Ecology and Environmental Impact of Myriophyllum heterophyllum, an Aggressive Invader in European Waterways


Abstract
:1. Introduction
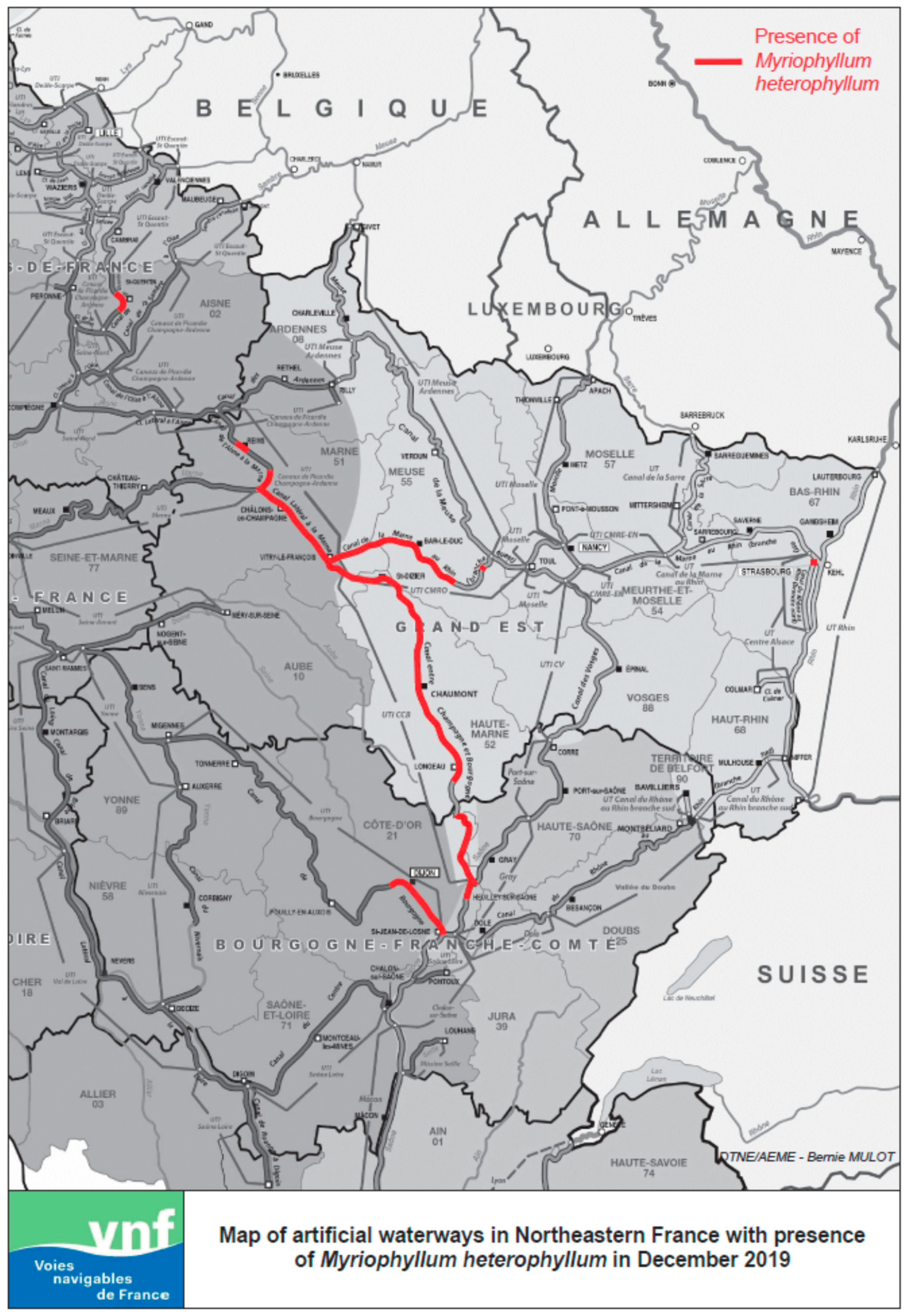
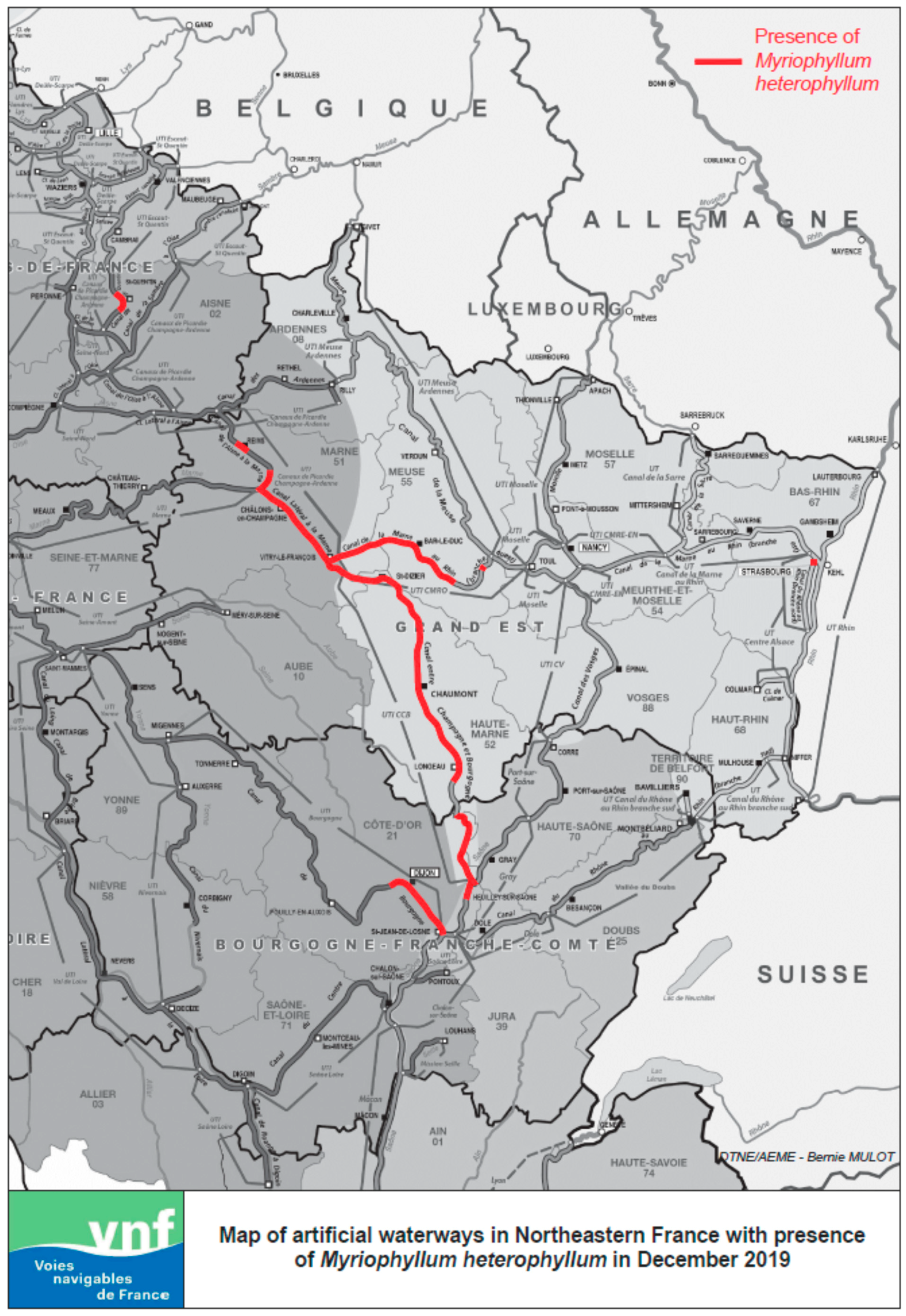
2. Origin and Invasive Spread
3. Morphological and Molecular Identification
4. Autecology and Synecology
4.1. Autecology
4.1.1. Habitat and Water Physicochemical Conditions
4.1.2. Light and Temperature Requirements
4.1.3. Nutrient Requirements
4.2. Synecology
4.2.1. Associated Plant Species and Competition
4.2.2. Herbivory
4.2.3. Pathogens and Toxic Interactions
5. Environmental Factors Influencing Growth and Dispersal
6. Regulation and Control Measures
6.1. Regulatory Demands and Challenges to Overcome
6.2. Mechanical Control
6.3. Biological Control
6.4. Chemical Control
6.5. Prevention of Spread
7. Conclusions
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Hussner, A. Alien aquatic plant species in European countries. Weed Res. 2012, 52, 297–306. [Google Scholar] [CrossRef]
- Anderson, L.; Fried, G.; Gunasekera, L.; Hussner, A.; Newman, J.; Starfinger, U.; Stiers, I.; van Valkenburg, J.; Tanner, R. Pest Risk Analysis for Myriophyllum Heterophyllum; European and Mediterranean Plant Protection Organization (EPPO): Paris, France, 2015; p. 32. [Google Scholar]
- EPPO. EPPO List of Invasive Alien Plants. Available online: https://www.eppo.int/ACTIVITIES/invasive_alien_plants/iap_lists (accessed on 17 January 2020).
- EU. Regulation (EU) No 1143/2014 of the European Parliament and of the Council of 22 October 2014 on the Prevention and Management of the Introduction and Spread of Invasive Alien Species. Available online: https://eur-lex.europa.eu/legal-content/EN/TXT/?uri=OJ:JOL_2014_317_R_0003 (accessed on 4 November 2019).
- EU. Commission Implementing Regulation (EU) 2017/1263 of 12 July 2017 Updating the List of Invasive Alien Species of Union Concern Established by Implementing Regulation (EU) 2016/1141 Pursuant to Regulation (EU) No 1143/2014 of the European Parliament and of the Council. Available online: https://op.europa.eu/en/publication-detail/-/publication/7ca17aa9-6788-11e7-b2f2-01aa75ed71a1 (accessed on 12 November 2019).
- Aiken, S.G. A conspectus of Myriophyllum (Haloragaceae) in North America. Brittonia 1981, 33, 57–69. [Google Scholar] [CrossRef]
- Chambers, P.A.; Barko, J.W.; Smith, C.S. Evaluation of invasions and declines of submersed aquatic macrophytes. J. Aquat. Plant Manag. 1993, 31, 218–220. [Google Scholar]
- Thum, R.A.; Zuellig, M.P.; Johnson, R.L.; Moody, M.L.; Vossbrinck, C. Molecular markers reconstruct the invasion history of variable leaf watermilfoil (Myriophyllum heterophyllum) and distinguish it from closely related species. Biol. Invasions 2011, 13, 1687–1709. [Google Scholar] [CrossRef]
- Les, D.H.; Mehrhoff, L.J. Introduction of nonindigenous aquatic vascular plants in Southern New England: A historical perspective. Biol. Invasions 1999, 1, 281–300. [Google Scholar] [CrossRef]
- Variable-Leaf Milfoil—Myriophyllum heterophyllum. Available online: https://www.nwcb.wa.gov/weeds/variable-leaf-milfoil (accessed on 12 January 2020).
- Canadensys: Myriophyllum heterophyllum. Available online: http://data.canadensys.net/vascan/taxon/6097?lang=en (accessed on 21 February 2020).
- Lebreton, A. Myriophyllum heterophyllum Michaux [Haloragaceae] en Haute-Vienne (Limousin, France), et situation de cette plante invasive en France et en Europe. EPPO Bull. 2013, 43, 180–192. [Google Scholar] [CrossRef]
- GBIF: Myriophyllum heterophyllum Michaux. Available online: https://www.gbif.org/species/100027408 (accessed on 21 February 2020).
- Tropicos: Myriophyllum heterophyllum. Available online: https://www.tropicos.org/nameSearch?name=myriophyllum%20heterophyllum (accessed on 21 February 2020).
- Yu, D.; Wang, D.; Li, Z.Y.; Funston, A.M. Taxonomic revision of the genus Myriophyllum (Haloragaceae) in China. Rhodora 2002, 104, 396–421. [Google Scholar]
- Jasprica, N.; Lasić, A.; Hafner, D.; Bratoš Cetinić, A. European invasion in progress: Myriophyllum heterophyllum Michx. (Haloragaceae) in Croatia. Nat. Croat. 2017, 26, 99–103. [Google Scholar] [CrossRef] [Green Version]
- Wimmer, W. Myriophyllum heterophyllum Michaux in Lower Saxony and Bremen and determination key for the vegetative stage. Florist. Rundbr. 1997, 31, 23–31. [Google Scholar]
- Myriophyllum heterophyllum—Verschiedenblättriges Tausendblatt. Available online: https://www.neobiota-austria.at/ms/neobiota-austria/neobiota_recht/neobiota_steckbriefe/tausendblatt/ (accessed on 12 January 2020).
- Myriophyllum heterophyllum Michx.—CH. Available online: https://www.infoflora.ch/de/flora/myriophyllum-heterophyllum.html. (accessed on 18 February 2020).
- Lukács, B.A.; Mesterházy, A.; Vidéki, R.; Király, G. Alien aquatic vascular plants in Hungary (Pannonian ecoregion): Historical aspects, data set and trends. Plant Biosyst. 2016, 150, 388–395. [Google Scholar] [CrossRef] [Green Version]
- Barina, Z. Flora of the Gerecse Mountains; Magyar Természettudományi Múzeum & Duna–Ipoly Nemzeti Park Igazgatóság: Budapest, Hungary, 2006; pp. 1–612. [Google Scholar]
- Cartografía y Medidas Para la Eliminación de Myriophyllum heterophyllum Michx. en Los Pozos de La Arboleda (Trapagaran, Bizkaia). Available online: https://www.researchgate.net/publication/331225278_Cartografia_y_medidas_para_la_eliminacion_de_Myriophyllum_heterophyllum_Michx_en_los_pozos_de_La_Arboleda_Trapagaran_Bizkaia (accessed on 23 March 2020).
- Cirujano, S.; Medina, L.; Stübing, G.; Peris, J.B. Myriophyllum heterophyllum Michx. (Haloragaceae), naturalized in Spain. An. Jard. Bot. Madr. 1997, 55, 164–165. [Google Scholar]
- Servei de Vida Silvestre. Actuaciones de Control de Especies Invasoras. In Informe Técnico 04/2018: Technical Report from Direcció General de Medi Natural i d’Avaluació Ambiental; Memoria Anual 2017; Generalitat Valenciana: Valencia, Spain, 2018. [Google Scholar]
- De Beer, D.; De Vlaeminck, R. Myriophyllum heterophyllum, een nieuwe invasieve waterplant (English: A new invasive aquatic plant). Dumortiera 2008, 94, 8–13. [Google Scholar]
- Myriophyllum heterophyllum. Available online: https://waarnemingen.be/species/17153/ (accessed on 14 March 2020).
- EPPO. Myriophyllum heterophyllum Michaux. EPPO Bull. 2016, 46, 20–24. [Google Scholar] [CrossRef] [Green Version]
- Myriophyllum heterophyllum Michx.—NL. Available online: https://www.verspreidingsatlas.nl/5500 (accessed on 18 February 2020).
- Peeters, G.M.T. Een vreemd vederkruid in het Zwartwater (English: An unusual water-milfoil at the Zwartwater pond): Myriophyllum heterophyllum Michx. Nat. Hist. Maandbl. 2004, 93, 251–252. [Google Scholar]
- Myriophyllum heterophyllum—Q-Bank. Available online: https://q-bankplants.eu/Biolomics.aspx?Table=Plants%20-%20Specimens (accessed on 18 February 2020).
- FloraWeb. Verbreitung der Farn- und Blütenpflanzen in Deutschland; Aggregiert im Raster der Topographischen Karte 1:25000. Datenstand 2013/Verbreitungsatlas. Available online: http://www.floraweb.de (accessed on 12 January 2020).
- Hussner, A.; Nienhaus, I.; Krause, T. Zur Verbreitung von Myriophyllum heterophyllum Michx. in Nordrhein-Westfalen. Flor. Rundbr. 2005, 39, 113–120. [Google Scholar]
- Myriophyllum heterophyllum—FR. Available online: http://especes-exotiques-envahissantes.fr/espece/myriophyllum-heterophyllum/ (accessed on 12 January 2020).
- Saint-Val, M. Myriophyllum heterophyllum. 2019. Available online: http://cbnbp.mnhn.fr/cbnbp/ressources/alaloupe/Myriophyllum_heterophyllum_Mathieu_Saint-Val.pdf (accessed on 11 March 2020).
- Moody, M.L.; Les, D.H. Systematics of the aquatic angiosperm genus Myriophyllum (Haloragaceae). Syst. Bot. 2010, 35, 121–139. [Google Scholar] [CrossRef]
- Thum, R.A.; Lennon, J.T.; Connor, J.; Smagula, A.P. A DNA fingerprinting approach for distinguishing native and non-native milfoils. Lake Reserv. Manag. 2006, 22, 1–6. [Google Scholar] [CrossRef] [Green Version]
- Moody, M.L.; Les, D.H. Evidence of hybridity in invasive watermilfoil (Myriophyllum) populations. Proc. Natl. Acad. Sci. USA 2002, 99, 14867–14871. [Google Scholar] [CrossRef] [Green Version]
- Tavalire, H.F.; Bugbee, G.E.; LaRue, E.A.; Thum, R.A. Hybridization, cryptic diversity, and invasiveness in introduced variable-leaf watermilfoil. Evol. Appl. 2012, 5, 892–900. [Google Scholar] [CrossRef]
- Thum, R.A.; Lennon, J.T. Is hybridization responsible for invasive growth of non-indigenous water-milfoils? Biol. Invasions 2006, 8, 1061–1066. [Google Scholar] [CrossRef]
- Environmental Fact Sheet—Variable Watermilfoil Myriophyllum heterophyllum. 2019. Available online: https://www.des.nh.gov/organization/commissioner/pip/factsheets/bb/documents/bb-23.pdf (accessed on 12 January 2020).
- GISD. Species Profile Myriophyllum heterophyllum. 2015. Available online: http://www.iucngisd.org/gisd/species.php?sc=1700 (accessed on 17 February 2020).
- Myriophyllum heterophyllum. Available online: http://alienplantsbelgium.be/content/myriophyllum-heterophyllum (accessed on 12 January 2020).
- Sculthorpe, C.D. The Biology of Aquatic Vascular Plants; Edward Arnold Publishers: London, UK, 1967; pp. 1–610. [Google Scholar]
- Zepeda, C.G.; Lot, A.; Antonio Nemiga, X.; Manjarrez, J. Seed bank and established vegetation in the last remnants of the Mexican Central Plateau wetlands: The Lerma marshes. Rev. Biol. Trop. 2014, 62, 455–472. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Heidbuechel, P.; Sachs, M.; Stanik, N.; Hussner, A. Species-specific fragmentation rate and colonization potential partly explain the successful spread of aquatic plants in lowland streams. Hydrobiologia 2019, 843, 107–123. [Google Scholar] [CrossRef]
- Pietsch, W.; Jentsch, H. Zur Soziologie und Ökologie von Myriophyllum heterophyllum Mich. in Mitteleuropa. Gleditschia 1984, 12, 303–335. [Google Scholar]
- June-Wells, M.; Gallagher, F.; Gibbons, J.; Bugbee, G. Water chemistry preferences of five nonnative aquatic macrophyte species in Connecticut: A preliminary risk assessment tool. Lake Reserv. Manag. 2013, 29, 303–316. [Google Scholar] [CrossRef] [Green Version]
- Thum, R.A.; Lennon, J.T. Comparative ecological niche models predict the invasive spread of variable-leaf milfoil (Myriophyllum heterophyllum) and its potential impact on closely related native species. Biol. Invasions 2010, 12, 133–143. [Google Scholar] [CrossRef]
- Hussner, A. Ökologische und Ökophysiologische Charakteristika Aquatischer Neophyten in Nordrhein-Westfalen. Ph.D. Thesis, Biology, Heinrich-Heine University, Düsseldorf, Germany, 2008; pp. 1–192. [Google Scholar]
- Hussner, A.; Jahns, P. European native Myriophyllum spicatum showed a higher HCO3− use capacity than alien invasive Myriophyllum heterophyllum. Hydrobiologia 2015, 746, 171–182. [Google Scholar] [CrossRef]
- Hussner, A.; Hoelken, H.P.; Jahns, P. Low light acclimated submerged freshwater plants show a pronounced sensitivity to increasing irradiances. Aquat. Bot. 2010, 93, 17–24. [Google Scholar] [CrossRef]
- Gross, E.M. Impact of reoligotrophication in Lake Constance on nutrient content and defensive phenolic compounds in Eurasian watermilfoil. Verh. Int. Verein Limnol. 2009, 30, 897–899. [Google Scholar] [CrossRef]
- Fornoff, F.; Gross, E. Induced defense mechanisms in an aquatic angiosperm to insect herbivory. Oecologia 2014, 175, 173–185. [Google Scholar] [CrossRef]
- Hilt, S.; Köhler, J.; Kozerski, H.-P.; van Nes, E.H.; Scheffer, M. Abrupt regime shifts in space and time along rivers and connected lake systems. Oikos 2011, 120, 766–775. [Google Scholar] [CrossRef]
- Hilt, S.; Gross, E.M. Can allelopathically active submerged macrophytes stabilise clear-water states in shallow lakes? Basic Appl. Ecol. 2008, 9, 422–432. [Google Scholar] [CrossRef] [Green Version]
- Bugbee, G.J.; White, J.C.; Krol, W.J. Control of variable watermilfoil in Bashan Lake, CT with 2,4-D: Monitoring of lake and well water. J. Aquat. Plant Manag. 2003, 41, 18–25. [Google Scholar]
- Taiz, L.; Zeiger, E.; Moller, I.M.; Murphy, A. Plant Physiology and Development, 6th ed.; Sinauer: Sunderland, MA, USA, 2014; pp. 1–761. [Google Scholar]
- Dülger, E.; Heidbüchel, P.; Schumann, T.; Mettler-Altmann, T.; Hussner, A. Interactive effects of nitrate concentrations and carbon dioxide on the stoichiometry, biomass allocation and growth rate of submerged aquatic plants. Freshw. Biol. 2017, 62, 1094–1104. [Google Scholar] [CrossRef]
- Banks, L.K.; Frost, P.C. Biomass loss and nutrient release from decomposing aquatic macrophytes: Effects of detrital mixing. Aquat. Sci. 2017, 79, 881–890. [Google Scholar] [CrossRef]
- Demars, B.O.L.; Edwards, A.C. Tissue nutrient concentrations in freshwater aquatic macrophytes: High inter-taxon differences and low phenotypic response to nutrient supply. Freshw. Biol. 2007, 52, 2073–2086. [Google Scholar] [CrossRef]
- Dülger, E.; Hussner, A. Differences in the growth and physiological response of eight Myriophyllum species to carbon dioxide depletion. Aquat. Bot. 2017, 139, 25–31. [Google Scholar] [CrossRef]
- Beer, S.; Wetzel, R.G. Photosynthesis in submersed macrophytes of a temperate lake. Plant Physiol. 1982, 70, 488–492. [Google Scholar] [CrossRef]
- Hempel, M.; Grossart, H.P.; Gross, E.M. Community composition of bacterial biofilms on two submerged macrophytes and an artificial substrate in a pre-alpine lake. Aquat. Microb. Ecol. 2009, 58, 79–94. [Google Scholar] [CrossRef]
- Casper, S.J.; Jentsch, H.; Gutte, P. Beiträge zur Taxonomie und Chorologie europäischer Wasser-und Sumpfpflanzen. Hercynia 1980, 17, 365–374. [Google Scholar]
- Hussner, A.; van Dam, H.; Vermaat, J.E.; Hilt, S. Comparison of native and neophytic aquatic macrophyte developments in a geothermally warmed river and thermally normal channels. Fund. Appl. Limnol. 2014, 185, 155–165. [Google Scholar] [CrossRef]
- Greulich, S.; Bornette, G. Being evergreen in an aquatic habitat with attenuated seasonal contrasts – a major competitive advantage? Plant Ecol. 2003, 167, 9–18. [Google Scholar] [CrossRef]
- Petruzzella, A.; Manschot, J.; van Leeuwen, C.H.A.; Grutters, B.M.C.; Bakker, E.S. Mechanisms of invasion resistance of aquatic plant communities. Front. Plant Sci. 2018, 9. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Schwoerer, T.; Morton, J.M. Human dimensions of aquatic invasive species in Alaska: Lessons learned while integrating economics, management and biology to incentivize early detection and rapid response. In Alaska: Economic, Environmental, and Social Issues; Nova Science Publishers Inc.: Hauppauge, NY, USA, 2018; pp. 1–46. [Google Scholar]
- Lodge, D.M. Herbivory on freshwater macrophytes. Aquat. Bot. 1991, 41, 195–224. [Google Scholar] [CrossRef]
- Newman, R.M. Invited review—Biological control of Eurasian watermilfoil by aquatic insects: Basic insights from an applied problem. Arch. Hydrobiol. 2004, 159, 145–184. [Google Scholar] [CrossRef]
- Newman, R.M. Herbivory and detritivory on fresh-water macrophytes by invertebrates: A review. J. N. Am. Benthol. Soc. 1991, 10, 89–114. [Google Scholar] [CrossRef]
- Wimmer, W.; Sprick, P. Records of weevils (Coleoptera: Curculionidae) on Myriophyllum species, with special regard to M. heterophyllum MICHAUX, in Lower Saxony, Germany. Braunschw. Naturk. Schr. 2000, 6, 123–130. [Google Scholar]
- Newman, R.M.; Gross, E.M.; Wimmer, W.; Sprick, P. Life history and developmental performance of the Eurasian milfoil weevil, Eubrychius velutus (Coleoptera: Curculionidae). Coleopt. Bull. 2006, 60, 170–176. [Google Scholar] [CrossRef]
- Marko, M.D.; Newman, R.M.; Gleason, F.K. Chemically mediated host-plant selection by the milfoil weevil: A freshwater insect-plant interaction. J. Chem. Ecol. 2005, 31, 2857–2876. [Google Scholar] [CrossRef]
- Gross, E.M.; Johnson, R.L.; Hairston, N.G., Jr. Experimental evidence for changes in submersed macrophyte species composition caused by the herbivore Acentria ephemerella (Lepidoptera). Oecologia 2001, 127, 105–114. [Google Scholar] [CrossRef] [Green Version]
- Johnson, R.L.; Gross, E.M.; Hairston, N.G., Jr. Decline of the invasive submersed macrophyte Myriophyllum spicatum (Haloragaceae) associated with herbivory by larvae of Acentria ephemerella (Lepidoptera). Aquat. Ecol. 1998, 31, 273–282. [Google Scholar] [CrossRef] [Green Version]
- Shearer, J.F. A historical perspective of pathogen biological control of aquatic plants. Weed Technol. 2010, 24, 202–207. [Google Scholar] [CrossRef]
- Al-Sammak, M.A.; Hoagland, K.D.; Cassada, D.; Snow, D.D. Co-occurrence of the cyanotoxins BMAA, DABA and anatoxin-a in Nebraska reservoirs, fish, and aquatic plants. Toxins 2014, 6, 488–508. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kuntz, K.; Heidbuechel, P.; Hussner, A. Effects of water nutrients on regeneration capacity of submerged aquatic plant fragments. Ann. Limnol. 2014, 50, 155–162. [Google Scholar] [CrossRef]
- Heidbuechel, P.; Hussner, A. Fragment type and water depth determine the regeneration and colonization success of submerged aquatic macrophytes. Aquat. Sci. 2019, 81. [Google Scholar] [CrossRef]
- Johnson, L.E.; Ricciardi, A.; Carlton, J.T. Overland dispersal of aquatic invasiv species: A risk assessment of transient recreational boating. Ecol. Appl. 2001, 11, 1789–1799. [Google Scholar] [CrossRef]
- García-Álvarez, A.; van Leeuwen, C.H.A.; Luque, C.J.; Hussner, A.; Vélez-Martín, A.; Pérez-Vázquez, A.; Green, A.J.; Castellanos, E.M. Internal transport of alien and native plants by geese and ducks: An experimental study. Freshw. Biol. 2015, 60, 1316–1329. [Google Scholar] [CrossRef] [Green Version]
- Coughlan, N.E.; Kelly, T.C.; Davenport, J.; Jansen, M.A.K. Humid microclimates within the plumage of mallard ducks (Anas platyrhynchos) can potentially facilitate long distance dispersal of propagules. Acta Oecol. 2015, 65–66, 17–23. [Google Scholar] [CrossRef]
- Barnes, M.A.; Jerde, C.L.; Keller, D.; Chadderton, W.L.; Howeth, J.G.; Lodge, D.M. Viability of aquatic plant fragments following desiccation. Invasive Plant Sci. Manag. 2013, 6, 320–325. [Google Scholar] [CrossRef]
- Heidbuechel, P.; Jahns, P.; Hussner, A. Chlorophyll fluorometry sheds light on the role of desiccation resistance for vegetative overland dispersal of aquatic plants. Freshw. Biol. 2019, 64, 1401–1415. [Google Scholar] [CrossRef]
- Milfoil Prevention. Available online: http://www.gmcg.org/advocacy/milfoil-prevention/ (accessed on 27 February 2020).
- EASIN. Available online: https://easin.jrc.ec.europa.eu/easin (accessed on 27 February 2020).
- Faulkner, K.T.; Robertson, M.P.; Wilson, J.R.U. Stronger regional biosecurity is essential to prevent hundreds of harmful biological invasions. Glob. Chang. Biol. 2020, 26, 2449–2462. [Google Scholar] [CrossRef]
- Hussner, A.; Stiers, I.; Verhofstad, M.J.J.M.; Bakker, E.S.; Grutters, B.M.C.; Haury, J.; van Valkenburg, J.L.C.H.; Brundu, G.; Newman, J.; Clayton, J.S.; et al. Management and control methods of invasive alien freshwater aquatic plants: A review. Aquat. Bot. 2017, 136, 112–137. [Google Scholar] [CrossRef]
- Louda, S.M.; Stiling, P. The double-edged sword of biological control in conservation and restoration. Conserv. Biol. 2004, 18, 50–53. [Google Scholar] [CrossRef]
- Ray, P. Microbial agents for control of aquatic weeds and their role in integrated management. CAB Rev. 2013, 8. [Google Scholar] [CrossRef]
- Gassmann, A.; Cock, M.J.W.; Shaw, R.; Evans, H.C. The potential for biological control of invasive alien aquatic weeds in Europe: A review. Hydrobiologia 2006, 570, 217. [Google Scholar] [CrossRef]
- Mafokoane, D.L.; Oberholzer, I.G.; Hill, M.P. The biology and laboratory host range of the weevil, Listronotus marginicollis (Hustache) (Coleoptera: Curculionidae), a natural enemy of the invasive aquatic weed, parrot’s feather, Myriophyllum aquaticum (Velloso) Verde (Haloragaceae): Short communication. Afr. Entomol. 2007, 15, 385–390. [Google Scholar]
- Havel, J.E.; Knight, S.E.; Maxson, K.A. A field test on the effectiveness of milfoil weevil for controlling Eurasian watermilfoil in Wisconsin lakes. Hydrobiologia 2017, 800, 81–97. [Google Scholar] [CrossRef]
- Newman, R.M.; Inglis, W.G. Distribution and abundance of the milfoil weevil, Euhrychiopsis lecontei, in Lake Minnetonka and relation to milfoil harvesting. J. Aquat. Plant Manag. 2009, 47, 21–25. [Google Scholar]
- Ward, D.M.; Newman, R.M. Fish predation on Eurasian watermilfoil (Myriophyllum spicatum) herbivores and indirect effects on macrophytes. Can. J. Fish. Aquat. Sci. 2006, 63, 1049–1057. [Google Scholar] [CrossRef] [Green Version]
- Marko, M.D.; White, J.C. Direct comparison of herbicidal or biological treatment on Myriophyllum spicatum control and biochemistry. Front. Plant Sci. 2018, 871. [Google Scholar] [CrossRef] [Green Version]
- Havel, J.E.; Knight, S.E.; Miazga, J.R. Abundance of milfoil weevil in Wisconsin lakes: Potential effects from herbicide control of Eurasian watermilfoil. Lake Reserv. Manag. 2017, 33, 270–279. [Google Scholar] [CrossRef]
- Kasinak, J.-M.E.; Bishop, C.J.; Wright, R.A.; Wilson, A.E. Grass carp do not consume the nuisance benthic cyanobacterium, Lyngbya wollei. J. Aquat. Plant Manag. 2015, 53, 74–80. [Google Scholar]
- Yu, J.; Zhen, W.; Guan, B.; Zhong, P.; Jeppesen, E.; Liu, Z. Dominance of Myriophyllum spicatum in submerged macrophyte communities associated with grass carp. Knowl. Manag. Aquat. Ecosyst. 2016. [Google Scholar] [CrossRef] [Green Version]
- Shearer, J.F.; Durham, B.D.; Harms, N. Screening of biological control pathogens isolated from Eurasian watermilfoil. J. Aquat. Plant Manag. 2011, 49, 118–121. [Google Scholar]
- Verma, U.; Charudattan, R. Host range of Mycoleptodiscus terrestris, a microbial herbicide candidate for Eurasian Watermilfoil, Myriophyllum spicatum. Biol. Control 1993, 3, 271–280. [Google Scholar] [CrossRef]
- Getsinger, K.D.; Netherland, M.D.; Grue, C.E.; Koschnick, T.J. Improvements in the use of aquatic herbicides and establishment of future research directions. J. Aquat. Plant Manag. 2008, 46, 32–41. [Google Scholar]
- Beets, J.; Heilman, M.; Netherland, M.D. Large-scale mesocosm evaluation of florpyrauxifen-benzyl, a novel arylpicolinate herbicide, on Eurasian and hybrid watermilfoil and seven native submersed plants. J. Aquat. Plant Manag. 2019, 57, 49–55. [Google Scholar]
- Parks, S.R.; McNair, J.N.; Hausler, P.; Tyning, P.; Thum, R.A. Divergent responses of cryptic invasive watermilfoil to treatment with auxinic herbicides in a large Michigan lake. Lake Reserv. Manag. 2016, 32, 366–372. [Google Scholar] [CrossRef]
- Glomski, L.M.; Netherland, M.D. Use of a small-scale primary screening method to predict effects of flumioxazin and carfentrazone-ethyl on native and invasive, submersed plants. J. Aquat. Plant Manag. 2013, 51, 45–48. [Google Scholar]
- Hussner, A.; Nehring, S.; Hilt, S. From first reports to successful control: A plea for improved management of alien aquatic plant species in Germany. Hydrobiologia 2014, 737, 321–331. [Google Scholar] [CrossRef]
- Cabomba and Myriophyllum in Trade: What’s in a Name? Robson Meeting Proceedings 2005. Available online: http://www.robsonmeeting.org/ (accessed on 28 February 2020).
- Ghahramanzadeh, R.; Esselink, G.; Kodde, L.P.; Duistermaat, H.; Van Valkenburg, J.; Marashi, S.H.; Smulders, M.J.M.; Van De Wiel, C.C.M. Efficient distinction of invasive aquatic plant species from non-invasive related species using DNA barcoding. Mol. Ecol. Resour. 2013, 13, 21–31. [Google Scholar] [CrossRef]
- Bruckerhoff, L.; Havel, J.; Knight, S. Survival of invasive aquatic plants after air exposure and implications for dispersal by recreational boats. Hydrobiologia 2015, 746, 113–121. [Google Scholar] [CrossRef]
- Johnstone, I.M.; Coffey, B.T.; Howard-Williams, C. The role of recreational boat traffic in interlake dispersal of macrophytes: A New Zealand case study. J. Environ. Manag. 1985, 20, 263–279. [Google Scholar]
- CABI. Datasheet Myriophyllum heterophyllum (Broadleaf Watermilfoil). Available online: https://www.cabi.org/isc/datasheet/34940#REF-DDB-27 (accessed on 14 March 2020).
- Stop Aquatic Hitchhikers! Available online: https://www.fws.gov/fisheries/ans/pdf_files/Stop_Aquatic_Hitchhikers_factsheet.pdf (accessed on 26 February 2020).
- Stop Aquatic Hitchhikers. Available online: http://www.env.gov.bc.ca/fw/fish/regulations/docs/1011/fa_AquaticHitchhikers.pdf (accessed on 26 February 2020).
- Check, Clean, Dry. Available online: http://www.nonnativespecies.org/checkcleandry/ (accessed on 26 February 2020).
- Be Plant Wise. Available online: http://www.nonnativespecies.org/beplantwise/ (accessed on 26 February 2020).
- Hippolite, J.; Te Kurapa, T.W.M. Aquatic Pest Report 2018; Bay of Plenty Regional Council, Ed.; Environmental Publication: Whakatane, New Zealand, 2018. [Google Scholar]
- Lake Stewards of Maine. Available online: https://www.lakestewardsofmaine.org/ (accessed on 27 February 2020).
- Bleu Laurentides. Available online: https://crelaurentides.org/dossiers/eau-lacs/bleu-laurentides (accessed on 27 February 2020).





{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Channel | Statistical Details | pH | Alk. (mM) | Cond. (µS cm−1) | N-NO2 (µg L−1) | N-NO3 (mg L−1) | N-NH4 (µg L−1) | P-PO4 µg L−1) | N/P (mol mol−1) | Cl (mg L−1) | SO4 (mg L−1) | Ca (mg L−1) | Mg (mg L−1) | Na (mg L−1) | K (mg L−1) |
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| CMRo | mean | 8.23 | 2.03 | 436 | 13.39 | 3.26 | 17.11 | 1.57 | 8343 | 6.30 | 11.92 | 80.53 | 5.02 | 2.80 | 2.54 |
| SD | 0.14 | 0.07 | 15 | 2.72 | 0.13 | 6.51 | 1.23 | 7958 | 3.99 | 0.30 | 3.40 | 0.12 | 0.79 | 3.70 | |
| min | 8.11 | 1.97 | 420 | 9.13 | 3.05 | 7.78 | 0.33 | 2085 | 4.10 | 11.60 | 77.64 | 4.85 | 2.20 | 0.73 | |
| max | 8.45 | 2.15 | 451 | 15.22 | 3.36 | 23.33 | 3.59 | 22099 | 13.40 | 12.40 | 86.39 | 5.13 | 4.15 | 9.15 | |
| CCB | mean | 8.32 | 1.72 | 431 | 12.17 | 2.51 | 10.42 | 5.42 | 1088 | 24.40 | 13.40 | 64.77 | 6.28 | 5.92 | 16.81 |
| SD | 0.04 | 0.18 | 26 | 4.81 | 0.25 | 6.59 | 1.90 | 188 | 5.29 | 0.32 | 6.07 | 3.62 | 2.61 | 3.87 | |
| min | 8.27 | 1.56 | 402 | 6.09 | 2.38 | 3.11 | 4.57 | 752 | 17.40 | 13.00 | 59.87 | 3.51 | 4.48 | 10.50 | |
| max | 8.37 | 1.96 | 470 | 18.26 | 2.96 | 18.67 | 8.81 | 1188 | 31.25 | 13.71 | 74.50 | 12.61 | 10.58 | 19.99 |
| Channel | Statistical Details | N-NO2 (µg L−1) | N-NO3 (mg L−1) | N-NH4 (µg L−1) | P-PO4 (µg L−1) | N/P (mol mol−1) | Cl (mg L−1) | SO4 (mg L−1) |
|---|---|---|---|---|---|---|---|---|
| CMRo | mean | 9.13 | 45.16 | 556.11 | 12.07 | 177 | 6.28 | 9.08 |
| SD | 10.25 | 31.93 | 128.04 | 7.70 | 147 | 2.16 | 4.57 | |
| min | 3.04 | 22.58 | 474.44 | 2.94 | 65 | 4.50 | 3.20 | |
| max | 24.35 | 90.32 | 746.67 | 18.93 | 389 | 9.40 | 13.30 | |
| CCB | mean | 7.30 | 364.45 | 239.24 | 8.03 | 225 | 8.68 | 13.08 |
| SD | 5.09 | 594.04 | 258.71 | 4.01 | 274 | 1.90 | 1.86 | |
| min | 3.04 | 54.19 | 26.44 | 4.57 | 57 | 5.91 | 10.06 | |
| max | 15.22 | 1422.58 | 688.33 | 13.38 | 710 | 10.60 | 14.88 |
| Channel | Statistical Details | C (mg g−1) | N (mg g−1) | P (mg g−1) | C/P (mol mol−1) | C/N (mol mol−1) | N/P (mol mol−1) | DMC (%) |
|---|---|---|---|---|---|---|---|---|
| CMRo | mean | 376.70 | 38.40 | 1.66 | 612.59 | 11.65 | 52.67 | 10.12 |
| SD | 30.82 | 7.21 | 0.38 | 146.70 | 1.39 | 10.58 | 3.00 | |
| min | 324.70 | 31.30 | 1.17 | 430.64 | 8.48 | 33.47 | 6.86 | |
| max | 432.00 | 55.50 | 2.24 | 841.53 | 13.17 | 69.72 | 17.83 | |
| CCB | mean | 374.55 | 38.00 | 1.98 | 566.58 | 11.72 | 47.65 | 7.37 |
| SD | 25.06 | 6.21 | 0.84 | 227.46 | 1.81 | 14.32 | 1.55 | |
| min | 324.60 | 29.80 | 1.09 | 255.47 | 8.90 | 22.12 | 5.81 | |
| max | 408.80 | 52.30 | 3.72 | 935.67 | 15.36 | 69.25 | 10.45 |
| Channel | Statistical Details | Chl a (mg g−1) | Chl b (mg g−1) | Chl a+b (mg g−1) | Chl a/b (mg mg−1) | Car (mg g−1) | Acy (mg g−1) | TPC (mg g−1) |
|---|---|---|---|---|---|---|---|---|
| CMRo | mean | 8.09 | 2.60 | 10.69 | 3.13 | 1.46 | 0.88 | 30.83 |
| SD | 1.27 | 0.43 | 1.63 | 0.30 | 0.32 | 0.69 | 8.28 | |
| min | 5.93 | 1.94 | 7.86 | 2.68 | 0.96 | 0.07 | 15.84 | |
| max | 10.43 | 3.38 | 13.37 | 3.54 | 2.05 | 2.10 | 39.64 | |
| CCB | mean | 8.46 | 2.89 | 11.36 | 3.03 | 1.49 | 0.76 | 59.73 |
| SD | 1.60 | 0.93 | 2.50 | 0.44 | 0.26 | 0.49 | 24.84 | |
| min | 7.07 | 2.05 | 9.12 | 2.51 | 1.02 | 0.08 | 31.30 | |
| max | 12.23 | 4.87 | 17.10 | 3.52 | 1.84 | 1.62 | 112.21 |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Gross, E.M.; Groffier, H.; Pestelard, C.; Hussner, A. Ecology and Environmental Impact of Myriophyllum heterophyllum, an Aggressive Invader in European Waterways. Diversity 2020, 12, 127. https://doi.org/10.3390/d12040127
Gross EM, Groffier H, Pestelard C, Hussner A. Ecology and Environmental Impact of Myriophyllum heterophyllum, an Aggressive Invader in European Waterways. Diversity. 2020; 12(4):127. https://doi.org/10.3390/d12040127
Chicago/Turabian StyleGross, Elisabeth M., Hélène Groffier, Cécile Pestelard, and Andreas Hussner. 2020. "Ecology and Environmental Impact of Myriophyllum heterophyllum, an Aggressive Invader in European Waterways" Diversity 12, no. 4: 127. https://doi.org/10.3390/d12040127
APA StyleGross, E. M., Groffier, H., Pestelard, C., & Hussner, A. (2020). Ecology and Environmental Impact of Myriophyllum heterophyllum, an Aggressive Invader in European Waterways. Diversity, 12(4), 127. https://doi.org/10.3390/d12040127