Limnological Characteristics and Diatom Dominants in Lakes of Northeastern Poland



Abstract
:1. Introduction
2. Materials and Methods
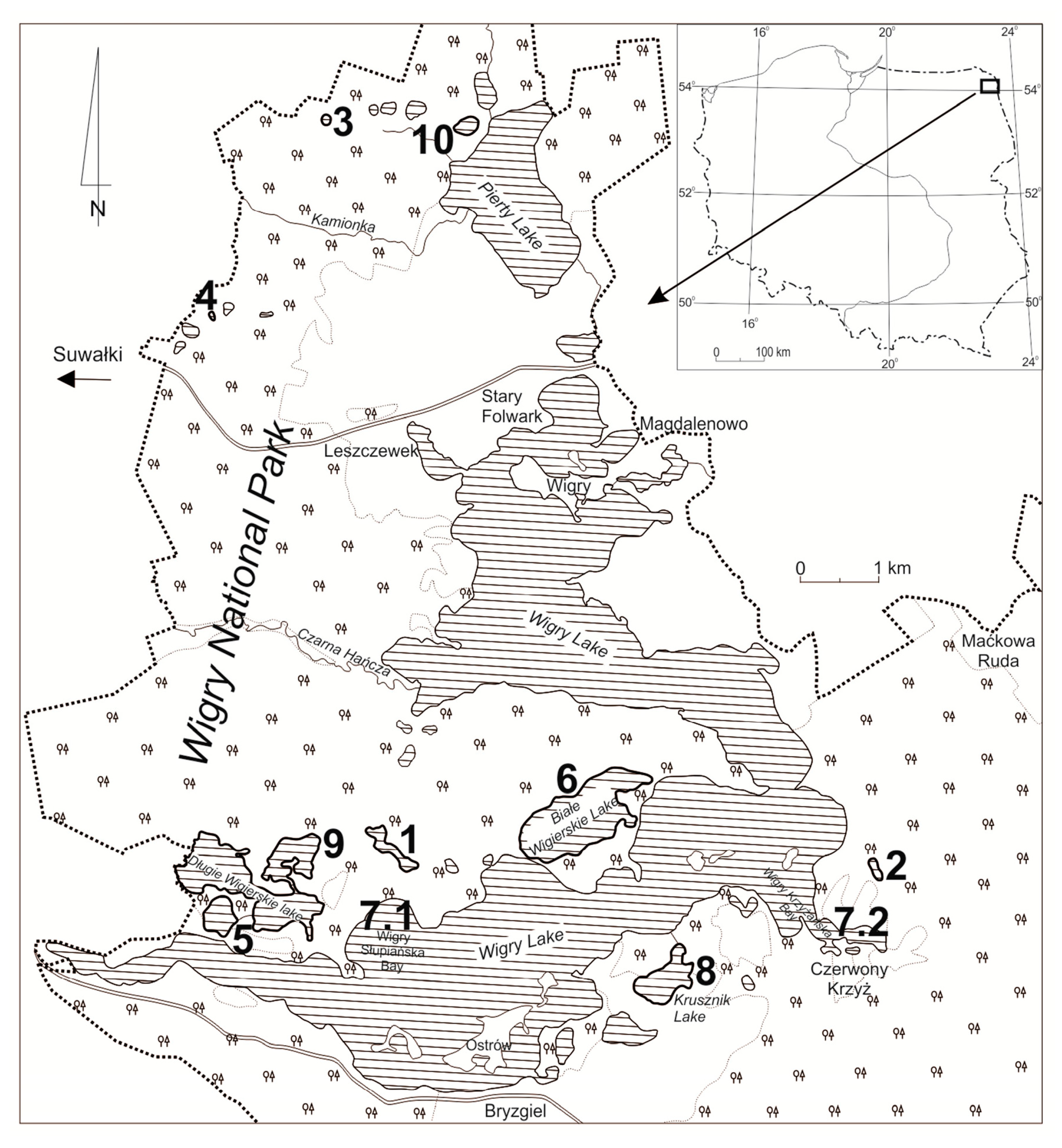
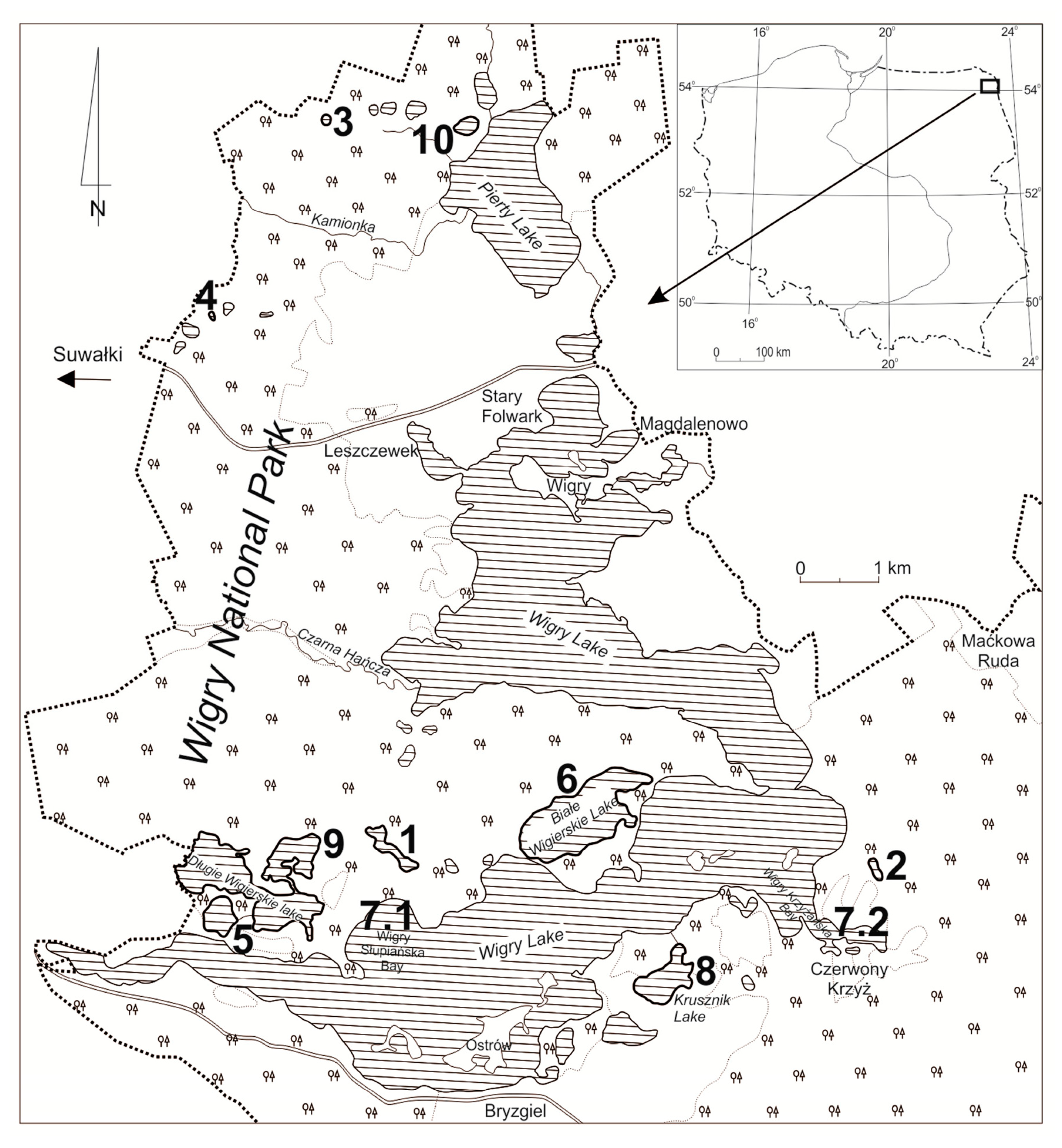
2.1. Study Area
2.2. Statistical Data
2.3. Diatom Data
3. Results
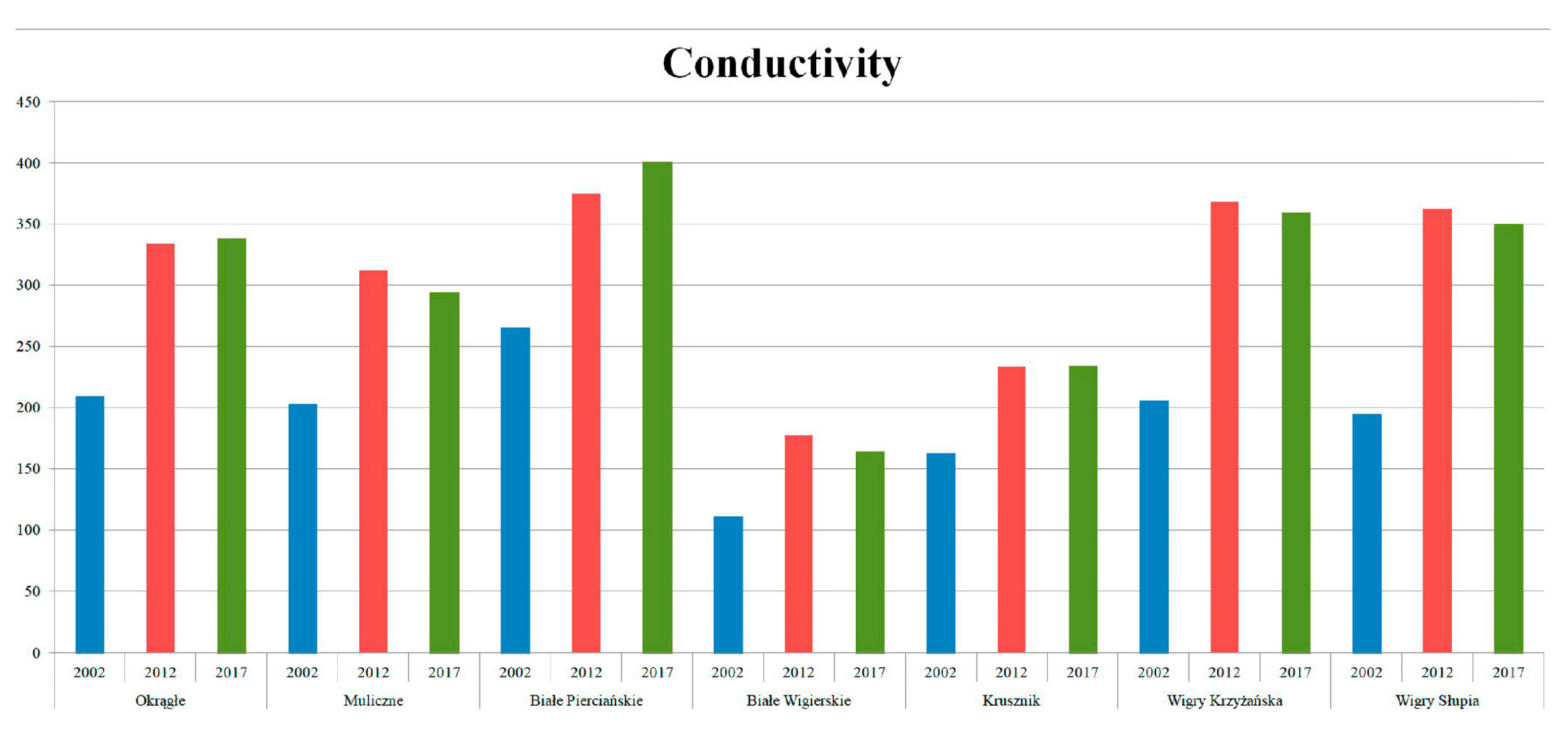
3.1. Physical and Chemical Data
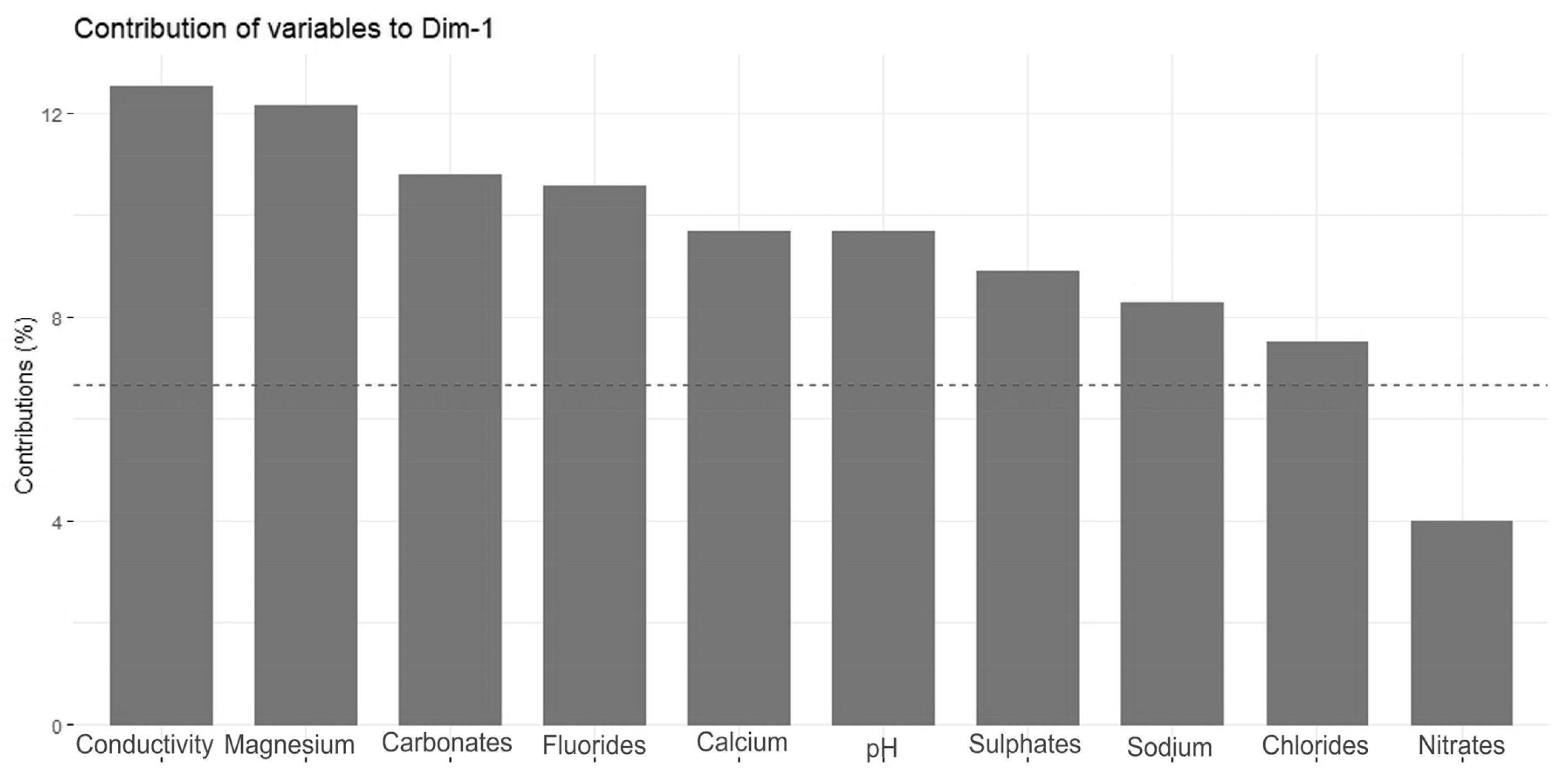
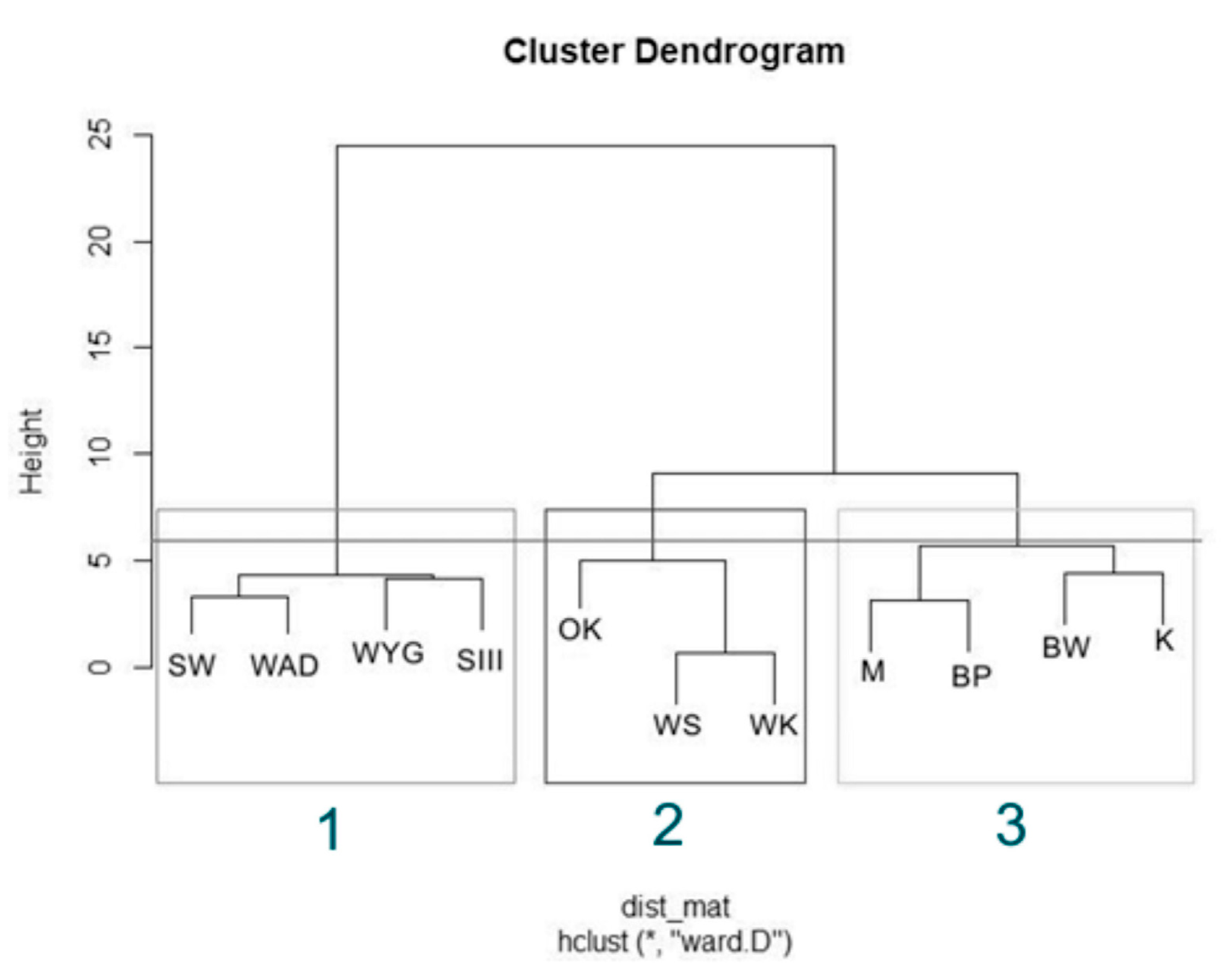
3.2. Statistical Data
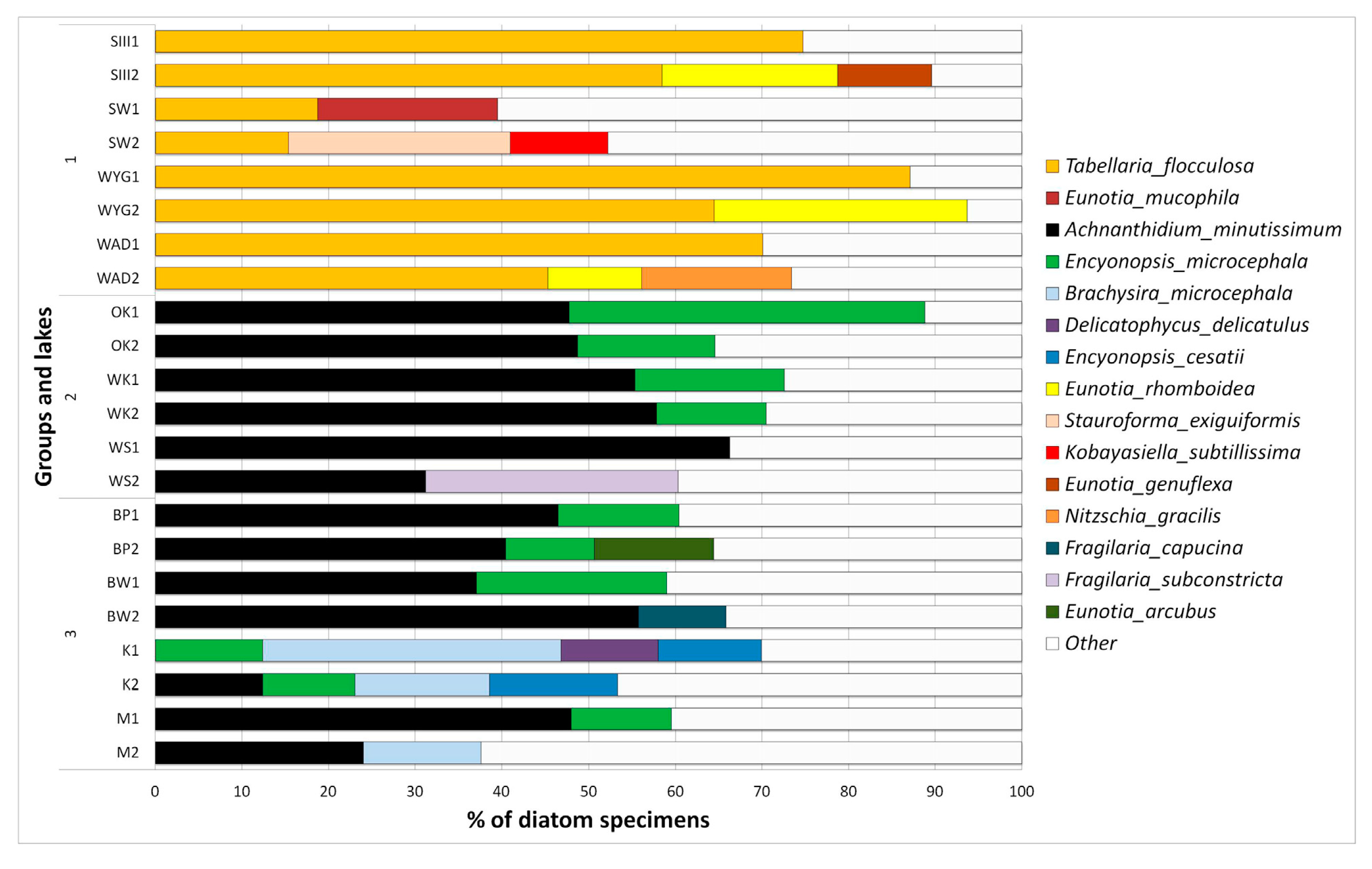
3.3. Diatom Data
4. Discussion
5. Conclusions
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Morandín-Ahuerma, I.; Contreras-Hernández, A.; Ayala-Ortiz, D.A.; Pérez-Maqueo, O. Socio—Ecosystemic sustainability. Sustainability 2019, 11, 3354. [Google Scholar] [CrossRef] [Green Version]
- Krztoń, W.; Kosiba, J.; Pociecha, A. The effect of cyanobacterial blooms on bio-and functional diversity of zooplankton communities. Biodivers. Conserv. 2019, 28, 1815–1835. [Google Scholar] [CrossRef] [Green Version]
- Ognjanova-Rumenova, N.; Wojtal, A.Z.; Sienkiewicz, E.; Botev, I.; Trichkova, T. Biodiversity of high mountain lakes in europe with special regards to Rila Mountains (Bulgaria) and Tatra Mountains (Poland). Diatoms Fundam. Appl. 2019, 335–354. [Google Scholar] [CrossRef]
- Strayer, D.L.; Findlay, S.E. Ecology of freshwater shore zones. Aquat. Sci. 2010, 72, 127–163. [Google Scholar] [CrossRef] [Green Version]
- Hofmann, G.; Werum, M.; Lange-Bertalot, H. Diatomeen im Süßwasser-Benthos von Mitteleuropa. Bestimmungsflora Kieselalgen für die Ökologische Praxis; Über, 700 der Häufigsten Arten und Ihrer Ökologie; Gantner: Nüziders, Austria, 2011; pp. 1–942. [Google Scholar]
- Marra, R.C.; Algarte, V.M.; Ludwig, T.A.; Padial, A.A. Diatom diversity at multiple scales in urban reservoirs in Southern Brazil reveals the likely role of trophic state. Limnologica 2018, 70, 49–57. [Google Scholar] [CrossRef]
- Stoermer, E.F.; Smol, J.P. Applications and uses of diatoms: Prologue. In The Diatoms: Applications for the Environmental and Earth Sciences, 1st ed.; Cambridge University Press: New York, NY, USA, 1999; pp. 3–10. [Google Scholar]
- Van Dam, H. On the use of measures of structure and diversity in applied diatom ecology. Nova Hedwig. 1982, 73, 97–115. [Google Scholar]
- Witkowski, A.; Radziejewska, T.; Wawrzyniak-Wydrowska, B.; Lange-Bertalot, H.; Bąk, M.; Gelbrecht, J. Living on the pH edge: Diatom assemblages of low–pH lakes in western Pomerania (NW Poland). In The Diatom World; Sekbach, J., Kociolek, P., Eds.; Springer: Dordrecht, The Netherlands, 2011; pp. 365–384. [Google Scholar]
- Rybak, M.; Poradowska, A.; Kochman-Kedziora, N.; Peszek, Ł.; Noga, T.; Stanek-Tarkowska, J. Okrzemki (bacillariophyta) torfowiska wołosate (bieszczadzki park narodowy). Rocz. Bieszcz. 2018, 26, 169–183. [Google Scholar]
- Górniak, A. Typologia i Aktualna Trofia Jezior WPN; Wydawnictwa–Uniwersytet w Białymstoku. Zakład Hydrobiology: Białystok, Poland, 2006; pp. 128–140. [Google Scholar]
- Górniak, A. Plan Ochrony dla Wigierskiego Parku Narodowego i Obszaru Natura 2000 Ostoja PLH200004. Available online: http://www.wigry.org.pl/inf_i_rozw/plany/materialy.htm (accessed on 4 August 2019).
- Tomaszewicz, G.H. Flora glonow wigierskiego parku narodowego. Fragm. Florist. Geobot. Pol. 1996, 3, 239–259. [Google Scholar]
- Rakowska, B. Preliminary elaboration of the diatom communities occuring in the benthos of the Kamionka River and Lake Czarne at the Pojezierze Suwalskie (1993–1994). Fragm. Florist. Geobot. Pol. 1996, 3, 221–238. [Google Scholar]
- Bobrukiewicz, K. Bory Tucholskie i Inne Obszary Leśne. Ochrona, Monitoring, Edukacja. Available online: https://books.google.cn/books/about/Bory_Tucholskie_i_inne_obszary_le%C5%9Bne.html?id=ejZ0PgAACAAJ&redir_esc=y (accessed on 12 July 2019).
- Andrzejczyk, T.; Brzeziecki, B. The structure and dynamics of old-growth Pinus sylvestris (L.) stands in the Wigry National Park, north-eastern Poland. Vegetatio 1995, 117, 81–94. [Google Scholar] [CrossRef]
- Drzymulska, D.; Zieliński, P. Phases and interruptions in postglacial development of humic lake margin (Lake Suchar Wielki, NE Poland). Limnol. Rev. 2014, 14, 13–20. [Google Scholar] [CrossRef] [Green Version]
- Górniak, A. Current Climatic Conditions of Lake Regions in Poland and Impacts on Their Functioning; Polish River Basins and Lakes–Part. I, 1–25; Springer: Cham, Switzerland, 2020. [Google Scholar] [CrossRef]
- Skrajna, T.; Kubicka, H. Segetal flora of the wigry national park. Acta Agrobot. 2011, 64, 119–132. [Google Scholar] [CrossRef] [Green Version]
- Kostrowicki, A.S.; Baumgart-Kotarba, M. Studium Geoekologiczne Rejonu Jezior Wigierskich; Zakład Narodowy im. Ossolińskich: Warszawa, Poland, 1988. [Google Scholar]
- Rutkowski, J.; Krzysztofiak, L. Jezioro Wigry: Historia Jeziora w Świetle Badań Geologicznych i Paleoekologicznych; Stowarzyszenie Człowiek i Przyroda: Podlaskie, Poland, 2009. [Google Scholar]
- Boggero, A.; Zaupa, S.; Musazzi, S.; Rogora, M.; Dumnicka, E.; Lami, A. Environmental factors as drivers for macroinvertebrate and diatom diversity in Alpine lakes: New insights from the Stelvio National Park (Italy). J. Limnol. 2019, 78, 147–162. [Google Scholar] [CrossRef]
- Tibshirani, R.; Walther, G. Cluster validation by prediction strength. J. Comput. Graph. Stat. 2005, 14, 511–528. [Google Scholar] [CrossRef]
- R Core Team. R: A Language and Environment for Statistical Computing. R Foundation for Statistical Computing, Vienna, Austria 2013. Available online: http://www.R-project.org (accessed on 2 March 2020).
- Husson, F.; Josse, J.; Le, S.; Mazet, J.; Husson, M.F. Package ‘FactoMineR’. Package R FactorMine 2019, 96, 698. [Google Scholar]
- Wickham, H. Ggplot2: Elegant Graphics for Data Analysis; Springer: Berlin/Heidelberg, Germany, 2016. [Google Scholar] [CrossRef] [Green Version]
- Kassambara, A.; Mundt, F. Factoextra: Extract and visualize the results of multivariate data analyses. R Package Version 2017, 1, 337–354. [Google Scholar]
- McNaughton, S.J.; Wolf, L.L. Dominance and the niche in ecological systems. Science 1970, 167, 131–139. [Google Scholar] [CrossRef]
- Picińska-Fałtynowicz, J.; Błachuta, J. Wytyczne Metodyczne do Przeprowadzenia Oceny Stanu Ekologicznego Jednolitych Części Wód Rzek i Jezior Oraz Potencjału Ekologicznego Sztucznych i Silnie Zmienionych Jednolitych Części Wód Płynących Polski na Podstawie Badań Fitobentosu; Wydawnictwo GIOŚ Warszawa: Warsaw, Poland, 2008; pp. 1–79. [Google Scholar]
- Zgrundo, A.; Peszek, Ł.; Poradowska, A. Podręcznik do Monitoringu i Oceny Rzecznych Jednolitych Części Wód Powierzchniowych na Podstawie Fitobentosu; Ocean Sense: Gdańsk, Poland, 2018. [Google Scholar]
- Kulikovskiy, M.S.; Lange-Bertalot, H.; Witkowski, A.; Dorofeyuk, N.I.; Genkal, S.I. Diatom assemblages from Sphagnum bogs of the world. Bibl. Diatomol. 2010, 55, 1–326. [Google Scholar]
- Wojtal, A.Z.; Ector, L.; Van de Vijver, B.; Morales, E.A.; Blanco, S.; Piatek, J.; Smieja, A. The Achnanthidium minutissimum complex (Bacillariophyceae) in southern Poland. Algol. Stud. 2011, 136, 211–238. [Google Scholar] [CrossRef]
- Lange-Bertalot, H.; Ulrich, S. Contributions to the taxonomy of needle-shaped Fragilaria and Ulnaria species. Lauterbornia 2014, 78, 1–73. [Google Scholar]
- Van de Vijver, B. Analysis of the type material of Navicula brachysira Brébisson with the description of Brachysira sandrae, a new raphid diatom (Bacillariophyceae) from Iles Kerguelen (TAAF, sub-Antarctica, southern Indian Ocean). Phytotaxa 2014, 184, 139–147. [Google Scholar] [CrossRef]
- Delgado, C.; Novais, M.H.; Blanco, S.; Almeida, S.F. Examination and comparison of Fragilaria candidagilae sp. nov. with type material of Fragilaria recapitellata, F. capucina, F. perminuta, F. intermedia and F. neointermedia (Fragilariales, Bacillariophyceae). Phytotaxa 2015, 231, 1–18. [Google Scholar] [CrossRef]
- Lengyel, E. Stress and Disturbance in Benthic Diatom Assemblages. Examples from Lakes and Streams and from the Field and Laboratory Studies. Ph.D. Thesis, University of Pannonia, Veszprém, Hungary, 2016. [Google Scholar] [CrossRef]
- Kennedy, B.; Allott, N. A review of the genus Brachysira in Ireland with the description of Brachysira praegeri and Brachysira conamarae, new raphid diatoms (Bacillariophyceae) from high status waterbodies. Phytotaxa 2017, 326, 1–27. [Google Scholar] [CrossRef]
- Heudre, D.; Wetzel, C.E.; Moreau, L.; Van de Vijver, B.; Ector, L. On the identity of the rare Fragilaria subconstricta (Fragilariaceae), with Fragilaria species forming ribbon–like colonies shortly reconsidered. Plant Ecol. Evol. 2019, 152, 327–339. [Google Scholar] [CrossRef]
- Bennett, J.R.; Cumming, B.F.; Ginn, B.K.; Smol, J.P. Broad-scale environmental response and niche conservatism in lacustrine diatom communities. Glob. Ecol. Biogeogr. 2010, 19, 724–732. [Google Scholar] [CrossRef]
- Schaumburg, J.; Schranz, C.; Hofmann, G.; Stelzer, D.; Schneider, S.; Schmedtje, U. Macrophytes and phytobenthos as indicators of ecological status in German lakes—A contribution to the implementation of the Water Framework Directive. Limnologica 2004, 34, 302–314. [Google Scholar] [CrossRef] [Green Version]
- Ciecierska, H.; Kolada, A. ESMI: A macrophyte index for assessing the ecological status of lakes. Environ. Monit. Assess. 2014, 186, 5501–5517. [Google Scholar] [CrossRef] [Green Version]
- Kłosowski, S.; Tomaszewicz, G.H.; Tomaszewicz, H. The expansion and decline of charophyte communities in lakes within the Sejny Lake District (north-eastern Poland) and changes in water chemistry. Limnologica 2006, 36, 234–240. [Google Scholar] [CrossRef]
- Siuda, W.; Kaliński, T.; Kauppinen, E.; Chróst, R.J. Eutrofizacja południowej części kompleksu wielkich jezior mazurskich w latach 1977–2011. Technol. Wody 2014, 3, 48–62. [Google Scholar]
- Bok, G.; Boguszewska, A.; Barańska, A.; Panasewicz, A.; Wiśniewska, M. Raport o Stanie Środowiska Województwa Podlaskiego 2015. Available online: http://www.wios.bialystok.pl/pdf/wios_raport2015_wersja_WWW.pdf (accessed on 6 June 2020).
- Panek, P.; Biliński, G.; Brodowska, M.; Chełstowska, A.; Czajka, J.; Dudek, M.; Fornal, B.; Kamińska, M.; Kasprowicz, H.; Kociuba, M.; et al. Stan Środowiska w Polsce. Raport 2018. Available online: http://www.gios.gov.pl/pl/aktualnosci/521-stan-srodowiska-w-polsce-raport-2018 (accessed on 12 May 2020).
- Górniak, A.; Jekatierynczuk-Rudczyk, E.; Dobrzyń, P. Hydrochemistry of three dystrophic lakes in Northeastern Poland. Acta Hydrochim. Hydrobiol. 1999, 27, 12–18. [Google Scholar] [CrossRef]
- Hervé, V.; Derr, J.; Douady, S.; Quinet, M.; Moisan, L.; Lopez, P.J. Multiparametric analyses reveal the pH-dependence of silicon biomineralization in diatoms. PLoS ONE 2012, 7, e46722. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lange-Bertalot, H.; Hofmann, G.; Werum, M.; Cantonati, M. Freshwater Benthic Diatoms of Central Europe: Over 800 Common Species Used in Ecological Assessment; Koeltz Botanical Books: Hochtaunuskreis, Germany, 2017. [Google Scholar]
- Bennion, H.; Sayer, C.D.; Tibby, J.; Carrick, H.J. Diatoms as indicatorsof environmental change in shallow lakes. In The Diatoms: Applications for the Environmental and Earth Sciences; Cambridge University Press: Cambridge, UK, 2010. [Google Scholar] [CrossRef]
- Pęczuła, W.; Grabowska, M.; Zieliński, P.; Karpowicz, M.; Danilczyk, M. Vertical distribution of expansive, bloom-forming algae Gonyostomum semen vs. plankton community and water chemistry in four small humic lakes. Knowl. Manag. Aquat. Ecosyst. 2018, 419, 28. [Google Scholar] [CrossRef]
- Karpowicz, M.; Zieliński, P.; Grabowska, M.; Ejsmont-Karabin, J.; Kozłowska, J.; Feniova, I. Effect of eutrophication and humification on nutrient cycles and transfer efficiency of matter in freshwater food webs. Hydrobiologia 2020. [Google Scholar] [CrossRef]
- Pestryakova, L.A.; Herzschuh, U.; Gorodnichev, R.; Wetterich, S. The sensitivity of diatom taxa from Yakutian lakes (north-eastern Siberia) to electrical conductivity and other environmental variables. Polar Res. 2018, 37, 1485625. [Google Scholar] [CrossRef] [Green Version]
- Szczęsny, B.; Majecki, J. Variations in the composition of Trichoptera communities associated with lakes of different trophy in the Wigry National Park (NE-Poland). Nova Suppl. Entomol. 2002, 15, 581–593. [Google Scholar]
- Van Dam, H.; Mertens, A.; Sinkeldam, J. A coded checklist and ecological indicator values of freshwater diatoms from The Netherlands. Neth. J. Aquat. Ecol. 1994, 28, 117–133. [Google Scholar] [CrossRef]
- Calapez, A.R.; Elias, C.L.; Almeida, S.F.; Feio, M.J. Extreme drought effects and recovery patterns in the benthic communities of temperate streams. Limnetica 2014, 33, 281–296. [Google Scholar] [CrossRef]
- Connell, J.H. Diversity in tropical rain forests and coral reefs. Science 1978, 199, 1302–1310. [Google Scholar] [CrossRef] [Green Version]
- Fox, J.W. The intermediate disturbance hypothesis should be abandoned. Trends Ecol. Evol. 2013, 28, 86–92. [Google Scholar] [CrossRef]
- Sheil, D.D.; Burslem, F.R.P. Defining and defending Connell’s intermediate disturbance hypothesis: A response to Fox. Trends Ecol. Evol. 2013, 28, 571–572. [Google Scholar] [CrossRef]
- Pandey, U. Anthropogenic drivers shift diatom dominance-diversity relationships and transparent exopolymeric particles production in River Ganga: Implication for natural cleaning of river water. Curr. Sci. 2017, 00113891, 113. [Google Scholar] [CrossRef]
- Schneider, S.C.; Hjermann, D.O.; Edvardsen, H. Do benthic algae provide important information over and above that provided by macrophytes and phytoplankton in lake status assessment?–Results from a case study in Norway. Limnologica 2019, 76, 28–40. [Google Scholar] [CrossRef]
- Taylor, D.R.; Aarssen, L.W.; Loehle, C. On the relationship between r/K selection and environmental carrying capacity: A new habitat templet for plant life history strategies. Oikos 1990, 58, 239–250. [Google Scholar] [CrossRef]
- Pither, J.; Aarssen, L.W. The evolutionary species pool hypothesis and patterns of freshwater diatom diversity along a pH gradient. J. Biogeogr. 2005, 32, 503–513. [Google Scholar] [CrossRef]
- Witkowski, A.; Bąk, M.; Kierzek, A. Zmiany warunków środowiskowych w Holocenie w jeziorze Wigry w świetle analizy diatomologicznej osadów. In Jezioro Wigry Historia Jeziora w Świetle Badań Geologicznych i Paleoekologicznych; Rutkowski, J., Krzysztofiak, L., Eds.; Stowarzyszenie “Człowiek i Przyroda”: Podlaskie, Poland, 2009; pp. 215–226. [Google Scholar]
- Zawisza, E.; Szeroczyńska, K. The development history of Wigry Lake as shown by subfossil Cladocera. Geochronometria 2007, 27, 67–74. [Google Scholar] [CrossRef] [Green Version]
- González-Trujillo, J.D.; Pedraza-Garzón, E.; Donato-Rondon, J.C.; Sabater, S. Ecoregional characteristics drive the distribution patterns of neotropical stream diatoms. J. Phycol. 2020. [Google Scholar] [CrossRef]
- Soininen, J.; Heino, J. Variation in niche parameters along the diversity gradient of unicellular eukaryote assemblages. Protist 2007, 158, 181–191. [Google Scholar] [CrossRef]
- Chen, W.; Wang, Q.; Xia, S.; Pei, G. Comparative responses of dominant and subordinate periphytic diatom species to environmental gradients in Donghu Lake, China. Int. Rev. Hydrobiol. 2016, 101, 182–192. [Google Scholar] [CrossRef]
- MacDougall, M.J.; Paterson, A.M.; Winter, J.G.; Jones, F.C.; Knopf, L.A.; Hall, R.I. Response of periphytic diatom communities to multiple stressors influencing lakes in the Muskoka River watershed, Ontario, Canada. Freshw. Sci. 2017, 36, 77–89. [Google Scholar] [CrossRef]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Lake | Lake Area [ha] | Max. Depth [m] | Coastline Length [m] | Direct Catchment [ha] | Catchment [ha] | Typology of Lakes | Lake Group |
|---|---|---|---|---|---|---|---|
| W | 2163.3 | 73 | 63920 | 5159.8 | 45293.1 | flow-through | Wigierskie |
| K | 26.7 | 18 | 2643 | 70.7 | 70.7 | exorheic | Wigierskie |
| BW | 99.9 | 34 | 5117 | 329.1 | 329.1 | exorheic | Wigierskie |
| M | 24.1 | 11.3 | 3175 | 191.2 | 191.2 | exorheic | Wigierskie |
| OK | 13.7 | 13 | 1459 | 28.5 | 906.8 | flow-through | Wigierskie |
| BP | 6.9 | 24 | 1011 | 50.4 | 50.4 | endorheic | Pierciańskie |
| WYG | 2 | 3 | 670 | 63.5 | 63.5 | endorheic | Wigierskie |
| SW | 8.44 | 9.6 | 2066 | 107.1 | 107.1 | endorheic | Wigierskie |
| WAD | 1.09 | 15 | 474 | 19.4 | 19.4 | endorheic | Pierciańskie |
| SIII | 0.44 | 4 | 320 | 32.2 | 32,2 | exorheic | Huciańskie |
| Lake | Chlorides | Carbonates | Sulphates | Nitrates | Ammonium | Magnesium | Phosphates | Calcium | pH | Conductivity | Dominance Index |
|---|---|---|---|---|---|---|---|---|---|---|---|
| [mg/L] | µS/cm | % | |||||||||
| OK | 8.45–9.99 | 129.30–248 | 28.37–30.92 | 0.48–1.75 | 0.04–0.83 | 11.58–15.58 | 0.002–0.02 | 46.81–73.62 | 7.68–8.28 | 338–385 | 68–89 |
| M | 3.52–3.85 | 126.70–239 | 21.35–24.06 | 0–0.37 | 0.02–0.22 | 10.14–14.45 | 0.002–0.011 | 40.21–68.25 | 7.52–8.26 | 294–342 | 38–59 |
| BP | 2.28–2.94 | 186.77–292 | 3.15–5.63 | 0–0.35 | 0.02–0.32 | 13.88–18.95 | 0.002–0.01 | 48.46–69.45 | 7.63–8.23 | 385–401 | 43–64 |
| BW | 2.68–3.35 | 74.81–137.4 | 6.35–6.81 | 0.00–0.36 | 0.01–0.13 | 4.93–6.53 | 0.002–0.03 | 23.85–36.66 | 7.32–8.15 | 164–185 | 59–66 |
| K | 4.24–4.52 | 101.49–239 | 12.36–22.23 | 0–0.35 | 0.036–0.23 | 7.66–14.45 | 0.002–0.01 | 30.45–68.25 | 7.51–8.26 | 234–342 | 30–47 |
| WK | 14.92–17.05 | 138.99–243 | 21.68–23.47 | 0–1.26 | 0.012–0.12 | 11.91–16.07 | 0.002–0.01 | 16.05–45.67 | 7.56–8.23 | 359–402 | 71–73 |
| WS | 14.47–16.62 | 135.36–237 | 21.92–23.57 | 0–1.20 | 0.01–0.12 | 11.80–16.08 | 0.003–0.03 | 16.08–43.93 | 7.60–8.15 | 350–388 | 61–73 |
| WYG | 1.40–2.03 | 8.44–35.71 | 0.02–1.23 | 0.00–0.40 | 0.20–1.26 | 0.00–0.49 | 0.01–0.03 | 0.85–2.01 | 3.7–6.11 | 20–25 | 89–94 |
| WAD | 1.27–1.35 | 17.56–18.60 | 0.02–0.56 | 0–0 | 0.00–0.20 | 0.77 | 0.005–0.04 | 2.67–3.35 | 4.65–5.31 | 20–23 | 63–76 |
| SIII | 0.86–8.44 | 9.04–18.45 | 0.02–1.11 | 0–0 | 0.08–0.20 | 0.00–0.46 | 0.02–0.05 | 0.59–1.74 | 3.6–6.8 | 22–23 | 79–83 |
| SW | 0.97–1.28 | 8.66–39.66 | 2.23–2.68 | 0.00–0.36 | 0.00–0.00 | 0.50–0.74 | 0.01–0.02 | 1.73–2.75 | 4.60–6.34 | 15–18 | 40–41 |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Eliasz-Kowalska, M.; Wojtal, A.Z. Limnological Characteristics and Diatom Dominants in Lakes of Northeastern Poland. Diversity 2020, 12, 374. https://doi.org/10.3390/d12100374
Eliasz-Kowalska M, Wojtal AZ. Limnological Characteristics and Diatom Dominants in Lakes of Northeastern Poland. Diversity. 2020; 12(10):374. https://doi.org/10.3390/d12100374
Chicago/Turabian StyleEliasz-Kowalska, Monika, and Agata Z. Wojtal. 2020. "Limnological Characteristics and Diatom Dominants in Lakes of Northeastern Poland" Diversity 12, no. 10: 374. https://doi.org/10.3390/d12100374
APA StyleEliasz-Kowalska, M., & Wojtal, A. Z. (2020). Limnological Characteristics and Diatom Dominants in Lakes of Northeastern Poland. Diversity, 12(10), 374. https://doi.org/10.3390/d12100374