
Molecular Mechanisms Responsible for the Therapeutic Potential of Mesenchymal Stem Cell-Derived Exosomes in the Treatment of Lung Fibrosis
 ,
,
Abstract
:1. Introduction
2. Therapeutic Potential of Mesenchymal Stem Cells-Derived Exosomes in Pulmonary Fibrosis
3. MSC-Exo-Dependent Suppression of Immune Cells in Fibrotic Lungs
4. MSC-Exo-Based Attenuation of Pulmonary Fibrosis Is Based on the Modulation of Wnt-Driven Signaling Pathways
5. The Role of MSC-Exo-Derived miRNAs in the Attenuation of Lung Fibrosis
6. Conclusions and Future Perspectives
Author Contributions
Funding
Data Availability Statement
Conflicts of Interest
References
- Sweis, J.J.G.; Sweis, N.W.G.; Alnaimat, F.; Jansz, J.; Liao, T.E.; Alsakaty, A.; Azam, A.; Elmergawy, H.; Hanson, H.A.; Ascoli, C.; et al. Immune-mediated lung diseases: A narrative review. Front. Med. 2023, 10, 1160755. [Google Scholar] [CrossRef]
- Koudstaal, T.; Funke-Chambour, M.; Kreuter, M.; Molyneaux, P.L.; Wijsenbeek, M.S. Pulmonary fibrosis: From pathogenesis to clinical decision-making. Trends Mol. Med. 2023, 29, 1076–1087. [Google Scholar] [CrossRef] [PubMed]
- Savin, I.A.; Zenkova, M.A.; Sen’kova, A.V. Bronchial Asthma, Airway Remodeling and Lung Fibrosis as Successive Steps of One Process. Int. J. Mol. Sci. 2023, 24, 16042. [Google Scholar] [CrossRef] [PubMed]
- Nedeva, D.; Kowal, K.; Mihaicuta, S.; Guidos Fogelbach, G.; Steiropoulos, P.; Jose Chong-Neto, H.; Tiotiu, A. Epithelial alarmins: A new target to treat chronic respiratory diseases. Expert. Rev. Respir. Med. 2023, 17, 773–786. [Google Scholar] [CrossRef] [PubMed]
- Mutsaers, S.E.; Miles, T.; Prêle, C.M.; Hoyne, G.F. Emerging role of immune cells as drivers of pulmonary fibrosis. Pharmacol. Ther. 2023, 252, 108562. [Google Scholar] [CrossRef]
- Willis, B.C.; Liebler, J.M.; Luby-Phelps, K.; Nicholson, A.G.; Crandall, E.D.; du Bois, R.M.; Borok, Z. Induction of epithelial-mesenchymal transition in alveolar epithelial cells by transforming growth factor-beta1: Potential role in idiopathic pulmonary fibrosis. Am. J. Pathol. 2005, 166, 1321–1332. [Google Scholar] [CrossRef] [PubMed]
- Selvarajah, B.; Platé, M.; Chambers, R.C. Pulmonary fibrosis: Emerging diagnostic and therapeutic strategies. Mol. Asp. Med. 2023, 94, 101227. [Google Scholar] [CrossRef] [PubMed]
- Kreuter, M.; Ladner, U.M.; Costabel, U.; Jonigk, D.; Heussel, C.P. The Diagnosis and Treatment of Pulmonary Fibrosis. Dtsch. Arztebl. Int. 2021, 118, 152–162. [Google Scholar] [CrossRef]
- Singh, V.; Ulasov, I.; Gupta, S.; Singh, A.; Roy, V.K.; Kharwar, R.K. Idiopathic Pulmonary Fibrosis: Where do We Stand and How Far to Go? Discov. Med. 2024, 36, 22–47. [Google Scholar] [CrossRef]
- Hadi, D.D.; Marsool, M.D.M.; Marsool, A.D.M.; Vora, N.; Al-Badri, S.G.; Al-Fatlawi, N.H.K.; Abbas Al Wssawi, A.F.; Al-Ibraheem, A.M.T.; Hamza, K.A.; Prajjwal, P.; et al. Idiopathic pulmonary fibrosis: Addressing the current and future therapeutic advances along with the role of Sotatercept in the management of pulmonary hypertension. Immun. Inflamm. Dis. 2023, 11, e1079. [Google Scholar] [CrossRef] [PubMed]
- Bando, M. Rethinking treatment strategies for idiopathic pulmonary fibrosis: Reevaluation of anti-inflammatory and immunosuppressive therapies. Respir. Investig. 2023, 61, 58–60. [Google Scholar] [CrossRef]
- Isshiki, T.; Naiel, S.; Vierhout, M.; Otsubo, K.; Ali, P.; Tsubouchi, K.; Yazdanshenas, P.; Kumaran, V.; Dvorkin-Gheva, A.; Kolb, M.R.J.; et al. Therapeutic strategies to target connective tissue growth factor in fibrotic lung diseases. Pharmacol. Ther. 2024, 253, 108578. [Google Scholar] [CrossRef]
- Abuserewa, S.T.; Duff, R.; Becker, G. Treatment of Idiopathic Pulmonary Fibrosis. Cureus 2021, 13, e15360. [Google Scholar] [CrossRef]
- Guo, H.; Sun, J.; Zhang, S.; Nie, Y.; Zhou, S.; Zeng, Y. Progress in understanding and treating idiopathic pulmonary fibrosis: Recent insights and emerging therapies. Front. Pharmacol. 2023, 14, 1205948. [Google Scholar] [CrossRef]
- Xu, Y.; Lan, P.; Wang, T. The Role of Immune Cells in the Pathogenesis of Idiopathic Pulmonary Fibrosis. Medicina 2023, 59, 1984. [Google Scholar] [CrossRef] [PubMed]
- Hussain, Y.; Khan, H. Immunosuppressive Drugs. In Encyclopedia of Infection and Immunity; Elsevier: Amsterdam, The Netherlands, 2022; pp. 726–740. [Google Scholar]
- Belkaid, Y.; Hand, T.W. Role of the microbiota in immunity and inflammation. Cell 2014, 157, 121–141. [Google Scholar] [CrossRef]
- Ikrama, M.; Usama, M.; Israr, S.; Humayon, M. Pulmonary fibrosis: Is stem cell therapy the way forward? J. Taibah Univ. Med. Sci. 2023, 19, 82–89. [Google Scholar] [CrossRef] [PubMed]
- Guo, Z.; Zhang, Y.; Yan, F. Potential of Mesenchymal Stem Cell-Based Therapies for Pulmonary Fibrosis. DNA Cell Biol. 2022, 41, 951–965. [Google Scholar] [CrossRef]
- Vats, A.; Chaturvedi, P. The Regenerative Power of Stem Cells: Treating Bleomycin-Induced Lung Fibrosis. Stem Cells Cloning 2023, 16, 43–59. [Google Scholar] [CrossRef]
- Chen, Y.; Liu, X.; Tong, Z. Mesenchymal Stem Cells in Radiation-Induced Pulmonary Fibrosis: Future Prospects. Cells 2022, 12, 6. [Google Scholar] [CrossRef] [PubMed]
- Harrell, C.R.; Djonov, V.; Volarevic, V. The Cross-Talk between Mesenchymal Stem Cells and Immune Cells in Tissue Repair and Regeneration. Int. J. Mol. Sci. 2021, 22, 2472. [Google Scholar] [CrossRef] [PubMed]
- Harrell, C.R.; Fellabaum, C.; Jovicic, N.; Djonov, V.; Arsenijevic, N.; Volarevic, V. Molecular Mechanisms Responsible for Therapeutic Potential of Mesenchymal Stem Cell-Derived Secretome. Cells 2019, 8, 467. [Google Scholar] [CrossRef]
- Meng, H.F.; Jin, J.; Wang, H.; Wang, L.S.; Wu, C.T. Recent advances in the therapeutic efficacy of hepatocyte growth factor gene-modified mesenchymal stem cells in multiple disease settings. J. Cell Mol. Med. 2022, 26, 4745–4755. [Google Scholar] [CrossRef] [PubMed]
- Harrell, C.R.; Sadikot, R.; Pascual, J.; Fellabaum, C.; Jankovic, M.G.; Jovicic, N.; Djonov, V.; Arsenijevic, N.; Volarevic, V. Mesenchymal Stem Cell-Based Therapy of Inflammatory Lung Diseases: Current Understanding and Future Perspectives. Stem Cells Int. 2019, 2019, 4236973. [Google Scholar] [CrossRef]
- Volarevic, V.; Markovic, B.S.; Gazdic, M.; Volarevic, A.; Jovicic, N.; Arsenijevic, N.; Armstrong, L.; Djonov, V.; Lako, M.; Stojkovic, M. Ethical and Safety Issues of Stem Cell-Based Therapy. Int. J. Med. Sci. 2018, 15, 36–45. [Google Scholar] [CrossRef] [PubMed]
- Harrell, C.R.; Volarevic, A.; Djonov, V.G.; Jovicic, N.; Volarevic, V. Mesenchymal Stem Cell: A Friend or Foe in Anti-Tumor Immunity. Int. J. Mol. Sci. 2021, 22, 12429. [Google Scholar] [CrossRef]
- Harrell, C.R.; Jovicic, N.; Djonov, V.; Arsenijevic, N.; Volarevic, V. Mesenchymal Stem Cell-Derived Exosomes and Other Extracellular Vesicles as New Remedies in the Therapy of Inflammatory Diseases. Cells 2019, 8, 1605. [Google Scholar] [CrossRef]
- Karn, V.; Ahmed, S.; Tsai, L.W.; Dubey, R.; Ojha, S.; Singh, H.N.; Kumar, M.; Gupta, P.K.; Sadhu, S.; Jha, N.K.; et al. Extracellular Vesicle-Based Therapy for COVID-19: Promises, Challenges and Future Prospects. Biomedicines 2021, 9, 1373. [Google Scholar] [CrossRef] [PubMed]
- Yuan, Y.G.; Wang, J.L.; Zhang, Y.X.; Li, L.; Reza, A.M.M.T.; Gurunathan, S. Biogenesis, Composition and Potential Therapeutic Applications of Mesenchymal Stem Cells Derived Exosomes in Various Diseases. Int. J. Nanomed. 2023, 18, 3177–3210. [Google Scholar] [CrossRef]
- Melo, M.M.; Cruz, F.F.; Rocco, P.R.M. Mesenchymal stromal cell therapy for chronic lung diseases: Experimental and clinical evidence. Expert. Rev. Respir. Med. 2023, 17, 223–235. [Google Scholar] [CrossRef] [PubMed]
- Li, S.; Zhang, J.; Feng, G.; Jiang, L.; Chen, Z.; Xin, W.; Zhang, X. The Emerging Role of Extracellular Vesicles from Mesenchymal Stem Cells and Macrophages in Pulmonary Fibrosis: Insights into miRNA Delivery. Pharmaceuticals 2022, 15, 1276. [Google Scholar] [CrossRef]
- Mansouri, N.; Willis, G.R.; Fernandez-Gonzalez, A.; Reis, M.; Nassiri, S.; Mitsialis, S.A.; Kourembanas, S. Mesenchymal stromal cell exosomes prevent and revert experimental pulmonary fibrosis through modulation of monocyte phenotypes. JCI Insight 2019, 4, e128060. [Google Scholar] [CrossRef]
- Lai, P.; Chen, X.; Guo, L.; Wang, Y.; Liu, X.; Liu, Y.; Zhou, T.; Huang, T.; Geng, S.; Luo, C.; et al. A potent immunomodulatory role of exosomes derived from mesenchymal stromal cells in preventing cGVHD. J. Hematol. Oncol. 2018, 11, 135. [Google Scholar] [CrossRef]
- Zhang, E.; Geng, X.; Shan, S.; Li, P.; Li, S.; Li, W.; Yu, M.; Peng, C.; Wang, S.; Shao, H.; et al. Exosomes derived from bone marrow mesenchymal stem cells reverse epithelial-mesenchymal transition potentially via attenuating Wnt/β-catenin signaling to alleviate silica-induced pulmonary fibrosis. Toxicol. Mech. Methods 2021, 31, 655–666. [Google Scholar] [CrossRef]
- Zhang, Z.; Ge, L.; Zhang, S.; Wang, J.; Jiang, W.; Xin, Q.; Luan, Y. The protective effects of MSC-EXO against pulmonary hypertension through regulating Wnt5a/BMP signalling pathway. J. Cell Mol. Med. 2020, 24, 13938–13948. [Google Scholar] [CrossRef]
- Bissonnette, E.Y.; Lauzon-Joset, J.F.; Debley, J.S.; Ziegler, S.F. Cross-Talk Between Alveolar Macrophages and Lung Epithelial Cells is Essential to Maintain Lung Homeostasis. Front. Immunol. 2020, 11, 583042. [Google Scholar] [CrossRef] [PubMed]
- Brune, K.; Frank, J.; Schwingshackl, A.; Finigan, J.; Sidhaye, V.K. Pulmonary epithelial barrier function: Some new players and mechanisms. Am. J. Physiol. Lung Cell Mol. Physiol. 2015, 308, L731-45. [Google Scholar] [CrossRef]
- Singh, S.; Anshita, D.; Ravichandiran, V. MCP-1: Function, regulation, and involvement in disease. Int. Immunopharmacol. 2021, 101, 107598. [Google Scholar] [CrossRef] [PubMed]
- Shen, B.; Liu, J.; Zhang, F.; Wang, Y.; Qin, Y.; Zhou, Z.; Qiu, J.; Fan, Y. CCR2 Positive Exosome Released by Mesenchymal Stem Cells Suppresses Macrophage Functions and Alleviates Ischemia/Reperfusion-Induced Renal Injury. Stem Cells Int. 2016, 2016, 1240301. [Google Scholar] [CrossRef] [PubMed]
- Liu, G.; Philp, A.M.; Corte, T.; Travis, M.A.; Schilter, H.; Hansbro, N.G.; Burns, C.J.; Eapen, M.S.; Sohal, S.S.; Burgess, J.K.; et al. Therapeutic targets in lung tissue remodelling and fibrosis. Pharmacol. Ther. 2021, 225, 107839. [Google Scholar] [CrossRef]
- Lei, L.; Zhao, C.; Qin, F.; He, Z.Y.; Wang, X.; Zhong, X.N. Th17 cells and IL-17 promote the skin and lung inflammation and fibrosis process in a bleomycin-induced murine model of systemic sclerosis. Clin. Exp. Rheumatol. 2016, 34, 14–22. [Google Scholar] [PubMed]
- Gurczynski, S.J.; Moore, B.B. IL-17 in the lung: The good, the bad, and the ugly. Am. J. Physiol. Lung Cell Mol. Physiol. 2018, 314, L6–L16. [Google Scholar] [CrossRef] [PubMed]
- Zhang, M.; Zhang, S. T Cells in Fibrosis and Fibrotic Diseases. Front. Immunol. 2020, 11, 1142. [Google Scholar] [CrossRef] [PubMed]
- Singh, R.; Alape, D.; de Lima, A.; Ascanio, J.; Majid, A.; Gangadharan, S.P. Regulatory T Cells in Respiratory Health and Diseases. Pulm. Med. 2019, 2019, 1907807. [Google Scholar] [CrossRef] [PubMed]
- Harrell, C.R.; Miloradovic, D.; Sadikot, R.; Fellabaum, C.; Markovic, B.S.; Miloradovic, D.; Acovic, A.; Djonov, V.; Arsenijevic, N.; Volarevic, V. Molecular and Cellular Mechanisms Responsible for Beneficial Effects of Mesenchymal Stem Cell-Derived Product "Exo-d-MAPPS" in Attenuation of Chronic Airway Inflammation. Anal. Cell Pathol. 2020, 2020, 3153891. [Google Scholar] [CrossRef]
- Harrell, C.R.; Fellabaum, C.; Markovic, B.S.; Arsenijevic, A.; Volarevic, V. Therapeutic potential of “Exosomes Derived Multiple Allogeneic Proteins Paracrine Signaling: Exosomes d-MAPPS” is based on the effects of exosomes, immunosuppressive and trophic factors. Serbian J. Exp. Clin. Res. 2019, 20, 189–197. [Google Scholar] [CrossRef]
- Li, R.; Kang, H.; Chen, S. From Basic Research to Clinical Practice: Considerations for Treatment Drugs for Silicosis. Int. J. Mol. Sci. 2023, 24, 8333. [Google Scholar] [CrossRef] [PubMed]
- Shi, J.; Li, F.; Luo, M.; Wei, J.; Liu, X. Distinct Roles of Wnt/β-Catenin Signaling in the Pathogenesis of Chronic Obstructive Pulmonary Disease and Idiopathic Pulmonary Fibrosis. Mediat. Inflamm. 2017, 2017, 3520581. [Google Scholar] [CrossRef]
- Martin-Medina, A.; Lehmann, M.; Burgy, O.; Hermann, S.; Baarsma, H.A.; Wagner, D.E.; De Santis, M.M.; Ciolek, F.; Hofer, T.P.; Frankenberger, M.; et al. Increased Extracellular Vesicles Mediate WNT5A Signaling in Idiopathic Pulmonary Fibrosis. Am. J. Respir. Crit. Care Med. 2018, 198, 1527–1538. [Google Scholar] [CrossRef]
- De Langhe, E.; Cailotto, F.; De Vooght, V.; Aznar-Lopez, C.; Vanoirbeek, J.A.; Luyten, F.P.; Lories, R.J. Enhanced endogenous bone morphogenetic protein signaling protects against bleomycin induced pulmonary fibrosis. Respir. Res. 2015, 16, 38. [Google Scholar] [CrossRef]
- Harrell, C.R.; Djonov, V.; Volarevic, V. Mesenchymal stem cell-derived microRNAs: Friends or foes of tumor cells? Histol. Histopathol. 2023, 38, 1373–1379. [Google Scholar] [PubMed]
- Xu, C.; Zhao, J.; Li, Q.; Hou, L.; Wang, Y.; Li, S.; Jiang, F.; Zhu, Z.; Tian, L. Exosomes derived from three-dimensional cultured human umbilical cord mesenchymal stem cells ameliorate pulmonary fibrosis in a mouse silicosis model. Stem Cell Res. Ther. 2020, 11, 503. [Google Scholar] [CrossRef] [PubMed]
- Xu, C.; Hou, L.; Zhao, J.; Wang, Y.; Jiang, F.; Jiang, Q.; Zhu, Z.; Tian, L. Exosomal let-7i-5p from three-dimensional cultured human umbilical cord mesenchymal stem cells inhibits fibroblast activation in silicosis through targeting TGFBR1. Ecotoxicol. Environ. Saf. 2022, 233, 113302. [Google Scholar] [CrossRef] [PubMed]
- Zhao, Y.; Du, L.; Sun, J.; Wang, X.; Cong, Z.; Chen, S.; Wang, F.; Li, Z. Exosomal miR-218 derived from mesenchymal stem cells inhibits endothelial-to-mesenchymal transition by epigenetically modulating of BMP2 in pulmonary fibrosis. Cell Biol. Toxicol. 2023, 39, 2919–2936. [Google Scholar] [CrossRef] [PubMed]
- Xie, L.; Long, X.; Mo, M.; Jiang, J.; Zhang, Q.; Long, M.; Li, M. Bone marrow mesenchymal stem cell-derived exosomes alleviate skin fibrosis in systemic sclerosis by inhibiting the IL-33/ST2 axis via the delivery of microRNA-214. Mol. Immunol. 2023, 157, 146–157. [Google Scholar] [CrossRef] [PubMed]
- Li, Y.; Shen, Z.; Jiang, X.; Wang, Y.; Yang, Z.; Mao, Y.; Wu, Z.; Li, G.; Chen, H. Mouse mesenchymal stem cell-derived exosomal miR-466f-3p reverses EMT process through inhibiting AKT/GSK3β pathway via c-MET in radiation-induced lung injury. J. Exp. Clin. Cancer Res. 2022, 41, 128. [Google Scholar] [CrossRef]
- Yun, E.; Kook, Y.; Yoo, K.H.; Kim, K.I.; Lee, M.S.; Kim, J.; Lee, A. Endothelial to Mesenchymal Transition in Pulmonary Vascular Diseases. Biomedicines 2020, 8, 639. [Google Scholar] [CrossRef] [PubMed]
- Mou, Y.; Wu, G.R.; Wang, Q.; Pan, T.; Zhang, L.; Xu, Y.; Xiong, W.; Zhou, Q.; Wang, Y. Macrophage-targeted delivery of siRNA to silence Mecp2 gene expression attenuates pulmonary fibrosis. Bioeng. Transl. Med. 2022, 7, e10280. [Google Scholar] [CrossRef]
- Li, D.; Guabiraba, R.; Besnard, A.G.; Komai-Koma, M.; Jabir, M.S.; Zhang, L.; Graham, G.J.; Kurowska-Stolarska, M.; Liew, F.Y.; McSharry, C.; et al. IL-33 promotes ST2-dependent lung fibrosis by the induction of alternatively activated macrophages and innate lymphoid cells in mice. J. Allergy Clin. Immunol. 2014, 134, 1422–1432.e11. [Google Scholar] [CrossRef]
- Wang, Y.; Li, S.; Zhao, J.; Li, Q.; Xu, C.; Wu, H.; Zhu, Z.; Tian, L. Snail-mediated partial epithelial mesenchymal transition augments the differentiation of local lung myofibroblast. Chemosphere 2021, 267, 128870. [Google Scholar] [CrossRef]
- Cai, L.; Wang, J.; Yi, X.; Yu, S.; Wang, C.; Zhang, L.; Zhang, X.; Cheng, L.; Ruan, W.; Dong, F.; et al. Nintedanib-loaded exosomes from adipose-derived stem cells inhibit pulmonary fibrosis induced by bleomycin. Pediatr. Res. 2024. [Google Scholar] [CrossRef] [PubMed]
- Rezabakhsh, A.; Sokullu, E.; Rahbarghazi, R. Applications, challenges and prospects of mesenchymal stem cell exosomes in regenerative medicine. Stem Cell Res. Ther. 2021, 12, 521. [Google Scholar] [CrossRef] [PubMed]
- Gowen, A.; Shahjin, F.; Chand, S.; Odegaard, K.E.; Yelamanchili, S.V. Mesenchymal Stem Cell-Derived Extracellular Vesicles: Challenges in Clinical Applications. Front. Cell Dev. Biol. 2020, 8, 149. [Google Scholar] [CrossRef] [PubMed]
- Xu, L.; Liu, Y.; Sun, Y.; Wang, B.; Xiong, Y.; Lin, W.; Wei, Q.; Wang, H.; He, W.; Wang, B.; et al. Tissue source determines the differentiation potentials of mesenchymal stem cells: A comparative study of human mesenchymal stem cells from bone marrow and adipose tissue. Stem Cell Res. Ther. 2017, 8, 275. [Google Scholar] [CrossRef] [PubMed]



{kind=link}
{kind=link}
| Target Cell(s) | Mechanism(s) of Action | Biological Effect(s) | Ref. No. |
|---|---|---|---|
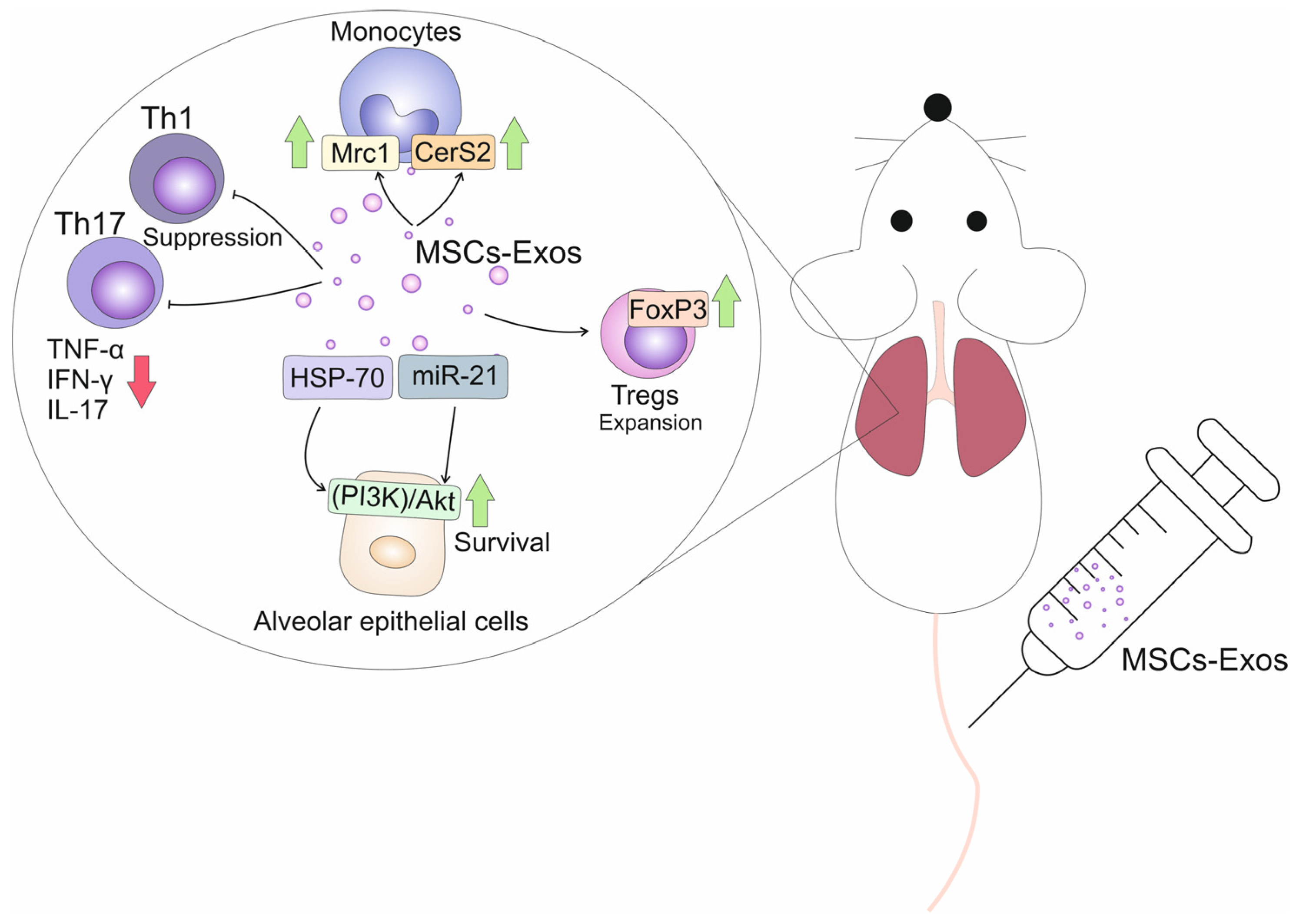
| Alveolar epithelial cells | HSP-70-dependent activation of PI3K/Akt- signaling pathway; | Improved survival and viability of alveolar epithelial cells | [33] |
| Monocytes/ macrophages | CCR2-based suppression of CCL2-driven signaling pathway; Reprogramming of “classical”, inflammatory M1 macrophages into anti-inflammatory, immunosuppressive M2 macrophages | Attenuated production of inflammatory and pro-fibrotic cytokines (TNF-α, IL-1β, IL-6, TGF-β); Increased synthesis of immunosuppressive IL-10 | [33] |
| Tregs; Th17 lymphocytes | IDO-dependent activation of GCN2 kinase and inhibition of Akt/mTOR2-driven signaling | Increased expansion of Tregs; Prevented differentiation of Tregs into Th17 cells; Suppressed production of IL-17 | [34] |
| Alveolar epithelial cells; lung-infiltrated immune cells | Inhibition of Wnt/β-catenin pathway | Increased expression of E-cadherin and cytokeratin 19 and reduced expression of α-SMA; Attenuated production of TGF-β | [35] |
| Fibroblasts | Down-regulated gene expression of β-catenin, cyclin D1 and TGF-β1; Activation of Wnt5a/BMP2-signaling pathway | Reduced collagen production and extracellular matrix deposition in inflamed lungs; | [36] |
| MSC-Derived miRNA | Animal Model | Target Cell(s) | Mechanism(s) of Action | Biological Effect(s) | Ref. No. |
|---|---|---|---|---|---|
| MiR-21 | Mice model of bleomycin-induced pulmonary fibrosis | Alveolar epithelial cells; Monocytes/ macrophages | Reduced synthesis of pro-apoptotic proteins; Increased synthesis of immunoregulatory proteins (Mrc1 and CerS2) | Improved viability of alveolar epithelial cells; Reduced production of TNF-α and IL-1β; Increased synthesis of IL-10 | [33] |
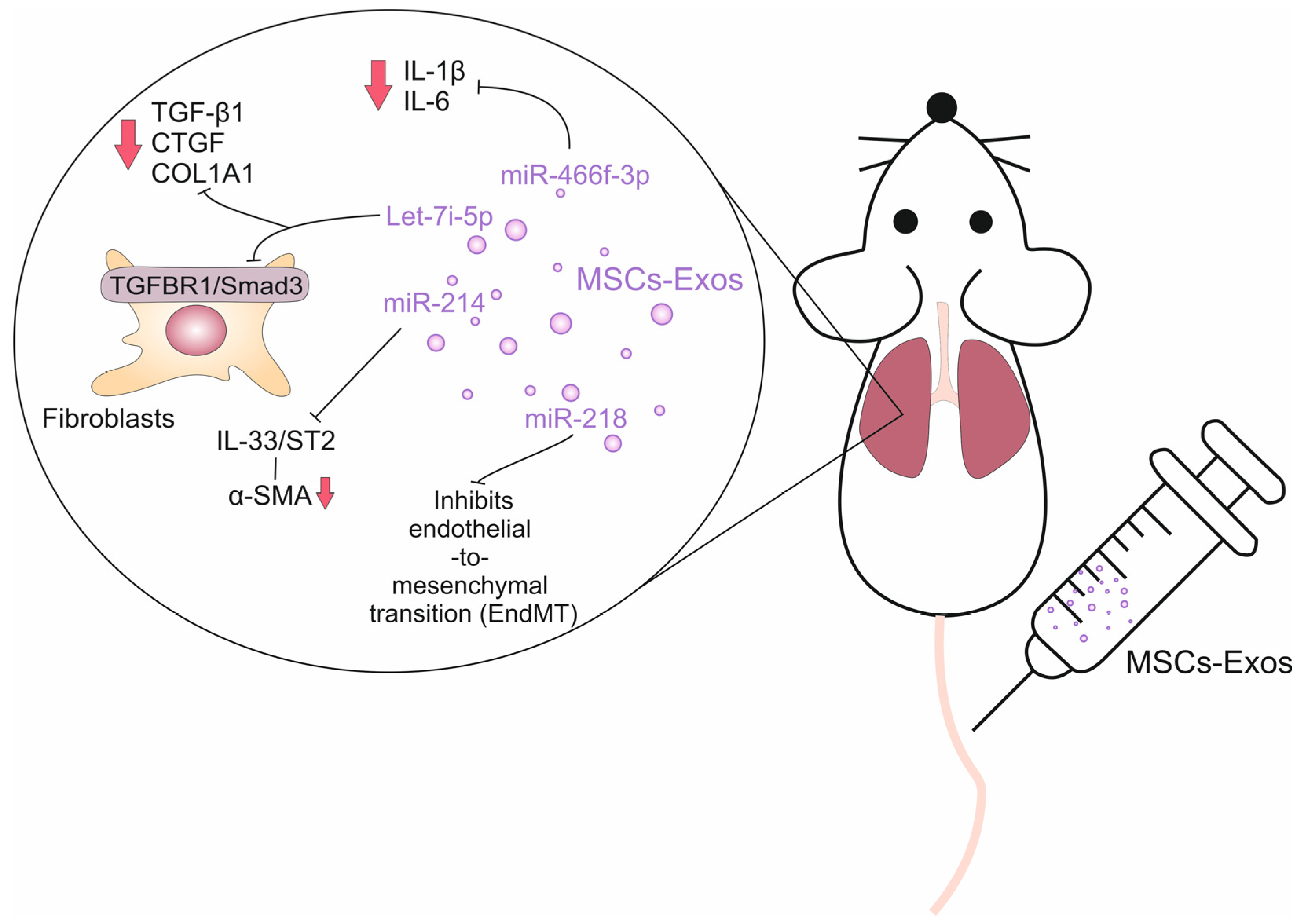
| Let-7i-5p | Mice model of silica-induced lung fibrosis | Lung fibroblasts; Alveolar epithelial cells; Lung-infiltrated monocytes | Inhibition of TGFBR1/Smad3 signaling pathway; Inhibition of epithelial-to-mesenchymal transition; Suppressed production of TNF-α and IL-1β | Reduced collagen production and extracellular matrix deposition in inflamed lungs; Preserved integrity of the epithelial barrier and reduced influx of circulating immune cells in the fibrotic lungs | [54] |
| MiR-218 | Mice model of bleomycin-induced pulmonary fibrosis | Endothelial cells | Increased BMP2 expression; Inhibition of endothelial-to-mesenchymal transition | Reduced extravasation of immune cells from the blood vessels n inflamed lungs | [55] |
| MiR-214 | Mice model of bleomycin-induced pulmonary fibrosis | Lung fibroblasts; Lung-infiltrated immune cells | Inhibition of IL-33/ST2 signaling pathway | Reduced collagen and α-SMA production; Attenuated production of pro-fibrotic TGF-β | [56] |
| miR-466f-3p | Radiation-induced lung fibrosis | Lung fibroblasts; Lung-infiltrated immune cells; Alveolar epithelial cells | Suppressed activation of AKT/GSK3β signaling pathway; Reduced synthesis of SNAIL protein | Reduced collagen production; Attenuated production of TNF-α and IL-1β; Inhibition of epithelial-to-mesenchymal transition | [57] |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Harrell, C.R.; Djonov, V.; Volarevic, A.; Arsenijevic, A.; Volarevic, V. Molecular Mechanisms Responsible for the Therapeutic Potential of Mesenchymal Stem Cell-Derived Exosomes in the Treatment of Lung Fibrosis. Int. J. Mol. Sci. 2024, 25, 4378. https://doi.org/10.3390/ijms25084378
Harrell CR, Djonov V, Volarevic A, Arsenijevic A, Volarevic V. Molecular Mechanisms Responsible for the Therapeutic Potential of Mesenchymal Stem Cell-Derived Exosomes in the Treatment of Lung Fibrosis. International Journal of Molecular Sciences. 2024; 25(8):4378. https://doi.org/10.3390/ijms25084378
Chicago/Turabian StyleHarrell, Carl Randall, Valentin Djonov, Ana Volarevic, Aleksandar Arsenijevic, and Vladislav Volarevic. 2024. "Molecular Mechanisms Responsible for the Therapeutic Potential of Mesenchymal Stem Cell-Derived Exosomes in the Treatment of Lung Fibrosis" International Journal of Molecular Sciences 25, no. 8: 4378. https://doi.org/10.3390/ijms25084378