
CANT1 Is Involved in Collagen Fibrogenesis in Tendons by Regulating the Synthesis of Dermatan/Chondroitin Sulfate Attached to the Decorin Core Protein

 , , , and
, , , and
Abstract
1. Introduction
2. Results
2.1. Cant1−/− Mice Exhibit Tendon Hypoplasia During Postnatal Development
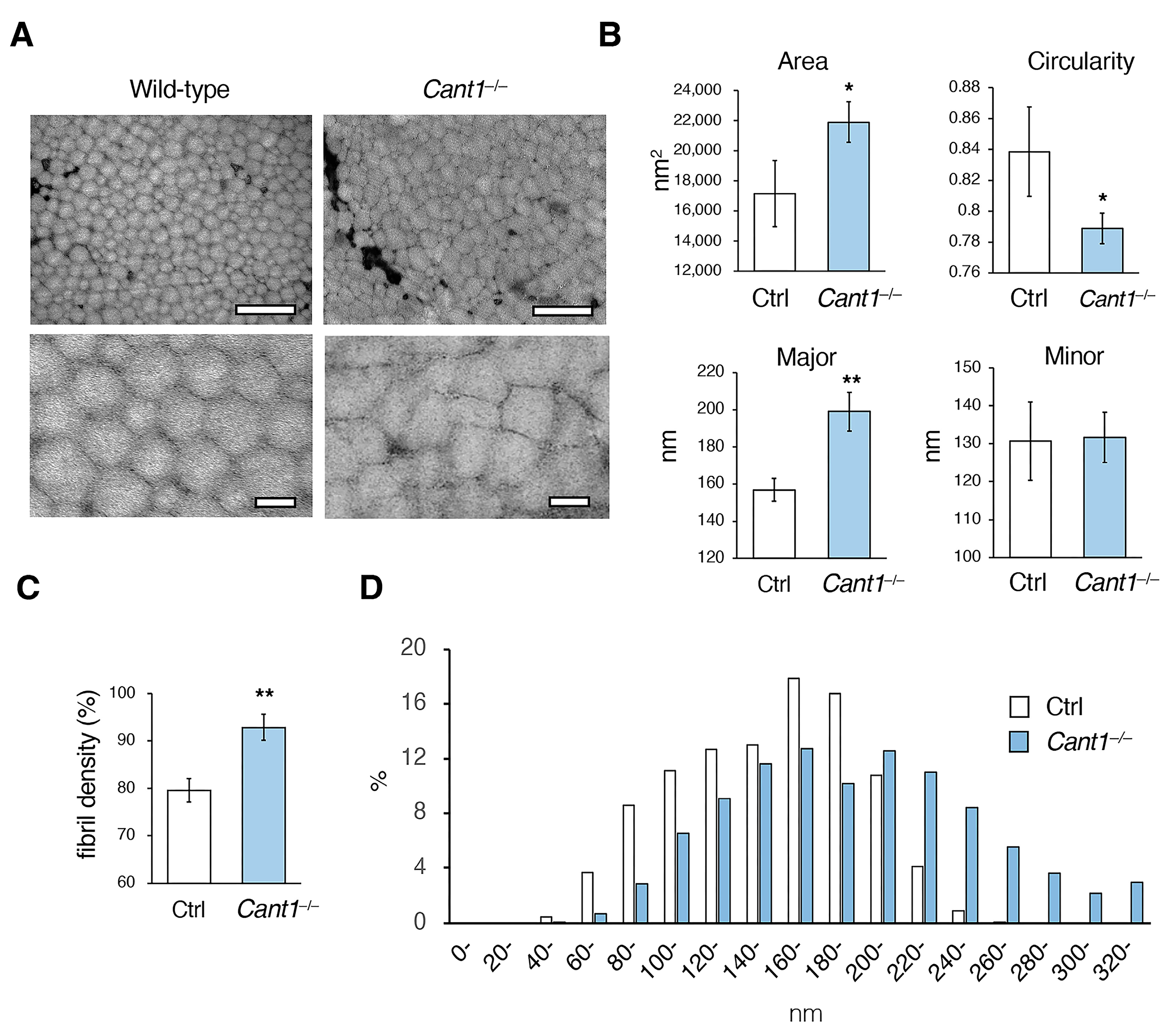
2.2. Fibrillogenesis Is Impaired in the Cant1−/− Tendons
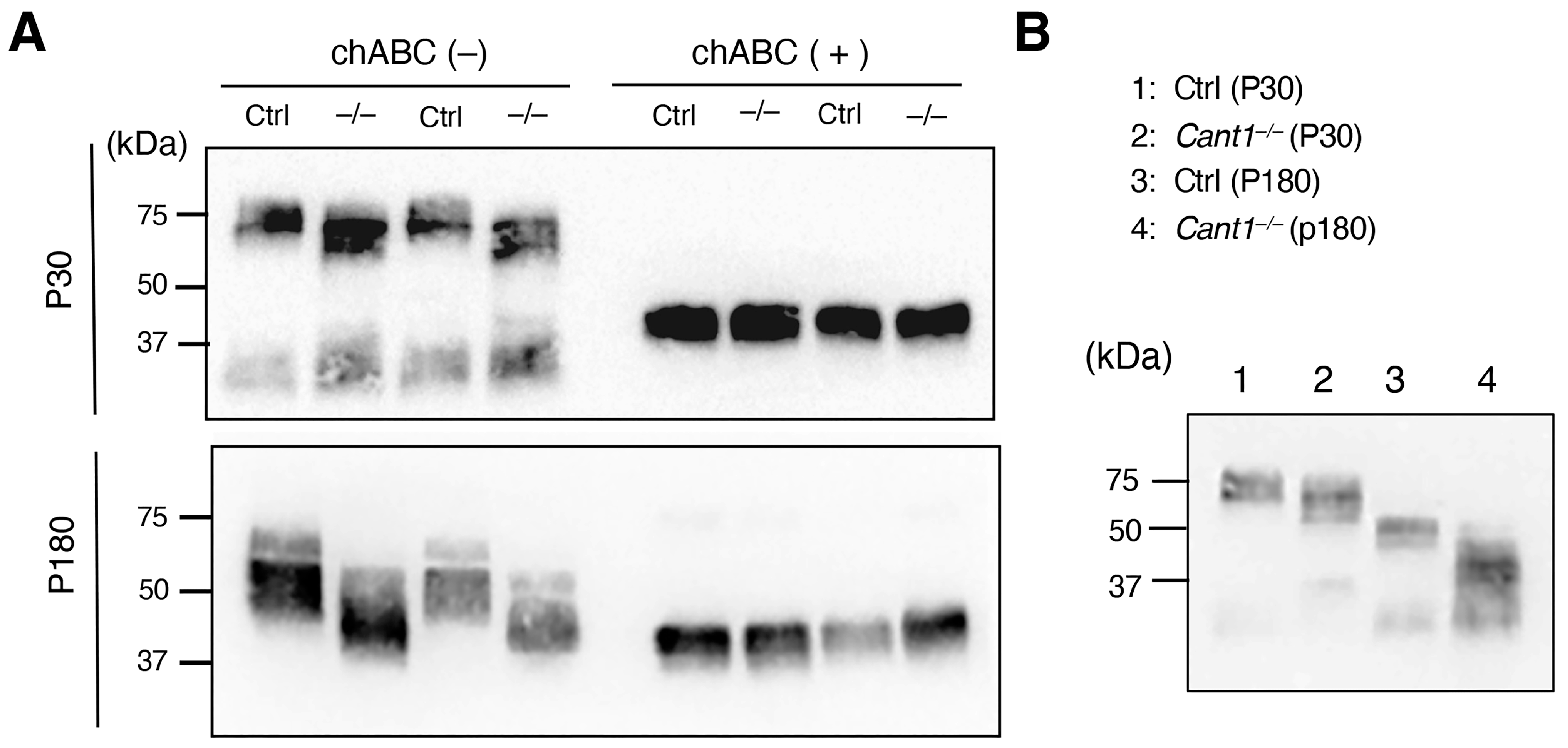
2.3. DS/CS Content Is Markedly Reduced, and the Molecular Weight of Decorin Is Correspondingly Decreased in the Cant1−/− Tendons
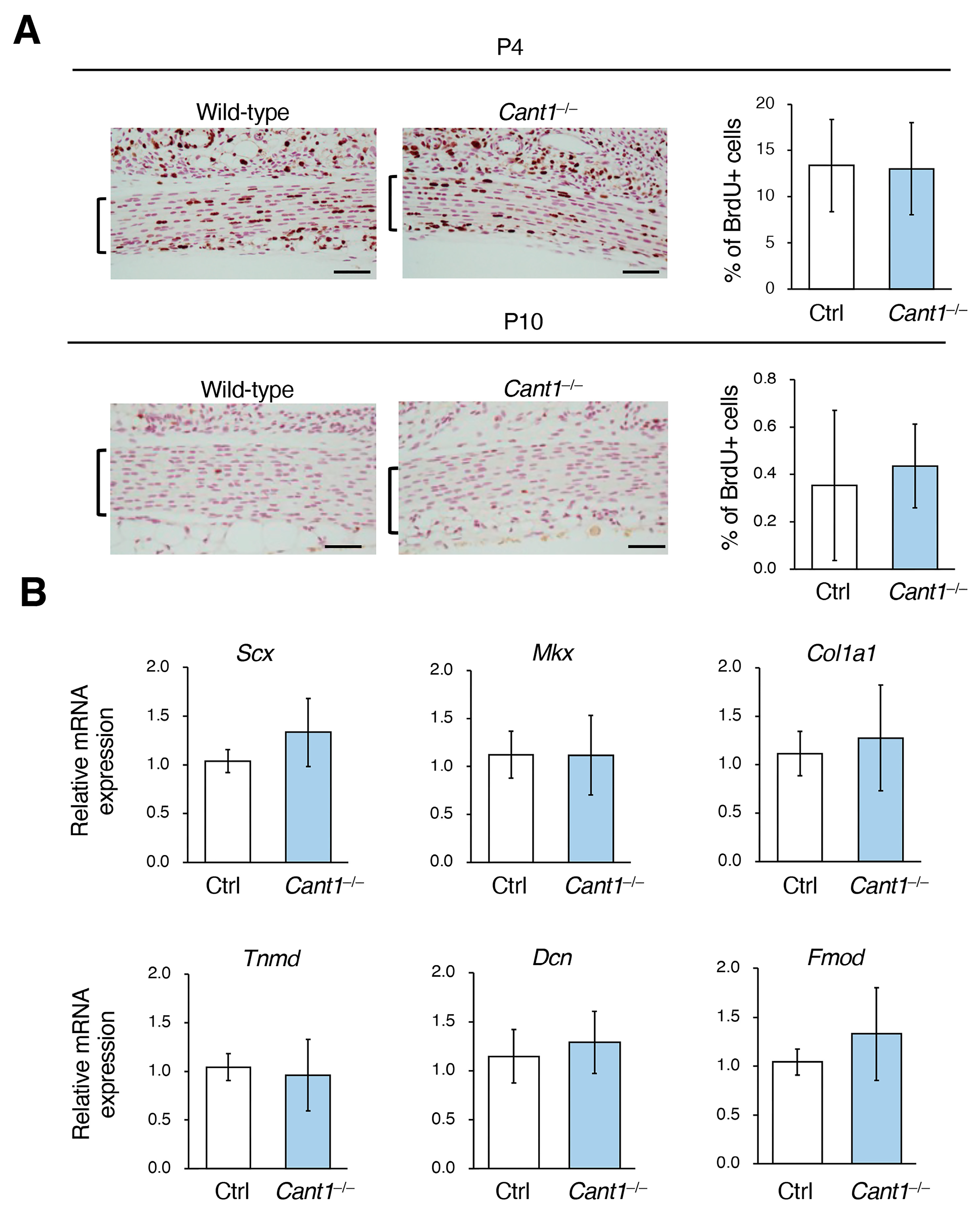
2.4. Proliferation and Differentiation Are Not Affected in the Cant1−/− Tendon Cells
3. Discussion
4. Materials and Methods
4.1. Animals
4.2. Macroscopic and Histological Analyses
4.3. Transmission Electron Microscopy (TEM)
4.4. Tensile Testing
4.5. Disaccharide Composition Analysis of the GAGs
4.6. Western Blot Analysis
4.7. Real-Time PCR
4.8. Statistical Analyses
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Kannus, P. Structure of the tendon connective tissue. Scand. J. Med. Sci. Sports 2000, 10, 312–320. [Google Scholar] [CrossRef] [PubMed]
- Birk, D.E.; Trelstad, R.L. Extracellular compartments in tendon morphogenesis: Collagen fibril, bundle, and macroaggregate formation. J. Cell Biol. 1986, 103, 231–240. [Google Scholar] [CrossRef] [PubMed]
- Zhang, G.; Young, B.B.; Ezura, Y.; Favata, M.; Soslowsky, L.J.; Chakravarti, S.; Birk, D.E. Development of tendon structure and function: Regulation of collagen fibrillogenesis. J. Musculoskelet. Neuronal Interact. 2005, 5, 5–21. [Google Scholar] [PubMed]
- Eisner, L.E.; Rosario, R.; Andarawis-Puri, N.; Arruda, E.M. The Role of the Non-collagenous extracellular matrix in tendon and ligament mechanical behavior: A Review. J. Biomech. Eng. 2022, 144, 050801. [Google Scholar] [CrossRef]
- Chen, S.; Birk, D.E. The regulatory roles of small leucine-rich proteoglycans in extracellular matrix assembly. FEBS J. 2013, 280, 2120–2137. [Google Scholar] [CrossRef]
- Matsushima, N.; Miyashita, H.; Kretsinger, R.H. Sequence features, structure, ligand interaction, and diseases in small leucine rich repeat proteoglycans. J. Cell Commun. Signal 2021, 15, 519–531. [Google Scholar] [CrossRef]
- Sugahara, K.; Kitagawa, H. Recent advances in the study of the biosynthesis and functions of sulfated glycosaminoglycans. Curr. Opin. Struct. Biol. 2000, 10, 518–527. [Google Scholar] [CrossRef]
- Mizumoto, S.; Sugahara, K. Glycosaminoglycan chain analysis and characterization (glycosylation/epimerization). Methods Mol. Biol. 2012, 836, 99–115. [Google Scholar]
- Kresse, H.; Schönherr, E. Proteoglycans of the extracellular matrix and growth control. J. Cell. Physiol. 2001, 189, 266–274. [Google Scholar] [CrossRef]
- Taipale, J.; Keski-Oja, J. Growth factors in the extracellular matrix. FEBS J. 1997, 11, 51–59. [Google Scholar] [CrossRef]
- Stanley, P. Golgi glycosylation. Cold Spring Harb. Perspect. Biol. 2011, 3, a005199. [Google Scholar] [CrossRef] [PubMed]
- Spiro, R.G. Protein glycosylation: Nature, distribution, enzymatic formation, and disease implications of glycopeptide bonds. Glycobiology 2002, 12, 43r–56r. [Google Scholar] [CrossRef] [PubMed]
- Parker, J.L.; Newstead, S. Gateway to the Golgi: Molecular mechanisms of nucleotide sugar transporters. Curr. Opin. Struct. Biol. 2019, 57, 127–134. [Google Scholar] [CrossRef] [PubMed]
- Ishida, N.; Kawakita, M. Molecular physiology and pathology of the nucleotide sugar transporter family (SLC35). Pflugers. Arch. 2004, 447, 768–775. [Google Scholar] [CrossRef]
- Maszczak-Seneczko, D.; Wiktor, M.; Skurska, E.; Wiertelak, W.; Olczak, M. Delivery of Nucleotide Sugars to the Mammalian Golgi: A Very Well (un)Explained Story. Int. J. Mol. Sci. 2022, 23, 8648. [Google Scholar] [CrossRef]
- Brandan, E.; Fleischer, B. Orientation and role of nucleosidediphosphatase and 5’-nucleotidase in Golgi vesicles from rat liver. Biochemistry 1982, 21, 4640–4645. [Google Scholar] [CrossRef]
- Kuhn, N.J.; White, A. The role of nucleoside diphosphatase in a uridine nucleotide cycle associated with lactose synthesis in rat mammary-gland Golgi apparatus. Biochem. J. 1977, 168, 423–433. [Google Scholar] [CrossRef]
- Smith, T.M.; Hicks-Berger, C.A.; Kim, S.; Kirley, T.L. Cloning, expression, and characterization of a soluble calcium-activated nucleotidase, a human enzyme belonging to a new family of extracellular nucleotidases. Arch. Biochem. Biophys. 2002, 406, 105–115. [Google Scholar] [CrossRef]
- Failer, B.U.; Braun, N.; Zimmermann, H. Cloning, expression, and functional characterization of a Ca(2+)-dependent endoplasmic reticulum nucleoside diphosphatase. J. Biol. Chem. 2002, 277, 36978–36986. [Google Scholar] [CrossRef]
- Murphy, D.M.; Ivanenkov, V.V.; Kirley, T.L. Bacterial expression and characterization of a novel, soluble, calcium-binding, and calcium-activated human nucleotidase. Biochemistry 2003, 42, 2412–2421. [Google Scholar] [CrossRef]
- Faivre, L.; Le Merrer, M.; Zerres, K.; Ben Hariz, M.; Scheffer, D.; Young, I.D.; Maroteaux, P.; Munnich, A.; Cormier-Daire, V. Clinical and genetic heterogeneity in Desbuquois dysplasia. Am. J. Med. Genet. A 2004, 128, 29–32. [Google Scholar] [CrossRef] [PubMed]
- Faivre, L.; Cormier-Daire, V.; Eliott, A.M.; Field, F.; Munnich, A.; Maroteaux, P.; Le Merrer, M.; Lachman, R. Desbuquois dysplasia, a reevaluation with abnormal and “normal” hands: Radiographic manifestations. Am. J. Med. Genet. A 2004, 124, 48–53. [Google Scholar] [CrossRef] [PubMed]
- Shohat, M.; Lachman, R.; Gruber, H.E.; Hsia, Y.E.; Golbus, M.S.; Witt, D.R.; Bodell, A.; Bryke, C.R.; Hogge, W.A.; Rimoin, D.L. Desbuquois syndrome: Clinical, radiographic, and morphologic characterization. Am. J. Med. Genet. 1994, 52, 9–18. [Google Scholar] [CrossRef] [PubMed]
- Piwar, H.; Ordak, M.; Bujalska-Zadrozny, M. Clinical and Genetic Insights into Desbuquois Dysplasia: Review of 111 Case Reports. Int. J. Mol. Sci. 2024, 25, 9700. [Google Scholar] [CrossRef]
- Kim, O.H.; Nishimura, G.; Song, H.R.; Matsui, Y.; Sakazume, S.; Yamada, M.; Narumi, Y.; Alanay, Y.; Unger, S.; Cho, T.J.; et al. A variant of Desbuquois dysplasia characterized by advanced carpal bone age, short metacarpals, and elongated phalanges: Report of seven cases. Am. J. Med. Genet. A 2010, 152, 875–885. [Google Scholar] [CrossRef]
- Furuichi, T.; Dai, J.; Cho, T.J.; Sakazume, S.; Ikema, M.; Matsui, Y.; Baynam, G.; Nagai, T.; Miyake, N.; Matsumoto, N.; et al. CANT1 mutation is also responsible for Desbuquois dysplasia, type 2 and Kim variant. J. Med. Genet. 2011, 48, 32–37. [Google Scholar] [CrossRef]
- Huber, C.; Oules, B.; Bertoli, M.; Chami, M.; Fradin, M.; Alanay, Y.; Al-Gazali, L.I.; Ausems, M.G.; Bitoun, P.; Cavalcanti, D.P.; et al. Identification of CANT1 mutations in Desbuquois dysplasia. Am. J. Hum. Genet. 2009, 85, 706–710. [Google Scholar] [CrossRef]
- Nizon, M.; Huber, C.; De Leonardis, F.; Merrina, R.; Forlino, A.; Fradin, M.; Tuysuz, B.; Abu-Libdeh, B.Y.; Alanay, Y.; Albrecht, B.; et al. Further delineation of CANT1 phenotypic spectrum and demonstration of its role in proteoglycan synthesis. Hum. Mutat. 2012, 33, 1261–1266. [Google Scholar] [CrossRef]
- Bui, C.; Huber, C.; Tuysuz, B.; Alanay, Y.; Bole-Feysot, C.; Leroy, J.G.; Mortier, G.; Nitschke, P.; Munnich, A.; Cormier-Daire, V. XYLT1 mutations in Desbuquois dysplasia type 2. Am. J. Hum. Genet. 2014, 94, 405–414. [Google Scholar] [CrossRef]
- Kodama, K.; Takahashi, H.; Oiji, N.; Nakano, K.; Okamura, T.; Niimi, K.; Takahashi, E.; Guo, L.; Ikegawa, S.; Furuichi, T. CANT1 deficiency in a mouse model of Desbuquois dysplasia impairs glycosaminoglycan synthesis and chondrocyte differentiation in growth plate cartilage. FEBS Open Bio 2020, 10, 1096–1103. [Google Scholar] [CrossRef]
- Paganini, C.; Monti, L.; Costantini, R.; Besio, R.; Lecci, S.; Biggiogera, M.; Tian, K.; Schwartz, J.M.; Huber, C.; Cormier-Daire, V.; et al. Calcium activated nucleotidase 1 (CANT1) is critical for glycosaminoglycan biosynthesis in cartilage and endochondral ossification. Matrix Biol. 2019, 81, 70–90. [Google Scholar] [CrossRef] [PubMed]
- Taieb, M.; Ghannoum, D.; Barré, L.; Ouzzine, M. Xylosyltransferase I mediates the synthesis of proteoglycans with long glycosaminoglycan chains and controls chondrocyte hypertrophy and collagen fibers organization of in the growth plate. Cell Death Dis. 2023, 14, 355. [Google Scholar] [CrossRef]
- Mis, E.K.; Liem, K.F., Jr.; Kong, Y.; Schwartz, N.B.; Domowicz, M.; Weatherbee, S.D. Forward genetics defines Xylt1 as a key, conserved regulator of early chondrocyte maturation and skeletal length. Dev. Biol. 2014, 385, 67–82. [Google Scholar] [CrossRef] [PubMed]
- Unger, S.; Ferreira, C.R.; Mortier, G.R.; Ali, H.; Bertola, D.R.; Calder, A.; Cohn, D.H.; Cormier-Daire, V.; Girisha, K.M.; Hall, C.; et al. Nosology of genetic skeletal disorders: 2023 revision. Am. J. Med. Genet. A 2023, 191, 1164–1209. [Google Scholar] [CrossRef] [PubMed]
- Screen, H.R.; Berk, D.E.; Kadler, K.E.; Ramirez, F.; Young, M.F. Tendon functional extracellular matrix. J. Orthop. Res. 2015, 33, 793–799. [Google Scholar] [CrossRef]
- Zhang, G.; Ezura, Y.; Chervoneva, I.; Robinson, P.S.; Beason, D.P.; Carine, E.T.; Soslowsky, L.J.; Iozzo, R.V.; Birk, D.E. Decorin regulates assembly of collagen fibrils and acquisition of biomechanical properties during tendon development. J. Cell. Biochem. 2006, 98, 1436–1449. [Google Scholar] [CrossRef]
- Danielson, K.G.; Baribault, H.; Holmes, D.F.; Graham, H.; Kadler, K.E.; Iozzo, R.V. Targeted disruption of decorin leads to abnormal collagen fibril morphology and skin fragility. J. Cell Biol. 1997, 136, 729–743. [Google Scholar] [CrossRef]
- Birk, D.E.; Nurminskaya, M.V.; Zycband, E.I. Collagen fibrillogenesis in situ: Fibril segments undergo post-depositional modifications resulting in linear and lateral growth during matrix development. Dev. Dyn. 1995, 202, 229–243. [Google Scholar] [CrossRef]
- Juneja, S.C.; Veillette, C. Defects in tendon, ligament, and enthesis in response to genetic alterations in key proteoglycans and glycoproteins: A review. Arthritis 2013, 2013, 154812. [Google Scholar] [CrossRef]
- Pacheco, B.; Malmström, A.; Maccarana, M. Two dermatan sulfate epimerases form iduronic acid domains in dermatan sulfate. J. Biol. Chem. 2009, 284, 9788–9795. [Google Scholar] [CrossRef]
- Maccarana, M.; Olander, B.; Malmström, J.; Tiedemann, K.; Aebersold, R.; Lindahl, U.; Li, J.P.; Malmström, A. Biosynthesis of dermatan sulfate: Chondroitin-glucuronate C5-epimerase is identical to SART2. J. Biol. Chem. 2006, 281, 11560–11568. [Google Scholar] [CrossRef] [PubMed]
- Müller, T.; Mizumoto, S.; Suresh, I.; Komatsu, Y.; Vodopiutz, J.; Dundar, M.; Straub, V.; Lingenhel, A.; Melmer, A.; Lechner, S.; et al. Loss of dermatan sulfate epimerase (DSE) function results in musculocontractural Ehlers-Danlos syndrome. Hum. Mo.l Genet. 2013, 22, 3761–3772. [Google Scholar] [CrossRef] [PubMed]
- Miyake, N.; Kosho, T.; Mizumoto, S.; Furuichi, T.; Hatamochi, A.; Nagashima, Y.; Arai, E.; Takahashi, K.; Kawamura, R.; Wakui, K.; et al. Loss-of-function mutations of CHST14 in a new type of Ehlers-Danlos syndrome. Hum. Mutat. 2010, 31, 966–974. [Google Scholar] [CrossRef]
- Dündar, M.; Müller, T.; Zhang, Q.; Pan, J.; Steinmann, B.; Vodopiutz, J.; Gruber, R.; Sonoda, T.; Krabichler, B.; Utermann, G.; et al. Loss of dermatan-4-sulfotransferase 1 function results in adducted thumb-clubfoot syndrome. Am. J. Hum. Genet. 2009, 85, 873–882. [Google Scholar] [CrossRef] [PubMed]
- Maccarana, M.; Kalamajski, S.; Kongsgaard, M.; Magnusson, S.P.; Oldberg, A.; Malmström, A. Dermatan sulfate epimerase 1-deficient mice have reduced content and changed distribution of iduronic acids in dermatan sulfate and an altered collagen structure in skin. Mol. Cell Biol. 2009, 29, 5517–5528. [Google Scholar] [CrossRef]
- Hirose, T.; Takahashi, N.; Tangkawattana, P.; Minaguchi, J.; Mizumoto, S.; Yamada, S.; Miyake, N.; Hayashi, S.; Hatamochi, A.; Nakayama, J.; et al. Structural alteration of glycosaminoglycan side chains and spatial disorganization of collagen networks in the skin of patients with mcEDS-CHST14. Biochim. Biophys. Acta Gen. Subj. 2019, 1863, 623–631. [Google Scholar] [CrossRef]
- Hirose, T.; Mizumoto, S.; Hashimoto, A.; Takahashi, Y.; Yoshizawa, T.; Nitahara-Kasahara, Y.; Takahashi, N.; Nakayama, J.; Takehana, K.; Okada, T.; et al. Systematic investigation of the skin in Chst14−/− mice: A model for skin fragility in musculocontractural Ehlers-Danlos syndrome caused by CHST14 variants (mcEDS-CHST14). Glycobiology 2021, 31, 137–150. [Google Scholar] [CrossRef]
- Robinson, P.S.; Huang, T.F.; Kazam, E.; Iozzo, R.V.; Birk, D.E.; Soslowsky, L.J. Influence of decorin and biglycan on mechanical properties of multiple tendons in knockout mice. J. Biomech. Eng. 2005, 127, 181–185. [Google Scholar] [CrossRef]
- Iozzo, R.V.; Moscatello, D.K.; McQuillan, D.J.; Eichstetter, I. Decorin is a biological ligand for the epidermal growth factor receptor. J. Biol. Chem. 1999, 274, 4489–4492. [Google Scholar] [CrossRef]
- Noble, N.A.; Harper, J.R.; Border, W.A. In vivo interactions of TGF-beta and extracellular matrix. Prog. Growth Factor Res. 1992, 4, 369–382. [Google Scholar] [CrossRef]
- Yamaguchi, Y.; Mann, D.M.; Ruoslahti, E. Negative regulation of transforming growth factor-beta by the proteoglycan decorin. Nature 1990, 346, 281–284. [Google Scholar] [CrossRef] [PubMed]
- Gallagher, J.T. Heparan sulfate: Growth control with a restricted sequence menu. J. Clin. Investig. 2001, 108, 357–361. [Google Scholar] [CrossRef] [PubMed]
- Knudson, C.B.; Knudson, W. Cartilage proteoglycans. Semin. Cell Dev. Biol. 2001, 12, 69–78. [Google Scholar] [CrossRef] [PubMed]
- Kiani, C.; Chen, L.; Wu, Y.J.; Yee, A.J.; Yang, B.B. Structure and function of aggrecan. Cell Res. 2002, 12, 19–32. [Google Scholar] [CrossRef]
- Calì, T.; Fedrizzi, L.; Ottolini, D.; Gomez-Villafuertes, R.; Mellström, B.; Naranjo, J.R.; Carafoli, E.; Brini, M. Ca2+-activated nucleotidase 1, a novel target gene for the transcriptional repressor DREAM (downstream regulatory element antagonist modulator), is involved in protein folding and degradation. J. Biol. Chem. 2012, 287, 18478–18491. [Google Scholar] [CrossRef]
- Harden, T.K.; Sesma, J.I.; Fricks, I.P.; Lazarowski, E.R. Signalling and pharmacological properties of the P2Y receptor. Acta Physiol. 2010, 199, 149–160. [Google Scholar] [CrossRef]
- Liu, T.; Li, Z.Z.; Sun, L.; Yang, K.; Chen, J.M.; Han, X.Y.; Qi, L.M.; Zhou, X.G.; Wang, P. Upregulated CANT1 is correlated with poor prognosis in hepatocellular carcinoma. BMC Cancer 2023, 23, 1007. [Google Scholar] [CrossRef]
- Yao, Q.; Yu, Y.; Wang, Z.; Zhang, M.; Ma, J.; Wu, Y.; Zheng, Q.; Li, J. CANT1 serves as a potential prognostic factor for lung adenocarcinoma and promotes cell proliferation and invasion in vitro. BMC Cancer 2022, 22, 117. [Google Scholar] [CrossRef]
- Marzin, P.; Cormier-Daire, V. New perspectives on the treatment of skeletal dysplasia. Ther. Adv. Endocrinol. Metab. 2020, 11, 2042018820904016. [Google Scholar] [CrossRef]
- Sabir, A.; Irving, M. Clinical trials in skeletal dysplasia: A paradigm for treating rare diseases. Br. Med. Bull. 2021, 139, 16–35. [Google Scholar] [CrossRef]



{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| GAG type | GAG level (pmol disaccharide/μg protein) | Cant1−/−/Ctrl ratio | p-Value | |
|---|---|---|---|---|
| Ctrl | Cant1−/− | |||
| CS/DS | 194 ± 32.7 | 59.4 ± 7.0 | 0.31 | 0.00020 |
| CS | 84.4 ± 26.0 | 47.4 ± 13.8 | 0.56 | 0.045 |
| DS | 94.2 ± 17.2 | 11.9 ± 6.7 | 0.13 | 0.00011 |
| HS | n.d. | n.d. | - | - |
| KS | n.d. | n.d. | - | - |
| HA | 13.4 ± 4.1 | 7.84 ± 1.7 | 0.59 | 0.048 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2025 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Yamashita, R.; Tsutsui, S.; Mizumoto, S.; Watanabe, T.; Yamamoto, N.; Nakano, K.; Yamada, S.; Okamura, T.; Furuichi, T. CANT1 Is Involved in Collagen Fibrogenesis in Tendons by Regulating the Synthesis of Dermatan/Chondroitin Sulfate Attached to the Decorin Core Protein. Int. J. Mol. Sci. 2025, 26, 2463. https://doi.org/10.3390/ijms26062463
Yamashita R, Tsutsui S, Mizumoto S, Watanabe T, Yamamoto N, Nakano K, Yamada S, Okamura T, Furuichi T. CANT1 Is Involved in Collagen Fibrogenesis in Tendons by Regulating the Synthesis of Dermatan/Chondroitin Sulfate Attached to the Decorin Core Protein. International Journal of Molecular Sciences. 2025; 26(6):2463. https://doi.org/10.3390/ijms26062463
Chicago/Turabian StyleYamashita, Rina, Saki Tsutsui, Shuji Mizumoto, Takafumi Watanabe, Noritaka Yamamoto, Kenta Nakano, Shuhei Yamada, Tadashi Okamura, and Tatsuya Furuichi. 2025. "CANT1 Is Involved in Collagen Fibrogenesis in Tendons by Regulating the Synthesis of Dermatan/Chondroitin Sulfate Attached to the Decorin Core Protein" International Journal of Molecular Sciences 26, no. 6: 2463. https://doi.org/10.3390/ijms26062463
APA StyleYamashita, R., Tsutsui, S., Mizumoto, S., Watanabe, T., Yamamoto, N., Nakano, K., Yamada, S., Okamura, T., & Furuichi, T. (2025). CANT1 Is Involved in Collagen Fibrogenesis in Tendons by Regulating the Synthesis of Dermatan/Chondroitin Sulfate Attached to the Decorin Core Protein. International Journal of Molecular Sciences, 26(6), 2463. https://doi.org/10.3390/ijms26062463