

The Development of Toxoplasma gondii Recombinant Trivalent Chimeric Proteins as an Alternative to Toxoplasma Lysate Antigen (TLA) in Enzyme-Linked Immunosorbent Assay (ELISA) for the Detection of Immunoglobulin G (IgG) in Small Ruminants

 ,
,  , , ,
, , ,
Abstract
1. Introduction
2. Results
2.1. Preliminary Evaluation of Single Recombinant Proteins Using IgG ELISA
2.2. Plasmid Construction, Expression, and Purification of the Recombinant Chimeric Proteins
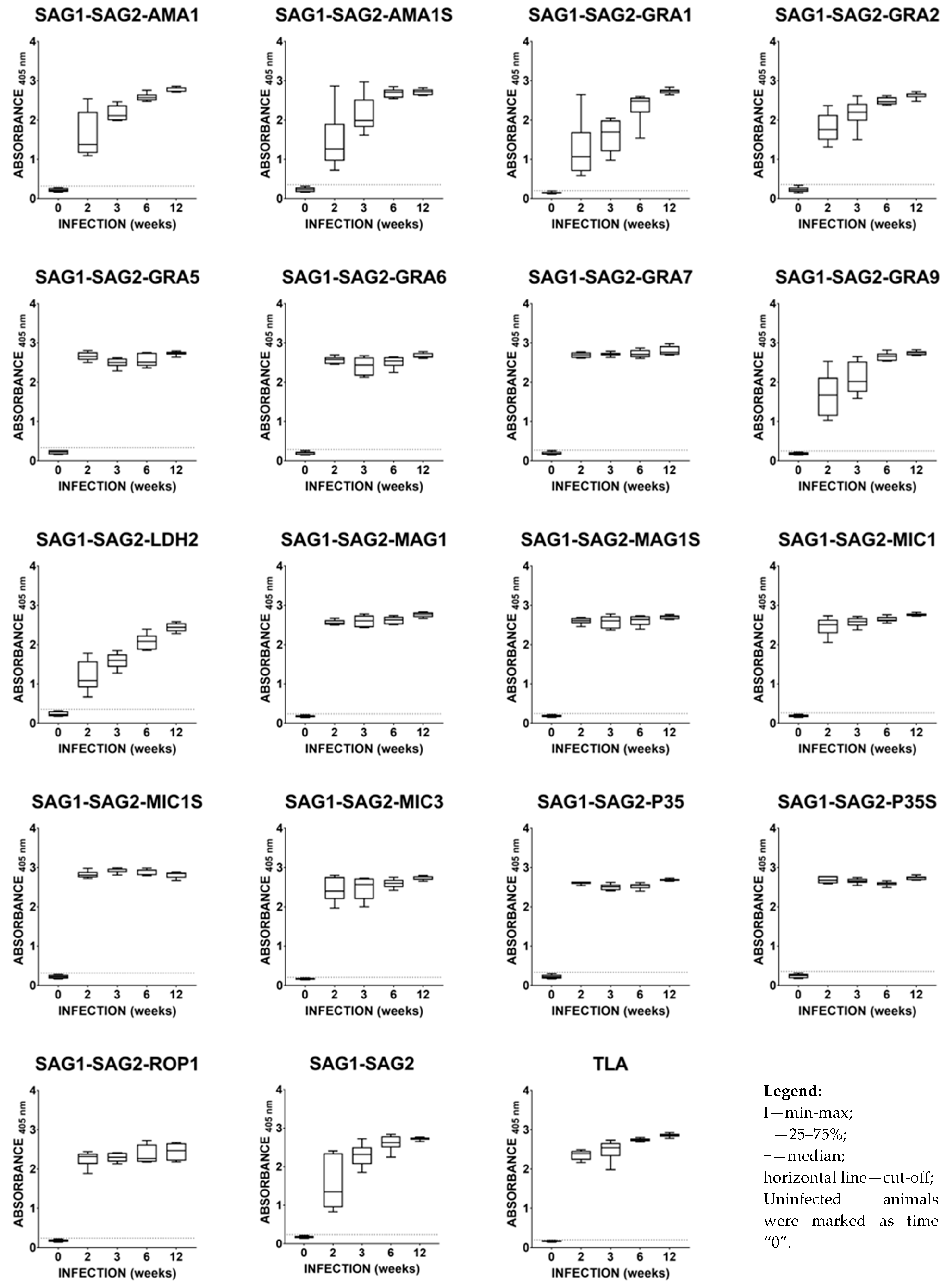
2.3. Immunoreactivities of Mouse Anti-T. gondii IgG Antibodies in ELISA
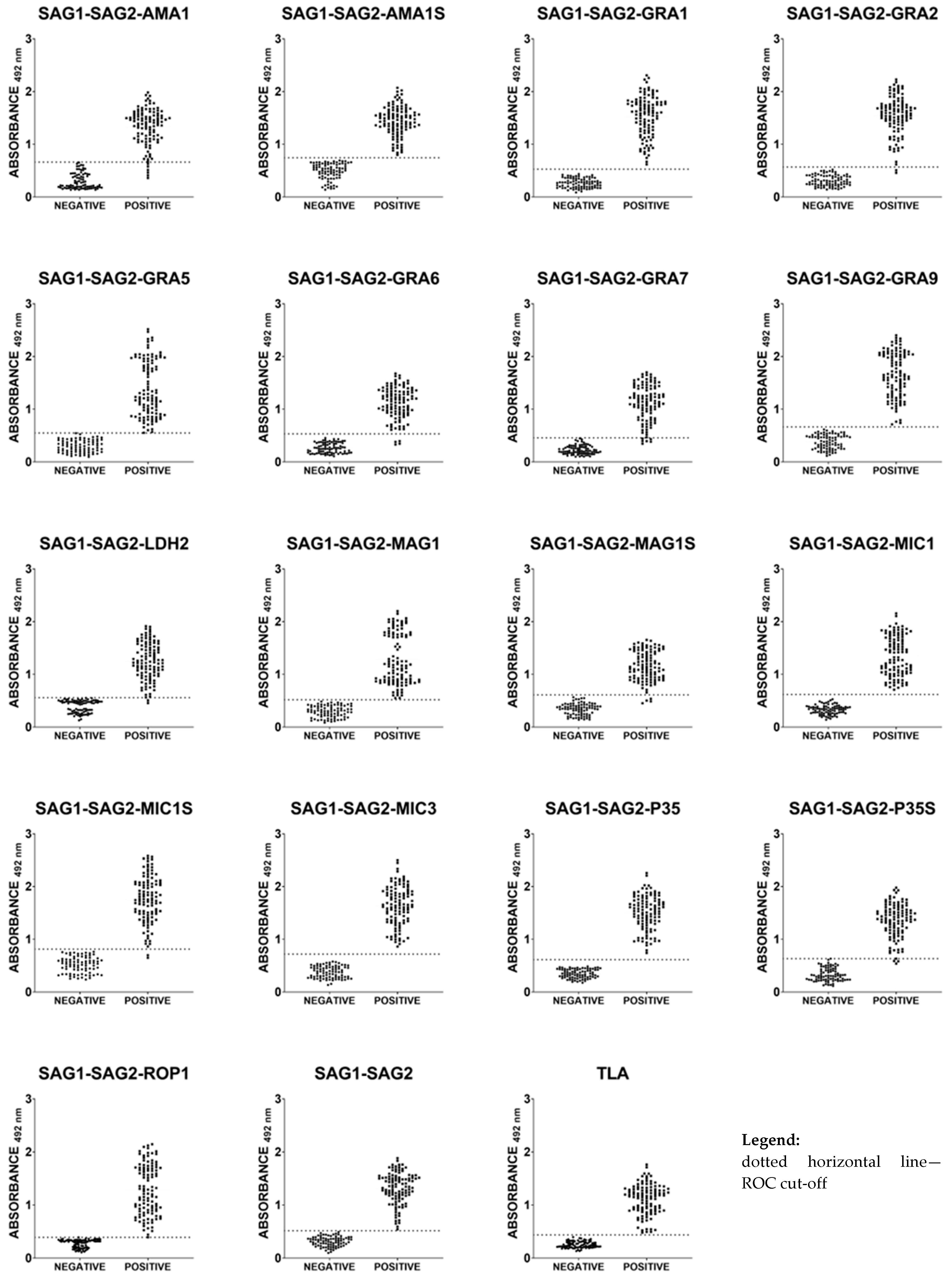
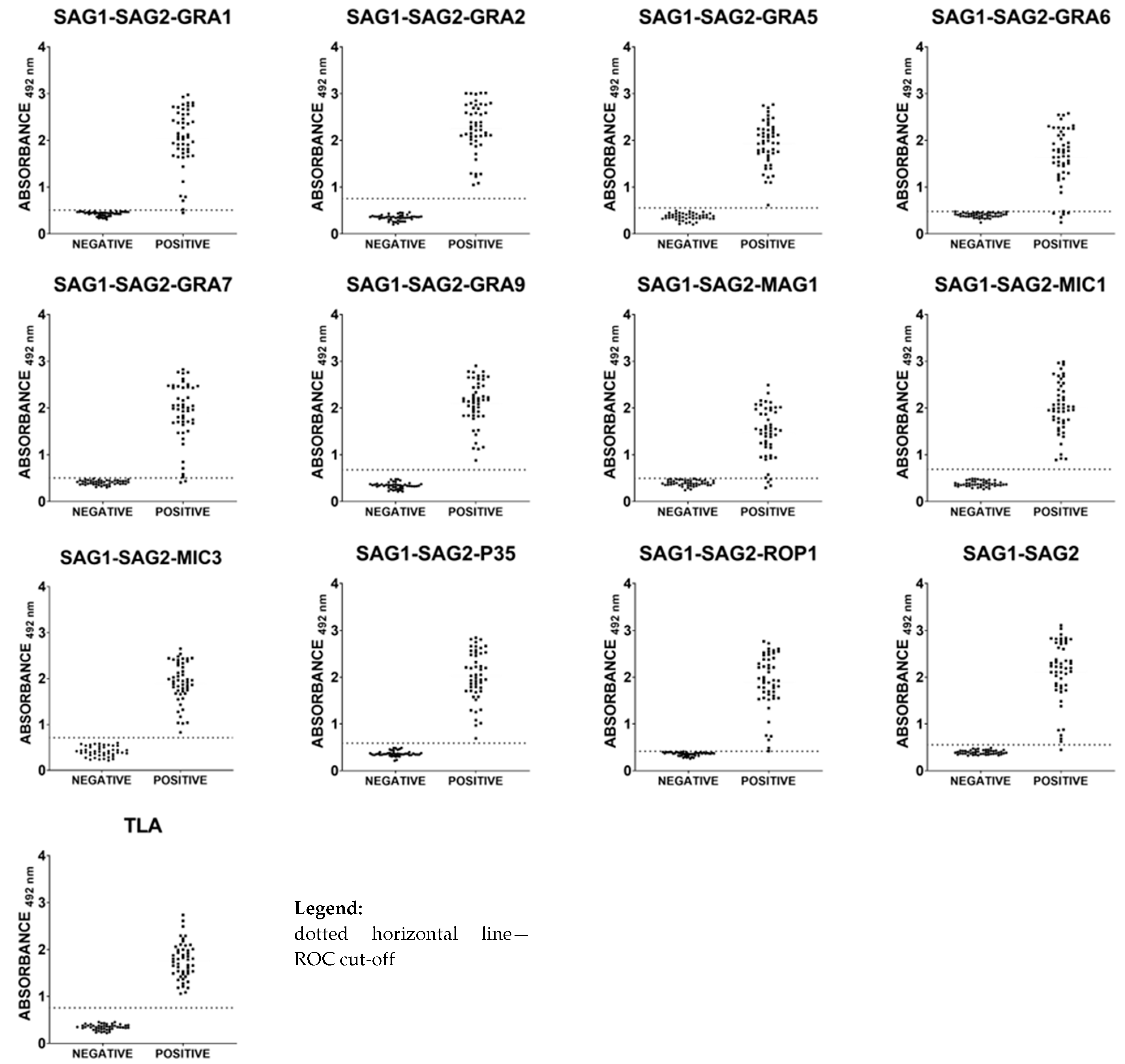
2.4. Immunoreactivities of Ovine Anti-T. gondii IgG Antibodies in ELISA with Recombinant Chimeric Proteins and TLA
2.5. Immunoreactivities of Caprine Anti-T. gondii IgG Antibodies in ELISA with Recombinant Chimeric Proteins and TLA
2.6. Total Immunoreactivity of Small Ruminants’ Anti-T. gondii IgG Antibodies in ELISA with Recombinant Chimeric Proteins and TLA
3. Discussion
4. Materials and Methods
4.1. T. gondii Single Recombinant Proteins
4.2. Ovine and Caprine Serum Samples
4.3. IgG ELISAs with Single Recombinant Proteins, Recombinant Chimeric Proteins, and TLA
- (a)
- single recombinant proteins at a final concentration of 5, 2.5, and 1 μg/mL for each recombinant protein in a coating buffer (0.05 M carbonate buffer, pH 9.6) for the preliminary test;
- (b)
- recombinant chimeric proteins at a final concentration of 2.5 μg/mL for each recombinant protein and 1 μg/mL for TLA in a coating buffer (0.05 M carbonate buffer, pH 9.6) for the testing of newly produced recombinant chimeric proteins.
4.4. Ethical Statements
4.5. Parasite
4.6. Animals
4.7. Mice Infection
4.8. Preparation of T. gondii Native Lysate Antigen (TLA)
4.9. IgG ELISA with Mouse Serum Samples
4.10. T. gondii RNA Isolation and cDNA Synthesis
4.11. Construction of the Recombinant Plasmids
4.12. Production and Purification of Recombinant Chimeric Proteins
4.13. Statistical Analysis
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Almeria, S.; Dubey, J.P. Foodborne Transmission of Toxoplasma gondii Infection in the Last Decade. An Overview. Res. Vet. Sci. 2021, 135, 371–385. [Google Scholar] [CrossRef] [PubMed]
- Ferra, B.T.; Holec-Gąsior, L.; Gatkowska, J.; Dziadek, B.; Dzitko, K.; Grąźlewska, W.; Lautenbach, D. The First Study on the Usefulness of Recombinant Tetravalent Chimeric Proteins Containing Fragments of SAG2, GRA1, ROP1 and AMA1 Antigens in the Detection of Specific Anti-Toxoplasma gondii Antibodies in Mouse and Human Sera. PLoS ONE 2019, 14, e0217866. [Google Scholar] [CrossRef]
- Ferra, B.; Holec-Gąsior, L.; Kur, J. A New Toxoplasma gondii Chimeric Antigen Containing Fragments of SAG2, GRA1, and ROP1 Proteins—Impact of Immunodominant Sequences Size on Its Diagnostic Usefulness. Parasitol. Res. 2015, 114, 3291–3299. [Google Scholar] [CrossRef] [PubMed][Green Version]
- Innes, E.A.; Bartley, P.M.; Buxton, D.; Katzer, F. Ovine Toxoplasmosis. Parasitology 2009, 136, 1887–1894. [Google Scholar] [CrossRef] [PubMed]
- Innes, E.A.; Vermeulen, A.N. Vaccination as a Control Strategy against the Coccidial Parasites Eimeria, Toxoplasma and Neospora. Parasitology 2006, 133, S145–S168. [Google Scholar] [CrossRef] [PubMed]
- Sam-Yellowe, T.Y. Rhoptry Organelles of the Apicomplexa: Their Role in Host Cell Invasion and Intracellular Survival. Parasitol. Today 1996, 12, 308–316. [Google Scholar] [CrossRef] [PubMed]
- Carruthers, V.B.; Sibley, L.D. Sequential Protein Secretion Front Three Distinct Organelles of Toxoplasma gondii Accompanies Invasion of Human Fibroblasts. Eur. J. Cell Biol. 1997, 73, 114–123. [Google Scholar] [PubMed]
- Carruthers, V.B.; Giddings, O.K.; Sibley, L.D. Secretion of Micronemal Proteins Is Associated with Toxoplasma Invasion of Host Cells. Cell Microbiol. 1999, 1, 225–235. [Google Scholar] [CrossRef] [PubMed]
- Lekutis, C.; Ferguson, D.J.P.; Grigg, M.E.; Camps, M.; Boothroyd, J.C. Surface Antigens of Toxoplasma gondii: Variations on a Theme. Int. J. Parasitol. 2001, 31, 1285–1292. [Google Scholar] [CrossRef]
- Joiner, K.A.; Roos, D.S. Secretory Traffic in the Eukaryotic Parasite Toxoplasma gondii. J. Cell Biol. 2002, 157, 557–563. [Google Scholar] [CrossRef]
- Mercier, C.; Dubremetz, J.F.; Rauscher, B.; Lecordier, L.; Sibley, L.D.; Cesbron-Delauw, M.F. Biogenesis of Nanotubular Network in Toxoplasma Parasitophorous Vacuole Induced by Parasite Proteins. Mol. Biol. Cell 2002, 13, 2397–2409. [Google Scholar] [CrossRef]
- Mercier, C.; Adjogble, K.D.Z.; Däubener, W.; Delauw, M.-F.-C. Dense Granules: Are They Key Organelles to Help Understand the Parasitophorous Vacuole of All Apicomplexa Parasites? Int. J. Parasitol. 2005, 35, 829–849. [Google Scholar] [CrossRef]
- Holec-Gasior, L. Toxoplasma gondii Recombinant Antigens as Tools for Serodiagnosis of Human Toxoplasmosis: Current Status of Studies. Clin. Vaccine Immunol. 2013, 20, 1343–1351. [Google Scholar] [CrossRef]
- Ferra, B.; Holec-Gasior, L.; Graźlewska, W. Toxoplasma gondii Recombinant Antigens in the Serodiagnosis of Toxoplasmosis in Domestic and Farm Animals. Animals 2020, 10, 1245. [Google Scholar] [CrossRef]
- Ferra, B.; Holec-Gasior, L.; Kur, J. Serodiagnosis of Toxoplasma gondii Infection in Farm Animals (Horses, Swine, and Sheep) by Enzyme-Linked Immunosorbent Assay Using Chimeric Antigens. Parasitol. Int. 2015, 64, 288–294. [Google Scholar] [CrossRef]
- Holec-Gąsior, L.; Ferra, B.; Grąźlewska, W. Toxoplasma gondii Tetravalent Chimeric Proteins as Novel Antigens for Detection of Specific Immunoglobulin G in Sera of Small Ruminants. Animals 2019, 9, 1146. [Google Scholar] [CrossRef]
- Holec-Gasior, L.; Ferra, B.; Hiszczyńska-Sawicka, E.; Kur, J. The Optimal Mixture of Toxoplasma gondii Recombinant Antigens (GRA1, P22, ROP1) for Diagnosis of Ovine Toxoplasmosis. Vet. Parasitol. 2014, 206, 146–152. [Google Scholar] [CrossRef]
- Kijlstra, A.; Jongert, E. Control of the Risk of Human Toxoplasmosis Transmitted by Meat. Int. J. Parasitol. 2008, 38, 1359–1370. [Google Scholar] [CrossRef]
- Kijlstra, A.; Jongert, E. Toxoplasma-Safe Meat: Close to Reality? Trends Parasitol. 2009, 25, 18–22. [Google Scholar] [CrossRef]
- Olivier, A.; Herbert, B.; Sava, B.; Pierre, C.; John, D.C.; Aline, D.K.; John, G.; Arie, H.; James, H.; Gunter, K.; et al. Surveillance and Monitoring of Toxoplasma in Humans, Food and Animals—Scientific Opinion of the Panel on Biological Hazards. EFSA J. 2007, 583, 1–64. [Google Scholar] [CrossRef]
- Jung, C.; Lee, C.Y.F.; Grigg, M.E. The SRS Superfamily of Toxoplasma Surface Proteins. Int. J. Parasitol. 2004, 34, 285–296. [Google Scholar] [CrossRef]
- Manger, I.D.; Hehl, A.B.; Boothroyd, J.C. The Surface of Toxoplasma Tachyzoites Is Dominated by a Family of Glycosylphosphatidylinositol-Anchored Antigens Related to SAG1. Infect. Immun. 1998, 66, 2237–2244. [Google Scholar] [CrossRef] [PubMed]
- Lekutis, C.; Ferguson, D.J.P.; Boothroyd, J.C. Toxoplasma gondii: Identification of a Developmentally Regulated Family of Genes Related to SAG2. Exp. Parasitol. 2000, 96, 89–96. [Google Scholar] [CrossRef] [PubMed]
- Bülow, R.; Boothroyd, J.C. Protection of Mice from Fatal Toxoplasma gondii Infection by Immunization with P30 Antigen in Liposomes. J. Immunol. 1991, 147, 3496–3500. [Google Scholar] [CrossRef] [PubMed]
- Nagel, S.D.; Boothroyd, J.C. The Major Surface Antigen, P30, of Toxoplasma gondii Is Anchored by a Glycolipid. J. Biol. Chem. 1989, 264, 5569–5574. [Google Scholar] [CrossRef] [PubMed]
- Singh, H.; Tewari, A.K.; Mishra, A.K.; Maharana, B.; Sudan, V.; Raina, O.K.; Rao, J.R. Detection of Antibodies to Toxoplasma gondii in Domesticated Ruminants by Recombinant Truncated SAG2 Enzyme-Linked Immunosorbent Assay. Trop. Anim. Health Prod. 2014, 47, 171–178. [Google Scholar] [CrossRef] [PubMed]
- Zhou, M.; Cao, S.; Sevinc, F.; Sevinc, M.; Ceylan, O.; Liu, M.; Wang, G.; Moumouni, P.F.A.; Jirapattharasate, C.; Suzuki, H.; et al. Enzyme-Linked Immunosorbent Assays Using Recombinant TgSAG2 and NcSAG1 to Detect Toxoplasma gondii and Neospora caninum-Specific Antibodies in Domestic Animals in Turkey. J. Vet. Med. Sci. 2016, 78, 1877–1881. [Google Scholar] [CrossRef] [PubMed]
- Fereig, R.M.; Mahmoud, H.Y.A.H.; Mohamed, S.G.A.; AbouLaila, M.R.; Abdel-Wahab, A.; Osman, S.A.; Zidan, S.A.; El-Khodary, S.A.; Mohamed, A.E.A.; Nishikawa, Y. Seroprevalence and Epidemiology of Toxoplasma gondii in Farm Animals in Different Regions of Egypt. Vet. Parasitol. Reg. Stud. Rep. 2016, 3–4, 1–6. [Google Scholar] [CrossRef] [PubMed]
- Tenter, A.M.; Vietmeyer, C.; Johnson, A.M. Development of ELISAs Based on Recombinant Antigens for the Detection of Toxoplasma gondii-Specific Antibodies in Sheep and Cats. Vet. Parasitol. 1992, 43, 189–201. [Google Scholar] [CrossRef]
- Sager, H.; Gloor, M.; Tenter, A.; Maley, S.; Hässig, M.; Gottstein, B. Immunodiagnosis of Primary Toxoplasma gondii Infection in Sheep by the Use of a P30 IgG Avidity ELISA. Parasitol. Res. 2003, 91, 171–174. [Google Scholar] [CrossRef]
- Tumurjav, B.; Nishikawa, Y.; Xuen, X. Seroprevalence of Ovine Toxoplasmosis in Mongolian Samples Using an ELISA with Recombinant Toxoplasma gondii Matrix Antigen 1. Mong. J. Agric. Sci. 2015, 15, 33–37. [Google Scholar] [CrossRef]
- Tumurjav, B.; Terkawi, M.A.; Zhang, H.; Zhang, G.; Jia, H.; Goo, Y.-K.; Yamagishi, J.; Nishikawa, Y.; Igarashi, I.; Sugimoto, C.; et al. Serodiagnosis of Ovine Toxoplasmosis in Mongolia by an Enzyme-Linked Immunosorbent Assay with Recombinant Toxoplasma gondii Matrix Antigen 1. Jpn. J. Vet. Res. 2010, 58, 111–119. [Google Scholar] [CrossRef] [PubMed]
- Kandemir, Ç.; Can, H.; Karakavuk, M.; Döşkaya, M.; Erkunt Alak, S.; Gürüz, A.Y.; Ün, C.; Taşkın, T.; Değirmenci Döşkaya, A. An In House ELISA Using Recombinant GRA1 and BAG1 Proteins as Antigen Successfully Detected Toxoplasma gondii Infection in a Flock of Sheep Suffering from Abortions. Res. Vet. Sci. 2023, 165, 105054. [Google Scholar] [CrossRef]
- Bachan, M.; Deb, A.R.; Maharana, B.R.; Sudhakar, N.R.; Sudan, V.; Saravanan, B.C.; Tewari, A.K. High Seroprevalence of Toxoplasma gondii in Goats in Jharkhand State of India. Vet. Parasitol. Reg. Stud. Rep. 2018, 12, 61–68. [Google Scholar] [CrossRef]
- Udonsom, R.; Supanta, J.; Tanglakmankhong, O.; Ngoenphisutsin, K.; Nishikawa, Y.; Fereig, R.M.; Jirapattharasate, C. Toxoplasma gondii and Neospora caninum Prevalence and Risk Factors on Goat Farms in Kanchanaburi Province, Thailand. Vet. Integr. Sci. 2021, 19, 65–74. [Google Scholar] [CrossRef]
- Jirapattharasate, C.; Udonsom, R.; Prachasuphap, A.; Jongpitisub, K.; Dhepakson, P. Development and Evaluation of Recombinant GRA8 Protein for the Serodiagnosis of Toxoplasma gondii Infection in Goats. BMC Vet. Res. 2021, 17, 1–9. [Google Scholar] [CrossRef] [PubMed]
- Velmurugan, G.V.; Tewari, A.K.; Rao, J.R.; Baidya, S.; Kumar, M.U.; Mishra, A.K. High-Level Expression of SAG1 and GRA7 Gene of Toxoplasma gondii (Izatnagar Isolate) and Their Application in Serodiagnosis of Goat Toxoplasmosis. Vet. Parasitol. 2008, 154, 185–192. [Google Scholar] [CrossRef]
- Cook, A.J.C.; Gilbert, R.E.; Buffolano, W.; Zufferey, J.; Petersen, E.; Jenum, P.A.; Foulon, W.; Semprini, A.E.; Dunn, D.T. Sources of Toxoplasma Infection in Pregnant Women: European Multicentre Case-Control Study. Br. Med. J. 2000, 321, 142–147. [Google Scholar] [CrossRef]
- Dziadek, B.; Gatkowska, J.; Brzostek, A.; Dziadek, J.; Dzitko, K.; Dlugonska, H. Toxoplasma gondii: The Immunogenic and Protective Efficacy of Recombinant ROP2 and ROP4 Rhoptry Proteins in Murine Experimental Toxoplasmosis. Exp. Parasitol. 2009, 123, 81–89. [Google Scholar] [CrossRef] [PubMed]
- Gatkowska, J.; Hiszczynska-Sawicka, E.; Kur, J.; Holec, L.; Dlugonska, H. Toxoplasma gondii: An Evaluation of Diagnostic Value of Recombinant Antigens in a Murine Model. Exp. Parasitol. 2006, 114, 220–227. [Google Scholar] [CrossRef]
- Ferra, B.; Holec-Gąsior, L.; Gatkowska, J.; Dziadek, B.; Dzitko, K. Toxoplasma gondii Recombinant Antigen AMA1: Diagnostic Utility of Protein Fragments for the Detection of IgG and IgM Antibodies. Pathogens 2020, 9, 43. [Google Scholar] [CrossRef] [PubMed]
- Hiszczyńska-Sawicka, E.; Brillowska-Dąbrowska, A.; Dąbrowski, S.; Pietkiewicz, H.; Myjak, P.; Kur, J. High Yield Expression and Single-Step Purification of Toxoplasma Gondii SAG1, GRA1, and GRA7 Antigens in Escherichia coli. Protein Expr. Purif. 2003, 27, 150–157. [Google Scholar] [CrossRef] [PubMed]
- Holec-Gasior, L.; Kur, J.; Hiszczyńska-Sawicka, E. GRA2 and ROP1 Recombinant Antigens as Potential Markers for Detection of Toxoplasma gondii-Specific Immunoglobulin G in Humans with Acute Toxoplasmosis. Clin. Vaccine Immunol. 2009, 16, 510–514. [Google Scholar] [CrossRef] [PubMed]
- Holec-Gasior, L.; Kur, J. Toxoplasma gondii: Recombinant GRA5 Antigen for Detection of Immunoglobulin G Antibodies Using Enzyme-Linked Immunosorbent Assay. Exp. Parasitol. 2010, 124, 272–278. [Google Scholar] [CrossRef]
- Hiszczyńska-Sawicka, E.; Kur, J.; Pietkiewicz, H.; Holec, L.; Gąsior, A.; Myjak, P. Efficient Production of the Toxoplasma gondii GRA6, P35 and SAG2 Recombinant Antigens and Their Applications in the Serodiagnosis of Toxoplasmosis. Acta Parasitol. 2005, 50, 249–254. [Google Scholar]
- Holec, L.; Hiszczyńska-Sawicka, E.; Gasior, A.; Brillowska-Dabrowska, A.; Kur, J. Use of MAG1 Recombinant Antigen for Diagnosis of Toxoplasma gondii Infection in Humans. Clin. Vaccine Immunol. 2007, 14, 220–225. [Google Scholar] [CrossRef]
- Holec, L.; Gąsior, A.; Brillowska-Dąbrowska, A.; Kur, J. Toxoplasma gondii: Enzyme-Linked Immunosorbent Assay Using Different Fragments of Recombinant Microneme Protein 1 (MIC1) for Detection of Immunoglobulin G Antibodies. Exp. Parasitol. 2008, 119, 1–6. [Google Scholar] [CrossRef]




{kind=link}
{kind=link}
{kind=link}
| Recombinant Protein | Antibodies Dilution | Animal Species | |||||
|---|---|---|---|---|---|---|---|
| Ovine | Caprine | ||||||
| Mean ± SD S+ | Mean ± SD S− | p Value | Mean ± SD S+ | Mean ± SD S− | p Value | ||
| AMA1 | 1:16,000 | 1.360 ± 0.271 | 0.506 ± 0.047 | 0.0058 ** | 2.339 ± 0.414 | 0.853 ± 0.057 | 0.0035 ** |
| 1:32,000 | 0.814 ± 0.175 | 0.322 ± 0.025 | 0.0086 ** | 1.541 ± 0.310 | 0.522 ± 0.025 | 0.0047 ** | |
| AMA1C | 1:16,000 | 1.124 ± 0.022 | 0.597 ± 0.153 | 0.0041 ** | 1.645 ± 0.232 | 0.841 ± 0.020 | 0.0039 ** |
| 1:32,000 | 0.559 ± 0.062 | 0.311 ± 0.092 | 0.0181 * | 0.997 ± 0.143 | 0.545 ± 0.034 | 0.0060 ** | |
| AMA1N | 1:16,000 | 1.087 ± 0.133 | 0.426 ± 0.064 | 0.0015 ** | 2.027 ± 0.388 | 0.723 ± 0.090 | 0.0048 ** |
| 1:32,000 | 0.583 ± 0.100 | 0.253 ± 0.037 | 0.0058 ** | 1.248 ± 0.259 | 0.410 ± 0.053 | 0.0053 ** | |
| BAG1 | 1:16,000 | 1.289 ± 0.314 | 0.680 ± 0.126 | 0.0356 * | 1.857 ± 0.233 | 1.354 ± 0.083 | 0.0245 * |
| 1:32,000 | 0.766 ± 0.206 | 0.395 ± 0.067 | 0.0411 * | 1.088 ± 0.108 | 0.723 ± 0.021 | 0.0045 ** | |
| BSR4 | 1:16,000 | 0.549 ± 0.262 | 0.223 ± 0.077 | 0.1076 | 1.080 ± 0.129 | 0.766 ± 0.049 | 0.0168 * |
| 1:32,000 | 0.312 ± 0.149 | 0.151 ± 0.044 | 0.1468 | 0.629 ± 0.064 | 0.464 ± 0.039 | 0.0186 * | |
| GRA1 | 1:16,000 | 1.049 ± 0.144 | 0.334 ± 0.037 | 0.0011 ** | 2.543 ± 0.222 | 0.599 ± 0.083 | 0.0001 *** |
| 1:32,000 | 0.606 ± 0.078 | 0.233 ± 0.045 | 0.0020 ** | 1.651 ± 0.163 | 0.350 ± 0.029 | 0.0002 *** | |
| GRA2ex2 | 1:16,000 | 1.411 ± 0.057 | 0.595 ± 0.138 | 0.0007 *** | 2.456 ± 0.567 | 0.929 ± 0.246 | 0.0128 * |
| 1:32,000 | 0.880 ± 0.056 | 0.357 ± 0.061 | 0.0004 *** | 1.558 ± 0.354 | 0.495 ± 0.133 | 0.0082 ** | |
| GRA4 | 1:16,000 | 1.471 ± 0.381 | 0.850 ± 0.026 | 0.0482 * | 2.667 ± 0.694 | 1.793 ± 0.221 | 0.1060 |
| 1:32,000 | 0.846 ± 0.278 | 0.505 ± 0.027 | 0.1020 | 1.805 ± 0.671 | 1.042 ± 0.150 | 0.1265 | |
| GRA5 | 1:16,000 | 1.550 ± 0.147 | 0.672 ± 0.018 | 0.0005 *** | 2.291 ± 0.445 | 0.804 ± 0.063 | 0.0046 ** |
| 1:32,000 | 0.905 ± 0.083 | 0.414 ± 0.013 | 0.0005 *** | 1.394 ± 0.323 | 0.539 ± 0.093 | 0.0117 * | |
| GRA6 | 1:16,000 | 1.442 ± 0.350 | 0.498 ± 0.030 | 0.0096 ** | 2.218 ± 0.076 | 0.610 ± 0.061 | <0.0001 **** |
| 1:32,000 | 0.871 ± 0.224 | 0.323 ± 0.021 | 0.0135 * | 1.381 ± 0.013 | 0.432 ± 0.042 | <0.0001 **** | |
| GRA7 | 1:16,000 | 1.298 ± 0.268 | 0.382 ± 0.033 | 0.0042 ** | 2.370 ± 0.450 | 1.032 ± 0.034 | 0.0068 ** |
| 1:32,000 | 0.845 ± 0.245 | 0.256 ± 0.021 | 0.0142 * | 1.473 ± 0.308 | 0.615 ± 0.034 | 0.0087 ** | |
| GRA9 | 1:16,000 | 1.255 ± 0.180 | 0.380 ± 0.039 | 0.0012 ** | 2.667 ± 0.694 | 1.019 ± 0.127 | 0.0155 * |
| 1:32,000 | 0.727 ± 0.114 | 0.250 ± 0.053 | 0.0028 ** | 1.805 ± 0.671 | 0.560 ± 0.029 | 0.0325 * | |
| LDH1 | 1:16 000 | 1.208 ± 0.674 | 0.324 ± 0.062 | 0.0862 | 1.569 ± 0.311 | 0.917 ± 0.195 | 0.0370 * |
| 1:32,000 | 0.588 ± 0.309 | 0.216 ± 0.046 | 0.1082 | 1.040 ± 0.345 | 0.571 ± 0.040 | 0.0795 | |
| LDH2 | 1:16,000 | 1.578 ± 0.291 | 0.565 ± 0.049 | 0.0040 ** | 1.847 ± 0.144 | 0.529 ± 0.061 | 0.0001 *** |
| 1:32,000 | 0.927 ± 0.173 | 0.353 ± 0.026 | 0.0047 ** | 1.168 ± 0.109 | 0.314 ± 0.020 | 0.0002 *** | |
| MAG1 | 1:16,000 | 1.152 ± 0.304 | 0.449 ± 0.024 | 0.0162 * | 1.626 ± 0.594 | 0.644 ± 0.083 | 0.0470 * |
| 1:32,000 | 0.666 ± 0.192 | 0.277 ± 0.019 | 0.0251 * | 0.954 ± 0.389 | 0.423 ± 0.048 | 0.0785 | |
| MIC1 | 1:16,000 | 1.233 ± 0.216 | 0.544 ± 0.077 | 0.0065 ** | 2.628 ± 0.445 | 0.885 ± 0.064 | 0.0026 ** |
| 1:32,000 | 0.741 ± 0.073 | 0.335 ± 0.006 | 0.0007 *** | 1.646 ± 0.365 | 0.475 ± 0.026 | 0.0052 ** | |
| MIC1ex2 | 1:16,000 | 1.486 ± 0.042 | 0.848 ± 0.069 | 0.0002 *** | 2.557 ± 0.418 | 0.957 ± 0.074 | 0.0028 ** |
| 1:32,000 | 0.865 ± 0.040 | 0.465 ± 0.015 | <0.0001 **** | 1.565 ± 0.361 | 0.496 ± 0.084 | 0.0075 ** | |
| MIC1ex3.4 | 1:16,000 | 1.105 ± 0.204 | 0.473 ± 0.138 | 0.0114 * | 2.186 ± 0.497 | 1.094 ± 0.117 | 0.0208 * |
| 1:32,000 | 0.667 ± 0.137 | 0.303 ± 0.094 | 0.0192 * | 1.351 ± 0.369 | 0.640 ± 0.092 | 0.0317 * | |
| MIC3 | 1:16,000 | 1.500 ± 0.357 | 0.506 ± 0.078 | 0.0093 ** | 2.671 ± 0.249 | 0.999 ± 0.111 | 0.0004 *** |
| 1:32,000 | 0.886 ± 0.187 | 0.322 ± 0.052 | 0.0073 ** | 1.654 ± 0.224 | 0.640 ± 0.107 | 0.0021 ** | |
| P35 | 1:16,000 | 2.410 ± 0.290 | 1.047 ± 0.054 | 0.0013 ** | 1.733 ± 0.080 | 0.841 ± 0.020 | <0.0001 **** |
| 1:32,000 | 1.613 ± 0.334 | 0.514 ± 0.026 | 0.0047 ** | 1.062 ± 0.034 | 0.393 ± 0.020 | <0.0001**** | |
| ROP1 | 1:16,000 | 1.695 ± 0.042 | 0.541 ± 0.148 | 0.0002 *** | 2.333 ± 0.439 | 0.800 ± 0.070 | 0.0039 ** |
| 1:32,000 | 0.968 ± 0.060 | 0.338 ± 0.087 | 0.0005 *** | 1.518 ± 0.377 | 0.491 ± 0.046 | 0.0094 ** | |
| SAG1 | 1:16,000 | 1.479 ± 0.430 | 0.508 ± 0.004 | 0.0174 * | 2.145 ± 0.129 | 0.865 ± 0.057 | <0.0001 **** |
| 1:32,000 | 0.873 ± 0.268 | 0.299 ± 0.007 | 0.0207 * | 1.189 ± 0.117 | 0.534 ± 0.046 | 0.0008 *** | |
| SAG2 | 1:16,000 | 1.885 ± 0.324 | 0.634 ± 0.028 | 0.0026 ** | 2.625 ± 0.330 | 0.718 ± 0.046 | 0.0006 *** |
| 1:32,000 | 1.118 ± 0.183 | 0.368 ± 0.015 | 0.0021 ** | 1.639 ± 0.294 | 0.518 ± 0.061 | 0.0030 ** | |
| SAG4 | 1:16,000 | 1.454 ± 0.527 | 0.700 ± 0.122 | 0.0730 | 1.625 ± 0.171 | 1.000 ± 0.025 | 0.0033 ** |
| 1:32,000 | 0.908 ± 0.369 | 0.444 ± 0.042 | 0.0961 | 0.974 ± 0.109 | 0.586 ± 0.017 | 0.0037 ** | |
| Recombinant Chimeric Protein | Amino Acid Residues | Additional Immunodominant Fragment | Amino Acid Residues | Number of Amino Acid Residues | Mw [kDa] | pI |
|---|---|---|---|---|---|---|
| SAG1-SAG2-AMA1 | SAG1 amino acid residues from 49-310 SAG2 amino acid residues from 30-170 | AMA1 | from 67-568 | 983 | 106.08 | 5.78 |
| SAG1-SAG2-AMA1S | AMA1S | from 67-483 | 898 | 96.76 | 5.97 | |
| SAG1-SAG2-GRA1 | GRA1 | from 24-190 | 648 | 68.05 | 5.00 | |
| SAG1-SAG2-GRA2 | GRA2 | from 51-185 | 616 | 64.84 | 6.60 | |
| SAG1-SAG2-GRA5 | GRA5 | from 26-120 | 576 | 60.53 | 5.93 | |
| SAG1-SAG2-GRA6 | GRA6 | from 30-228 | 681 | 70.92 | 5.75 | |
| SAG1-SAG2-GRA7 | GRA7 | from 27-236 | 691 | 73.38 | 5.61 | |
| SAG1-SAG2-GRA9 | GRA9 | from 21-318 | 779 | 83.43 | 5.69 | |
| SAG1-SAG2-LDH2 | LDH2 | from 2-326 | 806 | 85.33 | 6.26 | |
| SAG1-SAG2-MAG1 | MAG1 | from 30-452 | 904 | 96.34 | 5.24 | |
| SAG1-SAG2-MAG1S | MAG1S | from 30-222 | 674 | 70.82 | 5.09 | |
| SAG1-SAG2-MIC1 | MIC1 | from 25-456 | 913 | 96.30 | 5.71 | |
| SAG1-SAG2-MIC1S | MIC1ex2 | from 25-182 | 639 | 67.75 | 6.75 | |
| SAG1-SAG2-MIC3 | MIC3 | from 67-359 | 774 | 81.09 | 6.09 | |
| SAG1-SAG2-P35 | P35 | from 26-377 | 834 | 88.13 | 8.98 | |
| SAG1-SAG2-P35S | P35S | from 26-170 | 626 | 65.28 | 6.28 | |
| SAG1-SAG2-ROP1 | ROP1 | from 85-396 | 793 | 83.69 | 6.07 | |
| SAG1-SAG2 | - | - | 481 | 50.15 | 6.15 |
| Antigen | Calculated Cut-Off | ROC Cut-Off | Sensitivity [%] (95% CI) | Specificity [%] (95% CI) | Area under the Curve (95% CI) | p Value |
|---|---|---|---|---|---|---|
| SAG1-SAG2-AMA1 | 0.5962 | 0.6630 | 94.55 (88.61–97.48) | 100 (95.52–100) | 0.9909 (0.9816–1.000) | <0.0001 |
| SAG1-SAG2-AMA1S | 0.7912 | 0.7450 | 100 (96.63–100) | 100 (95.52–100) | 1.000 (1.000–1.000) | <0.0001 |
| SAG1-SAG2-GRA1 | 0.4376 | 0.5278 | 100 (96.63–100) | 100 (95.52–100) | 1.000 (1.000–1.000) | <0.0001 |
| SAG1-SAG2-GRA2 | 0.5133 | 0.5690 | 98.18 (93.61–99.68) | 100 (95.52–100) | 0.9990 (0.9970–1.000) | <0.0001 |
| SAG1-SAG2-GRA5 | 0.5271 | 0.5485 | 100 (96.63–100) | 100 (95.52–100) | 1.000 (1.000–1.000) | <0.0001 |
| SAG1-SAG2-GRA6 | 0.4525 | 0.5320 | 96.36 (91.02–98.58) | 100 (95.52–100) | 0.9928 (0.9847–1.000) | <0.0001 |
| SAG1-SAG2-GRA7 | 0.3914 | 0.4568 | 94.55 (88.61–97.48) | 100 (95.52–100) | 0.9978 (0.9949–1.000) | <0.0001 |
| SAG1-SAG2-GRA9 | 0.6491 | 0.6643 | 100 (96.63–100) | 100 (95.52–100) | 1.000 (1.000–1.000) | <0.0001 |
| SAG1-SAG2-LDH2 | 0.6246 | 0.5573 | 98.18 (93.61–99.68) | 100 (95.52–100) | 0.9990 (0.9970–1.000) | <0.0001 |
| SAG1-SAG2-MAG1 | 0.5095 | 0.5178 | 100 (96.63–100) | 100 (95.52–100) | 1.000 (1.000–1.000) | <0.0001 |
| SAG1-SAG2-MAG1S | 0.5563 | 0.6113 | 96.36 (91.02–98.58) | 100 (95.52–100) | 0.9971 (0.9934–1.000) | <0.0001 |
| SAG1-SAG2-MIC1 | 0.5001 | 0.6185 | 100 (96.63–100) | 100 (95.52–100) | 1.000 (1.000–1.000) | <0.0001 |
| SAG1-SAG2-MIC1S | 0.8130 | 0.8130 | 98.18 (93.61–99.68) | 100 (95.52–100) | 0.9968 (0.9919–1.000) | <0.0001 |
| SAG1-SAG2-MIC3 | 0.6005 | 0.7188 | 100 (96.63–100) | 100 (95.52–100) | 1.000 (1.000–1.000) | <0.0001 |
| SAG1-SAG2-P35 | 0.5134 | 0.6135 | 100 (96.63–100) | 100 (95.52–100) | 1.000 (1.000–1.000) | <0.0001 |
| SAG1-SAG2-P35S | 0.6213 | 0.6323 | 97.27 (92.29–99.26) | 100 (95.52–100) | 0.9991 (0.9976–1.000) | <0.0001 |
| SAG1-SAG2-ROP1 | 0.4276 | 0.3915 | 100 (96.63–100) | 100 (95.52–100) | 1.000 (1.000–1.000) | <0.0001 |
| SAG1-SAG2 | 0.4910 | 0.5168 | 100 (96.63–100) | 100 (95.52–100) | 1.000 (1.000–1.000) | <0.0001 |
| TLA | 0.3737 | 0.4378 | 100 (96.63–100) | 100 (95.52–100) | 1.000 (1.000–1.000) | <0.0001 |
| Antigen | Calculated Cut-Off | ROC Cut-Off | Sensitivity [%] (95% CI) | Specificity [%] (95% CI) | Area under the Curve (95% CI) | p Value |
|---|---|---|---|---|---|---|
| SAG1-SAG2-GRA1 | 0.5249 | 0.5095 | 98 (89.50–99.90) | 100 (92.13–100) | 0.9924 (0.9773–1.000) | <0.0001 |
| SAG1-SAG2-GRA2 | 0.4680 | 0.7560 | 100 (92.87–100) | 100 (92.13–100) | 1.000 (1.000–1.000) | <0.0001 |
| SAG1-SAG2-GRA5 | 0.5077 | 0.5558 | 100 (92.87–100) | 100 (92.13–100) | 1.000 (1.000–1.000) | <0.0001 |
| SAG1-SAG2-GRA6 | 0.5021 | 0.4810 | 90 (78.64–95.56) | 100 (92.13–100) | 0.9467 (0.8938–0.9995) | <0.0001 |
| SAG1-SAG2-GRA7 | 0.4981 | 0.5053 | 96 (86.54–98.36) | 100 (92.13–100) | 0.9829 (0.9581–1.000) | <0.0001 |
| SAG1-SAG2-GRA9 | 0.4721 | 0.6790 | 100 (92.87–100) | 100 (92.13–100) | 1.000 (1.000–1.000) | <0.0001 |
| SAG1-SAG2-MAG1 | 0.5132 | 0.4978 | 92 (81.16–96.85) | 100 (92.13–100) | 0.9556 (0.9050–1.000) | <0.0001 |
| SAG1-SAG2-MIC1 | 0.4992 | 0.6893 | 100 (92.87–100) | 100 (92.13–100) | 1.000 (1.000–1.000) | <0.0001 |
| SAG1-SAG2-MIC3 | 0.6300 | 0.7129 | 100 (92.87–100) | 100 (92.13–100) | 1.000 (1.000–1.1000) | <0.0001 |
| SAG1-SAG2-P35 | 0.4795 | 0.5898 | 100 (92.87–100) | 100 (92.13–100) | 1.000 (1.000–1.000) | <0.0001 |
| SAG1-SAG2-ROP1 | 0.4357 | 0.4158 | 100 (92.87–100) | 100 (92.13–100) | 1.000 (1.000–1.000) | <0.0001 |
| SAG1-SAG2 | 0.4799 | 0.5555 | 98 (89.50–99.90) | 100 (92.13–100) | 0.9973 (0.9915–1.000) | <0.0001 |
| TLA | 0.4652 | 0.7580 | 100 (92.87–100) | 100 (92.13–100) | 1.000 (1.000–1.000) | <0.0001 |
| Recombinant Chimeric Protein | Sheep Serum Samples | Goat Serum Samples | |||||
|---|---|---|---|---|---|---|---|
| r (95% CI) | r2 | p Value | r (95% CI) | r2 | p Value | ||
| SAG1-SAG2-GRA1 | vs. TLA vs. SAG1-SAG2 | 0.9301 (0.9081–0.9470) 0.9254 (0.9019–0.9433) | 0.8651 0.8563 | <0.0001 <0.0001 | 0.9634 (0.9455–0.9756) 0.9867 (0.9081–0.9912) | 0.9282 0.9737 | <0.0001 <0.0001 |
| SAG1-SAG2-GRA2 | vs. TLA vs. SAG1-SAG2 | 0.9254 (0.9020–0.9434) 0.9584 (0.9451–0.9686) | 0.8564 0.9186 | <0.0001 <0.0001 | 0.9635 (0.9456–0.9756) 0.9865 (0.9798–0.9910) | 0.9284 0.9733 | <0.0001 <0.0001 |
| SAG1-SAG2-GRA5 | vs. TLA vs. SAG1-SAG2 | 0.7883 (0.7279–0.8365) 0.7851 (0.7240–0.8340) | 0.6214 0.6164 | <0.0001 <0.0001 | 0.9659 (0.9492–0.9772) 0.9762 (0.9643–0.9841) | 0.9330 0.9529 | <0.0001 <0.0001 |
| SAG1-SAG2-GRA6 | vs. TLA vs. SAG1-SAG2 | 0.9372 (0.9174–0.9524) 0.9328 (0.9116–0.9491) | 0.8784 0.8702 | <0.0001 <0.0001 | 0.9119 (0.8703–0.9406) 0.9777 (0.9667–0.9851) | 0.8316 0.9559 | <0.0001 <0.0001 |
| SAG1-SAG2-GRA7 | vs. TLA vs. SAG1-SAG2 | 0.8883 (0.8542–0.9149) 0.9287 (0.9063–0.9459) | 0.7892 0.8625 | <0.0001 <0.0001 | 0.9383 (0.9086–0.9586) 0.9900 (0.9850–0.9933) | 0.8804 0.9801 | <0.0001 <0.0001 |
| SAG1-SAG2-GRA9 | vs. TLA vs. SAG1-SAG2 | 0.8750 (0.8371–0.9045) 0.8763 (0.8388–0.9056) | 0.7656 0.7680 | <0.0001 <0.0001 | 0.9680 (0.9523–0.9786) 0.9857 (0.9786–0.9905) | 0.9371 0.9717 | <0.0001 <0.0001 |
| SAG1-SAG2-MAG1 | vs. TLA vs. SAG1-SAG2 | 0.8091 (0.7539–0.8530) 0.7708 (0.7063–0.8226) | 0.6546 0.5941 | <0.0001 <0.0001 | 0.9031 (0.8577–0.9345) 0.9803 (0.9704–0.9868) | 0.8156 0.9609 | <0.0001 <0.0001 |
| SAG1-SAG2-MIC1 | vs. TLA vs. SAG1-SAG2 | 0.8774 (0.8402–0.9064) 0.8578 (0.8153–0.8912) | 0.7698 0.7359 | <0.0001 <0.0001 | 0.9618 (0.9430–0.9744) 0.9827 (0.9741–0.9885) | 0.9250 0.9657 | <0.0001 <0.0001 |
| SAG1-SAG2-MIC3 | vs. TLA vs. SAG1-SAG2 | 0.9413 (0.9227–0.9555) 0.9592 (0.9461–0.9692) | 0.8860 0.9201 | <0.0001 <0.0001 | 0.9565 (0.9352–0.9709) 0.9677 (0.9759–0.9892) | 0.9149 0.9677 | <0.0001 <0.0001 |
| SAG1-SAG2-P35 | vs. TLA vs. SAG1-SAG2 | 0.9487 (0.9323–0.9612) 0.9609 (0.9483–0.9705) | 0.9000 0.9233 | <0.0001 <0.0001 | 0.9667 (0.9503–0.9777) 0.9861 (0.9792–0.9908) | 0.9344 0.9724 | <0.0001 <0.0001 |
| SAG1-SAG2-ROP1 | vs. TLA vs. SAG1-SAG2 | 0.8082 (0.7528–0.8523) 0.8005 (0.7432–0.8462) | 0.6532 0.6409 | <0.0001 <0.0001 | 0.9657 (0.9489–0.9771) 0.9871 (0.9807–0.9914) | 0.9326 0.9744 | <0.0001 <0.0001 |
| SAG1-SAG2 | vs. TLA | 0.9260 (0.9027–0.9438) | 0.8574 | <0.0001 | 0.9388 (0.9093–0.9589) | 0.8813 | <0.0001 |
| Recombinant Chimeric Protein | Gene Fragment | Primer Sequence | Corresponding to Protein Residues |
|---|---|---|---|
| SAG1-SAG2-AMA1 | ama1 | SS-AMA1For 5′-CTCAACCATGGCGATCACGTCGGGGAATCCCTTTCA-3′ SS-AMA1Rev 5′-GAATTCGGATCCGATTCCCCCTCGACCATAACATGTG-3′ | 67-568 |
| SAG1-SAG2-AMA1S | sag1 | SS-SAG1For 5′-TGGACAGCCCAGATCCGGATCCCCCTCTTGTTGC-3′ SS-SAG1Rev 5′-TGGGCGCTGGCGTCTCAGCCGATTTTGCTGAC-3′ | 49-310 |
| sag2 | SS-SAG2For 5′-GTCAGCAAAATCGGCTGAGACGCCAGCGCCCA-3′ SS-S2/AMA1For 5′- AGGGATTCCCCGACGTCGTGAGAGACACAGGG-3′ | 30-170 | |
| ama1 | SSA-AMA1For 5′-CCCTGTGTCTCTCACGACGTCGGGGAATCCCT-3′ SSA-AMA1Rev 5′-ATCGGTACCCAGATCAGTGTTAGAGCCACATTCATTTTGTTCG-3′ | 67-483 | |
| SAG1-SAG2-GRA1 | gra1 | SS-GRA1For 5′-CTCAACCATGGCGATCGCTGCCGAAGGCG-3′ SS-GRA1Rev 5′-GAATTCGGATCCGATTCTCTCTCTCCTGTTAGGAACCCAAT-3′ | 24-190 |
| SAG1-SAG2-GRA2 | sag1 | SS-SAG1For 5′-TGGACAGCCCAGATCCGGATCCCCCTCTTGTTGC-3′ SS-SAG1Rev 5′-TGGGCGCTGGCGTCTCAGCCGATTTTGCTGAC-3′ | 49-310 |
| sag2 | SS-SAG2For 5′-GTCAGCAAAATCGGCTGAGACGCCAGCGCCCA-3′ SS-S2/GRA2Rev 5′-GGTGTATGTTCACCTTTTCCCGTGAGAGACACAGGGTC-3′ | 30-170 | |
| gra2 | SSG-GRA2For 5′-GACCCTGTGTCTCTCACGGGAAAAGGTGAACATACACC-3′ SSG-GRA2Rev 5′-ATCGGTACCCAGATCCTGCGAAAAGTCTGGGACGG-3′ | 51-185 | |
| SAG1-SAG2-GRA5 | gra5 | SS-GRA5For 5′-CTCAACCATGGCGATCGGTTCAACGCGTGACG-3′ SS-GRA5Rev 5′-GAATTCGGATCCGATTCTTCCTCGGCAACTTCTTCCT-3′ | 26-120 |
| SAG1-SAG2-GRA6 | gra6 | SS-GRA6For 5′-CTCAACCATGGCGATCATGGGTGTACTCGTCAATTCGTTG-3′ SS-GRA6Rev 5′-GAATTCGGATCCGATTCAAACACATTCACACGTTCCGG-3′ | 30-228 |
| SAG1-SAG2-GRA7 | gra7 | SS-GRA7For 5′-CTCAACCATGGCGATGGCCACCGCGTCAGAT-3′ SS-GRA7Rev 5′-GAATTCGGATCCGATTGGCGGGCATCCTCCC-3′ | 27-236 |
| SAG1-SAG2-GRA9 | gra9 | SS-GRA9For 5′-CTCAACCATGGCGATACTCGACCTTTTCCTCGGTGAA-3′ SS-GRA9Rev 5′-GAATTCGGATCCGATAGTCCTCGGTCTTCCTGCG-3′ | 21-318 |
| SAG1-SAG2-LDH2 | sag1 | SS-SAG1For 5′-TGGACAGCCCAGATCCGGATCCCCCTCTTGTTGC-3′ SS-SAG1Rev 5′-TGGGCGCTGGCGTCTCAGCCGATTTTGCTGAC-3′ | 49-310 |
| sag2 | SS-SAG2For 5′-GTCAGCAAAATCGGCTGAGACGCCAGCGCCCA-3′ SSL-S2/LDH2Rev 5′-TGCTAACGGTACCCGTCGTGAGAGACACAGGG-3′ | 30-170 | |
| ldh2 | SSL-LDH2For 5′-CCCTGTGTCTCTCACGACGGGTACCGTTAGCA-3′ SSL-LDH2Rev 5′-ATCGGTACCCAGATCACCCAGCGCCGCT-3′ | 2-326 | |
| SAG1-SAG2-MAG1 | mag1 | SS-MAG1For 5′-CTCAACCATGGCGATGAGCCAAAGGGTGCCAGAG-3′ SS-MAG1Rev 5′-GAATTCGGATCCGATGCTGCCTGTTCCGCTAAGAT-3′ | 30-452 |
| SAG1-SAG2-MAG1S | mag1 | SS-MAG1For 5′-CTCAACCATGGCGATGAGCCAAAGGGTGCCAGAG-3′ SS-MAG1SRev 5′-GAATTCGGATCCGATTTCTTGATGGCTTCCAACTGCT-3′ | 30-222 |
| SAG1-SAG2-MIC1 | mic1 | SS-MIC1For 5′-CTCAACCATGGCGATAGCGTCGCATTCTCATTCGC-3′ SS-MIC1Rev 5′-GAATTCGGATCCGATGCAGAGACGGCCGTAGG-3′ | 25-456 |
| SAG1-SAG2-MIC1S | sag1 | SS-SAG1For 5′-TGGACAGCCCAGATCCGGATCCCCCTCTTGTTGC-3′ SS-SAG1Rev 5′-TGGGCGCTGGCGTCTCAGCCGATTTTGCTGAC-3′ | 49-310 |
| sag2 | SS-SAG2For 5′-GTCAGCAAAATCGGCTGAGACGCCAGCGCCCA-3′ SSM-S2/MIC1Rev 5′-CGAATGAGAATGCGACGCCGTGAGAGACACAGGGT-3′ | 30-170 | |
| mic1 | SSM-MIC1For 5′-ACCCTGTGTCTCTCACGGCGTCGCATTCTCATTCG-3′ SSM-MIC1Rev 5′-ATCGGTACCCAGATCCTTCTCGTAACACCTCCACGCA-3′ | 25-182 | |
| SAG1-SAG2-MIC3 | sag1 | SS-SAG1For 5′-TGGACAGCCCAGATCCGGATCCCCCTCTTGTTGC-3′ SS-SAG1Rev 5′-TGGGCGCTGGCGTCTCAGCCGATTTTGCTGAC-3′ | 49-310 |
| sag2 | SS-SAG2For 5′-GTCAGCAAAATCGGCTGAGACGCCAGCGCCCA-3′ SSM-S2/MIC3Rev 5′-CCTGCTTGCTGGGGGACGTGAGAGACACAGG-3′ | 30-170 | |
| mic3 | SSM-MIC3For 5′-CCTGTGTCTCTCACGTCCCCCAGCAAGCAGG-3′ SSM-MIC3Rev 5′-ATCGGTACCCAGATCCTGCTTAATTTTCTCACACGTCACGG-3′ | 67-359 | |
| SAG1-SAG2-P35 | p35 | SS-P35For 5′-CTCAACCATGGCGATCGGTCCTTTGAGTTATCATCCAAGC-3′ SS-P35Rev 5′-GAATTCGGATCCGATTTCTGCGTCGTTACGGTGAATCT-3′ | 26-377 |
| SAG1-SAG2-P35S | sag1 | SS-SAG1For 5′-TGGACAGCCCAGATCCGGATCCCCCTCTTGTTGC-3′ SS-SAG1Rev 5′-TGGGCGCTGGCGTCTCAGCCGATTTTGCTGAC-3′ | 49-310 |
| sag2 | SS-SAG2For 5′-GTCAGCAAAATCGGCTGAGACGCCAGCGCCCA-3′ SSP-S2/P35Rev 5′-GATGATAACTCAAAGGACCCGTGAGAGACACAGGGTC-3′ | 30-170 | |
| p35 | SSP-P35For 5′-GACCCTGTGTCTCTCACGGGTCCTTTGAGTTATCATC-3′ SSP-P35Rev 5′-ATCGGTACCCAGATCAGCAGCTGTCGTGGTTGT-3′ | 26-170 | |
| SAG1-SAG2-ROP1 | sag1 | SS-SAG1For 5′-TGGACAGCCCAGATCCGGATCCCCCTCTTGTTGC-3′ SS-SAG1Rev 5′-TGGGCGCTGGCGTCTCAGCCGATTTTGCTGAC-3′ | 49-310 |
| sag2 | SS-SAG2For 5′-GTCAGCAAAATCGGCTGAGACGCCAGCGCCCA-3′ SSR-S2/ROP1Rev 5′-CGGGCCTCTGACAGGCGTGAGAGACACAGG-3′ | 30-170 | |
| rop1 | SSR-ROP1For 5′-CCTGTGTCTCTCACGCCTGTCAGAGGCCCG-3′ SSR-ROP1Rev 5′-ATCGGTACCCAGATCTTGCGATCCATCATCCTGCTCTC-3′ | 85-396 | |
| SAG1-SAG2 | sag1 | SS-SAG1For 5′-TGGACAGCCCAGATCCGGATCCCCCTCTTGTTGC-3′ SS-SAG1Rev 5′-TGGGCGCTGGCGTCTCAGCCGATTTTGCTGAC-3′ | 49-310 |
| sag2 | SS-SAG2For 5′-GTCAGCAAAATCGGCTGAGACGCCAGCGCCCA-3′ SS-SAG2Rev 5′-ATCGGTACCCAGATCCGTGAGAGACACAGGGTCAAAC-3′ | 30-170 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Ferra, B.T.; Chyb, M.; Sołowińska, K.; Holec-Gąsior, L.; Skwarecka, M.; Baranowicz, K.; Gatkowska, J. The Development of Toxoplasma gondii Recombinant Trivalent Chimeric Proteins as an Alternative to Toxoplasma Lysate Antigen (TLA) in Enzyme-Linked Immunosorbent Assay (ELISA) for the Detection of Immunoglobulin G (IgG) in Small Ruminants. Int. J. Mol. Sci. 2024, 25, 4384. https://doi.org/10.3390/ijms25084384
Ferra BT, Chyb M, Sołowińska K, Holec-Gąsior L, Skwarecka M, Baranowicz K, Gatkowska J. The Development of Toxoplasma gondii Recombinant Trivalent Chimeric Proteins as an Alternative to Toxoplasma Lysate Antigen (TLA) in Enzyme-Linked Immunosorbent Assay (ELISA) for the Detection of Immunoglobulin G (IgG) in Small Ruminants. International Journal of Molecular Sciences. 2024; 25(8):4384. https://doi.org/10.3390/ijms25084384
Chicago/Turabian StyleFerra, Bartłomiej Tomasz, Maciej Chyb, Karolina Sołowińska, Lucyna Holec-Gąsior, Marta Skwarecka, Karolina Baranowicz, and Justyna Gatkowska. 2024. "The Development of Toxoplasma gondii Recombinant Trivalent Chimeric Proteins as an Alternative to Toxoplasma Lysate Antigen (TLA) in Enzyme-Linked Immunosorbent Assay (ELISA) for the Detection of Immunoglobulin G (IgG) in Small Ruminants" International Journal of Molecular Sciences 25, no. 8: 4384. https://doi.org/10.3390/ijms25084384
APA StyleFerra, B. T., Chyb, M., Sołowińska, K., Holec-Gąsior, L., Skwarecka, M., Baranowicz, K., & Gatkowska, J. (2024). The Development of Toxoplasma gondii Recombinant Trivalent Chimeric Proteins as an Alternative to Toxoplasma Lysate Antigen (TLA) in Enzyme-Linked Immunosorbent Assay (ELISA) for the Detection of Immunoglobulin G (IgG) in Small Ruminants. International Journal of Molecular Sciences, 25(8), 4384. https://doi.org/10.3390/ijms25084384