. Longitudinal in vivo imaging of perineuronal nets. Neurophotonics 2023, 10, 015008. [Google Scholar] [CrossRef] [PubMed]
- Lemieux, S.P.; Lev-Ram, V.; Tsien, R.Y.; Ellisman, M.H. Perineuronal nets and the neuronal extracellular matrix can be imaged by genetically encoded labeling of HAPLN1 in vitro and in vivo. Neuroscience 2023. preprint. [Google Scholar] [CrossRef]
- Carstens, K.E.; Phillips, M.L.; Pozzo-Miller, L.; Weinberg, R.J.; Dudek, S.M. Perineuronal Nets Suppress Plasticity of Excitatory Synapses on CA2 Pyramidal Neurons. J. Neurosci. 2016, 36, 6312–6320. [Google Scholar] [CrossRef] [PubMed]
- O’Connor, A.M.; Burton, T.J.; Mansuri, H.; Hand, G.R.; Leamey, C.A.; Sawatari, A. Environmental Enrichment From Birth Impacts Parvalbumin Expressing Cells and Wisteria Floribunda Agglutinin Labelled Peri-Neuronal Nets within the Developing Murine Striatum. Front. Neuroanat. 2019, 13, 90. [Google Scholar] [CrossRef] [PubMed]
- Carulli, D.; Foscarin, S.; Faralli, A.; Pajaj, E.; Rossi, F. Modulation of semaphorin3A in perineuronal nets during structural plasticity in the adult cerebellum. Mol. Cell. Neurosci. 2013, 57, 10–22. [Google Scholar] [CrossRef]
- Wegrzyn, D.; Freund, N.; Faissner, A.; Juckel, G. Poly I:C Activated Microglia Disrupt Perineuronal Nets and Modulate Synaptic Balance in Primary Hippocampal Neurons in vitro. Front. Synaptic Neurosci. 2021, 13, 637549. [Google Scholar] [CrossRef]
- Van’t Spijker, H.M.; Rowlands, D.; Rossier, J.; Haenzi, B.; Fawcett, J.W.; Kwok, J.C.F. Neuronal Pentraxin 2 Binds PNNs and Enhances PNN Formation. Neural Plast. 2019, 2019, 6804575. [Google Scholar] [CrossRef]
- Crapser, J.D.; Ochaba, J.; Soni, N.; Reidling, J.C.; Thompson, L.M.; Green, K.N. Microglial depletion prevents extracellular matrix changes and striatal volume reduction in a model of Huntington’s disease. Brain 2020, 143, 266–288. [Google Scholar] [CrossRef]
- Stoyanov, S.; Sun, W.; Düsedau, H.P.; Cangalaya, C.; Choi, I.; Mirzapourdelavar, H.; Baidoe-Ansah, D.; Kaushik, R.; Neumann, J.; Dunay, I.R.; et al. Attenuation of the extracellular matrix restores microglial activity during the early stage of amyloidosis. Glia 2021, 69, 182–200. [Google Scholar] [CrossRef]
- Lendvai, D.; Morawski, M.; Négyessy, L.; Gáti, G.; Jäger, C.; Baksa, G.; Glasz, T.; Attems, J.; Tanila, H.; Arendt, T.; et al. Neurochemical mapping of the human hippocampus reveals perisynaptic matrix around functional synapses in Alzheimer’s disease. Acta Neuropathol. 2013, 125, 215–229. [Google Scholar] [CrossRef]
- Alcaide, J.; Guirado, R.; Crespo, C.; Blasco-Ibáñez, J.M.; Varea, E.; Sanjuan, J.; Nacher, J. Alterations of perineuronal nets in the dorsolateral prefrontal cortex of neuropsychiatric patients. Int. J. Bipolar Disord. 2019, 7, 24. [Google Scholar] [CrossRef]
- Matuszko, G.; Curreli, S.; Kaushik, R.; Becker, A.; Dityatev, A. Extracellular matrix alterations in the ketamine model of schizophrenia. Neuroscience 2017, 350, 13–22. [Google Scholar] [CrossRef]
- Allgäuer, L.; Cabungcal, J.-H.; Yzydorczyk, C.; Do, K.Q.; Dwir, D. Low protein-induced intrauterine growth restriction as a risk factor for schizophrenia phenotype in a rat model: Assessing the role of oxidative stress and neuroinflammation interaction. Transl. Psychiatry 2023, 13, 30. [Google Scholar] [CrossRef] [PubMed]
- Slaker, M.L.; Jorgensen, E.T.; Hegarty, D.M.; Liu, X.; Kong, Y.; Zhang, F.; Linhardt, R.J.; Brown, T.E.; Aicher, S.A.; Sorg, B.A. Cocaine Exposure Modulates Perineuronal Nets and Synaptic Excitability of Fast-Spiking Interneurons in the Medial Prefrontal Cortex. eNeuro 2018, 5, 30294670. [Google Scholar] [CrossRef]
- Wingert, J.C.; Anguiano, J.N.; Ramos, J.D.; Blacktop, J.M.; Gonzalez, A.E.; Churchill, L.; Sorg, B.A. Enhanced expression of parvalbumin and perineuronal nets in the medial prefrontal cortex after extended-access cocaine self-administration in rats. Addict. Biol. 2023, 28, e13334. [Google Scholar] [CrossRef]
- Sanchez-Hernandez, A.; Nicolas, C.; Gil-Miravet, I.; Guarque-Chabrera, J.; Solinas, M.; Miquel, M. Time-dependent regulation of perineuronal nets in the cerebellar cortex during abstinence of cocaine-self administration. Psychopharmacology 2021, 238, 1059–1068. [Google Scholar] [CrossRef] [PubMed]
- Jorgensen, E.T.; Gonzalez, A.E.; Harkness, J.H.; Hegarty, D.M.; Thakar, A.; Burchi, D.J.; Aadland, J.A.; Aicher, S.A.; Sorg, B.A.; Brown, T.E. Cocaine memory reactivation induces functional adaptations within parvalbumin interneurons in the rat medial prefrontal cortex. Addict. Biol. 2021, 26, e12947. [Google Scholar] [CrossRef] [PubMed]
- Carbo-Gas, M.; Moreno-Rius, J.; Guarque-Chabrera, J.; Vazquez-Sanroman, D.; Gil-Miravet, I.; Carulli, D.; Hoebeek, F.; De Zeeuw, C.; Sanchis-Segura, C.; Miquel, M. Cerebellar perineuronal nets in cocaine-induced pavlovian memory: Site matters. Neuropharmacology 2017, 125, 166–180. [Google Scholar] [CrossRef] [PubMed]
- Vazquez-Sanroman, D.; Leto, K.; Cerezo-Garcia, M.; Carbo-Gas, M.; Sanchis-Segura, C.; Carulli, D.; Rossi, F.; Miquel, M. The cerebellum on cocaine: Plasticity and metaplasticity. Addict. Biol. 2015, 20, 941–955. [Google Scholar] [CrossRef]
- Forostyak, S.; Forostyak, O.; Kwok, J.C.F.; Romanyuk, N.; Rehorova, M.; Kriska, J.; Dayanithi, G.; Raha-Chowdhury, R.; Jendelova, P.; Anderova, M.; et al. Transplantation of Neural Precursors Derived from Induced Pluripotent Cells Preserve Perineuronal Nets and Stimulate Neural Plasticity in ALS Rats. Int. J. Mol. Sci. 2020, 21, 9593. [Google Scholar] [CrossRef]
- Yang, T.; Hu, S.; Chang, W.-C.; Kao, H.-Y.; Wang, Y. Perineuronal Nets Degradation and Parvalbumin Interneuron Loss in a Mouse Model of DEPDC5-Related Epilepsy. Dev. Neurosci. 2022, 44, 671–677. [Google Scholar] [CrossRef] [PubMed]
- Giamanco, K.A.; Morawski, M.; Matthews, R.T. Perineuronal net formation and structure in aggrecan knockout mice. Neuroscience 2010, 170, 1314–1327. [Google Scholar] [CrossRef]
- Bertocchi, I.; Mele, P.; Ferrero, G.; Oberto, A.; Carulli, D.; Eva, C. NPY-Y1 receptor signaling controls spatial learning and perineuronal net expression. Neuropharmacology 2021, 184, 108425. [Google Scholar] [CrossRef]
- Rowlands, D.; Lensjø, K.K.; Dinh, T.; Yang, S.; Andrews, M.R.; Hafting, T.; Fyhn, M.; Fawcett, J.W.; Dick, G. Aggrecan Directs Extracellular Matrix-Mediated Neuronal Plasticity. J. Neurosci. 2018, 38, 10102–10113. [Google Scholar] [CrossRef] [PubMed]
- Ruzicka, J.; Dalecka, M.; Safrankova, K.; Peretti, D.; Jendelova, P.; Kwok, J.C.F.; Fawcett, J.W. Perineuronal nets affect memory and learning after synapse withdrawal. Transl. Psychiatry 2022, 12, 480. [Google Scholar] [CrossRef] [PubMed]
- Sonntag, M.; Blosa, M.; Schmidt, S.; Reimann, K.; Blum, K.; Eckrich, T.; Seeger, G.; Hecker, D.; Schick, B.; Arendt, T.; et al. Synaptic coupling of inner ear sensory cells is controlled by brevican-based extracellular matrix baskets resembling perineuronal nets. BMC Biol. 2018, 16, 99. [Google Scholar] [CrossRef] [PubMed]
- Cannarozzo, C.; Rubiolo, A.; Casarotto, P.; Castrén, E. Ketamine and its metabolite 2R,6R-hydroxynorketamine promote ocular dominance plasticity and release tropomyosin-related kinase B from inhibitory control without reducing perineuronal nets enwrapping parvalbumin interneurons. Eur. J. Neurosci. 2023, 57, 940–950. [Google Scholar] [CrossRef] [PubMed]
- Gogolla, N.; Caroni, P.; Lüthi, A.; Herry, C. Perineuronal Nets Protect Fear Memories from Erasure. Science 2009, 325, 1258–1261. [Google Scholar] [CrossRef]
- Shi, W.; Wei, X.; Wang, X.; Du, S.; Liu, W.; Song, J.; Wang, Y. Perineuronal nets protect long-term memory by limiting activity-dependent inhibition from parvalbumin interneurons. Proc. Natl. Acad. Sci. USA 2019, 116, 27063–27073. [Google Scholar] [CrossRef] [PubMed]
- Poli, A.; Viglione, A.; Mazziotti, R.; Totaro, V.; Morea, S.; Melani, R.; Silingardi, D.; Putignano, E.; Berardi, N.; Pizzorusso, T. Selective Disruption of Perineuronal Nets in Mice Lacking Crtl1 is Sufficient to Make Fear Memories Susceptible to Erasure. Mol. Neurobiol. 2023, 60, 4105–4119. [Google Scholar] [CrossRef] [PubMed]
- Cabungcal, J.-H.; Steullet, P.; Morishita, H.; Kraftsik, R.; Cuenod, M.; Hensch, T.K.; Do, K.Q. Perineuronal nets protect fast-spiking interneurons against oxidative stress. Proc. Natl. Acad. Sci. USA 2013, 110, 9130–9135. [Google Scholar] [CrossRef] [PubMed]
- Morawski, M.; Brückner, M.K.; Riederer, P.; Brückner, G.; Arendt, T. Perineuronal nets potentially protect against oxidative stress. Exp. Neurol. 2004, 188, 309–315. [Google Scholar] [CrossRef]
- Guirado, R.; Perez-Rando, M.; Sanchez-Matarredona, D.; Castrén, E.; Nacher, J. Chronic fluoxetine treatment alters the structure, connectivity and plasticity of cortical interneurons. Int. J. Neuropsychopharmacol. 2014, 17, 1635–1646. [Google Scholar] [CrossRef]
- Umemori, J.; Winkel, F.; Castrén, E.; Karpova, N.N. Distinct effects of perinatal exposure to fluoxetine or methylmercury on parvalbumin and perineuronal nets, the markers of critical periods in brain development. Int. J. Dev. Neurosci. 2015, 44, 55–64. [Google Scholar] [CrossRef]
- Jetsonen, E.; Didio, G.; Winkel, F.; Llach Pou, M.; Boj, C.; Kuczynski-Noyau, L.; Võikar, V.; Guirado, R.; Taira, T.; Lauri, S.E.; et al. Activation of TrkB in Parvalbumin interneurons is required for the promotion of reversal learning in spatial and fear memory by antidepressants. Neuropsychopharmacology 2023, 48, 1021–1030. [Google Scholar] [CrossRef] [PubMed]
- Murthy, S.; Kane, G.A.; Katchur, N.J.; Lara Mejia, P.S.; Obiofuma, G.; Buschman, T.J.; McEwen, B.S.; Gould, E. Perineuronal Nets, Inhibitory Interneurons, and Anxiety-Related Ventral Hippocampal Neuronal Oscillations Are Altered by Early Life Adversity. Biol. Psychiatry 2019, 85, 1011–1020. [Google Scholar] [CrossRef] [PubMed]
- Dityatev, A.; Brückner, G.; Dityateva, G.; Grosche, J.; Kleene, R.; Schachner, M. Activity-dependent formation and functions of chondroitin sulfate-rich extracellular matrix of perineuronal nets. Dev. Neurobiol. 2007, 67, 570–588. [Google Scholar] [CrossRef] [PubMed]
- Strackeljan, L.; Baczynska, E.; Cangalaya, C.; Baidoe-Ansah, D.; Wlodarczyk, J.; Kaushik, R.; Dityatev, A. Microglia Depletion-Induced Remodeling of Extracellular Matrix and Excitatory Synapses in the Hippocampus of Adult Mice. Cells 2021, 10, 1862. [Google Scholar] [CrossRef] [PubMed]
- Foscarin, S.; Ponchione, D.; Pajaj, E.; Leto, K.; Gawlak, M.; Wilczynski, G.M.; Rossi, F.; Carulli, D. Experience-Dependent Plasticity and Modulation of Growth Regulatory Molecules at Central Synapses. PLoS ONE 2011, 6, e16666. [Google Scholar] [CrossRef] [PubMed]
- Yang, S.; Gigout, S.; Molinaro, A.; Naito-Matsui, Y.; Hilton, S.; Foscarin, S.; Nieuwenhuis, B.; Tan, C.L.; Verhaagen, J.; Pizzorusso, T.; et al. Chondroitin 6-sulphate is required for neuroplasticity and memory in ageing. Mol. Psychiatry 2021, 26, 5658–5668. [Google Scholar] [CrossRef] [PubMed]
- Wegrzyn, D.; Manitz, M.; Kostka, M.; Freund, N.; Juckel, G.; Faissner, A. Poly I:C-induced maternal immune challenge reduces perineuronal net area and raises spontaneous network activity of hippocampal neurons in vitro. Eur. J. Neurosci. 2021, 53, 3920–3941. [Google Scholar] [CrossRef] [PubMed]
- Gottschling, C.; Wegrzyn, D.; Denecke, B.; Faissner, A. Elimination of the four extracellular matrix molecules tenascin-C, tenascin-R, brevican and neurocan alters the ratio of excitatory and inhibitory synapses. Sci. Rep. 2019, 9, 13939. [Google Scholar] [CrossRef]
- Morawski, M.; Dityatev, A.; Hartlage-Rübsamen, M.; Blosa, M.; Holzer, M.; Flach, K.; Pavlica, S.; Dityateva, G.; Grosche, J.; Brückner, G.; et al. Tenascin-R promotes assembly of the extracellular matrix of perineuronal nets via clustering of aggrecan. Philos. Trans. R. Soc. B Biol. Sci. 2014, 369, 20140046. [Google Scholar] [CrossRef]
- Klimczak, P.; Rizzo, A.; Castillo-Gómez, E.; Perez-Rando, M.; Gramuntell, Y.; Beltran, M.; Nacher, J. Parvalbumin Interneurons and Perineuronal Nets in the Hippocampus and Retrosplenial Cortex of Adult Male Mice After Early Social Isolation Stress and Perinatal NMDA Receptor Antagonist Treatment. Front. Synaptic Neurosci. 2021, 13, 733989. [Google Scholar] [CrossRef]
- Cope, E.C.; Zych, A.D.; Katchur, N.J.; Waters, R.C.; Laham, B.J.; Diethorn, E.J.; Park, C.Y.; Meara, W.R.; Gould, E. Atypical perineuronal nets in the CA2 region interfere with social memory in a mouse model of social dysfunction. Mol. Psychiatry 2022, 27, 3520–3531. [Google Scholar] [CrossRef] [PubMed]
- Faralli, A.; Dagna, F.; Albera, A.; Bekku, Y.; Oohashi, T.; Albera, R.; Rossi, F.; Carulli, D. Modifications of perineuronal nets and remodelling of excitatory and inhibitory afferents during vestibular compensation in the adult mouse. Brain Struct. Funct. 2016, 221, 3193–3209. [Google Scholar] [CrossRef]
- Christensen, A.C.; Lensjø, K.K.; Lepperød, M.E.; Dragly, S.-A.; Sutterud, H.; Blackstad, J.S.; Fyhn, M.; Hafting, T. Perineuronal nets stabilize the grid cell network. Nat. Commun. 2021, 12, 253. [Google Scholar] [CrossRef]
- Sun, Z.Y.; Bozzelli, P.L.; Caccavano, A.; Allen, M.; Balmuth, J.; Vicini, S.; Wu, J.; Conant, K. Disruption of perineuronal nets increases the frequency of sharp wave ripple events. Hippocampus 2018, 28, 42–52. [Google Scholar] [CrossRef] [PubMed]
- Orlando, C.; Ster, J.; Gerber, U.; Fawcett, J.W.; Raineteau, O. Perisynaptic Chondroitin Sulfate Proteoglycans Restrict Structural Plasticity in an Integrin-Dependent Manner. J. Neurosci. 2012, 32, 18009–18017. [Google Scholar] [CrossRef]
- Faini, G.; Aguirre, A.; Landi, S.; Lamers, D.; Pizzorusso, T.; Ratto, G.M.; Deleuze, C.; Bacci, A. Perineuronal nets control visual input via thalamic recruitment of cortical PV interneurons. eLife 2018, 7, e41520. [Google Scholar] [CrossRef] [PubMed]
- Al’joboori, Y.D.; Edgerton, V.R.; Ichiyama, R.M. Effects of Rehabilitation on Perineural Nets and Synaptic Plasticity Following Spinal Cord Transection. Brain Sci. 2020, 10, 824. [Google Scholar] [CrossRef] [PubMed]
- Milton, A.J.; Kwok, J.C.F.; McClellan, J.; Randall, S.G.; Lathia, J.D.; Warren, P.M.; Silver, D.J.; Silver, J. Recovery of Forearm and Fine Digit Function After Chronic Spinal Cord Injury by Simultaneous Blockade of Inhibitory Matrix Chondroitin Sulfate Proteoglycan Production and the Receptor PTPσ. J. Neurotrauma 2023, 40, 2500–2521. [Google Scholar] [CrossRef] [PubMed]
- Tsien, R.Y. Very long-term memories may be stored in the pattern of holes in the perineuronal net. Proc. Natl. Acad. Sci. USA 2013, 110, 12456–12461. [Google Scholar] [CrossRef] [PubMed]
- Ramsaran, A.I.; Wang, Y.; Golbabaei, A.; Aleshin, S.; De Snoo, M.L.; Yeung, B.A.; Rashid, A.J.; Awasthi, A.; Lau, J.; Tran, L.M.; et al. A shift in the mechanisms controlling hippocampal engram formation during brain maturation. Science 2023, 380, 543–551. [Google Scholar] [CrossRef] [PubMed]
- Huang, H.; Joffrin, A.M.; Zhao, Y.; Miller, G.M.; Zhang, G.C.; Oka, Y.; Hsieh-Wilson, L.C. Chondroitin 4-O-sulfation regulates hippocampal perineuronal nets and social memory. Proc. Natl. Acad. Sci. USA 2023, 120, e2301312120. [Google Scholar] [CrossRef] [PubMed]
- Burket, J.A.; Webb, J.D.; Deutsch, S.I. Perineuronal Nets and Metal Cation Concentrations in the Microenvironments of Fast-Spiking, Parvalbumin-Expressing GABAergic Interneurons: Relevance to Neurodevelopment and Neurodevelopmental Disorders. Biomolecules 2021, 11, 1235. [Google Scholar] [CrossRef]
- Chang, K.; Lin, L.; Cui, T.; Zhao, H.; Li, J.; Liu, C.; Gao, D.; Lu, S. Zinc-a2-Glycoprotein Acts as a Component of PNN to Protect Hippocampal Neurons from Apoptosis. Mol. Neurobiol. 2023. online ahead of print. [Google Scholar] [CrossRef]
- Wegrzyn, D.; Juckel, G.; Faissner, A. Structural and Functional Deviations of the Hippocampus in Schizophrenia and Schizophrenia Animal Models. Int. J. Mol. Sci. 2022, 23, 5482. [Google Scholar] [CrossRef] [PubMed]
- Miyamae, T.; Chen, K.; Lewis, D.A.; Gonzalez-Burgos, G. Distinct Physiological Maturation of Parvalbumin-Positive Neuron Subtypes in Mouse Prefrontal Cortex. J. Neurosci. 2017, 37, 4883–4902. [Google Scholar] [CrossRef]
- Cho, K.K.A.; Hoch, R.; Lee, A.T.; Patel, T.; Rubenstein, J.L.R.; Sohal, V.S. Gamma Rhythms Link Prefrontal Interneuron Dysfunction with Cognitive Inflexibility in Dlx5/6+/− Mice. Neuron 2015, 85, 1332–1343. [Google Scholar] [CrossRef] [PubMed]
- Maas, D.A.; Eijsink, V.D.; Spoelder, M.; Van Hulten, J.A.; De Weerd, P.; Homberg, J.R.; Vallès, A.; Nait-Oumesmar, B.; Martens, G.J.M. Interneuron hypomyelination is associated with cognitive inflexibility in a rat model of schizophrenia. Nat. Commun. 2020, 11, 2329. [Google Scholar] [CrossRef] [PubMed]
- Patrono, E.; Hrůzova, K.; Svoboda, J.; Stuchlík, A. The role of optogenetic stimulations of parvalbumin-positive interneurons in the prefrontal cortex and the ventral hippocampus on an acute MK-801 model of schizophrenia-like cognitive inflexibility. Schizophr. Res. 2023, 252, 198–205. [Google Scholar] [CrossRef]
- Kaar, S.J.; Angelescu, I.; Marques, T.R.; Howes, O.D. Pre-frontal parvalbumin interneurons in schizophrenia: A meta-analysis of post-mortem studies. J. Neural Transm. 2019, 126, 1637–1651. [Google Scholar] [CrossRef] [PubMed]
- Zhang, W.-J.; Shi, H.-Z.; Guo, M.-N.; Xu, L.-F.; Zhai, H.-R.; Liu, Z.-Z.; Zhu, Y.-Q.; Zhang, W.-N.; Wang, J. PGC-1α regulates critical period onset/closure, mediating cortical plasticity. Front. Mol. Neurosci. 2023, 16, 1149906. [Google Scholar] [CrossRef] [PubMed]
- Rankin-Gee, E.K.; McRae, P.A.; Baranov, E.; Rogers, S.; Wandrey, L.; Porter, B.E. Perineuronal net degradation in epilepsy. Epilepsia 2015, 56, 1124–1133. [Google Scholar] [CrossRef]
- Tewari, B.P.; Chaunsali, L.; Prim, C.E.; Sontheimer, H. A glial perspective on the extracellular matrix and perineuronal net remodeling in the central nervous system. Front. Cell. Neurosci. 2022, 16, 1022754. [Google Scholar] [CrossRef] [PubMed]
- Crapser, J.D.; Spangenberg, E.E.; Barahona, R.A.; Arreola, M.A.; Hohsfield, L.A.; Green, K.N. Microglia facilitate loss of perineuronal nets in the Alzheimer’s disease brain. EBioMedicine 2020, 58, 102919. [Google Scholar] [CrossRef]
- Wingert, J.C.; Ramos, J.D.; Reynolds, S.X.; Gonzalez, A.E.; Rose, R.M.; Hegarty, D.M.; Aicher, S.A.; Bailey, L.G.; Brown, T.E.; Abbas, A.I.; et al. Perineuronal nets in the rat medial prefrontal cortex alter hippocampal-prefrontal oscillations and reshape cocaine self-administration memories. Neuroscience 2024. preprint. [Google Scholar] [CrossRef]
- Sánchez-Ventura, J.; Giménez-Llort, L.; Penas, C.; Udina, E. Voluntary wheel running preserves lumbar perineuronal nets, enhances motor functions and prevents hyperreflexia after spinal cord injury. Exp. Neurol. 2021, 336, 113533. [Google Scholar] [CrossRef] [PubMed]
- Wang, H.; Rivenson, Y.; Jin, Y.; Wei, Z.; Gao, R.; Günaydın, H.; Bentolila, L.A.; Kural, C.; Ozcan, A. Deep learning enables cross-modality super-resolution in fluorescence microscopy. Nat. Methods 2019, 16, 103–110. [Google Scholar] [CrossRef] [PubMed]
- Oláh, S.; Füle, M.; Komlósi, G.; Varga, C.; Báldi, R.; Barzó, P.; Tamás, G. Regulation of cortical microcircuits by unitary GABA-mediated volume transmission. Nature 2009, 461, 1278–1281. [Google Scholar] [CrossRef] [PubMed]
- Karayannis, T.; Elfant, D.; Huerta-Ocampo, I.; Teki, S.; Scott, R.S.; Rusakov, D.A.; Jones, M.V.; Capogna, M. Slow GABA Transient and Receptor Desensitization Shape Synaptic Responses Evoked by Hippocampal Neurogliaform Cells. J. Neurosci. 2010, 30, 9898–9909. [Google Scholar] [CrossRef]
- Spoleti, E.; La Barbera, L.; Cauzzi, E.; De Paolis, M.L.; Saba, L.; Marino, R.; Sciamanna, G.; Di Lazzaro, V.; Keller, F.; Nobili, A.; et al. Dopamine neuron degeneration in the Ventral Tegmental Area causes hippocampal hyperexcitability in experimental Alzheimer’s Disease. Mol. Psychiatry 2024. [Google Scholar] [CrossRef] [PubMed]
- Cardarelli, R.A.; Martin, R.; Jaaro-Peled, H.; Sawa, A.; Powell, E.M.; O’Donnell, P. Dominant-Negative DISC1 Alters the Dopaminergic Modulation of Inhibitory Interneurons in the Mouse Prefrontal Cortex. Mol. Neuropsychiatry 2018, 4, 20–29. [Google Scholar] [CrossRef]
- Takiguchi, M.; Miyashita, K.; Yamazaki, K.; Funakoshi, K. Chondroitinase ABC Administration Facilitates Serotonergic Innervation of Motoneurons in Rats with Complete Spinal Cord Transection. Front. Integr. Neurosci. 2022, 16, 881632. [Google Scholar] [CrossRef]
- Dembitskaya, Y.; Gavrilov, N.; Kraev, I.; Doronin, M.; Tang, Y.; Li, L.; Semyanov, A. Attenuation of the extracellular matrix increases the number of synapses but suppresses synaptic plasticity through upregulation of SK channels. Cell Calcium 2021, 96, 102406. [Google Scholar] [CrossRef] [PubMed]
- Korotchenko, S.; Zanacchi, F.C.; Diaspro, A.; Dityatev, A. Zooming in on the (Peri)synaptic Extracellular Matrix. Neuromethods 2014, 84, 187–203. [Google Scholar] [CrossRef]
- Dzyubenko, E.; Rozenberg, A.; Hermann, D.M.; Faissner, A. Colocalization of synapse marker proteins evaluated by STED-microscopy reveals patterns of neuronal synapse distribution in vitro. J. Neurosci. Methods 2016, 273, 149–159. [Google Scholar] [CrossRef] [PubMed]
- Tewari, B.; Sontheimer, H. Protocol to Quantitatively Assess the Structural Integrity of Perineuronal Nets ex vivo. BIO-Protoc. 2019, 9, e3234. [Google Scholar] [CrossRef] [PubMed]
- Burket, J.A.; Urbano, M.R.; Deutsch, S.I. Sugarcoated Perineuronal Nets Regulate “GABAergic” Transmission: Bittersweet Hypothesis in Autism Spectrum Disorder. Clin. Neuropharmacol. 2017, 40, 120–130. [Google Scholar] [CrossRef]
- Berezin, V.; Walmod, P.S.; Filippov, M.; Dityatev, A. Targeting of ECM molecules and their metabolizing enzymes and receptors for the treatment of CNS diseases. In Progress in Brain Research; Elsevier: Amsterdam, The Netherlands, 2014; Volume 214, pp. 353–388. [Google Scholar] [CrossRef]
- Yick, L.-W.; So, K.-F.; Cheung, P.-T.; Wu, W.-T. Lithium Chloride Reinforces the Regeneration-Promoting Effect of Chondroitinase ABC on Rubrospinal Neurons after Spinal Cord Injury. J. Neurotrauma 2004, 21, 932–943. [Google Scholar] [CrossRef] [PubMed]
- Eisele, B.S.; Luka, Z.; Wu, A.J.; Yang, F.; Hale, A.T.; York, J.D. Sulfation of glycosaminoglycans depends on the catalytic activity of lithium-inhibited phosphatase BPNT2 in vitro. J. Biol. Chem. 2021, 297, 101293. [Google Scholar] [CrossRef]
- Zhao, R.-R.; Muir, E.M.; Alves, J.N.; Rickman, H.; Allan, A.Y.; Kwok, J.C.; Roet, K.C.D.; Verhaagen, J.; Schneider, B.L.; Bensadoun, J.-C.; et al. Lentiviral vectors express chondroitinase ABC in cortical projections and promote sprouting of injured corticospinal axons. J. Neurosci. Methods 2011, 201, 228–238. [Google Scholar] [CrossRef] [PubMed]
- Alves, J.N.; Muir, E.M.; Andrews, M.R.; Ward, A.; Michelmore, N.; Dasgupta, D.; Verhaagen, J.; Moloney, E.B.; Keynes, R.J.; Fawcett, J.W.; et al. AAV vector-mediated secretion of chondroitinase provides a sensitive tracer for axonal arborisations. J. Neurosci. Methods 2014, 227, 107–120. [Google Scholar] [CrossRef] [PubMed]
- Blanco, I.; Conant, K. Extracellular matrix remodeling with stress and depression: Studies in human, rodent and zebrafish models. Eur. J. Neurosci. 2021, 53, 3879–3888. [Google Scholar] [CrossRef] [PubMed]
- Paveliev, M.; Fenrich, K.K.; Kislin, M.; Kuja-Panula, J.; Kulesskiy, E.; Varjosalo, M.; Kajander, T.; Mugantseva, E.; Ahonen-Bishopp, A.; Khiroug, L.; et al. HB-GAM (pleiotrophin) reverses inhibition of neural regeneration by the CNS extracellular matrix. Sci. Rep. 2016, 6, 33916. [Google Scholar] [CrossRef] [PubMed]
- Rauvala, H.; Paveliev, M.; Kuja-Panula, J.; Kulesskaya, N. Inhibition and enhancement of neural regeneration by chondroitin sulfate proteoglycans. Neural Regen. Res. 2017, 12, 687. [Google Scholar] [CrossRef]
- Kulesskaya, N.; Molotkov, D.; Sliepen, S.; Mugantseva, E.; Garcia Horsman, A.; Paveliev, M.; Rauvala, H. Heparin-Binding Growth-Associated Molecule (Pleiotrophin) Affects Sensory Signaling and Selected Motor Functions in Mouse Model of Anatomically Incomplete Cervical Spinal Cord Injury. Front. Neurol. 2021, 12, 738800. [Google Scholar] [CrossRef]
- Kulesskaya, N.; Mugantseva, E.; Minkeviciene, R.; Acosta, N.; Rouhiainen, A.; Kuja-Panula, J.; Kislin, M.; Piirainen, S.; Paveliev, M.; Rauvala, H. Low-Molecular Weight Protamine Overcomes Chondroitin Sulfate Inhibition of Neural Regeneration. Front. Cell Dev. Biol. 2022, 10, 865275. [Google Scholar] [CrossRef] [PubMed]
- Alilain, W.J.; Horn, K.P.; Hu, H.; Dick, T.E.; Silver, J. Functional regeneration of respiratory pathways after spinal cord injury. Nature 2011, 475, 196–200. [Google Scholar] [CrossRef]
- Massey, J.M.; Hubscher, C.H.; Wagoner, M.R.; Decker, J.A.; Amps, J.; Silver, J.; Onifer, S.M. Chondroitinase ABC Digestion of the Perineuronal Net Promotes Functional Collateral Sprouting in the Cuneate Nucleus after Cervical Spinal Cord Injury. J. Neurosci. 2006, 26, 4406–4414. [Google Scholar] [CrossRef] [PubMed]
- Varbanov, H.; Jia, S.; Kochlamazashvili, G.; Bhattacharya, S.; Buabeid, M.A.; El Tabbal, M.; Hayani, H.; Stoyanov, S.; Sun, W.; Thiesler, H.; et al. Rescue of synaptic and cognitive functions in polysialic acid-deficient mice and dementia models by short polysialic acid fragments. Neurobiol. Dis. 2023, 180, 106079. [Google Scholar] [CrossRef]
- Wang, Y.; LeDue, J.M.; Murphy, T.H. Multiscale imaging informs translational mouse modeling of neurological disease. Neuron 2022, 110, 3688–3710. [Google Scholar] [CrossRef]
- Paveliev, M.; Kislin, M.; Molotkov, D.; Yuryev, M.; Rauvala, H.; Khiroug, L. Acute Brain Trauma in Mice Followed By Longitudinal Two-photon Imaging. J. Vis. Exp. 2014, 51559. [Google Scholar] [CrossRef]
- Sparks, F.T.; Liao, Z.; Li, W.; Grosmark, A.; Soltesz, I.; Losonczy, A. Hippocampal adult-born granule cells drive network activity in a mouse model of chronic temporal lobe epilepsy. Nat. Commun. 2020, 11, 6138. [Google Scholar] [CrossRef] [PubMed]
- Fu, S.; Shi, W.; Luo, T.; He, Y.; Zhou, L.; Yang, J.; Yang, Z.; Liu, J.; Liu, X.; Guo, Z.; et al. Field-dependent deep learning enables high-throughput whole-cell 3D super-resolution imaging. Nat. Methods 2023, 20, 459–468. [Google Scholar] [CrossRef] [PubMed]
- Peddie, C.J.; Genoud, C.; Kreshuk, A.; Meechan, K.; Micheva, K.D.; Narayan, K.; Pape, C.; Parton, R.G.; Schieber, N.L.; Schwab, Y.; et al. Volume electron microscopy. Nat. Rev. Methods Primer 2022, 2, 51. [Google Scholar] [CrossRef] [PubMed]
- Eisenstein, M. Seven technologies to watch in 2023. Nature 2023, 613, 794–797. [Google Scholar] [CrossRef]
- Micheva, K.D.; Gong, B.; Collman, F.; Weinberg, R.J.; Smith, S.J.; Trimmer, J.S.; Murray, K.D. Developing a Toolbox of Antibodies Validated for Array Tomography-Based Imaging of Brain Synapses. eNeuro 2023, 10. [Google Scholar] [CrossRef] [PubMed]
- Villanueva, C.B.; Stephensen, H.J.T.; Mokso, R.; Benraiss, A.; Sporring, J.; Goldman, S.A. Astrocytic engagement of the corticostriatal synaptic cleft is disrupted in a mouse model of Huntington’s disease. Proc. Natl. Acad. Sci. USA 2023, 120, e2210719120. [Google Scholar] [CrossRef]
- Tamada, H. Three-dimensional ultrastructure analysis of organelles in injured motor neuron. Anat. Sci. Int. 2023, 98, 360–369. [Google Scholar] [CrossRef]
- Haase, R.; Fazeli, E.; Legland, D.; Doube, M.; Culley, S.; Belevich, I.; Jokitalo, E.; Schorb, M.; Klemm, A.; Tischer, C. A Hitchhiker’s guide through the bio-image analysis software universe. FEBS Lett. 2022, 596, 2472–2485. [Google Scholar] [CrossRef] [PubMed]
- Belevich, I.; Jokitalo, E. DeepMIB: User-friendly and open-source software for training of deep learning network for biological image segmentation. PLoS Comput. Biol. 2021, 17, e1008374. [Google Scholar] [CrossRef] [PubMed]
- Hayashi, S.; Ohno, N.; Knott, G.; Molnár, Z. Correlative light and volume electron microscopy to study brain development. Microscopy 2023, 72, 279–286. [Google Scholar] [CrossRef] [PubMed]
- Berg, S.; Kutra, D.; Kroeger, T.; Straehle, C.N.; Kausler, B.X.; Haubold, C.; Schiegg, M.; Ales, J.; Beier, T.; Rudy, M.; et al. ilastik: Interactive machine learning for (bio)image analysis. Nat. Methods 2019, 16, 1226–1232. [Google Scholar] [CrossRef]
- Arzt, M.; Deschamps, J.; Schmied, C.; Pietzsch, T.; Schmidt, D.; Tomancak, P.; Haase, R.; Jug, F. LABKIT: Labeling and Segmentation Toolkit for Big Image Data. Front. Comput. Sci. 2022, 4, 777728. [Google Scholar] [CrossRef]
- St. Pierre, M.; Duck, S.A.; Nazareth, M.; Fung, C.; Jantzie, L.L.; Chavez-Valdez, R. Unbiased Quantitative Single-Cell Morphometric Analysis to Identify Microglia Reactivity in Developmental Brain Injury. Life 2023, 13, 899. [Google Scholar] [CrossRef] [PubMed]
- Zhang, J.; Xie, Y.; Wu, Q.; Xia, Y. Medical image classification using synergic deep learning. Med. Image Anal. 2019, 54, 10–19. [Google Scholar] [CrossRef]
- Jiang, N.; Yu, F. A Cell Counting Framework Based on Random Forest and Density Map. Appl. Sci. 2020, 10, 8346. [Google Scholar] [CrossRef]
- Jiang, N.; Yu, F. A Two-Path Network for Cell Counting. IEEE Access 2021, 9, 70806–70815. [Google Scholar] [CrossRef]
- Koyuncu, C.F.; Gunesli, G.N.; Cetin-Atalay, R.; Gunduz-Demir, C. DeepDistance: A multi-task deep regression model for cell detection in inverted microscopy images. Med. Image Anal. 2020, 63, 101720. [Google Scholar] [CrossRef]
- Wang, S.; Lin, B.; Lin, G.; Lin, R.; Huang, F.; Liu, W.; Wang, X.; Liu, X.; Zhang, Y.; Wang, F.; et al. Automated label-free detection of injured neuron with deep learning by two-photon microscopy. J. Biophotonics 2020, 13, e201960062. [Google Scholar] [CrossRef] [PubMed]
- Cai, Y.; Zhang, X.; Kovalsky, S.Z.; Ghashghaei, H.T.; Greenbaum, A. Detection and classification of neurons and glial cells in the MADM mouse brain using RetinaNet. PLoS ONE 2021, 16, e0257426. [Google Scholar] [CrossRef] [PubMed]
- Chen, D.; Nauen, D.W.; Park, H.-C.; Li, D.; Yuan, W.; Li, A.; Guan, H.; Kut, C.; Chaichana, K.L.; Bettegowda, C.; et al. Label-free imaging of human brain tissue at subcellular resolution for potential rapid intra-operative assessment of glioma surgery. Theranostics 2021, 11, 7222–7234. [Google Scholar] [CrossRef]
- Axer, M.; Amunts, K. Scale matters: The nested human connectome. Science 2022, 378, 500–504. [Google Scholar] [CrossRef] [PubMed]
- Traver, V.J.; Pla, F.; Miquel, M.; Carbo-Gas, M.; Gil-Miravet, I.; Guarque-Chabrera, J. Cocaine-Induced Preference Conditioning: A Machine Vision Perspective. Neuroinformatics 2019, 17, 343–359. [Google Scholar] [CrossRef] [PubMed]
- Campagner, A.; Ciucci, D.; Svensson, C.-M.; Figge, M.T.; Cabitza, F. Ground truthing from multi-rater labeling with three-way decision and possibility theory. Inf. Sci. 2021, 545, 771–790. [Google Scholar] [CrossRef]
- Isola, P.; Zhu, J.-Y.; Zhou, T.; Efros, A.A. Image-to-Image Translation with Conditional Adversarial Networks 2018. arXiv 2018, arXiv:1611.07004. [Google Scholar]
- Goodfellow, I.; Pouget-Abadie, J.; Mirza, M.; Xu, B.; Warde-Farley, D.; Ozair, S.; Courville, A.; Bengio, Y. Generative Adversarial Nets. In Proceedings of the Advances in Neural Information Processing Systems 27: Annual Conference on Neural Information Processing Systems 2014, Montreal, QC, Canada, 8–13 December 2014. [Google Scholar]
- Hinton, G.E.; Srivastava, N.; Krizhevsky, A.; Sutskever, I.; Salakhutdinov, R.R. Improving neural networks by preventing co-adaptation of feature detectors. arXiv 2012, arXiv:1207.0580. [Google Scholar]
- Ioffe, S.; Szegedy, C. Batch Normalization: Accelerating Deep Network Training by Reducing Internal Covariate Shift. arXiv 2015, arXiv:1502.03167. [Google Scholar]
- Nikolenko, S.; Kadurin, A.; Arkhangelskaya, E. Deep Learning; Piter: St. Petersburg, Russia, 2018. [Google Scholar]
- Krasilnikov, N. Digital Processing of 2D and 3D Images: A Tutorial; BHV-Petersburg: Moscow, Russia, 2011. [Google Scholar]
- Zeiler, M.D.; Krishnan, D.; Taylor, G.W.; Fergus, R. Deconvolutional networks. In Proceedings of the 2010 IEEE Computer Society Conference on Computer Vision and Pattern Recognition, San Francisco, CA, USA, 13–18 June 2010; IEEE: Piscataway, NJ, USA, 2010; pp. 2528–2535. [Google Scholar] [CrossRef]
- Ronneberger, O.; Fischer, P.; Brox, T. U-Net: Convolutional Networks for Biomedical Image Segmentation. In Medical Image Computing and Computer-Assisted Intervention—MICCAI 2015; Navab, N., Hornegger, J., Wells, W.M., Frangi, A.F., Eds.; Lecture Notes in Computer Science; Springer International Publishing: Cham, Switzerland, 2015; Volume 9351, pp. 234–241. [Google Scholar] [CrossRef]






Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Paveliev, M.; Egorchev, A.A.; Musin, F.; Lipachev, N.; Melnikova, A.; Gimadutdinov, R.M.; Kashipov, A.R.; Molotkov, D.; Chickrin, D.E.; Aganov, A.V. Perineuronal Net Microscopy: From Brain Pathology to Artificial Intelligence. Int. J. Mol. Sci. 2024, 25, 4227. https://doi.org/10.3390/ijms25084227
Paveliev M, Egorchev AA, Musin F, Lipachev N, Melnikova A, Gimadutdinov RM, Kashipov AR, Molotkov D, Chickrin DE, Aganov AV. Perineuronal Net Microscopy: From Brain Pathology to Artificial Intelligence. International Journal of Molecular Sciences. 2024; 25(8):4227. https://doi.org/10.3390/ijms25084227
Chicago/Turabian StylePaveliev, Mikhail, Anton A. Egorchev, Foat Musin, Nikita Lipachev, Anastasiia Melnikova, Rustem M. Gimadutdinov, Aidar R. Kashipov, Dmitry Molotkov, Dmitry E. Chickrin, and Albert V. Aganov. 2024. "Perineuronal Net Microscopy: From Brain Pathology to Artificial Intelligence" International Journal of Molecular Sciences 25, no. 8: 4227. https://doi.org/10.3390/ijms25084227
APA StylePaveliev, M., Egorchev, A. A., Musin, F., Lipachev, N., Melnikova, A., Gimadutdinov, R. M., Kashipov, A. R., Molotkov, D., Chickrin, D. E., & Aganov, A. V. (2024). Perineuronal Net Microscopy: From Brain Pathology to Artificial Intelligence. International Journal of Molecular Sciences, 25(8), 4227. https://doi.org/10.3390/ijms25084227