Isolation and Identification of Endophytic Bacteria from Mycorrhizal Tissues of Terrestrial Orchids from Southern Chile

 , and
, and
Abstract
:1. Introduction
2. Materials and Methods
2.1. Sampling
2.2. Isolation of Endophytic Bacteria
2.3. Molecular Identification of Bacteria
2.4. Screening of Plant Growth-Promoting Traits
2.5. Antifungal Activity
2.6. Data Analyses
3. Results
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Conflicts of Interest
References
- Roossinck, M.J. The good viruses: viral mutualistic symbioses. Nat. Rev. Microbiol. 2011, 9, 99. [Google Scholar] [CrossRef]
- Novotná, A.; Benítez, Á.; Herrera, P.; Cruz, D.; Filipczyková, E.; Suárez, J.P. High diversity of root-associated fungi isolated from three epiphytic orchids in southern Ecuador. Mycoscience 2018, 59, 24–32. [Google Scholar] [CrossRef]
- Strobel, G.A. Fungal Endophytes in Plants; MDPI: Basel, Switzerland, 2018. [Google Scholar]
- Glazebrook, J.; Roby, D. Plant biotic interactions: from conflict to collaboration. Plant J. 2018, 93, 589–591. [Google Scholar] [CrossRef] [Green Version]
- Liu, H.; Carvalhais, L.C.; Crawford, M.; Singh, E.; Dennis, P.G.; Pieterse, C.M.; Schenk, P.M. Inner plant values: diversity, colonization and benefits from endophytic bacteria. Front. Microbiol. 2017, 8, 2552. [Google Scholar] [CrossRef]
- Glick, B.R. Plant growth-promoting bacteria: mechanisms and applications. Scientifica 2012, 963401. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kloepper, J.; Schroth, M. Plant growth promoting rhizobacteria on radish. In Proceedings of the Fourth Conference Plant Pathogenic Bacteria, Angers, France, 27 August– 2 September 1978; Gilbert-Clarey: Tours, France, 1978; pp. 876–882. [Google Scholar]
- Compant, S.; Clément, C.; Sessitsch, A. Plant growth-promoting bacteria in the rhizo-and endosphere of plants: their role, colonization, mechanisms involved and prospects for utilization. Soil Biol. Biochem. 2010, 42, 669–678. [Google Scholar] [CrossRef] [Green Version]
- Rashid, S.; Charles, T.C.; Glick, B.R. Isolation and characterization of new plant growth-promoting bacterial endophytes. Appl. Soil Ecol. 2012, 61, 217–224. [Google Scholar] [CrossRef]
- Etesami, H.; Maheshwari, D.K. Use of plant growth promoting rhizobacteria (PGPRs) with multiple plant growth promoting traits in stress agriculture: action mechanisms and future prospects. Ecotox. Environ. Safe. 2018, 156, 225–246. [Google Scholar] [CrossRef] [PubMed]
- Senthilkumar, M.; Krishnamoorthy, R. Isolation and characterization of tomato leaf phyllosphere Methylobacterium and their effect on plant growth. Int. J. Curr. Microbiol. Appl. Sci. 2017, 6, 2121–2136. [Google Scholar] [CrossRef]
- Roberts, D.L.; Dixon, K.W. Orchids. Curr. Biol. 2008, 18, R325–R329. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Novoa, P.; Espejo, J.; Cisternas, M.; Rubio, M.; Dominguez, E. Guía de campo de las orquídeas chilenas; Segunda edición ampliada; Corporación Chilena de la Madera (CORMA): Santiago, Chile, 2015. [Google Scholar]
- Herrera, H.; García-Romera, I.; Meneses, C.; Pereira, G.; Arriagada, C. Orchid Mycorrhizal Interactions on the Pacific Side of the Andes from Chile. A Review. J. Soil Sci. Plant Nut. 2019, 19, 187–202. [Google Scholar] [CrossRef]
- Valadares, R.B.; Pereira, M.C.; Otero, J.T.; Cardoso, E.J. Narrow fungal mycorrhizal diversity in a population of the orchid Coppensia doniana. Biotropica 2012, 44, 114–122. [Google Scholar] [CrossRef]
- Bayman, P.; Otero, J.T. Microbial endophytes of orchid roots. In Microbial Root Endophytes; Springer: New York, NY, USA, 2006; pp. 153–177. [Google Scholar]
- Tsavkelova, E. Bacteria associated with orchid roots. In Bacteria in Agrobiology: Plant Growth Responses; Springer: New York, NY, USA; pp. 221–258.
- McCormick, M.K.; Taylor, D.L.; Whigham, D.F.; Burnett, R.K., Jr. Germination patterns in three terrestrial orchids relate to abundance of mycorrhizal fungi. J. Ecol. 2016, 104, 744–754. [Google Scholar] [CrossRef] [Green Version]
- Dearnaley, J.; Perotto, S.; Selosse, M.A. Structure and development of orchid mycorrhizas. In Molecular Mycorrhizal Symbiosis; Martin, E., Ed.; Springer: Berlin, Germany, 2016; pp. 63–86. [Google Scholar]
- Zettler, L.W.; Corey, L.L. Orchid Mycorrhizal Fungi: Isolation and Identification Techniques. In Orchid Propagation: From Laboratories to Greenhouses—Methods and Protocols; Springer: New York, NY, USA, 2018; pp. 27–59. [Google Scholar]
- McCormick, M.K.; Whigham, D.F.; Canchani-Viruet, A. Mycorrhizal fungi affect orchid distribution and population dynamics. New Phytol. 2018, 219, 1207–1215. [Google Scholar] [CrossRef] [Green Version]
- Huang, H.; Zi, X.-M.; Lin, H.; Gao, J.-Y. Host-specificity of symbiotic mycorrhizal fungi for enhancing seed germination, protocorm formation and seedling development of over-collected medicinal orchid, Dendrobium devonianum. J. Microbiol. 2018, 56, 42–48. [Google Scholar] [CrossRef]
- Girija, D.; Rajeevan, P.; Balakrishnan, S.; Panchami, P.; Mohan, M. 16S rRNA gene taxonomic profiling of endophytic bacteria associated with phalaenopsis roots. J. Hortic. Sci. 2018, 13, 103–107. [Google Scholar]
- Novotna, A.; Suárez, J. Molecular detection of bacteria associated with Serendipita sp., a mycorrhizal fungus from the orchid Stanhopea connata Klotzsch in southern Ecuador. Bot. Lett. 2018, 165, 307–313. [Google Scholar] [CrossRef]
- Faria, D.C.; Dias, A.C.F.; Melo, I.S.; de Carvalho Costa, F.E. Endophytic bacteria isolated from orchid and their potential to promote plant growth. World J. Microbiol. Biotechnol. 2013, 29, 217–221. [Google Scholar] [CrossRef]
- Tsavkelova, E.; Cherdyntseva, T.; Netrusov, A. Bacteria associated with the roots of epiphytic orchids. Microbiology 2004, 73, 710–715. [Google Scholar] [CrossRef]
- Zhang, X.; Gao, Z.; Zhang, M.; Jing, F.; Du, J.; Zhang, L. Analysis of endophytic actinobacteria species diversity in the stem of Gynura cusimbua by 16S rRNA gene clone library. Microbiology 2016, 85, 379–385. [Google Scholar] [CrossRef]
- Chen, J.; Hu, K.-X.; Hou, X.-Q.; Guo, S.-X. Endophytic fungi assemblages from 10 Dendrobium medicinal plants (Orchidaceae). World J. Microbiol. Biotechnol. 2011, 27, 1009–1016. [Google Scholar] [CrossRef]
- Larkin, M.A.; Blackshields, G.; Brown, N.; Chenna, R.; McGettigan, P.A.; McWilliam, H.; Valentin, F.; Wallace, I.M.; Wilm, A.; Lopez, R. Clustal W and Clustal X version 2.0. Bioinformatics 2007, 23, 2947–2948. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hall, T.A. BioEdit: a user-friendly biological sequence alignment editor and analysis program for Windows 95/98/NT. Nucl. Acids. Symp. Ser. 1999, 41, 95–98. [Google Scholar]
- Kumar, S.; Stecher, G.; Li, M.; Knyaz, C.; Tamura, K. MEGA X: molecular evolutionary genetics analysis across computing platforms. Mol. Biol. Evol. 2018, 35, 1547–1549. [Google Scholar] [CrossRef] [PubMed]
- Herrera, H.; Valadares, R.; Contreras, D.; Bashan, Y.; Arriagada, C. Mycorrhizal compatibility and symbiotic seed germination of orchids from the Coastal Range and Andes in south central Chile. Mycorrhiza 2017, 27, 175–188. [Google Scholar] [CrossRef] [PubMed]
- Ortiz, J.; Soto, J.; Almonacid, L.; Fuentes, A.; Campos-Vargas, R.; Arriagada, C. Alleviation of metal stress by Pseudomonas orientalis and Chaetomium cupreum strains and their effects on Eucalyptus globulus growth promotion. Plant Soil 2019, 436, 449–461. [Google Scholar] [CrossRef]
- Milagres, A.M.; Machuca, A.; Napoleao, D. Detection of siderophore production from several fungi and bacteria by a modification of chrome azurol S (CAS) agar plate assay. J. Microbiol. Methods 1999, 37, 1–6. [Google Scholar] [CrossRef]
- Petatan-Sagahon, I.; Anducho-Reyes, M.A.; Silva-Rojas, H.V.; Arana-Cuenca, A.; Tellez-Jurado, A.; Cárdenas-Álvarez, I.O.; Mercado-Flores, Y. Isolation of bacteria with antifungal activity against the phytopathogenic fungi Stenocarpella maydis and Stenocarpella macrospora. Int. J. Mol. Sci. 2011, 12, 5522–5537. [Google Scholar] [CrossRef]
- Sahin, N.; Tani, A.; Kotan, R.; Sedláček, I.; Kimbara, K.; Tamer, A.U. Pandoraea oxalativorans sp. nov., Pandoraea faecigallinarum sp. nov. and Pandoraea vervacti sp. nov., isolated from oxalate-enriched culture. Int. J. Syst. Evol. Microbiol. 2011, 61, 2247–2253. [Google Scholar] [CrossRef]
- Benidire, L.; Pereira, S.; Castro, P.M.; Boularbah, A. Assessment of plant growth promoting bacterial populations in the rhizosphere of metallophytes from the Kettara mine, Marrakech. Environ. Sci. Pollut. Res. 2016, 23, 21751–21765. [Google Scholar] [CrossRef]
- Leigh, M.B.; Prouzová, P.; Macková, M.; Macek, T.; Nagle, D.P.; Fletcher, J.S. Polychlorinated biphenyl (PCB)-degrading bacteria associated with trees in a PCB-contaminated site. Appl. Environ. Microbiol. 2006, 72, 2331–2342. [Google Scholar] [CrossRef] [Green Version]
- Chimwamurombe, P.M.; Grönemeyer, J.L.; Reinhold-Hurek, B. Isolation and characterization of culturable seed-associated bacterial endophytes from gnotobiotically grown Marama bean seedlings. FEMS Microbiol. Ecol. 2016, 92, fiw083. [Google Scholar] [CrossRef] [Green Version]
- Reinhold-Hurek, B.; Bünger, W.; Burbano, C.S.; Sabale, M.; Hurek, T. Roots shaping their microbiome: global hotspots for microbial activity. Annu. Rev. Phytopatol. 2015, 53, 403–424. [Google Scholar] [CrossRef]
- Tsavkelova, E.A.; Cherdyntseva, T.A.; Botina, S.G.; Netrusov, A.I. Bacteria associated with orchid roots and microbial production of auxin. Microbiol. Res. 2007, 162, 69–76. [Google Scholar] [CrossRef]
- Tan, X.M.; Wang, C.L.; Chen, X.M.; Zhou, Y.Q.; Wang, Y.Q.; Luo, A.X.; Liu, Z.H.; Guo, S.X. In vitro seed germination and seedling growth of an endangered epiphytic orchid, Dendrobium officinale, endemic to China using mycorrhizal fungi (Tulasnella sp.). Sci. Hortic. 2014, 165, 62–68. [Google Scholar] [CrossRef]
- Wilkinson, K.; Dixon, K.; Sivasithamparam, K. Interaction of soil bacteria, mycorrhizal fungi and orchid seed in relation to germination of Australian orchids. New Phytol. 1989, 112, 429–435. [Google Scholar] [CrossRef]
- Cameron, D.D.; Leake, J.R.; Read, D.J. Mutualistic mycorrhiza in orchids: evidence from plant–fungus carbon and nitrogen transfers in the green-leaved terrestrial orchid Goodyera repens. New Phytol. 2006, 171, 405–416. [Google Scholar] [CrossRef]
- Cameron, D.D.; Johnson, I.; Read, D.J.; Leake, J.R. Giving and receiving: measuring the carbon cost of mycorrhizas in the green orchid, Goodyera repens. New Phytol. 2008, 180, 176–184. [Google Scholar] [CrossRef]
- Ogura-Tsujita, Y.; Gebauer, G.; Hashimoto, T.; Umata, H.; Yukawa, T. Evidence for novel and specialized mycorrhizal parasitism: the orchid Gastrodia confusa gains carbon from saprotrophic Mycena. Proc. R. Soc. B. 2008, 276, 761–767. [Google Scholar] [CrossRef] [Green Version]
- Afzal, I.; Shinwari, Z.K.; Sikandar, S.; Shahzad, S. Plant beneficial endophytic bacteria: Mechanisms, diversity, host range and genetic determinants. Microbiol. Res. 2019, 221, 36–49. [Google Scholar] [CrossRef]
- Kuga, Y.; Sakamoto, N.; Yurimoto, H. Stable isotope cellular imaging reveals that both live and degenerating fungal pelotons transfer carbon and nitrogen to orchid protocorms. New Phytol. 2014, 202, 594–605. [Google Scholar] [CrossRef]
- Tsavkelova, E.; Cherdyntseva, T.; Lobakova, E.; Kolomeitseva, G.; Netrusov, A. Microbiota of the orchid rhizoplane. Microbiology 2001, 70, 492–497. [Google Scholar] [CrossRef]
- Tsavkelova, E.A.; Egorova, M.A.; Leontieva, M.R.; Malakho, S.G.; Kolomeitseva, G.L.; Netrusov, A.I. Dendrobium nobile Lindl. seed germination in co-cultures with diverse associated bacteria. Plant Growth Regul. 2016, 80, 79–91. [Google Scholar] [CrossRef]
- Sandhya, V.; Ali, S.Z.; Grover, M.; Reddy, G.; Venkateswarlu, B. Effect of plant growth promoting Pseudomonas spp. on compatible solutes, antioxidant status and plant growth of maize under drought stress. Plant Growth Regul. 2010, 62, 21–30. [Google Scholar] [CrossRef]
- Sharma, A.; Johri, B.; Sharma, A.; Glick, B. Plant growth-promoting bacterium Pseudomonas sp. strain GRP3 influences iron acquisition in mung bean (Vigna radiata L. Wilzeck). Soil Biol. Biochem. 2003, 35, 887–894. [Google Scholar] [CrossRef]
- Otieno, N.; Lally, R.D.; Kiwanuka, S.; Lloyd, A.; Ryan, D.; Germaine, K.J.; Dowling, D.N. Plant growth promotion induced by phosphate solubilizing endophytic Pseudomonas isolates. Front. Microbiol. 2015, 6, 745. [Google Scholar] [CrossRef] [Green Version]
- Raheem, A.; Shaposhnikov, A.; Belimov, A.A.; Dodd, I.C.; Ali, B. Auxin production by rhizobacteria was associated with improved yield of wheat (Triticum aestivum L.) under drought stress. Arch. Agron. Soil Sci. 2018, 64, 574–587. [Google Scholar] [CrossRef]
- Rath, M.; Mitchell, T.; Gold, S. Volatiles produced by Bacillus mojavensis RRC101 act as plant growth modulators and are strongly culture-dependent. Microbiol. Res. 2018, 208, 76–84. [Google Scholar] [CrossRef]
- Bharti, N.; Yadav, D.; Barnawal, D.; Maji, D.; Kalra, A. Exiguobacterium oxidotolerans, a halotolerant plant growth promoting rhizobacteria, improves yield and content of secondary metabolites in Bacopa monnieri (L.) Pennell under primary and secondary salt stress. World J. Microbiol. Biotechnol. 2013, 29, 379–387. [Google Scholar] [CrossRef]
- Guglielmetti, S.; Basilico, R.; Taverniti, V.; Arioli, S.; Piagnani, C.; Bernacchi, A. Luteibacter rhizovicinus MIMR1 promotes root development in barley (Hordeum vulgare L.) under laboratory conditions. World J. Microbiol. Biotechnol. 2013, 29, 2025–2032. [Google Scholar] [CrossRef]
- Galdiano, J.R.F.; Pedrinho, E.A.N.; Castellane, T.C.L.; Lemos, E.G.d.M. Auxin-producing bacteria isolated from the roots of Cattleya walkeriana, an endangered Brazilian orchid, and their role in acclimatization. Rev. Bras. Ciênc. Solo 2011, 35, 729–737. [Google Scholar]
- Herrera, H.; Novotná, A.; Ortiz, J.; Soto, J.; Arriagada, C. Isolation and identification of plant growth-promoting bacteria from rhizomes of Arachnitis uniflora, a fully mycoheterotrophic plant in southern Chile. Appl. Soil. Ecol. 2020, 149, 103512. [Google Scholar] [CrossRef]
- Rout, M.E.; Southworth, D. The root microbiome influences scales from molecules to ecosystems: the unseen majority1. Am. J. Bot. 2013, 100, 1689–1691. [Google Scholar] [CrossRef] [Green Version]
- Tsavkelova, E.; Cherdyntseva, T.; Netrusov, A. Auxin production by bacteria associated with orchid roots. Microbiology 2005, 74, 46–53. [Google Scholar] [CrossRef]
- Tsavkelova, E.A.; Cherdyntseva, T.A.; Klimova, S.Y.; Shestakov, A.I.; Botina, S.G.; Netrusov, A.I. Orchid-associated bacteria produce indole-3-acetic acid, promote seed germination, and increase their microbial yield in response to exogenous auxin. Arch. Microbiol. 2007, 188, 655–664. [Google Scholar] [CrossRef]
- Höppener-Ogawa, S.; de Boer, W.; Leveau, J.; Van Veen, J.; De Brandt, E.; Vanlaere, E.; Sutton, H.; Dare, D.J.; Vandamme, P. Collimonas arenae sp. nov. and Collimonas pratensis sp. nov., isolated from (semi-) natural grassland soils. Int. J. Syst. Evol. Microbiol. 2008, 58, 414–419. [Google Scholar] [CrossRef]
- Garbeva, P.; Hordijk, C.; Gerards, S.; De Boer, W. Volatiles produced by the mycophagous soil bacterium Collimonas. FEMS Microbiol. Ecol. 2014, 87, 639–649. [Google Scholar] [CrossRef] [Green Version]
- Poosakkannu, A.; Nissinen, R.; Kytöviita, M.M. Culturable endophytic microbial communities in the circumpolar grass, Deschampsia flexuosa in a sub-Arctic inland primary succession are habitat and growth stage specific. Environ. Microbiol. Rep. 2015, 7, 111–122. [Google Scholar] [CrossRef]
- de Boer, W.; Leveau, J.H.; Kowalchuk, G.A.; Gunnewiek, P.J.K.; Abeln, E.C.; Figge, M.J.; Sjollema, K.; Janse, J.D.; van Veen, J.A. Collimonas fungivorans gen. nov., sp. nov., a chitinolytic soil bacterium with the ability to grow on living fungal hyphae. Int. J. Syst. Evol. Microbiol. 2004, 54, 857–864. [Google Scholar] [CrossRef] [Green Version]
- Hoffman, M.T.; Gunatilaka, M.K.; Wijeratne, K.; Gunatilaka, L.; Arnold, A.E. Endohyphal bacterium enhances production of indole-3-acetic acid by a foliar fungal endophyte. PLoS ONE 2013, 8, e73132. [Google Scholar] [CrossRef]
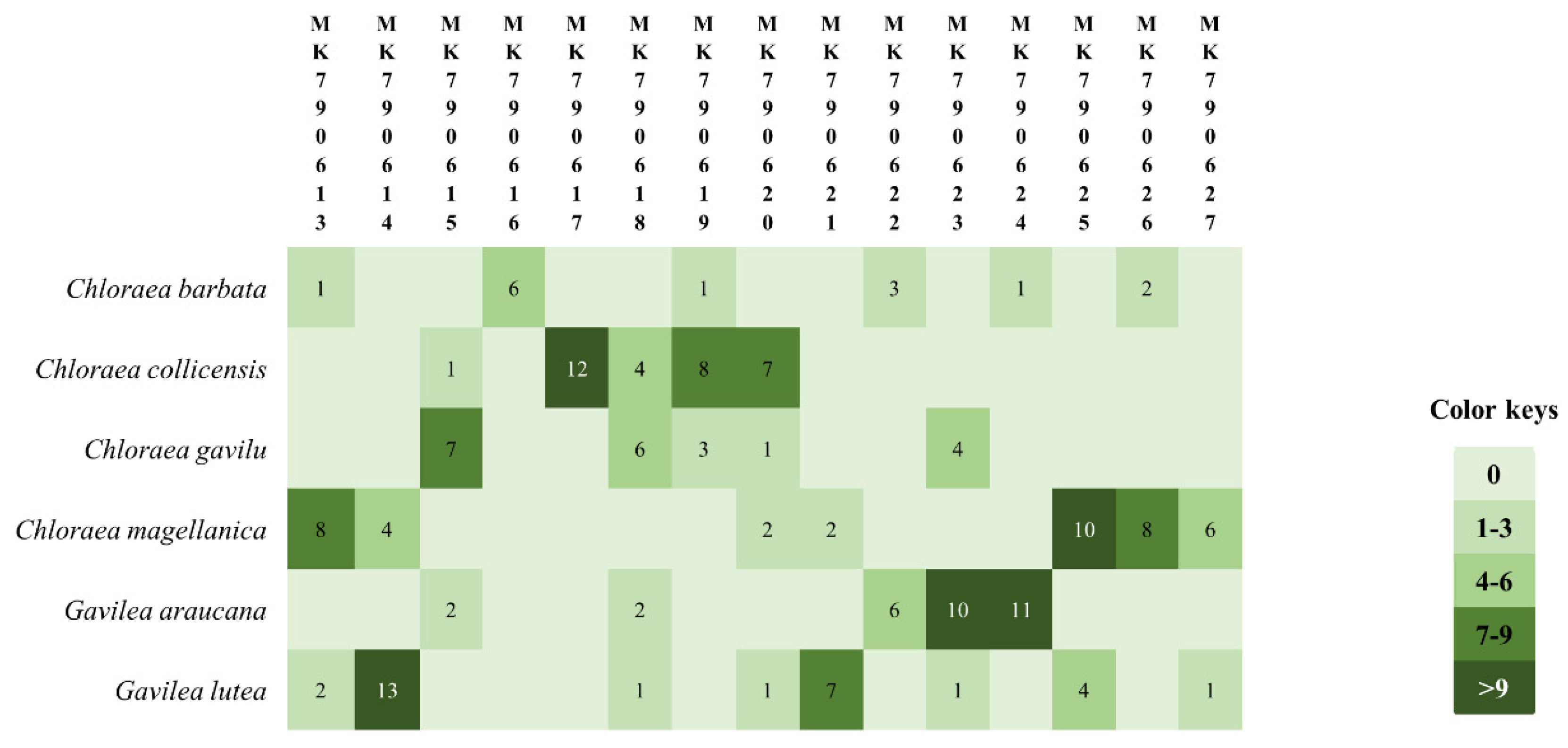


{kind=link}
{kind=link}
| Species | Location | Sample Site | Number of Root Samples |
|---|---|---|---|
| Chloraea barbata Lindl. | Imperial (38°43′31.5″ S 72°59′45.0″ W) | Grassland | 10 |
| Chloraea collicensis Kraenzl. | Cholchol (38°36′40.9″ S 72°49′16.0″ W) | Grassland | 10 |
| Chloraea gavilu Lindl. | Malalche (38°34′01.7″ S 72°56′57.3″ W) | Understorey exotic forest | 10 |
| Chloraea magellanica Hook. F. | Galletue (38°37′05.8″ S 71°26′02.4″ W) | Grassland | 10 |
| Gavilea Araucana (Phil.) M.N. Correa | Malalche (38°33′38.5″ S 72°56′19.5″ W) | Understorey native forest | 4 |
| Gavilea lutea (Pers.) M.N. Correa | Melipeuco (38°45′02.8″ S 71°36′09.8″ W) | Understorey native forest | 4 |
| Isolated Bacteria | GenBank Accession Number | Isolation Source | Close Relatives (Accession Number) | % Identity | Source | Reference |
|---|---|---|---|---|---|---|
| CM01 | MK790613 | Chloraea magellanica | Collimonas pratensis (KU311457) | 100 | Not defined | GenBank |
| GL01 | MK790614 | Gavilea lutea | Pseudomonas sp. (MK371074) | 100 | Permafrost soil | GenBank |
| CG01 | MK790615 | Chloraea gavilu | Pandoraea oxalativorans (CP011253) | 100 | Soil litter close to Oxalic sp. | Sahin et al. [36] |
| CB01 | MK790616 | Chloraea barbata | Pseudomonas koreensis (KT008003) | 100 | Rhizosphere from metallophyte plants | Benidire et al. [37] |
| CC01 | MK790617 | C. collicensis | Exiguobacterium aurantiacum (MH819695) | 100 | Coastal soil | Genbank |
| CC02 | MK790618 | C. collicensis | Dyella marensis (KF475806) | 100 | Tea roots | GenBank |
| CC03 | MK790619 | C. collicensis | Luteibacter rhizovicinus (AY785744) | 100 | Tree roots | Leigh et al. [38] |
| CC04 | MK790620 | C. collicensis | Bacillus sp.(KX959542) | 99 | Orchid mycorrhizal fungi | Novotna and Suárez [24] |
| GL02 | MK790621 | G. lutea | Pseudomonas sp. (KY849590) | 100 | Roots | GenBank |
| GA01 | MK790622 | G. araucana | Pseudomonas azotoformans (MF598585) | 100 | Sporomes of Lactarius salmonicolor | GenBank |
| GA02 | MK790623 | G. araucana | Pseudomonas sp. (MG833398) | 99 | Tailing soils | GenBank |
| GA03 | MK790624 | G. araucana | Pseudomonas azotoformans (MF598585) | 100 | Sporomes of Lactarius salmonicolor | GenBank |
| CM02 | MK790625 | C. magellanica | Chryseobacterium sp. (KC306432) | 99 | Frog skin | GenBank |
| CM03 | MK790626 | C. magellanica | Pseudomonas sp. (MH392636) | 99 | Water | GenBank |
| CM04 | MK790627 | C. magellanica | Pseudomonas costantinii (KP218045) | 100 | Pleurotus ostreatus meia | GenBank |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Herrera, H.; Sanhueza, T.; Novotná, A.; Charles, T.C.; Arriagada, C. Isolation and Identification of Endophytic Bacteria from Mycorrhizal Tissues of Terrestrial Orchids from Southern Chile. Diversity 2020, 12, 55. https://doi.org/10.3390/d12020055
Herrera H, Sanhueza T, Novotná A, Charles TC, Arriagada C. Isolation and Identification of Endophytic Bacteria from Mycorrhizal Tissues of Terrestrial Orchids from Southern Chile. Diversity. 2020; 12(2):55. https://doi.org/10.3390/d12020055
Chicago/Turabian StyleHerrera, Héctor, Tedy Sanhueza, Alžběta Novotná, Trevor C. Charles, and Cesar Arriagada. 2020. "Isolation and Identification of Endophytic Bacteria from Mycorrhizal Tissues of Terrestrial Orchids from Southern Chile" Diversity 12, no. 2: 55. https://doi.org/10.3390/d12020055