Seasonal Physiological Parameters and Phytotelmata Bacterial Diversity of Two Bromeliad Species (Aechmea gamosepala and Vriesea platynema) from the Atlantic Forest of Southern Brazil

 , ,
, , 
Abstract
:1. Introduction
2. Materials and Methods
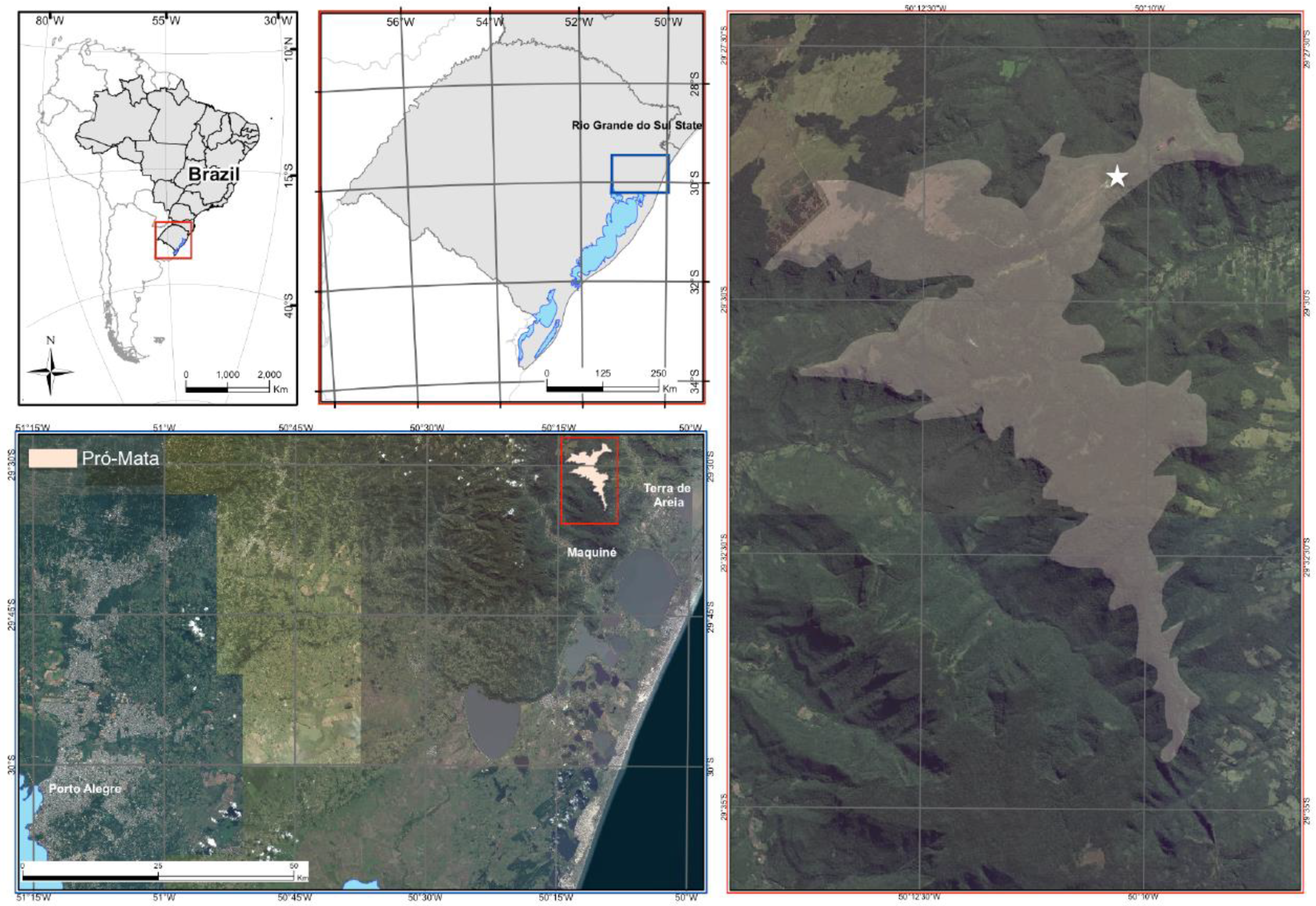
2.1. Sample Collection
2.2. Plant Nutritional Parameters
2.3. Analysis of Phytotelmata Microbial Communities
3. Results
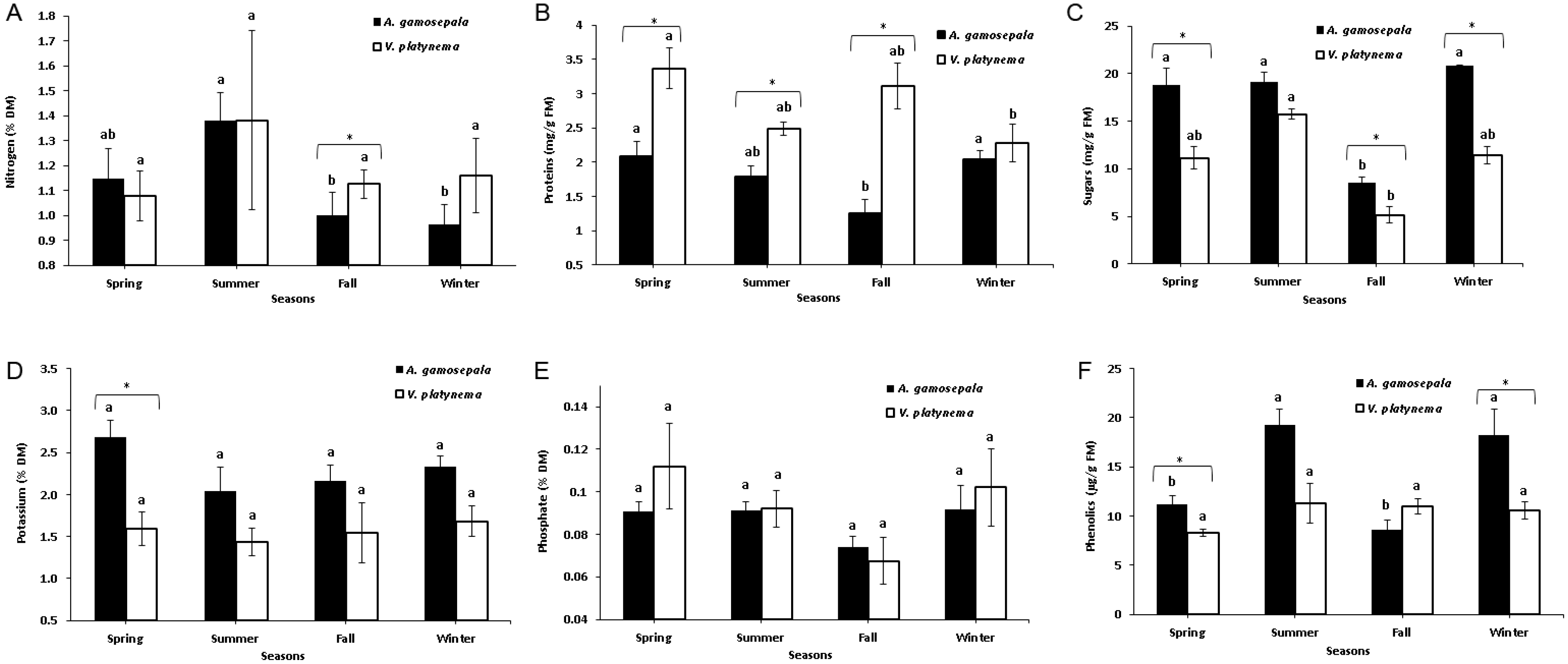
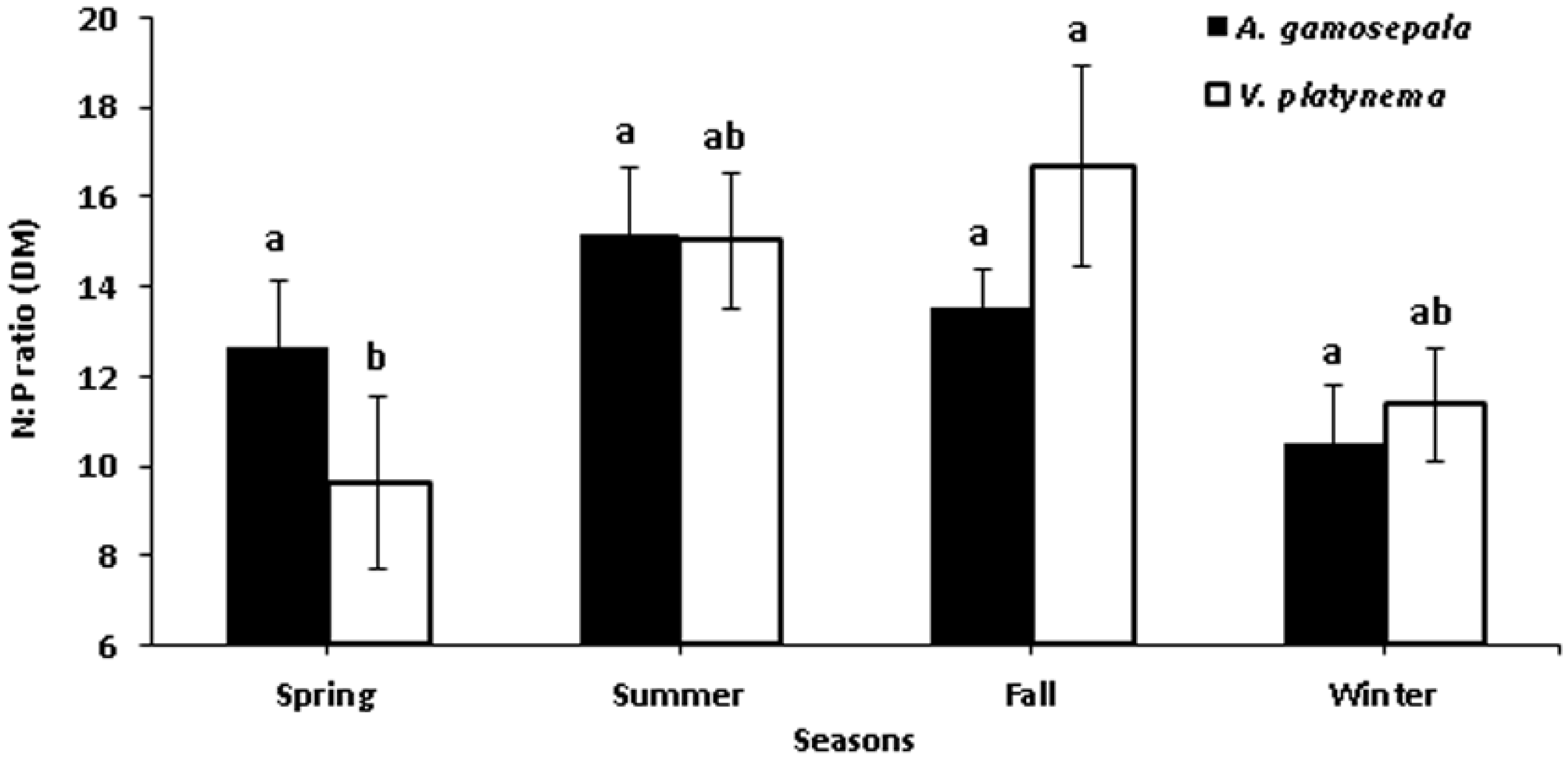
3.1. Physiological Parameters of V. platynema and A. gamosepala
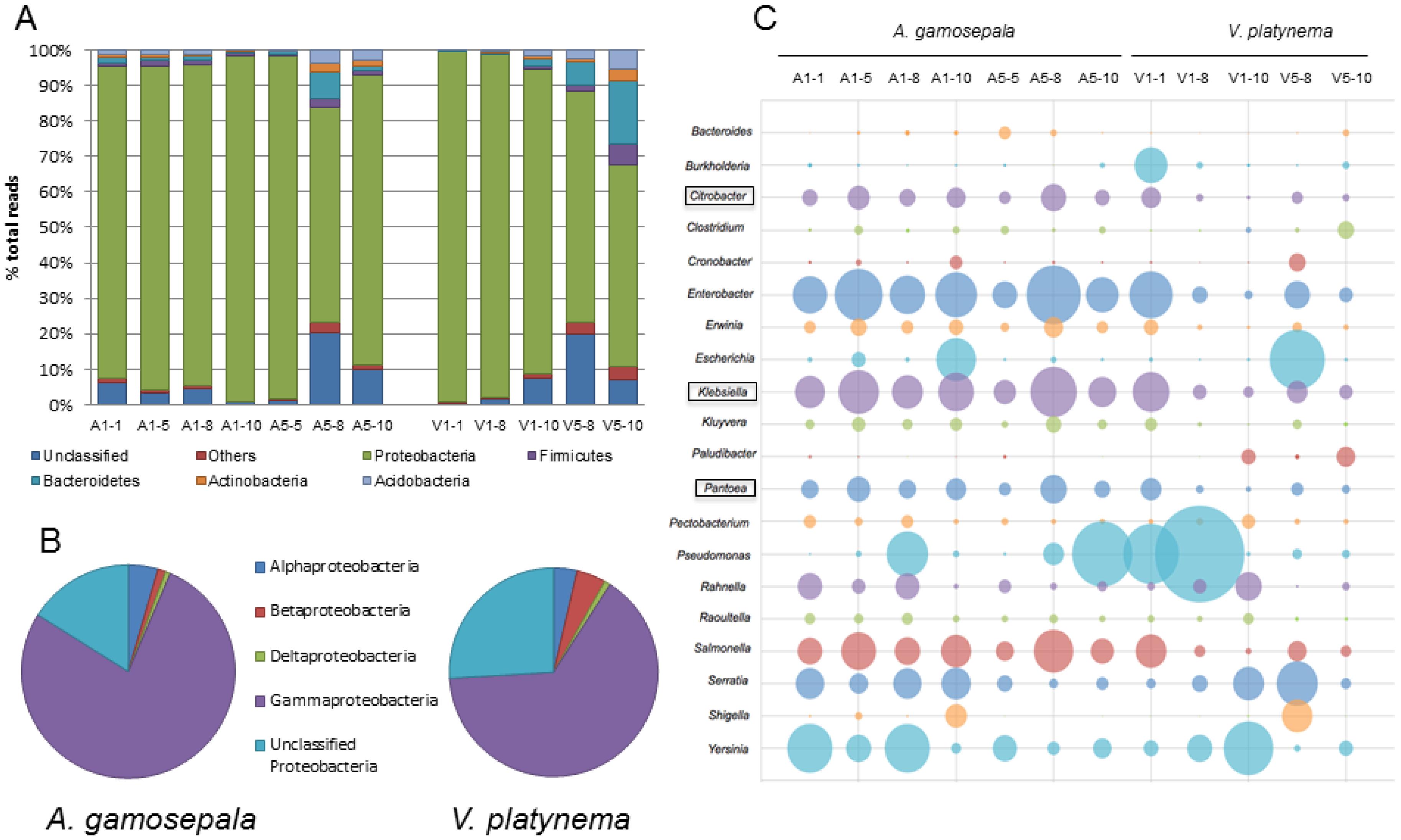
3.2. Microbial DNA Sequencing Data
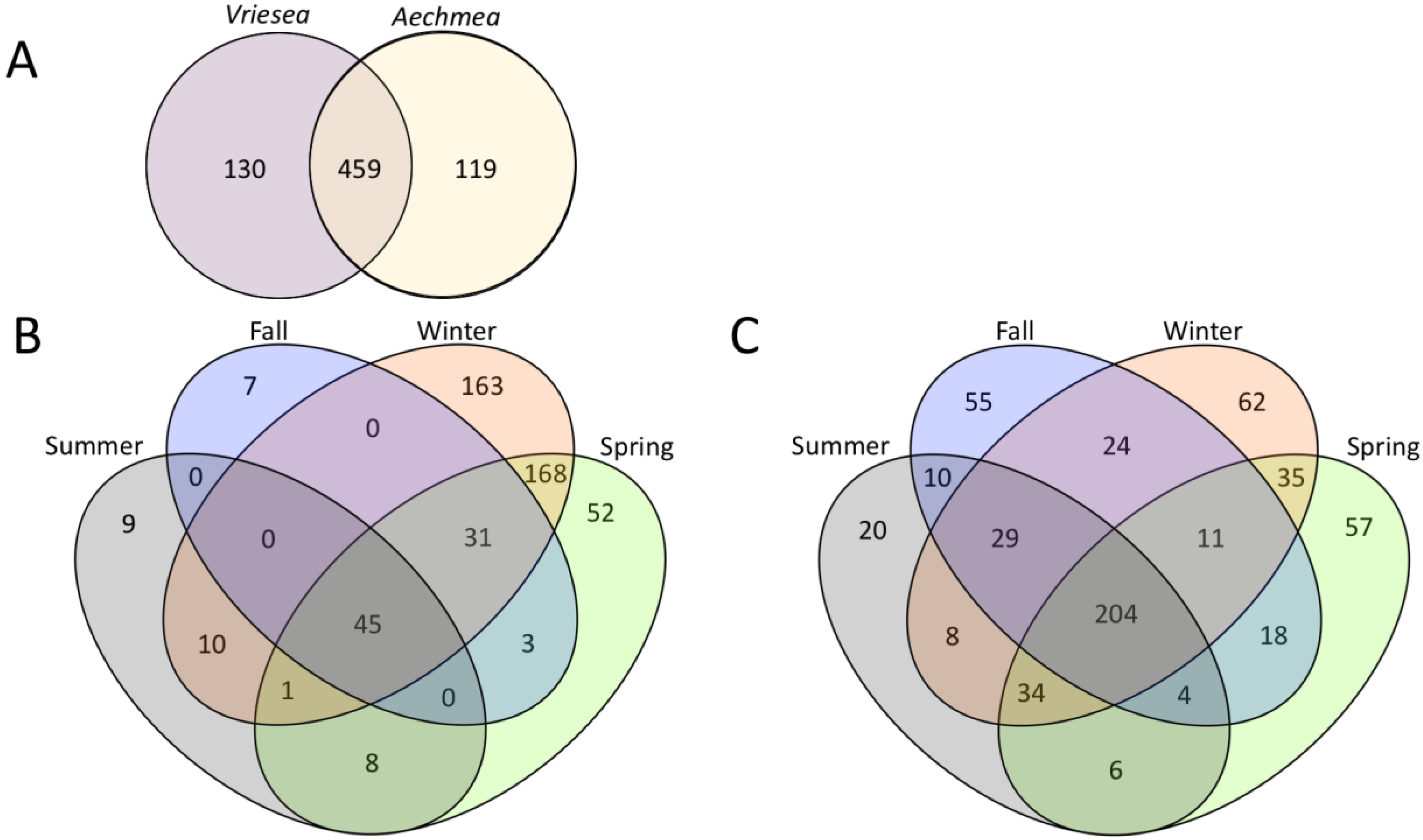
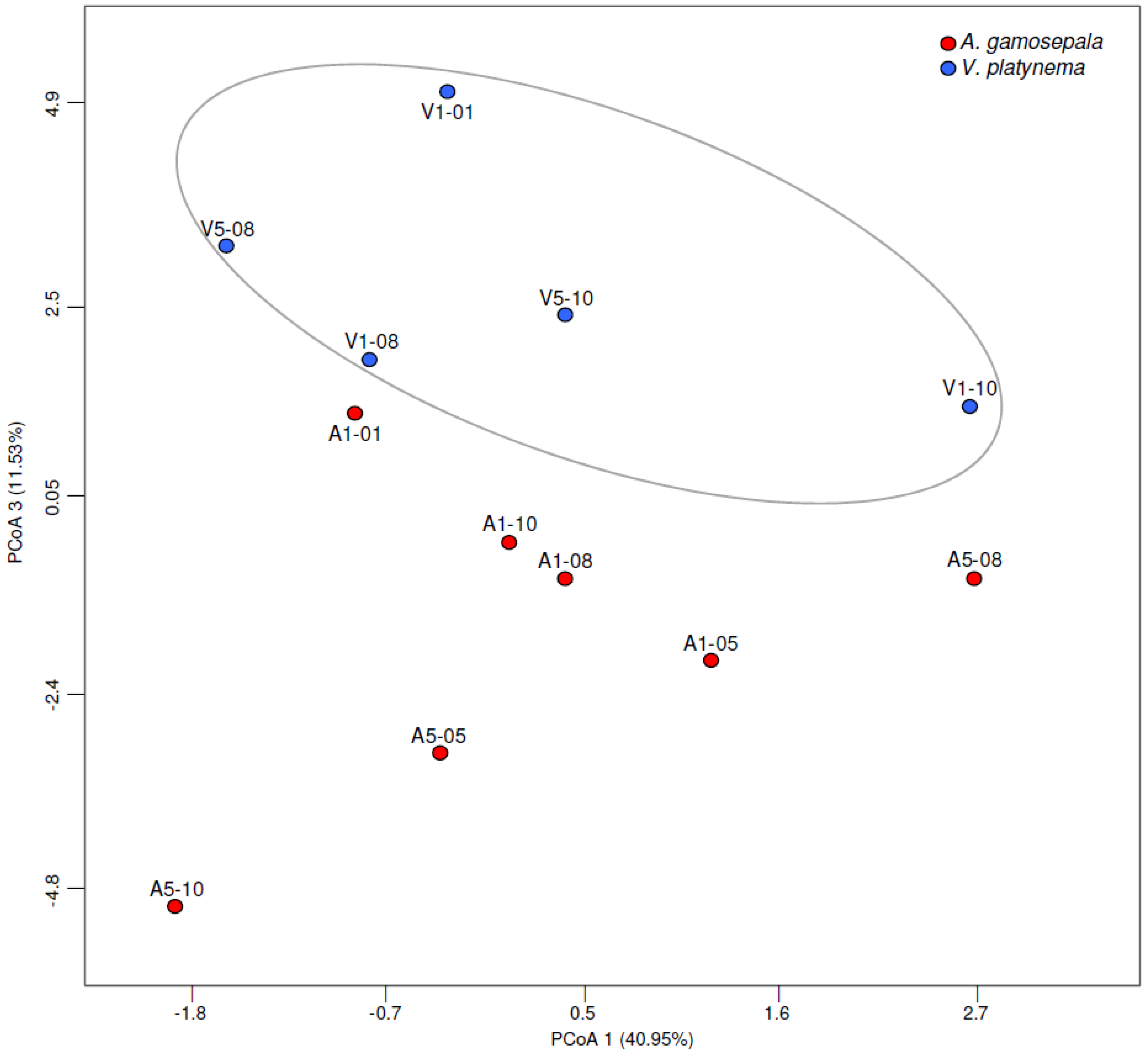
3.3. Differences in the Microbial Communities of A. gamosepala and V. platynema
4. Discussion
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Benzing, D.H.; Renfrow, A. The Mineral Nutrition of Bromeliaceae. Bot. Gaz. 1974, 135, 281–288. [Google Scholar] [CrossRef]
- Stuntz, S.; Zotz, G. Photosynthesis in vascular epiphytes—A survey of diverse taxonomical origin. Flora 2001, 196, 132–141. [Google Scholar] [CrossRef]
- Haubrich, C.S.; Pires, A.P.F.; Esteves, F.A.; Farjalla, V.F. Bottom-up regulation of bacterial growth in tropical phytotelm bromeliads. Hydrobiologia 2009, 632, 347–353. [Google Scholar] [CrossRef]
- Wanek, W.; Zotz, G. Are vascular epiphytes nitrogen or phosphorus limited? A study of plant 15N fractionation and foliar N:P stoichiometry with the tank bromeliad Vriesea sanguinolenta. New Phytol. 2011, 192, 462–470. [Google Scholar] [CrossRef] [PubMed]
- Medina, E.; Ziegler, H.; Luttge, U.; Trimborn, P.; Francisco, M. Light conditions during growth as reveled by 13C values of leaves of primitive cultivars of Ananas comosus, an obligate CAM species. Funct. Ecol. 1994, 8, 298–305. [Google Scholar] [CrossRef]
- Pop, M.; Janett, H.-P.; Luttge, U.; Medina, E. Metabolite gradients and carbohydrate translocation in rosette leaves of CAM and C3 bromeliads. New Phytol. 2003, 157, 649–656. [Google Scholar] [CrossRef]
- Crayn, D.M.; Winter, K.; Smith, J.A.C. Multiple origins of crassulacean acid metabolism and the epiphytic habit in the neotropical family Bromeliaceae. Proc. Natl. Acad. Sci. USA 2004, 101, 3703–3708. [Google Scholar] [CrossRef]
- Silvera, K.; Lasso, E. Ecophysiology and crassulacean acid metabolism of tropical epiphytes. In Tropical Tree Physiology: Adaptations and Responses in a Changing Environment; Goldstein, G., Santiago, L.S., Eds.; Springer: New York, NY, USA, 2016; pp. 25–43. [Google Scholar]
- Haslam, R.; Borland, A.; Maxswell, K.; Griffiths, H. Physiological responses of the CAM epiphyte Tillandsia usneoides L. (Bromeliaceae) to variations in light and water supply. J. Plant Physiol. 2003, 160, 627–634. [Google Scholar] [CrossRef]
- Benzing, D.H. Vascular Epiphytes. General Biology and Related Biota; Cambridge University Press: Cambridge, UK, 1990; p. 350. [Google Scholar]
- Wake, D.B. Adaptive radiation of salamanders in middle American cloud forests. Ann. Mo. Bot. Gard. 1987, 74, 242–264. [Google Scholar] [CrossRef]
- Laessle, A.M. A microlimnologicalstudy of Jamaican bromeliads. Ecology 1961, 24, 499–517. [Google Scholar] [CrossRef]
- Palacios-Vargas, J.G. Collembola asociados a Tillandsia (Bromeliaceae) en el Derrame Lavico del Chichinautzin, Morelos, Mexico. Southwest. Entomol. 1981, 6, 87–98. [Google Scholar]
- Fish, D. Phytotelmata: Terrestrial plants as hosts for aquatic insect communities. In Phytotelmata: Flora and Fauna; Frank, J.H., Lounibos, L.P., Eds.; Plexus: Medford, NJ, USA, 1983. [Google Scholar]
- Carrias, J.F.; Cussac, M.E.; Corbara, B. A preliminary study of freshwater protozoa in tank bromeliads. J. Trop. Ecol. 2001, 17, 611–617. [Google Scholar] [CrossRef]
- Landell, M.F.; Billodre, R.; Ramos, J.P.; Leoncine, O.; Vainstein, M.H.; Valente, P. Candida aechmeae sp. nov. and Candida vrieseae sp. nov., novel yeast species isolated from the phylloplane of bromeliads in Southern Brazil. Int. J. Syst. Evol. Microbiol. 2010, 60, 244–248. [Google Scholar] [CrossRef] [PubMed]
- Brouard, O.; Le Jeune, A.H.; Leroy, C.; Cereghino, R.; Roux, O.; Pelozuelo, L.; Dejean, A.; Corbara, B.; Carrias, J.F. Are algae relevant to the detritus-based food web in tank-bromeliads? PLoS ONE 2011, 6, e20129. [Google Scholar] [CrossRef]
- Simão, T.L.L.; Giongo-Borges, A.; Gano, K.A.; Davis-Richardson, A.G.; Brown, C.T.; Fagen, J.R.; Triplett, E.W.; Dias, R.; Mondin, C.A.; da Silva, R.M.; et al. Characterization of ciliate diversity in bromeliad tank waters from the Brazilian Atlantic Forest. Eur. J. Protistol. 2017, 61, 359–365. [Google Scholar] [CrossRef] [PubMed]
- Brighigna, L.; Montaini, P.; Favilli, F.; Trejo, A.C. Role of the nitrogen-fixing bacterial microflora in the epiphytism of Tillandsia (Bromeliaceae). Am. J. Bot. 1992, 79, 723–727. [Google Scholar] [CrossRef]
- Tsoktouridis, G.; Mantell, S.; Bantinaki, E.; Thangavelu, M. DNA profiling and strategies for in planta localisation of bacteria intimately associated with Billbergia magnifica ssp. acutisepalia. Plant Cell Tissue Organ Cult. 1998, 52, 41–45. [Google Scholar] [CrossRef]
- Goffredi, S.K.; Kantor, A.H.; Woodside, W.T. Aquatic microbial habitats within a neotropical rainforest: Bromeliads and pH-associated trends in bacterial diversity and composition. Microb. Ecol. 2010, 61, 529–542. [Google Scholar] [CrossRef]
- Pittl, E.; Innerebner, G.; Wanek, W.; Insam, H. Microbial communities of arboreal and ground soils in the Esquinas rainforest, Costa Rica. Plant Soil 2011, 329, 65–74. [Google Scholar] [CrossRef]
- Goffredi, S.K.; Jang, G.E.; Woodside, W.T.; Ussler, W., III. Bromeliad catchments as habitats for methanogenesis in tropical rainforest canopies. Front. Microbiol. 2011, 2, 256. [Google Scholar] [CrossRef]
- Giongo, A.; Beneduzi, A.; Gano, K.A.; Vargas, L.K.; Utz, L.; Passaglia, L.M.P. Characterization of plant growth-promoting bacteria inhabiting Vriesea gigantea Gaud. and Tillandsia aeranthos (Loiseleur) L.B. Smith (Bromeliaceae). Biota Neotrop. 2013, 13, 80–85. [Google Scholar] [CrossRef]
- Brandt, F.B.; Martinson, G.O.; Pommerenke, B.; Pump, J.; Conrad, R. Drying on archaeal community composition and methanogenesis in bromeliad tanks. FEMS Microbiol. Ecol. 2015, 91, 1–10. [Google Scholar] [CrossRef] [PubMed]
- Goffredi, S.K.; Jang, G.E.; Haroon, M.F. Transcriptomics in the tropics: Total RNA-based profiling of Costa Rican bromeliad-associated communities. Comput. Struct. Biotechnol. J. 2015, 13, 18–23. [Google Scholar] [CrossRef] [PubMed]
- Louca, S.; Jacques, S.M.S.; Pires, A.P.F.; Leal, J.S.; Srivastava, D.S.; Parfrey, L.W.; Farjalla, V.F.; Doebeli, M. High taxonomic variability despite stable functional structure across microbial communities. Nat. Ecol. Evol. 2016. [Google Scholar] [CrossRef] [PubMed]
- Brandt, F.B.; Martinson, G.O.; Conrad, R. Bromeliad tanks are unique habitats for microbial communities involved in methane turnover. Plant Soil 2017, 410, 167–179. [Google Scholar] [CrossRef]
- Inselsbacher, E.; Cambui, C.A.; Richter, A.; Stange, C.F.; Mercier, H.; Wanek, W. Microbial activities and foliar uptake of nitrogen in the epiphytic bromeliad Vriesea gigantea. New Phytol. 2007, 175, 311–320. [Google Scholar] [CrossRef] [PubMed]
- Cambuí, C.A.; Gaspar, M.; Mercier, H. Detection of urease in the cell wall and membranes from leaf tissues of bromeliad species. Physiol. Plant. 2009, 136, 86–93. [Google Scholar] [CrossRef]
- Louca, S.; Jacques, S.M.S.; Pires, A.P.F.; Leal, J.S.; González, A.L.; Doebeli, M.; Farjalla, V.F. Functional structure of the bromeliad tank microbiome is strongly shaped by local geochemical conditions. Environ. Microbiol. 2017, 19, 3132–3151. [Google Scholar] [CrossRef]
- Xu, J. Microbial ecology in the age of genomics and metagenomics concepts, tools, and recent advances. Mol. Ecol. 2006, 15, 1713–1731. [Google Scholar] [CrossRef]
- Mota, F.S. Estudos do clima do estado do Rio Grande do Sul, segundo o sistema de W. Koppen. Rev. Bras. Geogr. 1951, 13, 275–284. [Google Scholar]
- Bertoletti, J.J.; Teixeira, M.B. Centro de Pesquisas e Conservação da Natureza Pró-Mata. Termo de Referência. Div. Mus. Ciên. Tec. UBEA/PUCRS 1995, 2, 1–47. [Google Scholar]
- Allen, S.E. Chemical Analyses of Ecological Materials; Blackwell Scientific: Oxford, UK, 1974; p. 565. [Google Scholar]
- Tedesco, M.J.; Gianello, C.; Bissani, C.A.; Bohnen, H.; Volkweiss, S.J. Análise de Solo, Plantas e Outros Materiais, 2nd ed.; Universidade Federal do Rio Grande do Sul: Porto Alegre, Brazil, 1995; p. 174. [Google Scholar]
- Güsewell, S.; Koerselman, W. Variation in nitrogen and phosphorus concentrations of wetland plants. Perspect. Plant Ecol. Evol. Syst. 2002, 5, 37–61. [Google Scholar] [CrossRef]
- Yemm, E.W.; Willis, A.J. The estimation of carbohydrates in plant extracts by anthrone. Biochem. J. 1954, 57, 508–514. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Savio, L.E.B.; Astarita, L.V.; Santarém, E.L. Secondary metabolism in micropropagated Hypericum perforatum L. grown in non-aerated liquid. Plant Cell Tissue Organ Cult. 2012, 108, 465–472. [Google Scholar] [CrossRef]
- Bradford, M.M. A rapid sensitive method for the quantification of microgram quantities of protein utilising the principle of protein-Dye Binding. Anal. Biochem. 1976, 72, 248–254. [Google Scholar] [CrossRef]
- Bates, S.; Berg-Lyons, D.; Caporaso, J.G.; Walters, W.A.; Knight, R.; Fierer, N. Examining the global distribution of dominant archaeal populations in soil. ISME J. 2011, 5, 908–917. [Google Scholar] [CrossRef] [PubMed]
- Fagen, J.R.; Giongo, A.; Brown, C.T.; davis-Richardson, A.G.; Gano, K.A.; Triplett, E.W. Characterization of the Relative Abundance of the Citrus Pathogen Ca. Liberibacter asiaticus in the Microbiome of Its Insect Vector, Diaphorina citri, using High Throughput 16S rRNA Sequencing. Open Microbiol. J. 2012, 6, 29–33. [Google Scholar] [CrossRef] [PubMed]
- Huang, X.; Wang, J.; Aluru, S.; Yang, S.-P.; Hillier, L.D. PCAP: A Whole-Genome Assembly Program. Genome Res. 2003, 13, 2164–2170. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Giongo, A.; Crabb, D.B.; Davis-Richardson, A.G.; Chauliac, D.; Moberley, J.M.; Gano, K.A.; Mukherjee, N.; Casella, G.; Roesh, L.F.W.; Brandon, W.; et al. PANGEA: Pipeline for Analysis of Next GEneration Amplicons. ISME J. 2010, 4, 852–861. [Google Scholar] [CrossRef]
- Cole, J.R.; Chai, B.; Farris, R.J.; Wang, Q.; Kulam, S.A.; McGarrel, D.M.; Garrit, G.M.; Tiedje, J.M. The Ribosomal Database Project (RDP-II): Sequences and tools for high-throughput rRNA analysis. Nucleic Acids Res. 2005, 33, D294–D296. [Google Scholar] [CrossRef]
- Giongo, A.; Davis-Richardson, A.G.; Crabb, D.B.; Triplett, E.W. TaxCollector: Tools to modify existing 16S rRNA databases for the rapid classification at six taxonomic levels. Diversity 2010, 23, 1015–1025. [Google Scholar] [CrossRef]
- Hong, S.H.; Bunge, J.; Jeon, S.O.; Epistein, S.S. Predicting microbial species richness. Proc. Natl. Acad. Sci. USA 2006, 103, 117–122. [Google Scholar] [CrossRef] [PubMed]
- Meyer, F.; Paarmann, D.; D’Souza, M.; Olson, R.; Glass, E.M.; Paczian, T.; Rodriguez, A.; Stevens, R.; Wilke, A.; Wilkening, J.; et al. The metagenomics RAST server—A public resource for the automatic phylogenetic and functional analysis of metagenomes. BCM Bioinform. 2008, 9, 386. [Google Scholar] [CrossRef] [PubMed]
- Griffiths, H. Carbon dioxide concentration mechanisms and the evolution of CAM in vascular epiphytes. In Vascular Plants as Epiphytes Evolution and Ecophysiology; Lüttge, U., Ed.; Springer: Berlin, Germany, 1989; pp. 42–86. [Google Scholar]
- Pierce, S.; Winter, K.; Griffiths, H. The role of CAM in high rainfall cloud forests: An in situ comparison of photosynthetic pathways in Bromeliaceae. Plant Cell Environ. 2002, 25, 1181–1189. [Google Scholar] [CrossRef]
- Takahashi, C.A.; Mercier, H. Nitrogen metabolism in leaves of a tank epiphytic bromeliad: Characterization of a spatial and functional division. J. Plant Physiol. 2011, 11, 1208–1216. [Google Scholar] [CrossRef] [PubMed]
- Aerts, R.; Chapin, F.S. The mineral nutrition of wild plants revisited: A re-evaluation of processes and patterns. In Advances in Ecological Research; Academic Press: Cambridge, MA, USA, 2000; Volume 30, pp. 1–67. [Google Scholar]
- Winkler, U.; Zotz, G. Highly efficient uptake of phosphorus in epiphytic bromeliads. Ann. Bot. 2009, 103, 477–484. [Google Scholar] [CrossRef] [PubMed]
- Chapin, F.S. The mineral nutrition of wild plants. Ann. Rev. Ecol. Syst. 1980, 11, 233–260. [Google Scholar] [CrossRef]
- Cardelus, C.L.; Mack, M.C.; Woods, C.; DeMarco, J.; Treseder, K.K. The influence of tree species on canopy soil nutrient status in a tropical lowland wet forest in Costa Rica. Plant Soil 2009, 318, 47–61. [Google Scholar] [CrossRef]
- Wassen, M.J.; Venterink, H.O.; Lapshina, E.D.; Tanneberger, F. Endangered plants persist under phosphorus limitation. Nature 2005, 437, 547–550. [Google Scholar] [CrossRef]
- Koerselman, W.; Meuleman, A.F.M. The vegetation N:P ratio: A new tool to detect the nature of nutrient limitation. J. Appl. Ecol. 1996, 33, 1441–1450. [Google Scholar] [CrossRef]
- Farjalla, V.F.; Srivastava, D.S.; Marino, N.A.; Azevedo, F.D.; Dib, V.; Lopes, P.M.; Rosado, A.S.; Bozelli, R.L.; Esteves, F.A. Ecological determinism increases with organism size. Ecology 2012, 93, 1752–1759. [Google Scholar] [CrossRef] [PubMed]
- Carmo, F.L.; Santos, H.F.; Peixoto, R.S.; Rosado, A.S.; Araujo, F.V. Tank bromeliad water: Similar or distinct environments for research of bacterial bioactives? Braz. J. Microbiol. 2014, 45, 185–192. [Google Scholar] [CrossRef] [PubMed]
- Pedraza, R.O. Recent advances in nitrogen-fixing acetic acid bacteria. Int. J. Food Microbiol. 2008, 125, 25–35. [Google Scholar] [CrossRef] [PubMed]
- Jain, A.; Das, S. Insight into the interaction between plants and associated fluorescent Pseudomonas spp. Int. J. Agron. 2016. [Google Scholar] [CrossRef]
- Bodenhausen, N.; Horton, M.W.; Bergelson, J. Bacterial communities associated with the leaves and the roots of Arabidopsis thaliana. PLoS ONE 2013, 8, e56329. [Google Scholar] [CrossRef] [PubMed]
- Hagler, A.N.; Rosa, C.A.; Morais, P.B.; Mendonça-Hagler, L.C.; Franco, G.M.; Araujo, F.V.; Soares, C.A. Yeasts and coliform bacteria of water accumulated in bromeliads of mangrove and sand dune ecosystems of southeast Brazil. Can. J. Microbiol. 1993, 39, 973–977. [Google Scholar] [CrossRef]
- Bashan, Y.; de-Bashan, L.E.; Prabhu, S.R.; Hernandez, J.P. Advances in plant growth-promoting bacterial inoculant technology: Formulations and practical perspectives (1998–2013). Plant Soil 2014, 378, 1–33. [Google Scholar] [CrossRef]
- Ramesh, A.; Sharma, S.K.; Sharma, M.P.; Yadav, N.; Om, J.P. Plant growth-promoting traits in Enterobacter cloacae subsp. dissolvens MDSR9 isolated from soybean rhizosphere and its impact on growth and nutrition of soybean and wheat upon inoculation. Agric. Res. 2014, 3, 53–66. [Google Scholar] [CrossRef]
- Martinson, G.O.; Werner, F.A.; Scherber, C.; Conrad, R.; Corre, M.D.; Flessa, H.; Wolf, K.; Gradstein, S.R.; Veldkamp, E. Methane emissions from tank bromeliads in neotropical forests. Nat. Geosci. 2010, 3, 766–769. [Google Scholar] [CrossRef]
- Jousset, A.; Bienhold, C.; Chatzinotas, A.; Gallien, L.; Gobet, A.; Kurm, V.; Rillig, M.C.; Rivett, D.W.; Salles, J.F.; van der Heijden, M.G.; et al. Where less may be more: How the rare biosphere pulls ecosystems strings. ISME J. 2017, 11, 853–862. [Google Scholar] [CrossRef]
- Ahemad, M.; Kibret, M. Mechanisms and applications of plant growth promoting rhizobacteria: Current perspective. J. King Saud Univ. Sci. 2014, 26, 1–20. [Google Scholar] [CrossRef] [Green Version]
- Sogin, M.L.; Morrison, H.G.; Huber, J.A.; Welch, M.D.; Huse, S.M.; Neal, P.R.; Arrieta, J.M.; Herndl, G.J. Microbial diversity in the deep sea and the underexplored “rare biosphere”. Proc. Natl. Acad. Sci. USA 2006, 103, 12115–12120. [Google Scholar] [CrossRef] [PubMed]
- Richardson, B.A. The bromeliad microcosm and the assessment of faunal diversity in a neotropical forest. Biotropica 1999, 31, 321–336. [Google Scholar] [CrossRef]
- Leroy, C.; Carrias, J.-F.; Céréghino, R.; Corbara, B. The contribution of microorganisms and metazoans to mineral nutrition in bromeliads. J. Plant Ecol. 2015, 9, 241–255. [Google Scholar] [CrossRef]
- Normand, P.; Benson, D.R.; Berry, A.M.; Tisa, L.S. The family Frankiaceae. In The Prokaryotes; Rosenberg, E., DeLong, E.F., Lory, S., Stackebrandt, E., Thompson, F., Eds.; Springer: New York, NY, USA, 2015; pp. 339–356. [Google Scholar]
- Lorenzo, N.; Mantuano, D.; Mantovani, A. Comparative leaf ecophysiology and anatomy of seedlings, young and adult individuals of the epiphytic aroid Anthurium scandens (Aubl.) Engl. Environ. Exp. Bot. 2010, 68, 314–322. [Google Scholar] [CrossRef]
- Mardegan, S.F.; Nardoto, G.B.; Higuchi, N.; Reinert, F.; Martinelli, L.A. Variation in nitrogen use strategies and photosynthetic pathways among vascular epiphytes in the Brazilian Central Amazon. Rev. Bras. Bot. 2011, 34, 21–30. [Google Scholar] [CrossRef] [Green Version]
- Silby, M.W.; Winstanley, C.; Godfrey, S.A.C.; Levy, S.B.; Jackson, R.W. Pseudomonas genome: Diverse and adaptable. FEMS Microbiol. Rev. 2011, 35, 652–680. [Google Scholar] [CrossRef]
- Fox, A.R.; Soto, G.; Valverde, C.; Russo, D.; Lagares, A., Jr.; Zorreguieta, A.; Alleva, K.; Pascuan, C.; Frare, R.; Mercado-Blanco, J.; et al. Major cereal crops benefit from biological nitrogen fixation when inoculated with the nitrogen-fixing bacterium Pseudomonas protegens Pf-5 X940. Environ. Microbiol. 2016, 18, 3522–3534. [Google Scholar] [CrossRef]
- Patten, C.L.; Jeong, H.; Blakney, A.J.; Wallace, N. Draft genome sequence of a diazotrophic, plant growth-promoting rhizobacterium of the Pseudomonas syringae complex. Genome Announc. 2016, 4, e01023-16. [Google Scholar] [CrossRef]
- Saha, C.; Mukherjee, G.; Agarwal-Banka, P.; Seal, A. A consortium of non-rhizobial endophytic microbes from Typha angustifolia functions as probiotic in rice and improves nitrogen metabolism. Plant Biol. 2016, 18, 938–946. [Google Scholar] [CrossRef]
- Coenye, T.; Vandamme, P. Diversity and significance of Burkholderia species occupying diverse ecological niches. Environ. Microbiol. 2003, 5, 719–729. [Google Scholar] [CrossRef] [PubMed]
- Cheynier, V.; Comte, G.; Davies, K.M.; Latanzio, V.; Martens, S. Plant phenolics: Recent advances on their biosynthesis, genetics, and ecophysiology. Plant Physiol. Biochem. 2013, 72, 1–20. [Google Scholar] [CrossRef] [PubMed]
- Sartor, T.; Xavier, V.B.; Falcão, M.A.; Mondin, C.A.; Dos Santos, M.A. Seasonal changes in phenolic compounds and in the biological activities of Baccharis dentata (Vell.) G.M. Barroso. Ind. Crops Prod. 2013, 1, 355–359. [Google Scholar] [CrossRef]
- Ambrosini, A.; Giongo, A.; Beneduzi, A.; Colbachini, N.; Friedisch, L.; Passaglia, L.M.P. LBactérias promotoras de crescimento vegetal em Vriesea gigantea Gaudchi. (Bromeliaceae). Rev. Bras. Biociências 2007, 5, 1169–1170. [Google Scholar]








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Bromeliad species | Plant ID | Season | Air Temperature (°C) | Plant Height at Tree Stem (m) | Water Tank | |||
|---|---|---|---|---|---|---|---|---|
| Sample | pH | Temperature (°C) | Volume Collected (mL) | |||||
| Aechmea gamosepala | A1 | A1-1 | Summer | 20 | 2.8 | 6.5 | 20 | 20 |
| A1 | A1-5 | Fall | 13 | 6 | 14 | 13 | ||
| A1 | A1-8 | Winter | 10 | 6 | 5 | 5 | ||
| A1 | A1-10 | Spring | 17 | 5.5 | 16 | 17 | ||
| A5 | A5-5 | Fall | 13 | 2.4 | 6 | 14 | 13 | |
| A5 | A5-8 | Winter | 10 | 6 | 5 | 5 | ||
| A5 | A5-10 | Spring | 17 | 5.5 | 16 | 17 | ||
| Vriesea platynema | V1 | V1-1 | Summer | 20 | 2.5 | 6.5 | 20 | 29 |
| V1 | V1-8 | Winter | 10 | 6 | 5 | 5 | ||
| V1 | V1-10 | Spring | 17 | 5.5 | 16 | 17 | ||
| V5 | V5-8 | Winter | 10 | 2.4 | 6 | 5 | 5 | |
| V5 | V5-10 | Spring | 17 | 5.5 | 16 | 17 | ||
| Seasons | Sites | A. gamosepala | V. platynema |
|---|---|---|---|
| Spring | 1 | 10.236 c | 11.399 a |
| 2 | 15.641 b | 9.673 a | |
| 3 | 18.324 a | 11.450 a | |
| 4 | 19.839 a | 6.910 a | |
| 5 | 19.424 a | 5.810 b | |
| Summer | 1 | 15.139 a | na |
| 2 | 16.02 a | 14.602 a | |
| 3 | na | na | |
| 4 | 18.887b | na | |
| 5 | na | 12.721 a | |
| Fall | 1 | 15.954 a | 1.380 c |
| 2 | 6.250 bc | 2.813 c | |
| 3 | 7.061 b | 2.558 c | |
| 4 | 4.819 c | 5.499 b | |
| 5 | 7.624 b | 12.099 a | |
| Winter | 1 | 16.208 c | 20.817 a |
| 2 | 19.143 b | 10.528 b | |
| 3 | 18.795 b | 6,928 c | |
| 4 | 18,569 b | 8.736 cd | |
| 5 | 22.813 a | 11.454 b |
| Specimen | Relative Abundance | p | ||||||
|---|---|---|---|---|---|---|---|---|
| A1-1 | A1-2 | A1-3 | A1-4 | A5-1 | A5-2 | A5-3 | ||
| Order | ||||||||
| Neisseriales | 0.01 | 0.01 | 0.03 | 0.00 | 0.14 | 0.00 | 0.01 | 0.05 |
| Families | ||||||||
| Acidimicrobiaceae | 0.0054 | 0.0044 | 0.0052 | 0 | 0.0058 | 0.0788 | 0.0064 | 0.031 |
| Clostridiales Family XI. Incertae Sedis | 0 | 0 | 0 | 0 | 0.0006 | 0.0043 | 0.0013 | 0.018 |
| Frankiaceae | 0.0014 | 0 | 0 | 0 | 0.0012 | 0.0064 | 0.0090 | 0.042 |
| Micrococcaceae | 0 | 0 | 0.0026 | 0 | 0.0175 | 0.0043 | 0.0141 | 0.028 |
| Nakamurellaceae | 0.0054 | 0 | 0.0052 | 0 | 0.0064 | 0.0383 | 0.0128 | 0.031 |
| Neisseriaceae | 0.0054 | 0.0088 | 0 | 0 | 0.0263 | 0.1427 | 0.0090 | 0.048 |
| Oxalobacteraceae | 0.0216 | 0.0263 | 0.0262 | 0.0016 | 0.0292 | 0.5026 | 0.1863 | 0.032 |
| Phyllobacteriaceae | 0.0027 | 0.0044 | 0.0026 | 0 | 0.0035 | 0.0639 | 0.0077 | 0.048 |
| Genera | ||||||||
| Acinetobacter | 0.0230 | 0.0482 | 0.0735 | 0.0377 | 0.0158 | 0.0085 | 0.0180 | 0.05 |
| Amycolatopsis | 0 | 0 | 0 | 0 | 0.0006 | 0.0021 | 0.0090 | 0.031 |
| Arthrobacter | 0 | 0 | 0.0026 | 0 | 0.0164 | 0.0043 | 0.0128 | 0.018 |
| Buttiauxella | 0.3636 | 0.6743 | 0.5116 | 0.7680 | 0.3640 | 0.2023 | 0.3238 | 0.05 |
| Candidatus Cuticobacterium | 0.0230 | 0.0350 | 0.0367 | 0.0442 | 0.0181 | 0.0106 | 0.0218 | 0.028 |
| Candidatus Nardonella | 0.0865 | 0.1708 | 0.1181 | 0.1818 | 0.0748 | 0.0596 | 0.0604 | 0.048 |
| Candidatus Stammerula | 0.1162 | 0.1314 | 0.1522 | 0.2243 | 0.0841 | 0.0511 | 0.0758 | 0.034 |
| Cronobacter * | 0.0784 | 0.2977 | 1.3799 | 0.1015 | 0.0631 | 0.0256 | 0.0398 | 0.034 |
| Escherichia * | 0.2054 | 1.7340 | 13.4552 | 0.3078 | 0.1928 | 0.0937 | 0.1028 | 0.034 |
| Massilia | 0.0014 | 0.0044 | 0.0026 | 0 | 0.0041 | 0.0383 | 0.0270 | 0.019 |
| Nakamurella | 0.0054 | 0 | 0.0052 | 0 | 0.0058 | 0.0277 | 0.0103 | 0.034 |
| Solibium | 0 | 0 | 0 | 0 | 0.0012 | 0.0043 | 0.0013 | 0.034 |
| Xanthomonas | 0.0014 | 0.0044 | 0.0052 | 0.0016 | 0 | 0 | 0.0013 | 0.032 |

© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Giongo, A.; Medina-Silva, R.; Astarita, L.V.; Borges, L.G.d.A.; Oliveira, R.R.; Simão, T.L.L.; Gano, K.A.; Davis-Richardson, A.G.; Brown, C.T.; Fagen, J.R.; et al. Seasonal Physiological Parameters and Phytotelmata Bacterial Diversity of Two Bromeliad Species (Aechmea gamosepala and Vriesea platynema) from the Atlantic Forest of Southern Brazil. Diversity 2019, 11, 111. https://doi.org/10.3390/d11070111
Giongo A, Medina-Silva R, Astarita LV, Borges LGdA, Oliveira RR, Simão TLL, Gano KA, Davis-Richardson AG, Brown CT, Fagen JR, et al. Seasonal Physiological Parameters and Phytotelmata Bacterial Diversity of Two Bromeliad Species (Aechmea gamosepala and Vriesea platynema) from the Atlantic Forest of Southern Brazil. Diversity. 2019; 11(7):111. https://doi.org/10.3390/d11070111
Chicago/Turabian StyleGiongo, Adriana, Renata Medina-Silva, Leandro V. Astarita, Luiz Gustavo dos A. Borges, Rafael R. Oliveira, Taiz L. L. Simão, Kelsey A. Gano, Austin G. Davis-Richardson, Christopher T. Brown, Jennie R. Fagen, and et al. 2019. "Seasonal Physiological Parameters and Phytotelmata Bacterial Diversity of Two Bromeliad Species (Aechmea gamosepala and Vriesea platynema) from the Atlantic Forest of Southern Brazil" Diversity 11, no. 7: 111. https://doi.org/10.3390/d11070111
APA StyleGiongo, A., Medina-Silva, R., Astarita, L. V., Borges, L. G. d. A., Oliveira, R. R., Simão, T. L. L., Gano, K. A., Davis-Richardson, A. G., Brown, C. T., Fagen, J. R., Arzivenco, P. M., Neto, C. P., Abichequer, A. D., Lindholz, C. G., Baptista-Silva, A., Mondin, C. A., Utz, L. R. P., Triplett, E. W., & Eizirik, E. (2019). Seasonal Physiological Parameters and Phytotelmata Bacterial Diversity of Two Bromeliad Species (Aechmea gamosepala and Vriesea platynema) from the Atlantic Forest of Southern Brazil. Diversity, 11(7), 111. https://doi.org/10.3390/d11070111