
Environmental Exposure to Persistent Organic Pollutants and Its Association with Endometriosis Risk: Implications in the Epithelial–Mesenchymal Transition Process

 , , , , and
, , , , and 
Abstract
1. Introduction
2. Results
2.1. Adipose Tissue Concentrations of Selected POPs
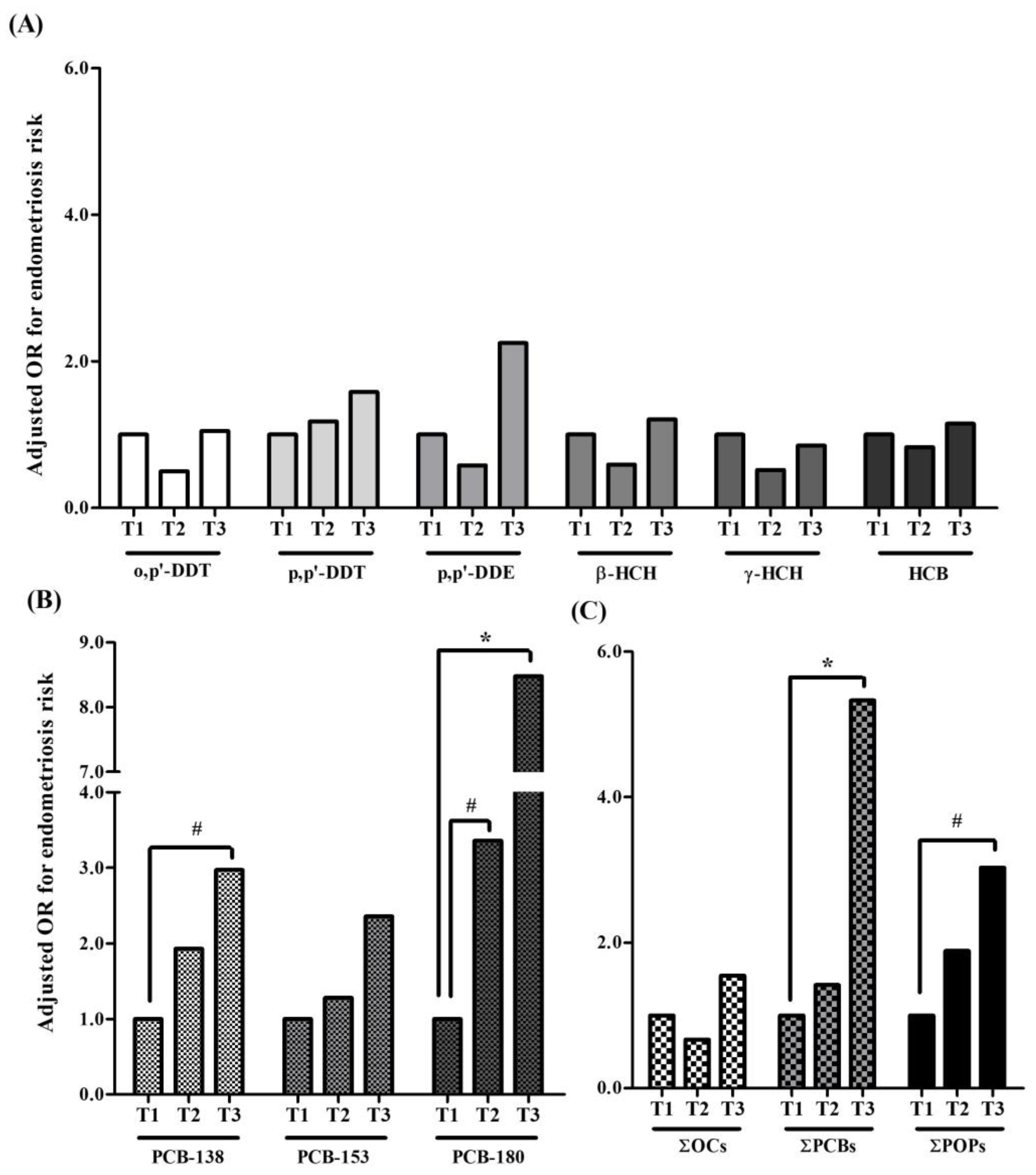
2.2. Associations between Chronic Adipose Tissue POP Concentrations and Endometriosis Risk
2.3. Associations between Adipose Tissue Levels of POPs and Expression Profile of EMT-Related Genes
3. Discussion
4. Materials and Methods
4.1. Study Population and Sample Collection
4.2. Adipose Tissue Sample Extraction and Chemical Analyses
4.3. RNA Isolation and Quantitative Real-Time Polymerase Chain Reaction (qRT-PCR)
4.4. Statistical Analysis
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Buck Louis, G.M.; Hediger, M.L.; Peterson, C.M.; Croughan, M.; Sundaram, R.; Stanford, J.; Chen, Z.; Fujimoto, V.Y.; Varner, M.W.; Trumble, A.; et al. Incidence of endometriosis by study population and diagnostic method: The ENDO study. Fertil. Steril. 2011, 96, 360–365. [Google Scholar] [CrossRef] [PubMed]
- Eisenberg, V.H.; Weil, C.; Chodick, G.; Shalev, V. Epidemiology of endometriosis: A large population-based database study from a healthcare provider with 2 million members. BJOG Int. J. Obstet. Gynaecol. 2018, 125, 55–62. [Google Scholar] [CrossRef]
- Zondervan, K.T.; Becker, C.M.; Missmer, S.A. Endometriosis. N. Engl. J. Med. 2020, 382, 1244–1256. [Google Scholar] [CrossRef]
- Vercellini, P.; Viganò, P.; Somigliana, E.; Fedele, L. Endometriosis: Pathogenesis and treatment. Nat. Rev. Endocrinol. 2014, 10, 261–275. [Google Scholar] [CrossRef] [PubMed]
- Chen, S.; Liu, Y.; Zhong, Z.; Wei, C.; Liu, Y.; Zhu, X. Peritoneal immune microenvironment of endometriosis: Role and therapeutic perspectives. Front. Immunol. 2023, 14, 1134663. [Google Scholar] [CrossRef] [PubMed]
- Giudice, L.C.; Kao, L.C. Endometriosis. Lancet 2004, 364, 1789–1799. [Google Scholar] [CrossRef]
- Eskenazi, B.; Warner, M.L. Epidemiology of endometriosis. Obstet. Gynecol. Clin. N. Am. 1997, 24, 235–258. [Google Scholar] [CrossRef]
- Matias-Guiu, X.; Stewart, C.J.R. Endometriosis-associated ovarian neoplasia. Pathology 2018, 50, 190–204. [Google Scholar] [CrossRef] [PubMed]
- Ruderman, R.; Pavone, M.E. Ovarian cancer in endometriosis: An update on the clinical and molecular aspects. Minerva Ginecol. 2017, 69, 286–294. [Google Scholar] [CrossRef]
- Benagiano, G.; Brosens, I.; Habiba, M. Structural and molecular features of the endomyometrium in endometriosis and adenomyosis. Hum. Reprod. Updat. 2014, 20, 386–402. [Google Scholar] [CrossRef]
- Kvaskoff, M.; Mu, F.; Terry, K.L.; Harris, H.R.; Poole, E.M.; Farland, L.; Missmer, S.A. Endometriosis: A high-risk population for major chronic diseases? Hum. Reprod. Updat. 2015, 21, 500–516. [Google Scholar] [CrossRef] [PubMed]
- Smarr, M.M.; Kannan, K.; Buck Louis, G.M. Endocrine disrupting chemicals and endometriosis. Fertil. Steril. 2016, 106, 959–966. [Google Scholar] [CrossRef] [PubMed]
- Kahn, L.G.; Philippat, C.; Nakayama, S.F.; Slama, R.; Trasande, L. Endocrine-disrupting chemicals: Implications for human health. Lancet Diabetes Endocrinol. 2020, 8, 703–718. [Google Scholar] [CrossRef] [PubMed]
- La Merrill, M.; Emond, C.; Kim, M.J.; Antignac, J.P.; Le Bizec, B.; Clément, K.; Birnbaum, L.S.; Barouki, R. Toxicological function of adipose tissue: Focus on persistent organic pollutants. Environ. Health Perspect. 2013, 121, 162–169. [Google Scholar] [CrossRef] [PubMed]
- Kohlmeier, L.; Kohlmeier, M. Adipose tissue as a medium for epidemiologic exposure assessment. Environ. Health Perspect. 1995, 103 (Suppl. S3), 99–106. [Google Scholar] [CrossRef] [PubMed]
- Artacho-Cordón, F.; Fernández-Rodríguez, M.; Garde, C.; Salamanca, E.; Iribarne-Durán, L.M.; Torné, P.; Expósito, J.; Papay-Ramírez, L.; Fernández, M.F.; Olea, N.; et al. Serum and adipose tissue as matrices for assessment of exposure to persistent organic pollutants in breast cancer patients. Environ. Res. 2015, 142, 633–643. [Google Scholar] [CrossRef] [PubMed]
- Bokobza, E.; Hinault, C.; Tiroille, V.; Clavel, S.; Bost, F.; Chevalier, N. The Adipose Tissue at the Crosstalk Between EDCs and Cancer Development. Front. Endocrinol. 2021, 12, 691658. [Google Scholar] [CrossRef] [PubMed]
- Bonefeld Jørgensen, E.C.; Autrup, H.; Hansen, J.C. Effect of toxaphene on estrogen receptor functions in human breast cancer cells. Carcinogenesis 1997, 18, 1651–1654. [Google Scholar] [CrossRef] [PubMed]
- Bonefeld-Jørgensen, E.C.; Andersen, H.R.; Rasmussen, T.H.; Vinggaard, A.M. Effect of highly bioaccumulated polychlorinated biphenyl congeners on estrogen and androgen receptor activity. Toxicology 2001, 158, 141–153. [Google Scholar] [CrossRef]
- Grünfeld, H.T.; Bonefeld-Jorgensen, E.C. Effect of in vitro estrogenic pesticides on human oestrogen receptor alpha and beta mRNA levels. Toxicol. Lett. 2004, 151, 467–480. [Google Scholar] [CrossRef]
- Sonnenschein, C.; Soto, A.M. An updated review of environmental estrogen and androgen mimics and antagonists. J. Steroid Biochem. Mol. Biol. 1998, 65, 143–150. [Google Scholar] [CrossRef] [PubMed]
- Soto, A.M.; Michaelson, C.L.; Prechtl, N.V.; Weill, B.C.; Sonnenschein, C.; Olea-Serrano, F.; Olea, N. Assays to measure estrogen and androgen agonists and antagonists. Adv. Exp. Med. Biol. 1998, 444, 9–23; discussion 23–28. [Google Scholar] [CrossRef] [PubMed]
- Arrebola, J.P.; Belhassen, H.; Artacho-Cordón, F.; Ghali, R.; Ghorbel, H.; Boussen, H.; Perez-Carrascosa, F.M.; Expósito, J.; Hedhili, A.; Olea, N. Risk of female breast cancer and serum concentrations of organochlorine pesticides and polychlorinated biphenyls: A case-control study in Tunisia. Sci. Total Environ. 2015, 520, 106–113. [Google Scholar] [CrossRef] [PubMed]
- Pastor-Barriuso, R.; Fernández, M.F.; Castaño-Vinyals, G.; Whelan, D.; Pérez-Gómez, B.; Llorca, J.; Villanueva, C.M.; Guevara, M.; Molina-Molina, J.M.; Artacho-Cordón, F.; et al. Total Effective Xenoestrogen Burden in Serum Samples and Risk for Breast Cancer in a Population-Based Multicase-Control Study in Spain. Environ. Health Perspect. 2016, 124, 1575–1582. [Google Scholar] [CrossRef] [PubMed]
- Costa, E.M.; Spritzer, P.M.; Hohl, A.; Bachega, T.A. Effects of endocrine disruptors in the development of the female reproductive tract. Arq. Bras. Endocrinol. Metabol. 2014, 58, 153–161. [Google Scholar] [CrossRef] [PubMed]
- Génard-Walton, M.; Warembourg, C.; Duros, S.; Mercier, F.; Lefebvre, T.; Guivarc’h-Levêque, A.; Le Martelot, M.T.; Le Bot, B.; Jacquemin, B.; Chevrier, C.; et al. Serum persistent organic pollutants and diminished ovarian reserve: A single-exposure and mixture exposure approach from a French case-control study. Hum. Reprod. 2023, 38, 701–715. [Google Scholar] [CrossRef] [PubMed]
- Lefebvre, T.; Fréour, T.; Ploteau, S.; Marchand, P.; Le Bizec, B.; Antignac, J.P.; Cano-Sancho, G. Mixtures of persistent organic pollutants and ovarian function in women undergoing IVF. Reprod. Biomed. Online 2023, 46, 129–137. [Google Scholar] [CrossRef] [PubMed]
- Syed, S.; Qasim, S.; Ejaz, M.; Sammar; Khan, N.; Ali, H.; Zaker, H.; Hatzidaki, E.; Mamoulakis, C.; Tsatsakis, A.; et al. Effects of Dichlorodiphenyltrichloroethane on the Female Reproductive Tract Leading to Infertility and Cancer: Systematic Search and Review. Toxics 2023, 11, 725. [Google Scholar] [CrossRef] [PubMed]
- Eick, S.M.; Tan, Y.; Taibl, K.R.; Barry Ryan, P.; Barr, D.B.; Hüls, A.; Eatman, J.A.; Panuwet, P.; D’Souza, P.E.; Yakimavets, V.; et al. Prenatal exposure to persistent and non-persistent chemical mixtures and associations with adverse birth outcomes in the Atlanta African American Maternal-Child Cohort. J. Expo. Sci. Environ. Epidemiol. 2023. [Google Scholar] [CrossRef] [PubMed]
- Yang, Q.; Zhao, Y.; Qiu, X.; Zhang, C.; Li, R.; Qiao, J. Association of serum levels of typical organic pollutants with polycystic ovary syndrome (PCOS): A case-control study. Hum. Reprod. 2015, 30, 1964–1973. [Google Scholar] [CrossRef]
- Trabert, B.; Chen, Z.; Kannan, K.; Peterson, C.M.; Pollack, A.Z.; Sun, L.; Buck Louis, G.M. Persistent organic pollutants (POPs) and fibroids: Results from the ENDO study. J. Expo. Sci. Environ. Epidemiol. 2015, 25, 278–285. [Google Scholar] [CrossRef] [PubMed]
- Wen, X.; Xiong, Y.; Qu, X.; Jin, L.; Zhou, C.; Zhang, M.; Zhang, Y. The risk of endometriosis after exposure to endocrine-disrupting chemicals: A meta-analysis of 30 epidemiology studies. Gynecol. Endocrinol. Off. J. Int. Soc. Gynecol. Endocrinol. 2019, 35, 645–650. [Google Scholar] [CrossRef]
- Ploteau, S.; Antignac, J.P.; Volteau, C.; Marchand, P.; Vénisseau, A.; Vacher, V.; Le Bizec, B. Distribution of persistent organic pollutants in serum, omental, and parietal adipose tissue of French women with deep infiltrating endometriosis and circulating versus stored ratio as new marker of exposure. Environ. Int. 2016, 97, 125–136. [Google Scholar] [CrossRef]
- Porpora, M.G.; Ingelido, A.M.; di Domenico, A.; Ferro, A.; Crobu, M.; Pallante, D.; Cardelli, M.; Cosmi, E.V.; De Felip, E. Increased levels of polychlorobiphenyls in Italian women with endometriosis. Chemosphere 2006, 63, 1361–1367. [Google Scholar] [CrossRef]
- Vichi, S.; Medda, E.; Ingelido, A.M.; Ferro, A.; Resta, S.; Porpora, M.G.; Abballe, A.; Nisticò, L.; De Felip, E.; Gemma, S.; et al. Glutathione transferase polymorphisms and risk of endometriosis associated with polychlorinated biphenyls exposure in Italian women: A gene-environment interaction. Fertil. Steril. 2012, 97, 1143–1151.e3. [Google Scholar] [CrossRef]
- Tsukino, H.; Hanaoka, T.; Sasaki, H.; Motoyama, H.; Hiroshima, M.; Tanaka, T.; Kabuto, M.; Niskar, A.S.; Rubin, C.; Patterson, D.G., Jr.; et al. Associations between serum levels of selected organochlorine compounds and endometriosis in infertile Japanese women. Environ. Res. 2005, 99, 118–125. [Google Scholar] [CrossRef]
- Niskar, A.S.; Needham, L.L.; Rubin, C.; Turner, W.E.; Martin, C.A.; Patterson, D.G., Jr.; Hasty, L.; Wong, L.Y.; Marcus, M. Serum dioxins, polychlorinated biphenyls, and endometriosis: A case-control study in Atlanta. Chemosphere 2009, 74, 944–949. [Google Scholar] [CrossRef] [PubMed]
- Trabert, B.; De Roos, A.J.; Schwartz, S.M.; Peters, U.; Scholes, D.; Barr, D.B.; Holt, V.L. Non-dioxin-like polychlorinated biphenyls and risk of endometriosis. Environ. Health Perspect. 2010, 118, 1280–1285. [Google Scholar] [CrossRef]
- Klemmt, P.A.B.; Starzinski-Powitz, A. Molecular and Cellular Pathogenesis of Endometriosis. Curr. Womens Health Rev. 2018, 14, 106–116. [Google Scholar] [CrossRef]
- Yang, Y.M.; Yang, W.X. Epithelial-to-mesenchymal transition in the development of endometriosis. Oncotarget 2017, 8, 41679–41689. [Google Scholar] [CrossRef]
- Artacho-Cordón, F.; Ríos-Arrabal, S.; Lara, P.C.; Artacho-Cordón, A.; Calvente, I.; Núñez, M.I. Matrix metalloproteinases: Potential therapy to prevent the development of second malignancies after breast radiotherapy. Surg. Oncol. 2012, 21, e143–e151. [Google Scholar] [CrossRef]
- Radisky, E.S.; Radisky, D.C. Matrix metalloproteinase-induced epithelial-mesenchymal transition in breast cancer. J. Mammary Gland Biol. Neoplasia 2010, 15, 201–212. [Google Scholar] [CrossRef]
- Lamouille, S.; Xu, J.; Derynck, R. Molecular mechanisms of epithelial-mesenchymal transition. Nat. Rev. Mol. Cell Biol. 2014, 15, 178–196. [Google Scholar] [CrossRef]
- Konrad, L.; Dietze, R.; Riaz, M.A.; Scheiner-Bobis, G.; Behnke, J.; Horné, F.; Hoerscher, A.; Reising, C.; Meinhold-Heerlein, I. Epithelial-Mesenchymal Transition in Endometriosis-When Does It Happen? J. Clin. Med. 2020, 9, 1915. [Google Scholar] [CrossRef]
- Chattopadhyay, I.; Ambati, R.; Gundamaraju, R. Exploring the Crosstalk between Inflammation and Epithelial-Mesenchymal Transition in Cancer. Mediat. Inflamm. 2021, 2021, 9918379. [Google Scholar] [CrossRef]
- Wu, J.; Li, X.; Huang, H.; Xia, X.; Zhang, M.; Fang, X. TET1 may contribute to hypoxia-induced epithelial to mesenchymal transition of endometrial epithelial cells in endometriosis. PeerJ 2020, 8, e9950. [Google Scholar] [CrossRef]
- Lih Yuan, T.; Sulaiman, N.; Nur Azurah, A.G.; Maarof, M.; Rabiatul Adawiyah, R.; Yazid, M.D. Oestrogen-induced epithelial-mesenchymal transition (EMT) in endometriosis: Aetiology of vaginal agenesis in Mayer-Rokitansky-Küster-Hauser (MRKH) syndrome. Front. Physiol. 2022, 13, 937988. [Google Scholar] [CrossRef]
- Xiong, W.; Zhang, L.; Liu, H.; Li, N.; Du, Y.; He, H.; Zhang, Z.; Liu, Y. E(2)-mediated EMT by activation of β-catenin/Snail signalling during the development of ovarian endometriosis. J. Cell. Mol. Med. 2019, 23, 8035–8045. [Google Scholar] [CrossRef]
- Pujals, M.; Resar, L.; Villanueva, J. HMGA1, Moonlighting Protein Function, and Cellular Real Estate: Location, Location, Location! Biomolecules 2021, 11, 1334. [Google Scholar] [CrossRef]
- Gorbacheva, A.M.; Uvarova, A.N.; Ustiugova, A.S.; Bhattacharyya, A.; Korneev, K.V.; Kuprash, D.V.; Mitkin, N.A. EGR1 and RXRA transcription factors link TGF-β pathway and CCL2 expression in triple negative breast cancer cells. Sci. Rep. 2021, 11, 14120. [Google Scholar] [CrossRef]
- Sommerova, L.; Ondrouskova, E.; Vojtesek, B.; Hrstka, R. Suppression of AGR2 in a TGF-β-induced Smad regulatory pathway mediates epithelial-mesenchymal transition. BMC Cancer 2017, 17, 546. [Google Scholar] [CrossRef]
- Tu, C.F.; Wu, M.Y.; Lin, Y.C.; Kannagi, R.; Yang, R.B. FUT8 promotes breast cancer cell invasiveness by remodeling TGF-β receptor core fucosylation. Breast Cancer Res. BCR 2017, 19, 111. [Google Scholar] [CrossRef]
- Yang, W.; Cui, X.; Sun, D.; Sun, G.; Yan, Z.; Wei, M.; Wang, Z.; Yu, W. POU5F1 promotes the proliferation, migration, and invasion of gastric cancer cells by reducing the ubiquitination level of TRAF6. Cell Death Dis. 2023, 14, 802. [Google Scholar] [CrossRef]
- Song, C.; Zhou, C. HOXA10 mediates epithelial-mesenchymal transition to promote gastric cancer metastasis partly via modulation of TGFB2/Smad/METTL3 signaling axis. J. Exp. Clin. Cancer Res. CR 2021, 40, 62. [Google Scholar] [CrossRef]
- Moncho-Amor, V.; Pintado-Berninches, L.; Ibañez de Cáceres, I.; Martín-Villar, E.; Quintanilla, M.; Chakravarty, P.; Cortes-Sempere, M.; Fernández-Varas, B.; Rodriguez-Antolín, C.; de Castro, J.; et al. Role of Dusp6 Phosphatase as a Tumor Suppressor in Non-Small Cell Lung Cancer. Int. J. Mol. Sci. 2019, 20, 2036. [Google Scholar] [CrossRef]
- Song, S.; Zhang, R.; Cao, W.; Fang, G.; Yu, Y.; Wan, Y.; Wang, C.; Li, Y.; Wang, Q. Foxm1 is a critical driver of TGF-β-induced EndMT in endothelial cells through Smad2/3 and binds to the Snail promoter. J. Cell. Physiol. 2019, 234, 9052–9064. [Google Scholar] [CrossRef]
- Li, J.; Zhang, Y.; Ye, F.; Qian, P.; Qin, Z.; Li, D.; Ye, L.; Feng, L. DKK1 Promotes Epithelial-Mesenchymal Transition and Cisplatin Resistance in Gastric Cancer via Activation of the PI3K/AKT Pathway. Cancers 2023, 15, 4756. [Google Scholar] [CrossRef]
- Qi, L.; Sun, B.; Liu, Z.; Li, H.; Gao, J.; Leng, X. Dickkopf-1 inhibits epithelial-mesenchymal transition of colon cancer cells and contributes to colon cancer suppression. Cancer Sci. 2012, 103, 828–835. [Google Scholar] [CrossRef]
- Zhang, B.; Zhang, M.; Li, Q.; Yang, Y.; Shang, Z.; Luo, J. TPX2 mediates prostate cancer epithelial-mesenchymal transition through CDK1 regulated phosphorylation of ERK/GSK3β/SNAIL pathway. Biochem. Biophys. Res. Commun. 2021, 546, 1–6. [Google Scholar] [CrossRef]
- Li, X.; Xu, Y.; Chen, Y.; Chen, S.; Jia, X.; Sun, T.; Liu, Y.; Li, X.; Xiang, R.; Li, N. SOX2 promotes tumor metastasis by stimulating epithelial-to-mesenchymal transition via regulation of WNT/β-catenin signal network. Cancer Lett. 2013, 336, 379–389. [Google Scholar] [CrossRef]
- Lee, M.M.; Wong, Y.H. CCR1-mediated activation of Nuclear Factor-kappaB in THP-1 monocytic cells involves Pertussis Toxin-insensitive Galpha(14) and Galpha(16) signaling cascades. J. Leukoc. Biol. 2009, 86, 1319–1329. [Google Scholar] [CrossRef]
- Liu, B.; Wang, Z.; Cheng, S.; Du, L.; Yin, Y.; Yang, Z.; Zhou, J. miR-379 inhibits cell proliferation and epithelial-mesenchymal transition by targeting CHUK through the NF-κB pathway in non-small cell lung cancer. Mol. Med. Rep. 2019, 20, 1418–1428. [Google Scholar] [CrossRef]
- Yang, M.; Yao, P.; Lang, X.; Li, X.; Zhang, D. Ribonucleotide reductase subunit M2 promotes proliferation and epithelial-mesenchymal transition via the JAK2/STAT3 signaling pathway in retinoblastoma. Bioengineered 2021, 12, 12800–12811. [Google Scholar] [CrossRef]
- Gogola-Mruk, J.; Hoffmann-Młodzianowska, M.; Kamińska, K.; Ptak, A. Mixtures of persistent organic pollutants increase ovarian granulosa tumor cell line migration and spheroid invasion by upregulating MMP2 expression and activity via IGF1R. Toxicology 2021, 452, 152715. [Google Scholar] [CrossRef]
- Zucchini-Pascal, N.; Peyre, L.; de Sousa, G.; Rahmani, R. Organochlorine pesticides induce epithelial to mesenchymal transition of human primary cultured hepatocytes. Food Chem. Toxicol. Int. J. Publ. Br. Ind. Biol. Res. Assoc. 2012, 50, 3963–3970. [Google Scholar] [CrossRef]
- Peinado, F.M.; Olivas-Martínez, A.; Iribarne-Durán, L.M.; Ubiña, A.; León, J.; Vela-Soria, F.; Fernández-Parra, J.; Fernández, M.F.; Olea, N.; Freire, C.; et al. Cell cycle, apoptosis, cell differentiation, and lipid metabolism gene expression in endometriotic tissue and exposure to parabens and benzophenones. Sci. Total Environ. 2023, 879, 163014. [Google Scholar] [CrossRef]
- Peinado, F.M.; Olivas-Martínez, A.; Lendínez, I.; Iribarne-Durán, L.M.; León, J.; Fernández, M.F.; Sotelo, R.; Vela-Soria, F.; Olea, N.; Freire, C.; et al. Expression Profiles of Genes Related to Development and Progression of Endometriosis and Their Association with Paraben and Benzophenone Exposure. Int. J. Mol. Sci. 2023, 24, 6678. [Google Scholar] [CrossRef]
- Lavogina, D.; Visser, N.; Samuel, K.; Davey, E.; Björvang, R.D.; Hassan, J.; Koponen, J.; Rantakokko, P.; Kiviranta, H.; Rinken, A.; et al. Endocrine disrupting chemicals interfere with decidualization of human primary endometrial stromal cells in vitro. Front. Endocrinol. 2022, 13, 903505. [Google Scholar] [CrossRef]
- Hu, T.; Yao, M.; Fu, X.; Chen, C.; Wu, R. Polychlorinated biphenyl 104 promotes migration of endometrial stromal cells in endometriosis. Toxicol. Lett. 2018, 290, 19–28. [Google Scholar] [CrossRef]
- Willing, C.; Peich, M.; Danescu, A.; Kehlen, A.; Fowler, P.A.; Hombach-Klonisch, S. Estrogen-independent actions of environmentally relevant AhR-agonists in human endometrial epithelial cells. Mol. Hum. Reprod. 2011, 17, 115–126. [Google Scholar] [CrossRef]
- Ploteau, S.; Cano-Sancho, G.; Volteau, C.; Legrand, A.; Vénisseau, A.; Vacher, V.; Marchand, P.; Le Bizec, B.; Antignac, J.P. Associations between internal exposure levels of persistent organic pollutants in adipose tissue and deep infiltrating endometriosis with or without concurrent ovarian endometrioma. Environ. Int. 2017, 108, 195–203. [Google Scholar] [CrossRef]
- Porpora, M.G.; Medda, E.; Abballe, A.; Bolli, S.; De Angelis, I.; di Domenico, A.; Ferro, A.; Ingelido, A.M.; Maggi, A.; Panici, P.B.; et al. Endometriosis and organochlorinated environmental pollutants: A case-control study on Italian women of reproductive age. Environ. Health Perspect. 2009, 117, 1070–1075. [Google Scholar] [CrossRef]
- Matta, K.; Vigneau, E.; Cariou, V.; Mouret, D.; Ploteau, S.; Le Bizec, B.; Antignac, J.P.; Cano-Sancho, G. Associations between persistent organic pollutants and endometriosis: A multipollutant assessment using machine learning algorithms. Environ. Pollut. 2020, 260, 114066. [Google Scholar] [CrossRef]
- Buck Louis, G.M.; Chen, Z.; Peterson, C.M.; Hediger, M.L.; Croughan, M.S.; Sundaram, R.; Stanford, J.B.; Varner, M.W.; Fujimoto, V.Y.; Giudice, L.C.; et al. Persistent lipophilic environmental chemicals and endometriosis: The ENDO Study. Environ. Health Perspect. 2012, 120, 811–816. [Google Scholar] [CrossRef]
- Martínez-Zamora, M.A.; Mattioli, L.; Parera, J.; Abad, E.; Coloma, J.L.; van Babel, B.; Galceran, M.T.; Balasch, J.; Carmona, F. Increased levels of dioxin-like substances in adipose tissue in patients with deep infiltrating endometriosis. Hum. Reprod. 2015, 30, 1059–1068. [Google Scholar] [CrossRef]
- De Felip, E.; Porpora, M.G.; di Domenico, A.; Ingelido, A.M.; Cardelli, M.; Cosmi, E.V.; Donnez, J. Dioxin-like compounds and endometriosis: A study on Italian and Belgian women of reproductive age. Toxicol. Lett. 2004, 150, 203–209. [Google Scholar] [CrossRef]
- Pauwels, A.; Schepens, P.J.; D’Hooghe, T.; Delbeke, L.; Dhont, M.; Brouwer, A.; Weyler, J. The risk of endometriosis and exposure to dioxins and polychlorinated biphenyls: A case-control study of infertile women. Hum. Reprod. 2001, 16, 2050–2055. [Google Scholar] [CrossRef]
- Fierens, S.; Mairesse, H.; Heilier, J.F.; De Burbure, C.; Focant, J.F.; Eppe, G.; De Pauw, E.; Bernard, A. Dioxin/polychlorinated biphenyl body burden, diabetes and endometriosis: Findings in a population-based study in Belgium. Biomark. Biochem. Indic. Expo. Response Susceptibility Chem. 2003, 8, 529–534. [Google Scholar] [CrossRef]
- Lebel, G.; Dodin, S.; Ayotte, P.; Marcoux, S.; Ferron, L.A.; Dewailly, E. Organochlorine exposure and the risk of endometriosis. Fertil. Steril. 1998, 69, 221–228. [Google Scholar] [CrossRef]
- Bratton, M.R.; Frigo, D.E.; Segar, H.C.; Nephew, K.P.; McLachlan, J.A.; Wiese, T.E.; Burow, M.E. The organochlorine o,p’-DDT plays a role in coactivator-mediated MAPK crosstalk in MCF-7 breast cancer cells. Environ. Health Perspect. 2012, 120, 1291–1296. [Google Scholar] [CrossRef]
- Nieto-Nicolau, N.; de la Torre, R.M.; Fariñas, O.; Savio, A.; Vilarrodona, A.; Casaroli-Marano, R.P. Extrinsic modulation of integrin α6 and progenitor cell behavior in mesenchymal stem cells. Stem Cell Res. 2020, 47, 101899. [Google Scholar] [CrossRef]
- Chiappini, F.; Ceballos, L.; Olivares, C.; Bastón, J.I.; Miret, N.; Pontillo, C.; Zárate, L.; Singla, J.J.; Farina, M.; Meresman, G.; et al. Endocrine disruptor hexachlorobenzene induces cell migration and invasion, and enhances aromatase expression levels in human endometrial stromal cells. Food Chem. Toxicol. Int. J. Publ. Br. Ind. Biol. Res. Assoc. 2022, 162, 112867. [Google Scholar] [CrossRef]
- Rivero, J.; Henríquez-Hernández, L.A.; Luzardo, O.P.; Pestano, J.; Zumbado, M.; Boada, L.D.; Valerón, P.F. Differential gene expression pattern in human mammary epithelial cells induced by realistic organochlorine mixtures described in healthy women and in women diagnosed with breast cancer. Toxicol. Lett. 2016, 246, 42–48. [Google Scholar] [CrossRef]
- Frigo, D.E.; Tang, Y.; Beckman, B.S.; Scandurro, A.B.; Alam, J.; Burow, M.E.; McLachlan, J.A. Mechanism of AP-1-mediated gene expression by select organochlorines through the p38 MAPK pathway. Carcinogenesis 2004, 25, 249–261. [Google Scholar] [CrossRef]
- Ambolet-Camoit, A.; Ottolenghi, C.; Leblanc, A.; Kim, M.J.; Letourneur, F.; Jacques, S.; Cagnard, N.; Guguen-Guillouzo, C.; Barouki, R.; Aggerbeck, M. Two persistent organic pollutants which act through different xenosensors (alpha-endosulfan and 2,3,7,8 tetrachlorodibenzo-p-dioxin) interact in a mixture and downregulate multiple genes involved in human hepatocyte lipid and glucose metabolism. Biochimie 2015, 116, 79–91. [Google Scholar] [CrossRef]
- Padilla-Banks, E.; Jefferson, W.N.; Papas, B.N.; Suen, A.A.; Xu, X.; Carreon, D.V.; Willson, C.J.; Quist, E.M.; Williams, C.J. Endocrine disruption of early uterine differentiation causes adenocarcinoma mediated by Wnt/β-catenin- and PI3K/AKT signaling. bioRxiv 2022. [Google Scholar] [CrossRef]
- Ricciardi, M.; Zanotto, M.; Malpeli, G.; Bassi, G.; Perbellini, O.; Chilosi, M.; Bifari, F.; Krampera, M. Epithelial-to-mesenchymal transition (EMT) induced by inflammatory priming elicits mesenchymal stromal cell-like immune-modulatory properties in cancer cells. Br. J. Cancer 2015, 112, 1067–1075. [Google Scholar] [CrossRef]
- Suarez-Carmona, M.; Lesage, J.; Cataldo, D.; Gilles, C. EMT and inflammation: Inseparable actors of cancer progression. Mol. Oncol. 2017, 11, 805–823. [Google Scholar] [CrossRef]
- Lv, N.; Gao, Y.; Guan, H.; Wu, D.; Ding, S.; Teng, W.; Shan, Z. Inflammatory mediators, tumor necrosis factor-α and interferon-γ, induce EMT in human PTC cell lines. Oncol. Lett. 2015, 10, 2591–2597. [Google Scholar] [CrossRef]
- Peinado, F.M.; Artacho-Cordón, F.; Barrios-Rodríguez, R.; Arrebola, J.P. Influence of polychlorinated biphenyls and organochlorine pesticides on the inflammatory milieu. A systematic review of in vitro, in vivo and epidemiological studies. Environ. Res. 2020, 186, 109561. [Google Scholar] [CrossRef]
- Thompson, L.A.; Ikenaka, Y.; Sobhy Darwish, W.; Nakayama, S.M.M.; Mizukawa, H.; Ishizuka, M. Effects of the organochlorine p,p’-DDT on MCF-7 cells: Investigating metabolic and immune modulatory transcriptomic changes. Environ. Toxicol. Pharmacol. 2019, 72, 103249. [Google Scholar] [CrossRef]
- Osman, A.M.; van Loveren, H. Phosphoproteomic analysis of mouse thymoma cells treated with tributyltin oxide: TBTO affects proliferation and energy sensing pathways. Toxicol. Sci. Off. J. Soc. Toxicol. 2012, 126, 84–100. [Google Scholar] [CrossRef]
- Ono, Y.J.; Hayashi, M.; Tanabe, A.; Hayashi, A.; Kanemura, M.; Terai, Y.; Ohmichi, M. Estradiol-mediated hepatocyte growth factor is involved in the implantation of endometriotic cells via the mesothelial-to-mesenchymal transition in the peritoneum. Am. J. Physiol. Endocrinol. Metab. 2015, 308, E950–E959. [Google Scholar] [CrossRef]
- Luo, M.; Hou, L.; Li, J.; Shao, S.; Huang, S.; Meng, D.; Liu, L.; Feng, L.; Xia, P.; Qin, T.; et al. VEGF/NRP-1axis promotes progression of breast cancer via enhancement of epithelial-mesenchymal transition and activation of NF-κB and β-catenin. Cancer Lett. 2016, 373, 1–11. [Google Scholar] [CrossRef]
- Clere, N.; Lauret, E.; Malthiery, Y.; Andriantsitohaina, R.; Faure, S. Estrogen receptor alpha as a key target of organochlorines to promote angiogenesis. Angiogenesis 2012, 15, 745–760. [Google Scholar] [CrossRef][Green Version]
- Zárate, L.V.; Pontillo, C.A.; Español, A.; Miret, N.V.; Chiappini, F.; Cocca, C.; Álvarez, L.; de Pisarev, D.K.; Sales, M.E.; Randi, A.S. Angiogenesis signaling in breast cancer models is induced by hexachlorobenzene and chlorpyrifos, pesticide ligands of the aryl hydrocarbon receptor. Toxicol. Appl. Pharmacol. 2020, 401, 115093. [Google Scholar] [CrossRef]
- Quintana, P.J.; Delfino, R.J.; Korrick, S.; Ziogas, A.; Kutz, F.W.; Jones, E.L.; Laden, F.; Garshick, E. Adipose tissue levels of organochlorine pesticides and polychlorinated biphenyls and risk of non-Hodgkin’s lymphoma. Environ. Health Perspect. 2004, 112, 854–861. [Google Scholar] [CrossRef]
- Vela-Soria, F.; García-Villanova, J.; Mustieles, V.; de Haro, T.; Antignac, J.P.; Fernandez, M.F. Assessment of perfluoroalkyl substances in placenta by coupling salt assisted liquid-liquid extraction with dispersive liquid-liquid microextraction prior to liquid chromatography-tandem mass spectrometry. Talanta 2021, 221, 121577. [Google Scholar] [CrossRef]
- Roy, D.; Morgan, M.; Yoo, C.; Deoraj, A.; Roy, S.; Yadav, V.K.; Garoub, M.; Assaggaf, H.; Doke, M. Integrated Bioinformatics, Environmental Epidemiologic and Genomic Approaches to Identify Environmental and Molecular Links between Endometriosis and Breast Cancer. Int. J. Mol. Sci. 2015, 16, 25285–25322. [Google Scholar] [CrossRef]


{kind=link}
| Cases (n = 34) | Controls (n = 75) | |||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|
| % >LOD | Mean | SD | Percentiles | Mean | SD | Percentiles | ||||||
| 25th | 50th | 75th | 25th | 50th | 75th | p-Value | ||||||
| o,p’-DDT | 96.30 | 0.89 | 1.19 | 0.28 | 0.46 | 0.83 | 0.50 | 0.38 | 0.27 | 0.40 | 0.60 | 0.360 |
| p,p’-DDT | 89.90 | 3.50 | 4.94 | 0.94 | 1.72 | 3.61 | 1.84 | 1.84 | 0.19 | 1.40 | 2.53 | 0.065 |
| p,p’-DDE | 100.00 | 282.05 | 552.90 | 48.03 | 118.52 | 226.42 | 103.35 | 84.47 | 41.96 | 74.33 | 136.76 | 0.067 |
| β-HCH | 100.00 | 32.79 | 45.86 | 4.31 | 14.88 | 37.12 | 16.67 | 17.18 | 5.46 | 11.09 | 22.34 | 0.261 |
| γ-HCH | 100.00 | 25.10 | 34.27 | 5.07 | 11.27 | 28.91 | 12.80 | 12.53 | 4.60 | 8.32 | 16.80 | 0.204 |
| HCB | 100.00 | 15.43 | 14.81 | 5.05 | 6.88 | 24.57 | 9.99 | 7.32 | 5.26 | 7.62 | 11.68 | 0.513 |
| ∑OCPs 1 | - | 1.16 | 1.93 | 0.24 | 0.55 | 1.13 | 0.47 | 0.35 | 0.21 | 0.35 | 0.60 | 0.072 |
| PCB-138 | 99.10 | 30.41 | 30.83 | 10.39 | 20.29 | 38.95 | 16.85 | 12.33 | 7.76 | 13.74 | 24.03 | 0.015 |
| PCB-153 | 100.00 | 39.77 | 41.32 | 12.47 | 26.76 | 45.48 | 21.90 | 16.99 | 9.21 | 18.50 | 31.05 | 0.012 |
| PCB-180 | 100.00 | 64.52 | 87.79 | 18.28 | 39.04 | 70.14 | 31.25 | 28.72 | 9.82 | 20.54 | 50.62 | 0.006 |
| ∑PCBs 1 | - | 0.36 | 0.42 | 0.11 | 0.25 | 0.37 | 0.19 | 0.15 | 0.07 | 0.15 | 0.27 | 0.009 |
| ∑POPs 1 | - | 1.51 | 2.15 | 0.43 | 0.76 | 1.78 | 0.66 | 0.46 | 0.30 | 0.52 | 0.92 | 0.018 |
| OR | IC 95% | p-Value | OR 1 | IC 95% | p-Value | |||
|---|---|---|---|---|---|---|---|---|
| o,p’-DDT | 1.45 | 0.87 | 2.42 | 0.152 | 1.35 | 0.78 | 2.34 | 0.284 |
| p,p’-DDT | 1.62 | 1.14 | 2.32 | 0.008 | 1.63 | 1.13 | 2.34 | 0.008 |
| p,p’-DDE | 1.49 | 0.98 | 2.26 | 0.061 | 1.42 | 0.90 | 2.26 | 0.135 |
| β-HCH | 1.11 | 0.83 | 1.48 | 0.499 | 0.99 | 0.72 | 1.36 | 0.935 |
| γ-HCH | 1.25 | 0.84 | 1.86 | 0.268 | 1.08 | 0.69 | 1.67 | 0.747 |
| HCB | 1.26 | 0.75 | 2.13 | 0.382 | 1.16 | 0.63 | 2.10 | 0.636 |
| ∑OCPs 1 | 1.54 | 0.98 | 2.42 | 0.060 | 1.44 | 0.87 | 2.40 | 0.159 |
| PCB-138 | 1.60 | 0.98 | 2.62 | 0.059 | 1.65 | 0.93 | 2.92 | 0.086 |
| PCB-153 | 1.59 | 0.99 | 2.53 | 0.054 | 1.65 | 0.95 | 2.87 | 0.075 |
| PCB-180 | 1.63 | 1.08 | 2.46 | 0.021 | 1.89 | 1.09 | 3.27 | 0.023 |
| ∑PCBs 1 | 1.64 | 1.04 | 2.58 | 0.034 | 1.78 | 1.02 | 3.12 | 0.044 |
| ∑POPs 1 | 1.76 | 1.07 | 2.88 | 0.025 | 1.75 | 0.99 | 3.10 | 0.056 |
| o’p’-DDT | p’,p’-DDT | p’p’-DDE | β-HCH | γ-HCH | HCB | ∑OCPs | PCB-138 | PCB-153 | PCB-180 | ∑PCBs | ∑POPs | |||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| CORE EMT MARKERS | CLDN7 a | ρ | −0.46 | −0.41 | −0.49 | −0.26 | −0.18 | −0.29 | −0.45 | −0.18 | −0.16 | −0.16 | −0.15 | −0.43 |
| p-value | 0.011 | 0.026 | 0.005 | 0.157 | 0.346 | 0.124 | 0.013 | 0.341 | 0.396 | 0.407 | 0.415 | 0.018 | ||
| ITGB2 a | ρ | 0.59 | 0.52 | 0.56 | 0.22 | 0.13 | 0.33 | 0.49 | 0.25 | 0.23 | 0.20 | 0.20 | 0.49 | |
| p-value | 0.001 | 0.004 | 0.001 | 0.247 | 0.493 | 0.072 | 0.006 | 0.176 | 0.223 | 0.300 | 0.289 | 0.006 | ||
| MMP1 b | ≤0.12 | 0.47 (0.53) | 1.64 (3.44) | 102.46 (171.71) | 21.97 (64.91) | 13.04 (46.69) | 6.98 (22.06) | 0.58 (0.80) | 20.44 (31.95) | 26.82 (41.64) | 49.88 (52.72) | 0.27 (0.34) | 0.84 (1.27) | |
| >0.12 | 0.51 (0.85) | 1.93 (2.69) | 134.57 (369.65) | 14.00 (28.98) | 10.56 (20.85) | 6.78 (11.77) | 0.53 (1.36) | 20.04 (27.62) | 25.60 (30.15) | 34.41 (39.29) | 0.23 (0.21) | 0.74 (1.82) | ||
| p-value | 0.885 | 0.548 | 0.468 | 0.520 | 0.373 | 0.787 | 0.694 | 0.663 | 0.604 | 0.310 | 0.373 | 0.983 | ||
| TIMP1 b | ≤0.39 | 0.58 (0.89) | 1.43 (3.10) | 102.46 (222.07) | 14.66 (28.98) | 10.97 (19.73) | 6.44 (17.31) | 0.48 (1.42) | 19.16 (16.06) | 25.41 (19.93) | 34.41 (34.22) | 0.22 (0.19) | 0.68 (1.82) | |
| >0.39 | 0.47 (0.57) | 1.93 (2.56) | 134.57 (163.70) | 19.34 (64.91) | 13.41 (47.86) | 7.87 (24.58) | 0.58 (0.73) | 22.82 (32.08) | 30.94 (41.84) | 51.76 (85.41) | 0.28 (0.42) | 0.77 (1.26) | ||
| p-value | 0.724 | 0.633 | 0.633 | 0.604 | 0.633 | 0.443 | 0.604 | 0.290 | 0.221 | 0.310 | 0.290 | 0.548 | ||
| TGF-β/PDGFRα SIGNALING PATHWAYS | AGR2 b | ≤0.10 | 0.58 (1.91) | 1.93 (5.68) | 136.30 (220.21) | 22.16 (59.93) | 16.70 (46.20) | 12.68 (21.97) | 0.73 (1.69) | 20.44 (33.71) | 26.28 (42.24) | 34.41 (55.54) | 0.22 (0.35) | 0.84 (1.83) |
| >0.10 | 0.45 (0.38) | 1.43 (1.75) | 72.36 (183.63) | 12.22 (23.46) | 9.19 (15.64) | 6.78 (5.33) | 0.34 (1.22) | 20.14 (9.93) | 26.82 (8.86) | 49.88 (47.68) | 0.25 (0.12) | 0.62 (1.43) | ||
| p-value | 0.237 | 0.191 | 0.395 | 0.237 | 0.191 | 0.548 | 0.351 | 0.724 | 0.633 | 0.310 | 0.548 | 0.694 | ||
| DUSP6 a | ρ | −0.29 | −0.31 | −0.29 | −0.31 | −0.27 | −0.23 | −0.31 | −0.20 | −0.23 | −0.13 | −0.18 | −0.29 | |
| p-value | 0.123 | 0.101 | 0.121 | 0.098 | 0.150 | 0.224 | 0.098 | 0.291 | 0.226 | 0.502 | 0.345 | 0.117 | ||
| FOXM1 a | ρ | 0.61 | 0.54 | 0.53 | 0.35 | 0.27 | 0.39 | 0.50 | 0.28 | 0.27 | 0.25 | 0.27 | 0.51 | |
| p-value | <0.001 | 0.002 | 0.003 | 0.059 | 0.150 | 0.034 | 0.005 | 0.134 | 0.146 | 0.177 | 0.142 | 0.004 | ||
| FUT8 b | ≤0.09 | 0.41 (0.77) | 1.41 (3.02) | 75.28 (212.33) | 14.33 (27.30) | 10.76 (18.37) | 6.50 (15.54) | 0.33 (1.37) | 19.04 (14.66) | 25.50 (19.28) | 34.82 (35.86) | 0.23 (0.20) | 0.54 (1.74) | |
| >0.09 | 0.57 (0.72) | 2.50 (2.51) | 171.75 (212.60) | 25.05 (75.48) | 18.88 (57.28) | 10.83 (34.83) | 0.79 (0.96) | 33.05 (56.55) | 42.33 (77.34) | 60.48 (114.59) | 0.36 (0.65) | 1.11 (1.83) | ||
| p-value | 0.598 | 0.291 | 0.312 | 0.253 | 0.312 | 0.538 | 0.235 | 0.078 | 0.086 | 0.173 | 0.173 | 0.173 | ||
| HMGA1 a | ρ | 0.57 | 0.50 | 0.56 | 0.34 | 0.27 | 0.35 | 0.51 | 0.29 | 0.28 | 0.25 | 0.27 | 0.53 | |
| p-value | 0.001 | 0.005 | 0.001 | 0.067 | 0.152 | 0.061 | 0.004 | 0.119 | 0.134 | 0.183 | 0.156 | 0.003 | ||
| HOXA10 a | ρ | 0.32 | 0.27 | 0.43 | 0.20 | 0.17 | 0.13 | 0.37 | 0.20 | 0.18 | 0.16 | 0.20 | 0.37 | |
| p-value | 0.084 | 0.154 | 0.018 | 0.286 | 0.379 | 0.500 | 0.043 | 0.296 | 0.332 | 0.396 | 0.295 | 0.047 | ||
| PDGFRA a | ρ | −0.15 | −0.20 | −0.08 | −0.27 | −0.26 | −0.19 | −0.17 | −0.19 | −0.20 | −0.17 | −0.19 | −0.18 | |
| p-value | 0.433 | 0.291 | 0.686 | 0.148 | 0.173 | 0.303 | 0.358 | 0.316 | 0.295 | 0.373 | 0.307 | 0.331 | ||
| POU5F1 b | ≤0.04 | 0.45 (0.53) | 1.33 (3.11) | 72.36 (223.61) | 9.54 (32.61) | 6.01 (20.85) | 6.44 (11.84) | 0.27 (1.43) | 20.04 (16.18) | 25.6 (20.13) | 35.24 (31.88) | 0.23 (0.20) | 0.50 (1.93) | |
| >0.04 | 0.51 (1.59) | 2.42 (4.79) | 136.3 (160.27) | 21.97 (53.72) | 13.41 (41.65) | 8.98 (24.75) | 0.73 (0.65) | 22.82 (31.95) | 30.94 (41.64) | 51.76 (84.05) | 0.28 (0.41) | 0.84 (1.17) | ||
| p-value | 0.443 | 0.130 | 0.272 | 0.078 | 0.152 | 0.419 | 0.178 | 0.419 | 0.330 | 0.494 | 0.419 | 0.290 | ||
| RXRA a | ρ | 0.03 | 0.01 | 0.00 | 0.00 | 0.01 | −0.03 | −0.01 | −0.06 | −0.04 | 0.00 | −0.02 | −0.03 | |
| p-value | 0.857 | 0.961 | 0.987 | 0.983 | 0.948 | 0.886 | 0.964 | 0.754 | 0.843 | 0.995 | 0.931 | 0.886 | ||
| WNT/β-CATENIN SIGNALING PATHWAY | BMI1 b | ≤0.58 | 0.51 (0.51) | 1.83 (3.02) | 153.84 (204.98) | 19.34 (20.73) | 13.04 (15.80) | 8.98 (16.18) | 0.58 (1.33) | 20.14 (13.49) | 26.82 (17.49) | 40.76 (33.44) | 0.25 (0.20) | 0.84 (1.38) |
| >0.58 | 0.47 (0.92) | 1.81 (3.11) | 78.19 (185.17) | 15.11 (64.91) | 11.39 (48.99) | 6.44 (33.57) | 0.36 (1.23) | 20.44 (62.07) | 26.70 (80.99) | 37.32 (110.51) | 0.25 (0.68) | 0.58 (1.99) | ||
| p-value | 0.395 | 0.604 | 0.290 | 0.983 | 0.983 | 0.694 | 0.373 | 0.787 | 0.756 | 0.724 | 0.756 | 0.395 | ||
| DKK1 a | ρ | 0.43 | 0.39 | 0.51 | 0.22 | 0.14 | 0.11 | 0.43 | 0.09 | 0.07 | −0.02 | 0.03 | 0.40 | |
| p-value | 0.017 | 0.034 | 0.004 | 0.243 | 0.469 | 0.572 | 0.017 | 0.644 | 0.714 | 0.899 | 0.872 | 0.029 | ||
| CDK1 a | ρ | 0.18 | 0.07 | 0.14 | −0.08 | −0.11 | 0.03 | 0.11 | 0.15 | 0.13 | 0.15 | 0.10 | 0.12 | |
| p-value | 0.355 | 0.709 | 0.466 | 0.673 | 0.546 | 0.856 | 0.546 | 0.434 | 0.505 | 0.431 | 0.607 | 0.511 | ||
| SOX2 b | ≤0.06 | 0.45 (0.53) | 1.33 (2.77) | 72.36 (222.07) | 12.22 (32.49) | 9.19 (20.70) | 6.44 (11.84) | 0.30 (1.42) | 20.04 (13.49) | 25.60 (17.49) | 35.24 (31.88) | 0.23 (0.19) | 0.50 (1.82) | |
| >0.06 | 0.51 (1.59) | 2.42 (4.79) | 136.3 (163.70) | 21.97 (57.03) | 13.41 (43.55) | 8.98 (24.75) | 0.73 (0.65) | 22.82 (35.82) | 30.94 (45.47) | 51.76 (85.41) | 0.28 (0.44) | 0.84 (1.17) | ||
| p-value | 0.576 | 0.191 | 0.494 | 0.206 | 0.351 | 0.468 | 0.351 | 0.633 | 0.520 | 0.694 | 0.633 | 0.419 | ||
| NOTCH/NF-κB SIGNALING PATHWAY | CCR1 a | ρ | 0.60 | 0.56 | 0.56 | 0.33 | 0.24 | 0.27 | 0.51 | 0.18 | 0.18 | 0.06 | 0.13 | 0.48 |
| p-value | <0.001 | 0.001 | 0.001 | 0.070 | 0.206 | 0.154 | 0.004 | 0.332 | 0.349 | 0.765 | 0.492 | 0.008 | ||
| CHUK a | ρ | 0.13 | 0.05 | 0.14 | −0.16 | −0.19 | −0.07 | 0.06 | −0.01 | −0.04 | −0.03 | −0.07 | 0.06 | |
| p-value | 0.505 | 0.793 | 0.445 | 0.393 | 0.327 | 0.718 | 0.741 | 0.938 | 0.825 | 0.890 | 0.732 | 0.761 | ||
| FOS b | ≤0.10 | 0.58 (0.78) | 1.64 (2.91) | 134.57 (175.97) | 19.34 (58.01) | 13.04 (42.16) | 6.78 (21.76) | 0.53 (0.98) | 20.44 (30.51) | 26.82 (39.83) | 40.76 (58.48) | 0.25 (0.36) | 0.74 (1.55) | |
| >0.10 | 0.47 (0.89) | 1.83 (3.38) | 98.56 (433.72) | 15.11 (26.53) | 11.39 (18.25) | 6.98 (8.98) | 0.58 (1.95) | 20.04 (15.97) | 26.70 (21.60) | 35.24 (58.33) | 0.25 (0.23) | 0.77 (2.13) | ||
| p-value | 0.700 | 0.910 | 0.700 | 0.803 | 0.769 | 0.455 | 0.734 | 0.512 | 0.455 | 0.769 | 0.603 | 0.734 | ||
| JUN b | ≤0.10 | 0.58 (1.13) | 2.12 (3.75) | 118.52 (260.86) | 17.23 (54.56) | 12.21 (39.36) | 8.42 (18.39) | 0.50 (1.49) | 20.28 (28.56) | 27.43 (33.01) | 34.82 (55.10) | 0.24 (0.27) | 0.71 (1.90) | |
| >0.10 | 0.42 (0.30) | 1.60 (1.33) | 126.2 (176.58) | 15.76 (24.21) | 11.84 (16.70) | 5.05 (17.87) | 0.58 (0.67) | 20.29 (46.32) | 26.76 (59.60) | 50.92 (84.81) | 0.26 (0.50) | 0.81 (1.02) | ||
| p-value | 0.482 | 0.542 | 0.778 | 0.963 | 0.888 | 0.223 | 0.851 | 0.851 | 0.888 | 0.373 | 0.606 | 0.888 | ||
| INFLAMMATION SIGNALING PATHWAY | IL1RL1 a | ρ | −0.01 | −0.06 | 0.02 | −0.13 | −0.14 | −0.26 | −0.03 | −0.14 | −0.14 | 0.01 | −0.08 | −0.01 |
| p-value | 0.950 | 0.759 | 0.917 | 0.496 | 0.467 | 0.173 | 0.859 | 0.477 | 0.456 | 0.964 | 0.675 | 0.965 | ||
| IL6ST a | ρ | 0.25 | 0.14 | 0.24 | −0.14 | −0.18 | −0.02 | 0.14 | 0.02 | −0.01 | −0.04 | −0.04 | 0.14 | |
| p-value | 0.186 | 0.462 | 0.200 | 0.469 | 0.341 | 0.927 | 0.469 | 0.919 | 0.960 | 0.827 | 0.827 | 0.468 | ||
| RRM2 a | ρ | 0.58 | 0.50 | 0.46 | 0.32 | 0.21 | 0.43 | 0.43 | 0.26 | 0.28 | 0.30 | 0.29 | 0.46 | |
| p-value | 0.001 | 0.005 | 0.012 | 0.088 | 0.263 | 0.019 | 0.018 | 0.173 | 0.139 | 0.107 | 0.123 | 0.010 | ||
| TNFRSF1B b | ≤0.10 | 0.36 (0.49) | 1.38 (2.43) | 64.58 (203.62) | 14.00 (27.98) | 10.56 (18.17) | 6.56 (15.30) | 0.30 (1.33) | 20.04 (19.78) | 26.28 (23.82) | 37.32 (39.44) | 0.25 (0.20) | 0.58 (1.69) | |
| >0.10 | 0.63 (1.58) | 2.88 (4.66) | 164.28 (262.20) | 23.82 (86.83) | 17.93 (65.67) | 12.68 (27.80) | 0.74 (1.33) | 24.04 (56.94) | 30.94 (82.43) | 51.96 (121.48) | 0.27 (0.69) | 0.85 (2.43) | ||
| p-value | 0.081 | 0.032 | 0.081 | 0.108 | 0.167 | 0.541 | 0.067 | 0.402 | 0.428 | 0.635 | 0.667 | 0.118 | ||
| ESTROGEN/GROWTH FACTOR SIGNALING PATHWAY | EGF b | ≤0.10 | 0.63 (1.92) | 2.57 (5.65) | 153.84 (204.98) | 21.97 (45.83) | 13.41 (36.52) | 10.24 (18.15) | 0.74 (1.63) | 20.14 (29.38) | 26.82 (39.20) | 40.76 (66.48) | 0.26 (0.39) | 0.84 (1.75) |
| >0.10 | 0.47 (0.28) | 1.81 (2.08) | 78.19 (183.63) | 14.66 (32.33) | 10.97 (20.70) | 6.44 (12.59) | 0.36 (1.22) | 20.44 (19.16) | 26.70 (24.00) | 37.32 (56.53) | 0.25 (0.26) | 0.58 (1.47) | ||
| p-value | 0.395 | 0.272 | 0.330 | 0.419 | 0.548 | 0.206 | 0.272 | 0.443 | 0.443 | 0.520 | 0.443 | 0.373 | ||
| ESR1 a | ρ | 0.37 | 0.33 | 0.43 | 0.12 | 0.05 | 0.09 | 0.35 | 0.05 | 0.03 | 0.00 | 0.02 | 0.33 | |
| p-value | 0.044 | 0.075 | 0.018 | 0.513 | 0.791 | 0.627 | 0.060 | 0.775 | 0.855 | 0.985 | 0.902 | 0.078 | ||
| FGF2 b | ≤0.12 | 0.58 (1.97) | 1.43 (5.73) | 102.46 (222.07) | 14.00 (23.46) | 10.56 (15.64) | 6.44 (18.76) | 0.48 (1.73) | 18.92 (25.94) | 25.41 (29.95) | 34.41 (31.36) | 0.22 (0.20) | 0.68 (1.82) | |
| >0.12 | 0.47 (0.55) | 1.93 (2.20) | 134.57 (183.09) | 19.34 (59.93) | 13.41 (46.20) | 7.87 (21.90) | 0.58 (1.14) | 22.82 (32.08) | 30.94 (41.84) | 51.96 (85.98) | 0.28 (0.42) | 0.77 (1.39) | ||
| p-value | 0.950 | 0.820 | 0.852 | 0.395 | 0.351 | 0.663 | 0.694 | 0.310 | 0.272 | 0.310 | 0.351 | 0.576 | ||
| HYPOXIA SIGNALING PATHWAY | ANG a | ρ | 0.41 | 0.37 | 0.37 | 0.20 | 0.14 | 0.23 | 0.31 | 0.17 | 0.16 | −0.01 | 0.05 | 0.30 |
| p-value | 0.024 | 0.047 | 0.043 | 0.290 | 0.471 | 0.228 | 0.094 | 0.357 | 0.398 | 0.940 | 0.775 | 0.113 | ||
| ANGPT1 a | ρ | −0.03 | −0.11 | −0.05 | −0.26 | −0.27 | −0.28 | −0.13 | −0.25 | −0.28 | −0.19 | −0.24 | −0.14 | |
| p-value | 0.883 | 0.549 | 0.783 | 0.166 | 0.145 | 0.130 | 0.506 | 0.178 | 0.131 | 0.302 | 0.202 | 0.472 | ||
| FLT1 b | ≤0.35 | 0.36 (2.00) | 1.38 (6.07) | 64.58 (223.61) | 14.66 (52.56) | 10.97 (38.61) | 5.79 (19.19) | 0.30 (1.74) | 18.48 (28.02) | 25.41 (31.74) | 34.41 (37.37) | 0.22 (0.21) | 0.58 (1.99) | |
| >0.35 | 0.51 (0.51) | 2.42 (1.97) | 136.30 (158.76) | 19.34 (25.46) | 13.41 (19.73) | 8.98 (21.59) | 0.73 (1.09) | 22.82 (23.45) | 30.94 (30.57) | 51.76 (73.53) | 0.27 (0.31) | 0.85 (1.38) | ||
| p-value | 0.419 | 0.330 | 0.272 | 0.633 | 0.604 | 0.237 | 0.272 | 0.141 | 0.191 | 0.237 | 0.290 | 0.206 | ||
| VEGFA b | ≤0.92 | 0.47 (0.60) | 1.81 (3.44) | 78.19 (222.07) | 14.66 (32.33) | 10.97 (20.70) | 6.44 (12.59) | 0.36 (1.42) | 20.44 (19.16) | 26.28 (24.00) | 35.24 (64.59) | 0.22 (0.26) | 0.62 (1.47) | |
| >0.92 | 0.58 (0.85) | 1.83 (2.69) | 136.30 (166.54) | 22.16 (45.93) | 16.70 (36.52) | 10.24 (24.05) | 0.58 (1.14) | 20.14 (23.58) | 26.82 (29.39) | 48.07 (37.71) | 0.27 (0.24) | 0.85 (1.75) | ||
| p-value | 0.443 | 0.494 | 0.548 | 0.443 | 0.576 | 0.120 | 0.468 | 0.419 | 0.443 | 0.310 | 0.237 | 0.395 |
| Cases (n = 34) | Controls (n = 75) | p-Value | |
|---|---|---|---|
| n (%) | n (%) | ||
| Age (years) 1 | 35.9 ± 5.3 | 35.1 ± 8.2 | 0.967 |
| Height (m) 1 | 1.6 ± 0.1 | 1.6 ± 0.1 | 0.365 |
| Weight (kg) 1 | 69.5 ± 15.7 | 65.7 ± 13.1 | 0.600 |
| Body mass index (kg/m2) 1 | 25.9 ± 5.5 | 24.6 ± 4.8 | 0.288 |
| Normal weight (BMI < 25) | 18 (52.9) | 46 (61.3) | 0.166 |
| Overweight (BMI 25–30) | 7 (20.6) | 20 (26.7) | |
| Obese (BMI > 30) | 9 (26.5) | 9 (12.0) | |
| Residence | 0.152 | ||
| Rural | 10 (29.4) | 13 (17.3) | |
| Urban | 24 (70.6) | 62 (82.7) | |
| Educational level | 0.949 | ||
| Less than university degree | 22 (64.7) | 49 (65.3) | |
| University degree | 12 (35.3) | 26 (34.7) | |
| Working outside home | 0.766 | ||
| Yes | 24 (70.6) | 55 (73.3) | |
| No | 10 (29.4) | 20 (26.7) | |
| Current Smoker | 0.549 | ||
| Yes | 24 (70.6) | 57 (76.0) | |
| No | 10 (29.4) | 18 (24.0) | |
| Parity | 0.593 | ||
| Nulliparous | 14 (41.2) | 35 (46.7) | |
| Primiparous/multiparous | 20 (58.8) | 40 (53.3) | |
| Intensity of menstrual bleeding | 0.542 | ||
| Mild | 11 (32.4) | 20 (26.7) | |
| Moderate/Severe | 23 (67.6) | 55 (73.3) | |
| rASRM endometriosis classification | - | ||
| I | 10 (29.4) | - | |
| II | 12 (35.3) | - | |
| III | 8 (23.5) | - | |
| IV | 4 (11.8) | - |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Martín-Leyva, A.; Peinado, F.M.; Ocón-Hernández, O.; Olivas-Martínez, A.; Luque, A.; León, J.; Lendínez, I.; Cardona, J.; Lara-Ramos, A.; Olea, N.; et al. Environmental Exposure to Persistent Organic Pollutants and Its Association with Endometriosis Risk: Implications in the Epithelial–Mesenchymal Transition Process. Int. J. Mol. Sci. 2024, 25, 4420. https://doi.org/10.3390/ijms25084420
Martín-Leyva A, Peinado FM, Ocón-Hernández O, Olivas-Martínez A, Luque A, León J, Lendínez I, Cardona J, Lara-Ramos A, Olea N, et al. Environmental Exposure to Persistent Organic Pollutants and Its Association with Endometriosis Risk: Implications in the Epithelial–Mesenchymal Transition Process. International Journal of Molecular Sciences. 2024; 25(8):4420. https://doi.org/10.3390/ijms25084420
Chicago/Turabian StyleMartín-Leyva, Ana, Francisco M. Peinado, Olga Ocón-Hernández, Alicia Olivas-Martínez, Antonio Luque, Josefa León, Inmaculada Lendínez, Jesús Cardona, Ana Lara-Ramos, Nicolás Olea, and et al. 2024. "Environmental Exposure to Persistent Organic Pollutants and Its Association with Endometriosis Risk: Implications in the Epithelial–Mesenchymal Transition Process" International Journal of Molecular Sciences 25, no. 8: 4420. https://doi.org/10.3390/ijms25084420
APA StyleMartín-Leyva, A., Peinado, F. M., Ocón-Hernández, O., Olivas-Martínez, A., Luque, A., León, J., Lendínez, I., Cardona, J., Lara-Ramos, A., Olea, N., Fernández, M. F., & Artacho-Cordón, F. (2024). Environmental Exposure to Persistent Organic Pollutants and Its Association with Endometriosis Risk: Implications in the Epithelial–Mesenchymal Transition Process. International Journal of Molecular Sciences, 25(8), 4420. https://doi.org/10.3390/ijms25084420