
Eldecalcitol Induces Minimodeling-Based Bone Formation and Inhibits Sclerostin Synthesis Preferentially in the Epiphyses Rather than the Metaphyses of the Long Bones in Rats
 , ,
, ,
Abstract
1. Introduction
2. Results
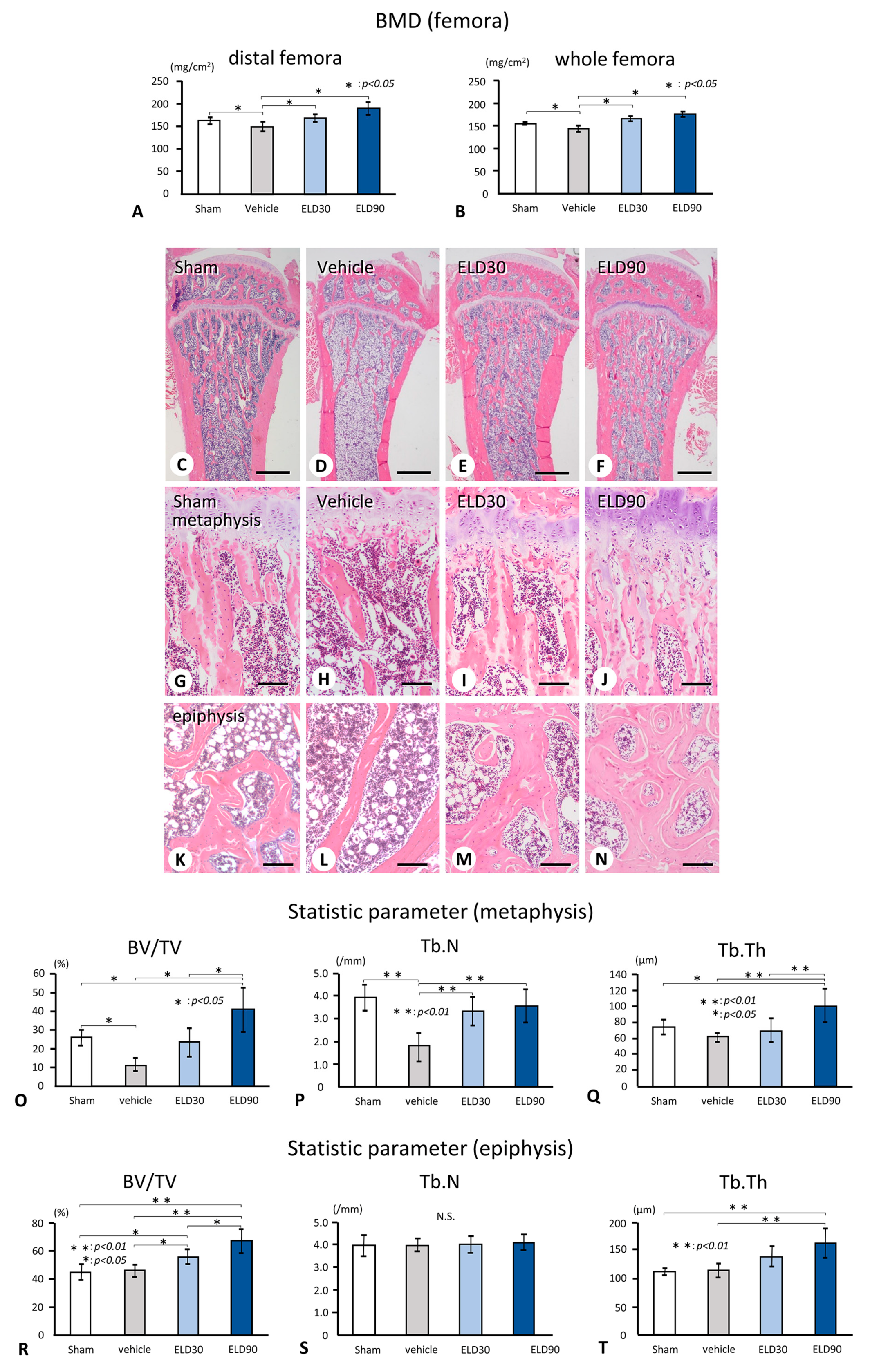
2.1. Femoral BMD and Bone Histomorphometrical Analyses of the Tibial Metapsyses and Epiphyses in Sham, Vehicle, ELD30, and ELD90 Groups
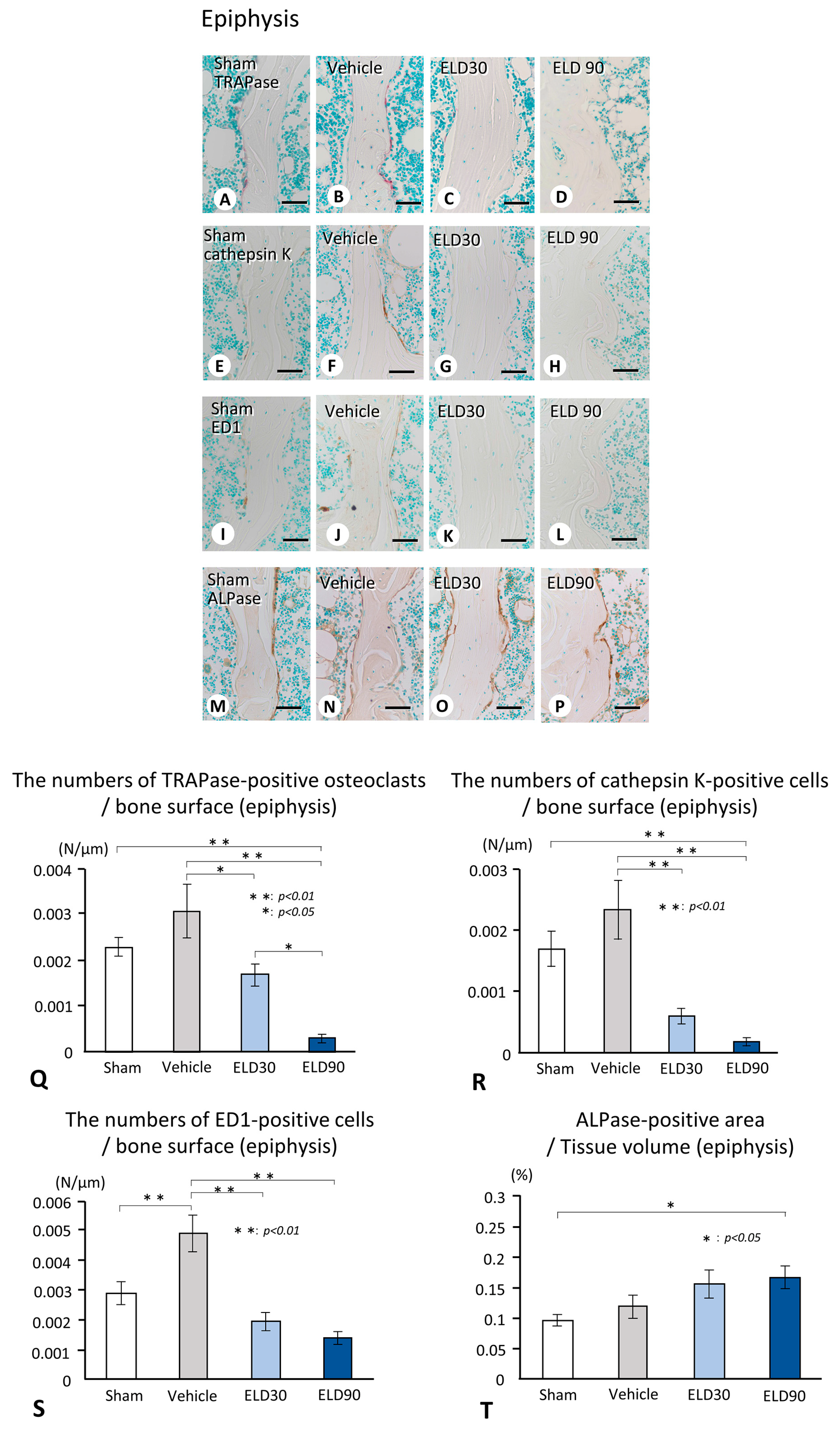
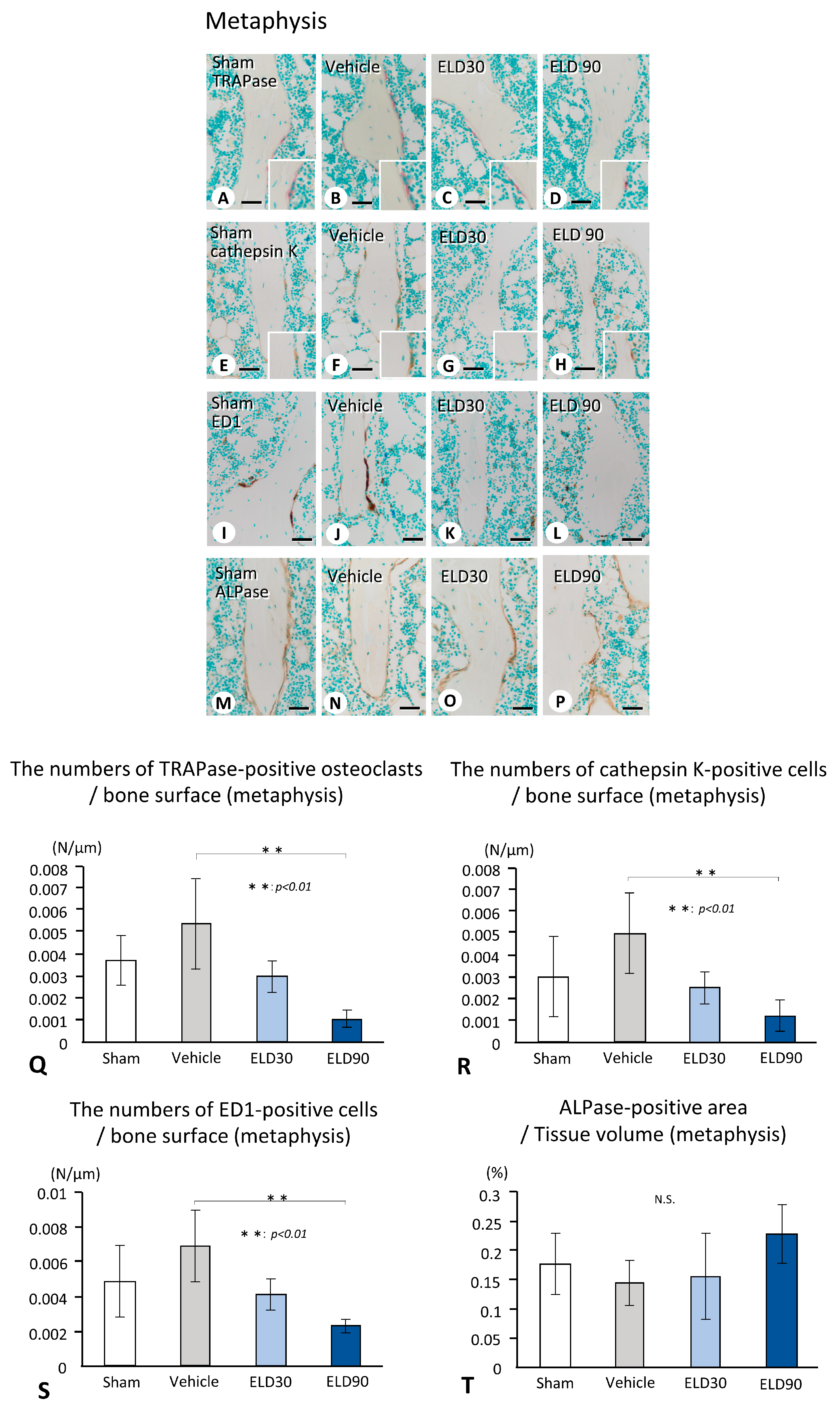
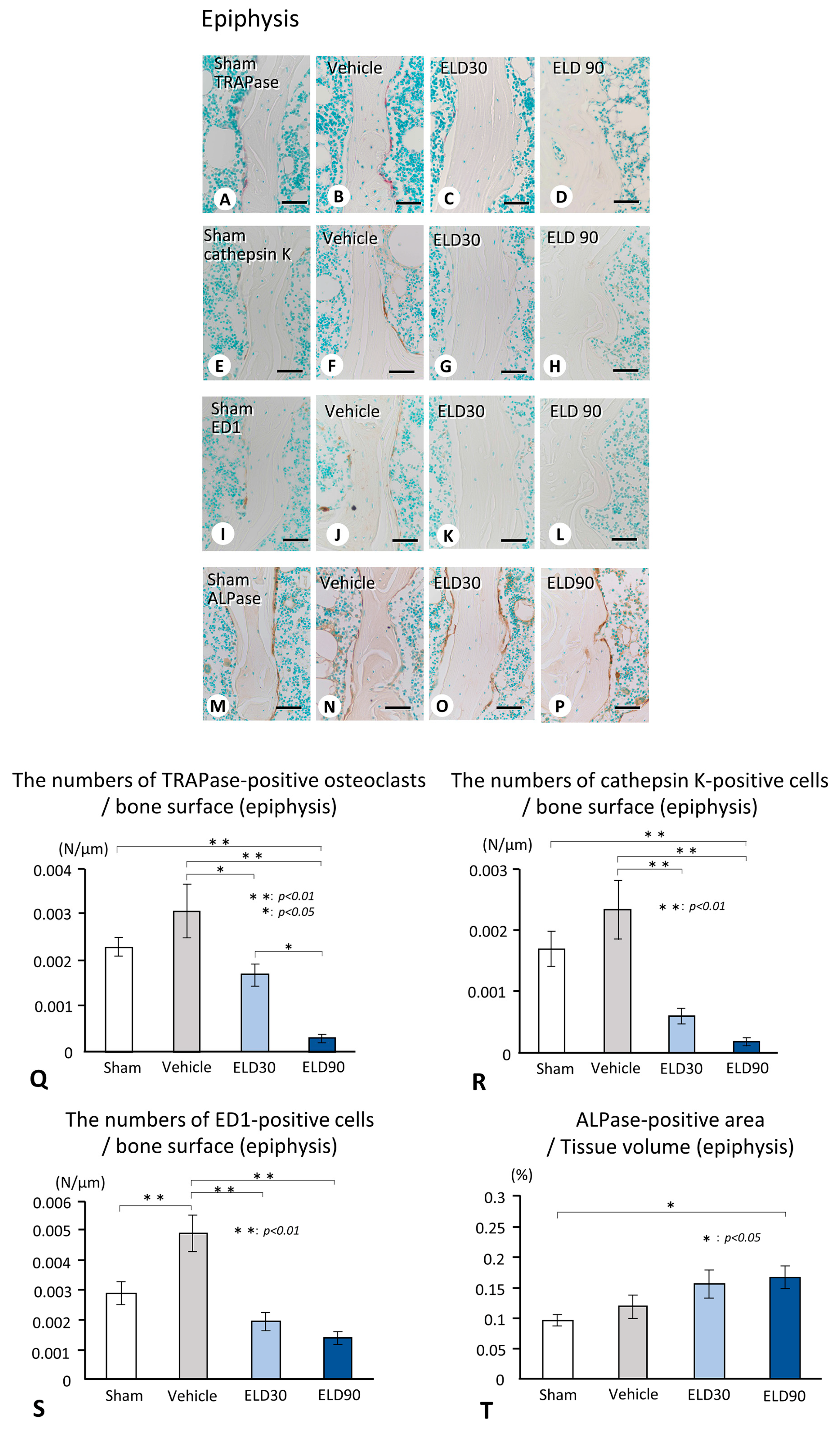
2.2. The Distribution of TRAPase-Reactive, Cathepsin K-Positive, and ED1-Immunoreactive Osteoclasts, and ALPase-Positive Osteoblasts in the ROI of Metaphysis and Epiphysis
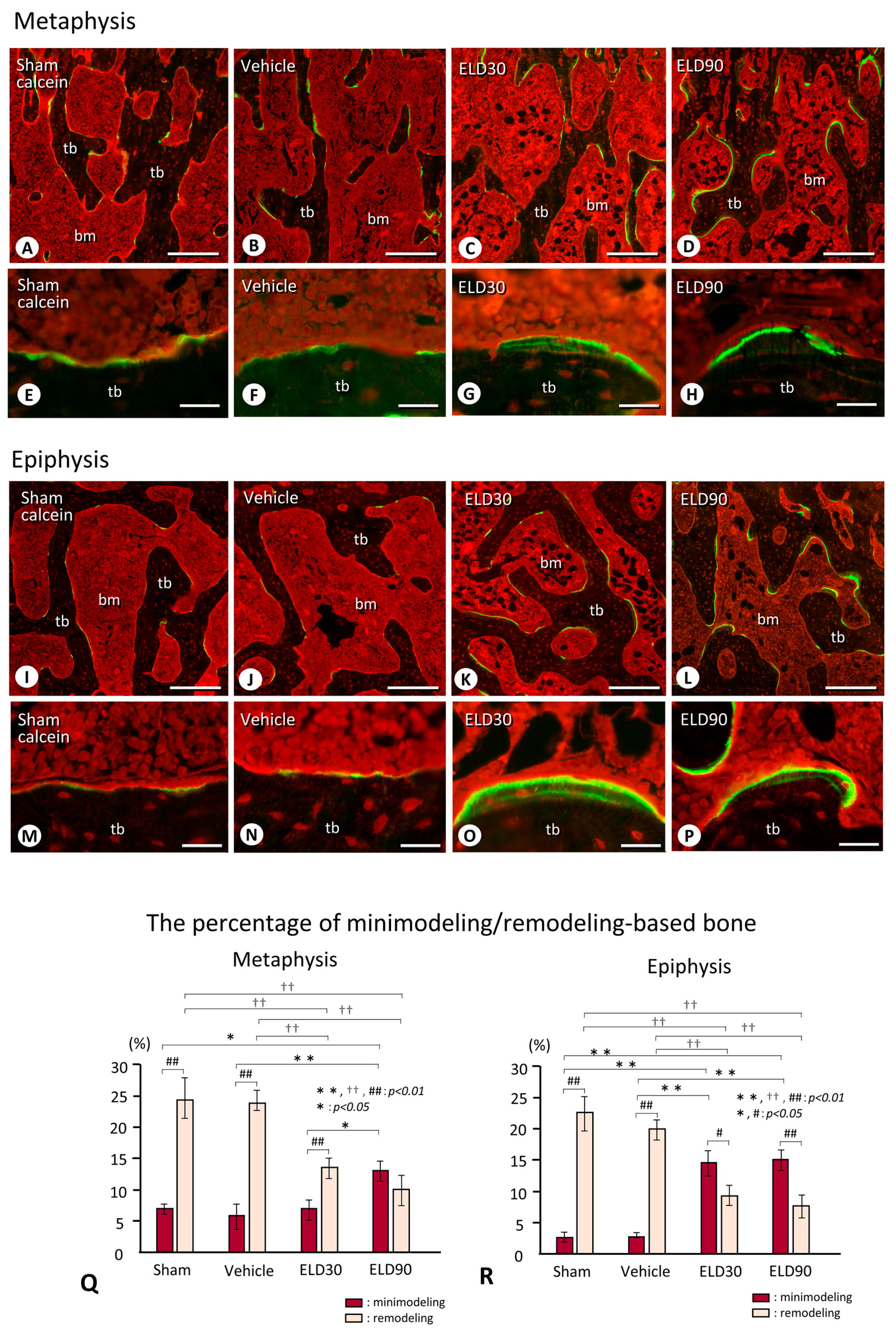
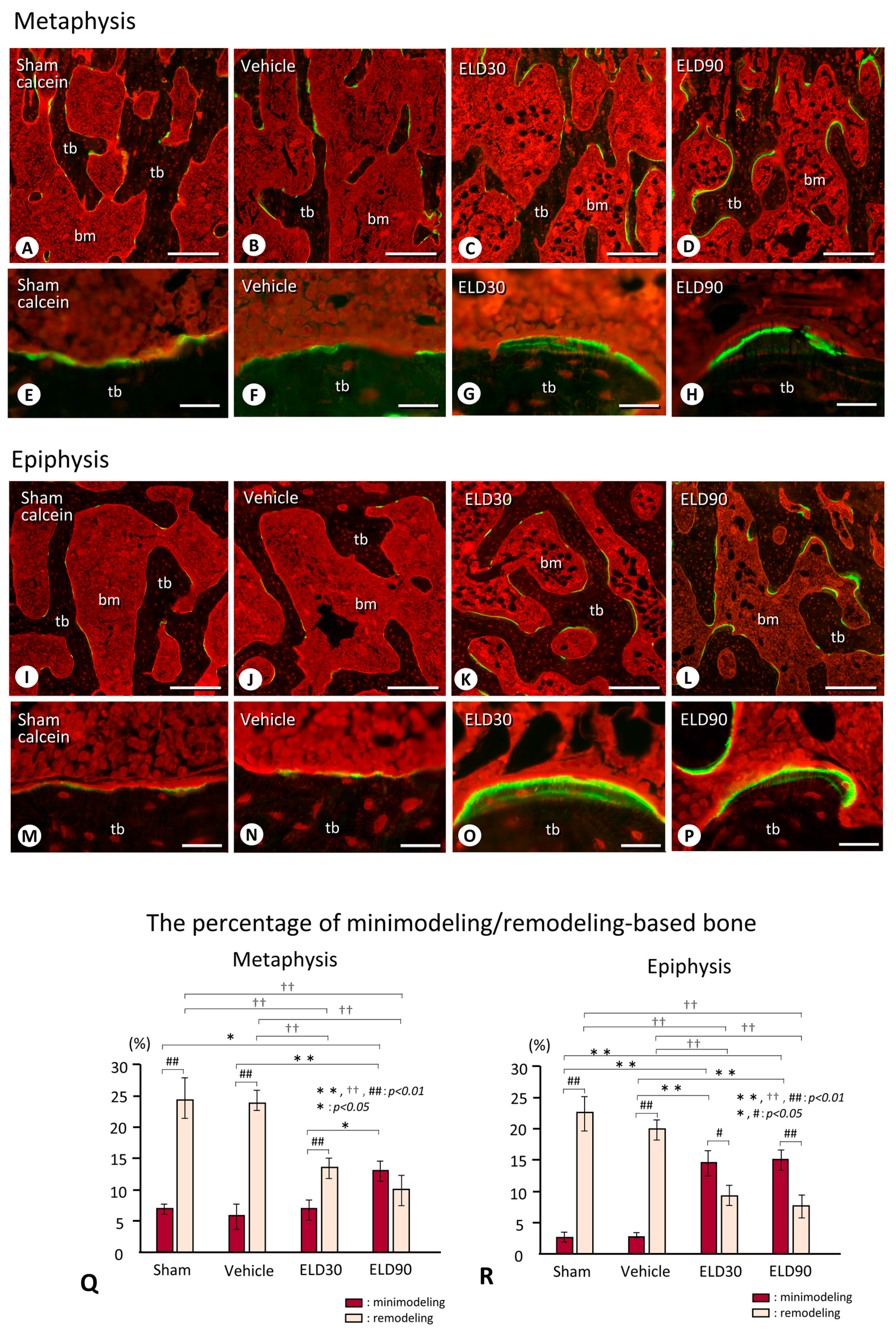
2.3. Minimodeling-Based Bone Formation and Remodeling-Based Bone Formation in Tibial Epiphyses and Metaphyses
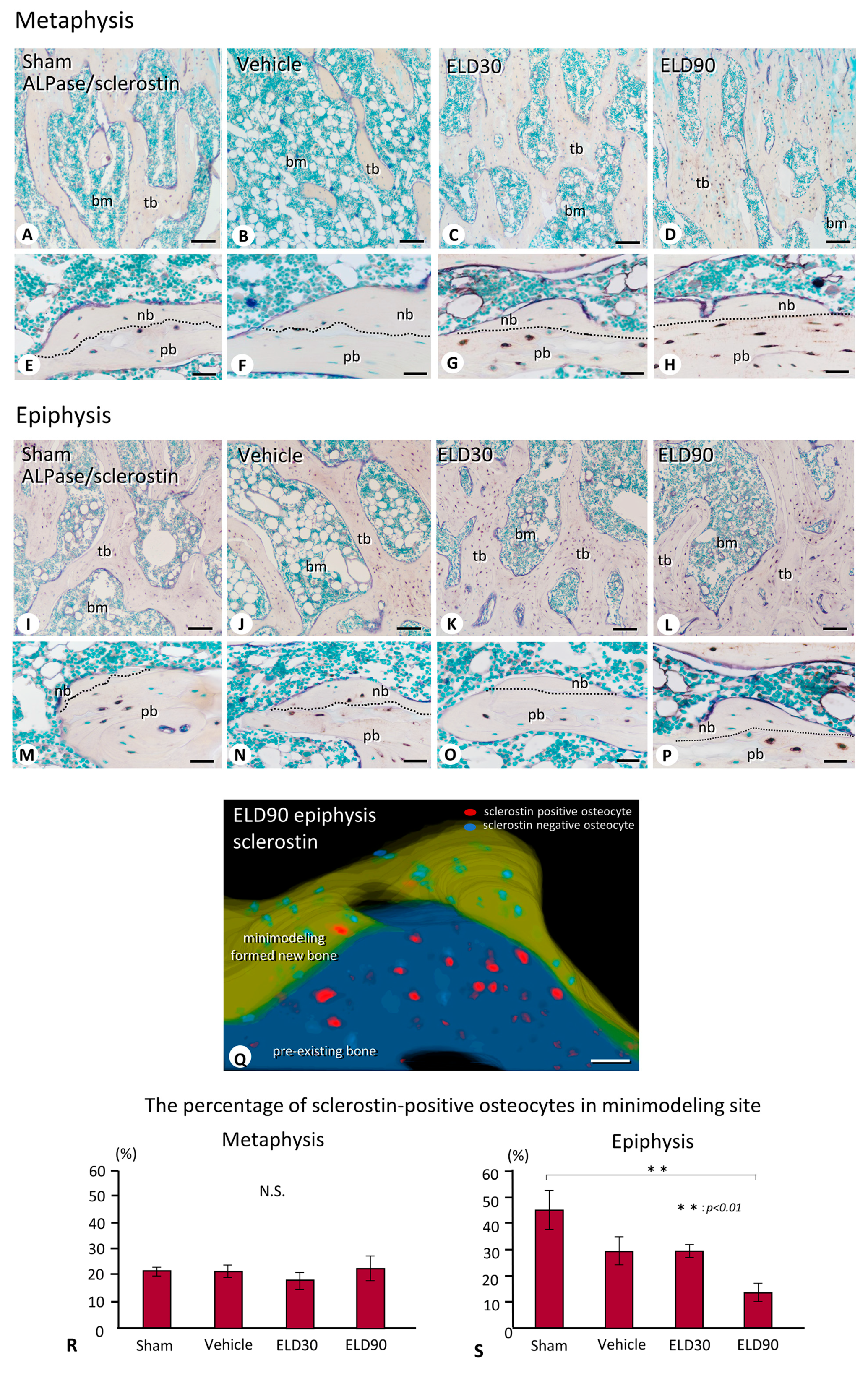
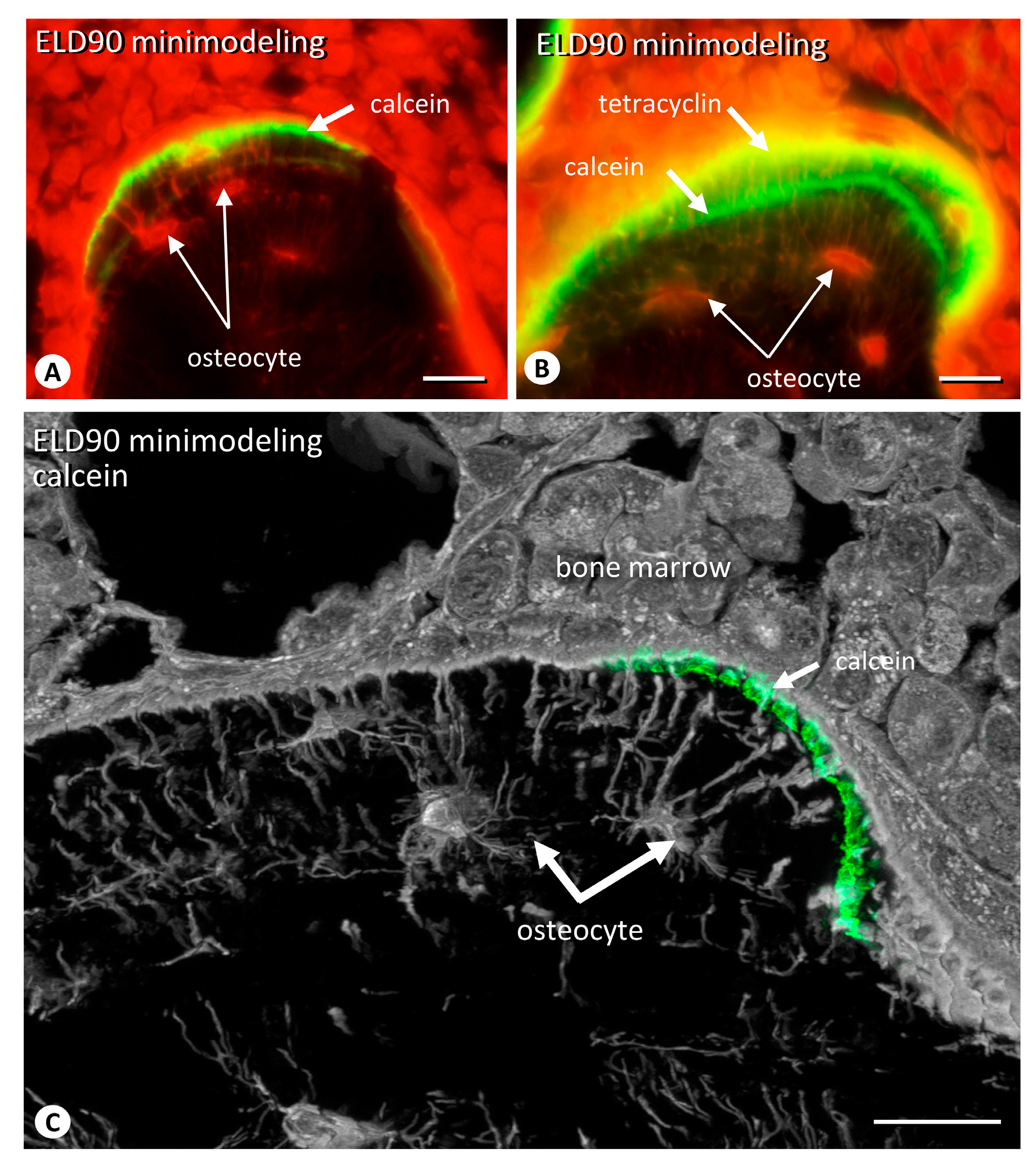
2.4. The Distribution of Sclerostin-Positive Osteocytes in Minimodeling-Induced New Bone
2.5. The Distribution of Osteocytic Lacunar Canalicular Network in Minimodeling-Induced New Bone
3. Discussion
4. Material and Methods
4.1. Animals and Tissue Preparation
4.2. Detection of BMD
4.3. Immunostaining for Tissue-Non-Specific Alkaline Phosphatase (ALPase), ED1, Cathepsin K, and Sclerostin
4.4. Enzyme Histochemistry for Tartrate-Resistant acid Phosphatase (TRAPase)
4.5. Bone Histomorphometry of BV/TV, Tb.N, Tb.Th, ALPase-Positive Osteoblastic Area, and the Analyses of TRAPase-Reactive, ED1-Positive, and Cathepsin K-Reactive Cells in Metaphyses and Epiphyses
4.6. Quantification of the Frequency of Minimodeling-Based/Remodeling-Based Bone in Metaphyses and Epiphyses
4.7. Quantification of the Percentage of Sclerostin-Positive Osteocytes in Minimodeling-Based and Remodeling-Based Bone in Metaphyses and Epiphyses
4.8. Statistical Analysis
4.9. Three-Dimensional Reconstruction of Minimodeling-Based Bone Formation including Sclerostin-Positive/Negative Osteocytes
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Frost, H.M. Dynamics of bone remodeling. In Bone Biodynamics; Frost, H.M., Ed.; Little and Brown Co.: Boston, MA, USA, 1964; pp. 315–333. [Google Scholar]
- Jee, W.S. The Skeletal tissues. In Histology, Cell and Tissue Biology, 5th ed.; Weiss, L., Ed.; The MacMillan Press: London, UK, 1983. [Google Scholar]
- Frost, H.M. Tetracycline-based histological analysis of bone remodeling. Calcif. Tissue. Res. 1969, 3, 211–237. [Google Scholar] [CrossRef] [PubMed]
- Frost, H.M. Skeletal structural adaptations to mechanical usage (SATMU): 2. Redefining Wolff’s law: The bone modeling problem. Anat. Rec. 1990, 226, 403–413. [Google Scholar] [CrossRef] [PubMed]
- Nordin, B.E.; Need, A.G.; Chatterton, B.E.; Horowitz, M.; Morris, H.A. The relative contributions of age and years since menopause to postmenopausal bone loss. J. Clin. Endocrinol. Metab. 1990, 70, 83–88. [Google Scholar] [CrossRef] [PubMed]
- Hattner, R.; Epker, B.N.; Frost, H.M. Suggested sequential mode of control of changes in cell behaviour in adult bone remodelling. Nature 1965, 206, 489–490. [Google Scholar] [CrossRef] [PubMed]
- Kobayashi, S.; Takahashi, H.E.; Ito, A.; Saito, N.; Nawata, M.; Horiuchi, H.; Ohta, H.; Ito, A.; Iorio, R.; Yamamoto, N.; et al. Trabecular minimodeling in human iliac bone. Bone 2003, 32, 163–169. [Google Scholar] [CrossRef]
- Tanaka, Y.; Nakamura, T.; Nishida, S.; Suzuki, K.; Takeda, S.; Sato, K.; Nishii, Y. Effects of a synthetic vitamin D analog, ED-71, on bone dynamics and strength in cancellous and cortical bone in prednisolone-treated rats. J. Bone. Miner. Res. 1996, 11, 325–336. [Google Scholar] [CrossRef] [PubMed]
- Uchiyama, Y.; Higuchi, Y.; Takeda, S.; Masaki, T.; Shira-Ishi, A.; Sato, K.; Kubodera, N.; Ikeda, K.; Ogata, E. ED-71, a vitamin D analog, is a more potent inhibitor of bone resorption than alfacalcidol in an estrogen-deficient rat model of osteoporosis. Bone 2002, 30, 582–588. [Google Scholar] [CrossRef] [PubMed]
- Matsumoto, T.; Miki, T.; Hagino, H.; Sugimoto, T.; Okamoto, S.; Hirota, T.; Tanigawara, Y.; Hayashi, Y.; Fukunaga, M.; Shiraki, M.; et al. A new active vitamin D, ED-71, increases bone mass in osteoporotic patients under vitamin D supplementation: A randomized, double-blind, placebo-controlled clinical trial. J. Clin. Endocrinol. Metab. 2005, 90, 5031–5036. [Google Scholar] [CrossRef]
- Matsumoto, T.; Takano, T.; Yamakido, S.; Takahashi, F.; Tsuji, N. Comparison of the effects of eldecalcitol and alfacalcidol on bone and calcium metabolism. J. Steroid. Biochem. Mol. Biol. 2010, 121, 261–264. [Google Scholar] [CrossRef] [PubMed]
- Matsumoto, T.; Ito, M.; Hayashi, Y.; Hirota, T.; Tanigawara, Y.; Sone, T.; Fukunaga, M.; Shiraki, M.; Nakamura, T. A new active vitamin D3 analog, eldecalcitol, prevents the risk of osteoporotic fractures—A randomized, active comparator, double-blind study. Bone 2011, 49, 605–612. [Google Scholar] [CrossRef]
- Miyamoto, K.; Murayama, E.; Ochi, K.; Watanabe, H.; Kubodera, N. Synthetic studies of vitamin D analogues XIV Synthesis and calcium regulating activity of vitamin D3 bearing a hydroxyalkoxy group at the 2β-position. Chem. Pharm. Bull. 1993, 41, 1111–1113. [Google Scholar] [CrossRef] [PubMed]
- Freitas, P.H.L.; Hasegawa, T.; Takeda, S.; Sasaki, M.; Tabata, C.; Oda, K.; Li, M.; Saito, H.; Amizuka, N. Eldecalcitol, a second-generation vitamin D analog, drives bone minimodeling and reduces osteoclastic number in trabecular bone of ovariectomized rats. Bone 2011, 49, 335–342. [Google Scholar] [CrossRef]
- Hasegawa, T.; Yamamoto, T.; Sakai, S.; Miyamoto, Y.; Hongo, H.; Qiu, Z.; Abe, M.; Takeda, S.; Oda, K.; Freitas, P.H.L.; et al. Histological effects of the combined administration of eldecalcitol and a parathyroid hormone in the metaphyseal trabeculae of ovariectomized rats. J. Histochem. Cytochem. 2019, 67, 169–184. [Google Scholar] [CrossRef]
- Parfitt, A.M. The physiologic and clinical significance of bone histomorphometricaal data. In Bone Histomorphometry: Techniques and Interpretation; Recker, R.R., Ed.; CRC Press: Boca Raton, FL, USA, 1983; pp. 143–223. [Google Scholar]
- Jee, W.S.; Tian, X.Y.; Setterberg, R.B. Cancellous bone minimodeling-based formation: A Frost, Takahashi legacy. J. Musculoskelet. Neuronal. Interact. 2007, 7, 232–239. [Google Scholar] [CrossRef] [PubMed]
- Dempster, D.W.; Birchman, R.; Xu, R.; Lindsay, R.; Shen, V. Temporal changes in cancellous bone structure of rats immediately after ovariectomy. Bone 1995, 16, 157–161. [Google Scholar] [CrossRef]
- Hilliam, R.A.; Skerry, T.M. Inhibition of bone resorption and stimulation of formation by mechanical loading of the modeling rat ulna in vivo. J. Bone. Miner. Res. 1995, 10, 683–689. [Google Scholar] [CrossRef] [PubMed]
- Baldock, P.A.; Morris, H.A.; Need, A.G.; Moore, R.J.; Durbridge, T.C. Variation in the short-term changes in bone cell activity in three regions of the distal femur immediately following ovariectomy. J. Bone. Miner. Res. 1998, 13, 1451–1457. [Google Scholar] [CrossRef]
- Baldock, P.A.; Need, A.G.; Moore, R.J.; Durbridge, T.C.; Morris, H.A. Discordance between bone turnover and bone loss: Effects of aging and ovariectomy in the rat. J. Bone. Miner. Res. 1999, 14, 1442–1448. [Google Scholar] [CrossRef] [PubMed]
- Cosman, F.; Crittenden, D.B.; Adachi, J.D.; Binkley, N.; Czerwinski, E.; Ferrari, S.; Hofbauer, L.C.; Lau, E.; Lewiecki, E.M.; Miyauchi, A.; et al. Romosozumab treatment in postmenopausal women with osteoporosis. N. Engl. J. Med. 2016, 375, 1532–1543. [Google Scholar] [CrossRef]
- Bandeira, L.; Lewiecki, E.M.; Bilezikian, J.P. Romosozumab for the treatment of osteoporosis. Expert. Opin. Biol. Ther. 2017, 17, 255–263. [Google Scholar] [CrossRef]
- Ominsky, M.S.; Boyd, S.K.; Varela, A.; Jolette, J.; Felx, M.; Doyle, N.; Mellal, N.; Smith, S.Y.; Locher, K.; Buntich, S.; et al. Romosozumab improves bone mass and strength while maintaining bone quality in ovariectomized cynomolgus monkeys. J. Bone. Miner. Res. 2017, 32, 788–801. [Google Scholar] [CrossRef] [PubMed]
- Chavassieux, P.; Chapurlat, R.; Portero-Muzy, N.; Roux, J.P.; Garcia, P.; Brown, J.P.; Libanati, C.; Boyce, R.W.; Wang, A.; Grauer, A. Bone-forming and antiresorptive effects of romosozumab in postmenopausal women with osteoporosis: Bone histomorphometry and microcomputed tomography analysis after 2 and 12 months of treatment. J. Bone. Miner. Res. 2019, 34, 1597–1608. [Google Scholar] [CrossRef]
- Plotkin, L.I.; Bellido, T. Osteocytic signalling pathways as therapeutic targets for bone fragility. Nat. Rev. Endocrinol. 2016, 12, 593–605. [Google Scholar] [CrossRef] [PubMed]
- Hasegawa, T.; Hongo, H.; Yamamoto, T.; Abe, M.; Yoshino, H.; Haraguchi-Kitakamae, M.; Ishizu, H.; Shimizu, T.; Iwasaki, N.; Amizuka, N. Matrix vesicle-mediated mineralization and osteocytic regulation of bone mineralization. Int. J. Mol. Sci. 2022, 23, 9941. [Google Scholar] [CrossRef]
- Hikata, T.; Hasegawa, T.; Horiuchi, K.; Fujita, N.; Iwanami, A.; Watanabe, K.; Ishii, K.; Nakamura, M.; Amizuka, N.; Matsumoto, M. Histomorphometric analysis of minimodeling in the vertebrae in postmenopausal patients treated with anti-osteoporotic agents. Bone. Rep. 2016, 5, 286–291. [Google Scholar] [CrossRef] [PubMed]
- Winkler, D.G.; Sutherland, M.K.; Geoghegan, J.C.; Yu, C.; Hayes, T.; Skonier, J.E.; Shpektor, D.; Jonas, M.; Kovacevich, B.R.; Staehling-Hampton, K.; et al. Osteocyte control of bone formation via sclerostin, a novel BMP antagonist. EMBO J. 2003, 22, 6267–6276. [Google Scholar] [CrossRef] [PubMed]
- Van Bezooijen, R.L.; Roelen, B.A.; Visser, A.; van der Wee-Pals, L.; de Wilt, E.; Karperien, M.; Hamersma, H.; Papapoulos, S.E.; ten Dijke, P.; Löwik, C.W. Sclerostin is an osteocyte-expressed negative regulator of bone formation, but not a classical BMP antagonist. J. Exp. Med. 2004, 199, 805–814. [Google Scholar] [CrossRef]
- Li, X.; Zhang, Y.; Kang, H.; Liu, W.; Liu, P.; Zhang, J.; Harris, S.E.; Wu, D. Sclerostin binds to LRP5/6 and antagonizes canonical Wnt signaling. J. Biol. Chem. 2005, 280, 19883–19887. [Google Scholar] [CrossRef] [PubMed]
- Poole, K.E.; van Bezooijen, R.L.; Loveridge, N.; Hamersma, H.; Papapoulos, S.E.; Löwik, C.W.; Reeve, J. Sclerostin is a delayed secreted product of osteocytes that inhibits bone formation. FASEB J. 2005, 19, 1842–1844. [Google Scholar] [CrossRef]
- Silvestrini, G.; Ballanti, P.; Leopizzi, M.; Sebastiani, M.; Berni, S.; Di Vito, M.; Bonucci, E. Effects of intermittent parathyroid hormone (PTH) administration on SOST mRNA and protein in rat bone. J. Mol. Histol. 2007, 38, 261–269. [Google Scholar] [CrossRef]
- Robling, A.G.; Niziolek, P.J.; Baldridge, L.A.; Condon, K.W.; Allen, M.R.; Alam, I.; Mantila, S.M.; Gluhak-Heinrich, J.; Bellido, T.M.; Harris, S.E.; et al. Mechanical stimulation of bone in vivo reduces osteocyte expression of Sost/sclerostin. J. Biol. Chem. 2008, 283, 5866–5875. [Google Scholar] [CrossRef] [PubMed]
- Hasegawa, T.; Tokunaga, S.; Yamamoto, T.; Sakai, M.; Hongo, H.; Kawata, T.; Amizuka, N. Evocalcet rescues secondary hyperparathyroidism-driven cortical porosity in chronic kidney disease male rats. Endocrinology 2023, 164, bqad022. [Google Scholar] [CrossRef]
- Oda, K.; Amaya, Y.; Fukushi-Irié, M.; Kinameri, Y.; Ohsuye, K.; Kubota, I.; Fujimura, S.; Kobayashi, J. A general method for rapid purification of soluble versions of glycosylphosphatidylinositol-anchored proteins expressed in insect cells: An application for human tissue-nonspecific alkaline phosphatase. J. Biochem. 1999, 126, 694–699. [Google Scholar] [CrossRef] [PubMed]
- Parfitt, A.M.; Drezner, M.K.; Glorieux, F.H.; Kanis, J.A.; Malluche, H.; Meunier, P.J.; Ott, S.M.; Recker, R.R. Bone histomorphometry: Standardization of nomenclature, symbols, and units. Report of the ASBMR Histomorphometry Nomenclature Committee. J. Bone. Miner. Res. 1987, 2, 595–610. [Google Scholar] [CrossRef] [PubMed]
- Schindelin, J.; Arganda-Carreras, I.; Frise, E.; Kaynig, V.; Longair, M.; Pietzsch, T.; Preibisch, S.; Rueden, C.; Saalfeld, S.; Schmid, B.; et al. Fiji: An open-source platform for biological-image analysis. Nat. Methods 2012, 9, 676–682. [Google Scholar] [CrossRef] [PubMed]




{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Experimental Group | Operation | Treatment |
|---|---|---|
| Sham | Sham | Vehicle |
| Vehicle | OVX | Vehicle |
| ELD30 | OVX | ELD 30 ng/kgBW |
| ELD90 | OVX | ELD 90 ng/kgBW |
| Tissue | Target region | Analysis item |
| Left femur | Whole and distal | BMD |
| Right and left tibiae | Metaphysis | Histological assessment |
| Epiphysis | Bone histomorphometry |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Hasegawa, T.; Yamamoto, T.; Hongo, H.; Yamamoto, T.; Haraguchi-Kitakamae, M.; Ishizu, H.; Shimizu, T.; Saito, H.; Sakai, S.; Yogo, K.; et al. Eldecalcitol Induces Minimodeling-Based Bone Formation and Inhibits Sclerostin Synthesis Preferentially in the Epiphyses Rather than the Metaphyses of the Long Bones in Rats. Int. J. Mol. Sci. 2024, 25, 4257. https://doi.org/10.3390/ijms25084257
Hasegawa T, Yamamoto T, Hongo H, Yamamoto T, Haraguchi-Kitakamae M, Ishizu H, Shimizu T, Saito H, Sakai S, Yogo K, et al. Eldecalcitol Induces Minimodeling-Based Bone Formation and Inhibits Sclerostin Synthesis Preferentially in the Epiphyses Rather than the Metaphyses of the Long Bones in Rats. International Journal of Molecular Sciences. 2024; 25(8):4257. https://doi.org/10.3390/ijms25084257
Chicago/Turabian StyleHasegawa, Tomoka, Tomomaya Yamamoto, Hiromi Hongo, Tsuneyuki Yamamoto, Mai Haraguchi-Kitakamae, Hotaka Ishizu, Tomohiro Shimizu, Hitoshi Saito, Sadaoki Sakai, Kenji Yogo, and et al. 2024. "Eldecalcitol Induces Minimodeling-Based Bone Formation and Inhibits Sclerostin Synthesis Preferentially in the Epiphyses Rather than the Metaphyses of the Long Bones in Rats" International Journal of Molecular Sciences 25, no. 8: 4257. https://doi.org/10.3390/ijms25084257
APA StyleHasegawa, T., Yamamoto, T., Hongo, H., Yamamoto, T., Haraguchi-Kitakamae, M., Ishizu, H., Shimizu, T., Saito, H., Sakai, S., Yogo, K., Matsumoto, Y., & Amizuka, N. (2024). Eldecalcitol Induces Minimodeling-Based Bone Formation and Inhibits Sclerostin Synthesis Preferentially in the Epiphyses Rather than the Metaphyses of the Long Bones in Rats. International Journal of Molecular Sciences, 25(8), 4257. https://doi.org/10.3390/ijms25084257