Patterns of Distribution of Phoretic Deutonymphs of Uropodina on Longhorn Beetles in Białowieża Primeval Forest, Central Europe


Abstract
:
1. Introduction
2. Materials and Methods
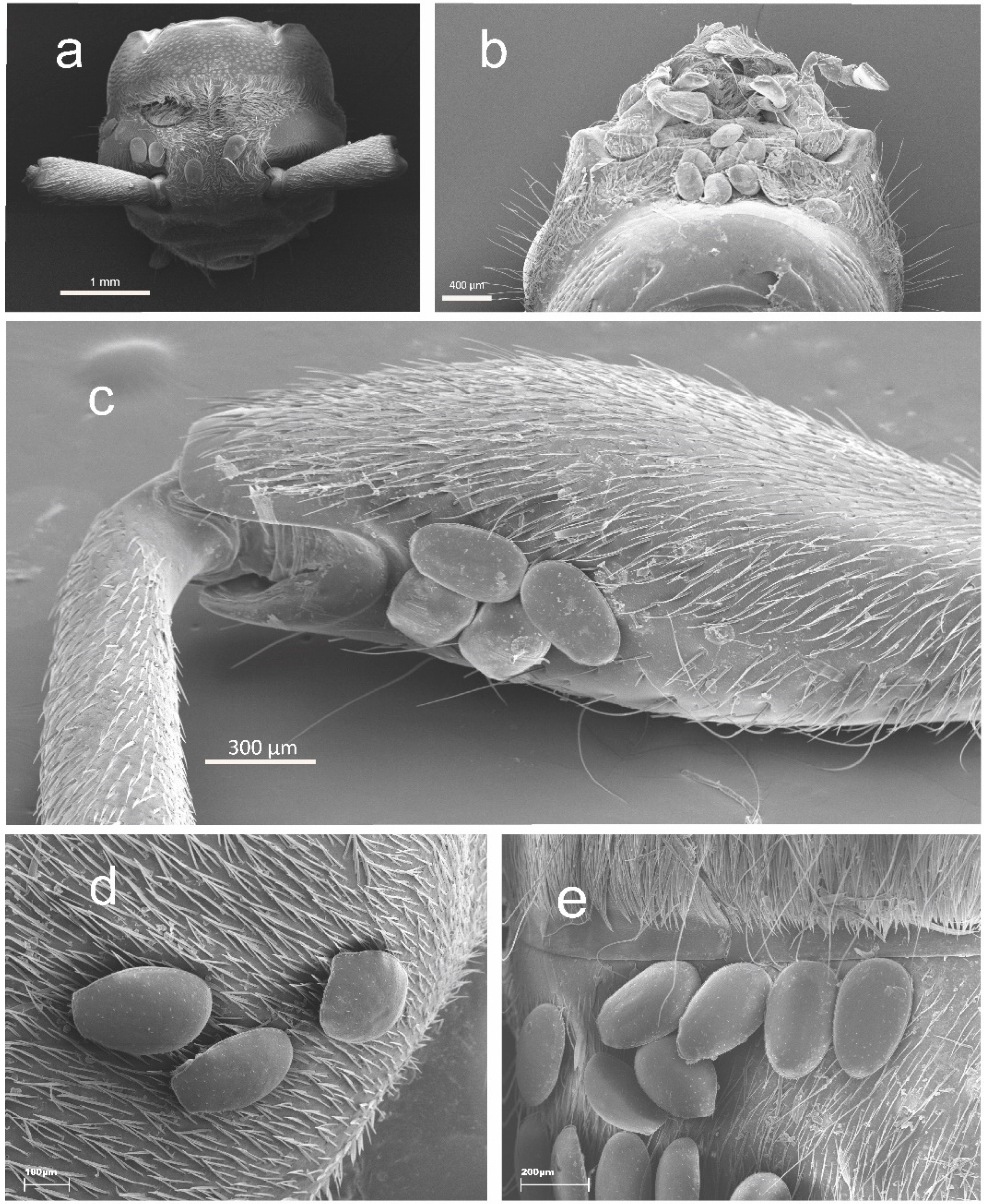
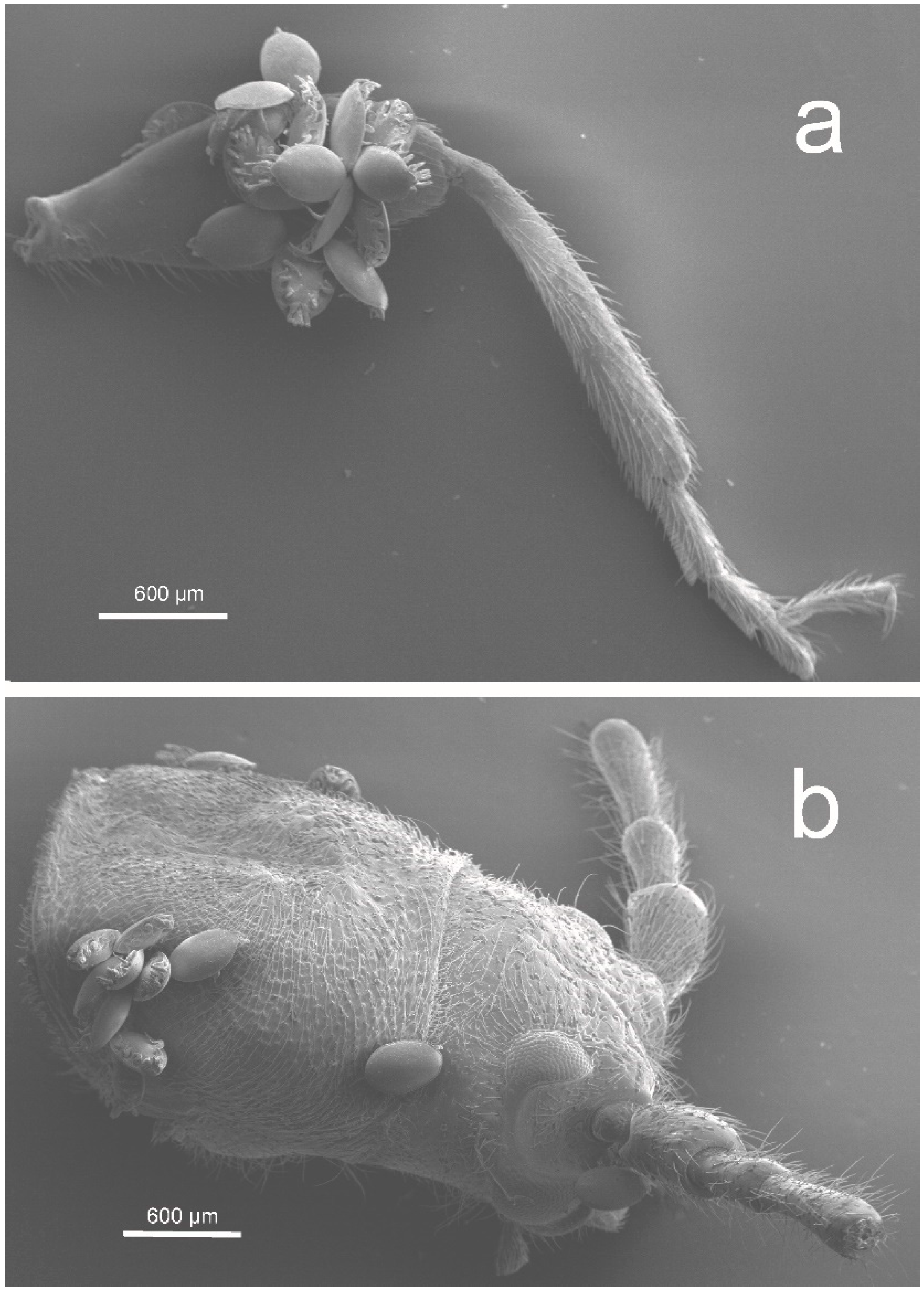
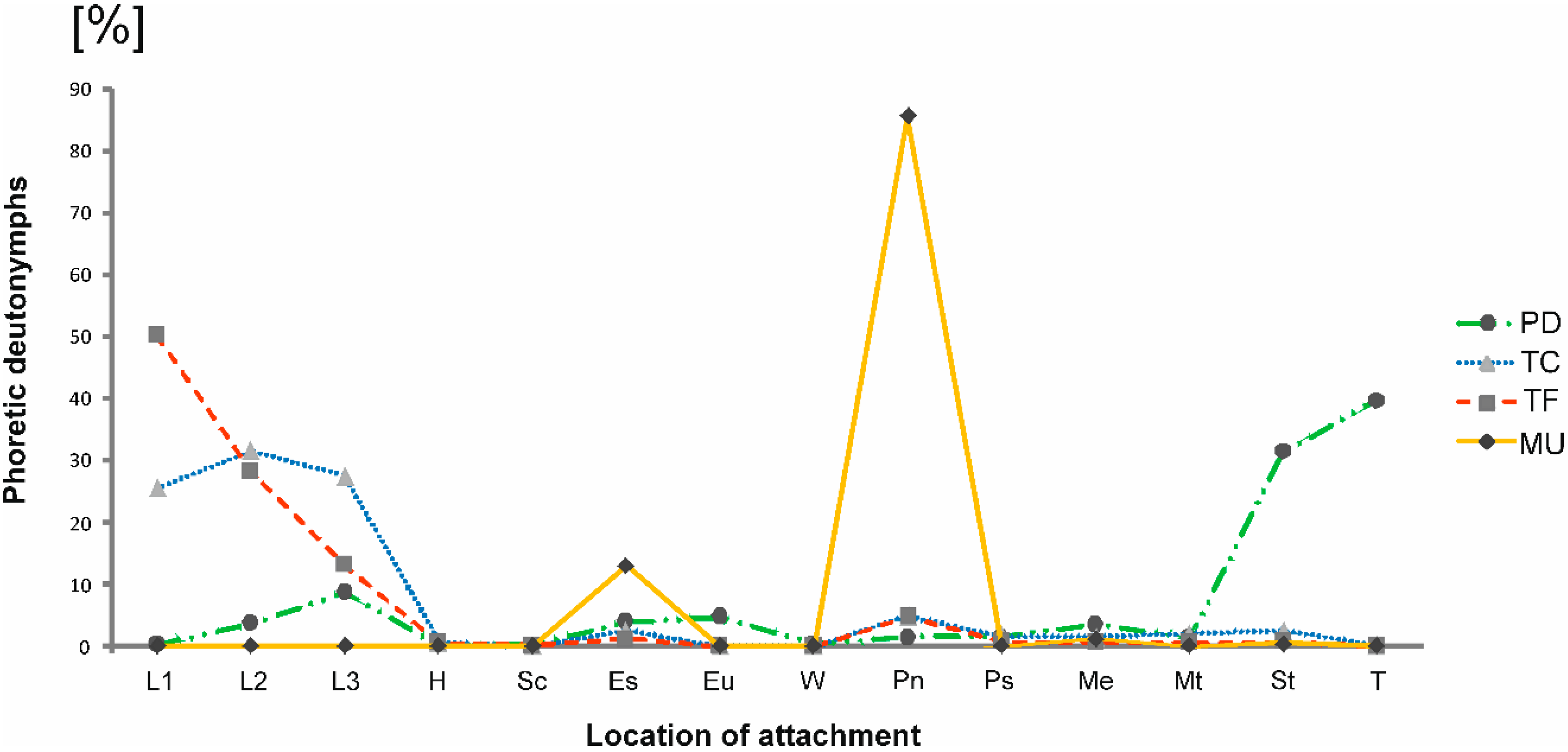
3. Results
3.1. Distribution of Oodinychus ovalis on Monochamus sartor urussovii
3.2. Distribution of Trichouropoda sociata on Plagionotus detritus
3.3. Distribution of Trichouropoda shcherbakae on Tetropium castaneum
3.4. Distribution of Trichouropoda shcherbakae on Tetropium fuscum
4. Discussion
5. Conclusions
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Walter, D.E.; Proctor, H.C. Mites: Ecology, Evolution & Behaviour. Life at a Microscale, 2nd ed.; Springer: Berlin/Heidelberg, Germany, 2013; pp. 1–494. [Google Scholar]
- Bajerlein, D.; Witaliński, W. Anatomy and fine structure of pedicellar glands in phoretic deutonymphs of uropodid mites (Acari: Mesostigmata). Arthropod Struct. Dev. 2012, 41, 245–257. [Google Scholar] [CrossRef] [PubMed]
- Bajerlein, D.; Witaliński, W.; Adamski, Z. Morphological diversity of pedicels in phoretic deutonymphs of Uropodina mites (Acari: Mesostigmata). Arthropod Struct. Dev. 2013, 42, 185–196. [Google Scholar] [CrossRef] [PubMed]
- Athias-Binche, F. Dispersal in varying environments: The case of phoretic uropodid mites. Can. J. Zool. 1993, 71, 1793–1798. [Google Scholar] [CrossRef]
- Hunter, P.E.; Rosario, R.M.T. Associations of Mesostigmata with other arthropods. Annu. Rev. Entomol. 1988, 33, 393–417. [Google Scholar] [CrossRef]
- Vitzthum, H. Acari als Commensalen von Ipiden. (Der Acarologischen Beobachtungen 11. Reihe.). Zool. Jahrb. Abt. Syst. Geogr. Biol. Tiere 1926, 52, 407–503. [Google Scholar]
- Gwiazdowicz, D.J. Mesostigmatid mites (Acari) associated with Scolytidae in Poland. In Selected Problems of Acarological Research in Forests, 1st ed.; Gwiazdowicz, D.J., Ed.; Wydawnictwo Uniwersytetu Przyrodniczego w Poznaniu: Poznań, Poland, 2008; pp. 59–95. [Google Scholar]
- Moser, J.C.; Konrad, H.; Blomquist, S.R.; Kirisits, T. Do mites phoretic on elm bark beetles contribute to the transmission of Dutch elm disease? Naturwissenschaften 2009, 97, 219–227. [Google Scholar] [CrossRef] [Green Version]
- Gwiazdowicz, D.J.; Kamczyc, J.; Teodorowicz, E.; Błoszyk, J. Mite communities (Acari, Mesostigmata) associated with Ips typographus (Coleoptera, Scolytidae) in managed and natural Norway spruce stands in Central Europe. Open Life Sci. 2012, 7, 910–916. [Google Scholar] [CrossRef] [Green Version]
- Gwiazdowicz, D.J.; Błoszyk, J.; Gdula, A.K. Alpha diversity of mesostigmatid mites associated with the bark beetle Ips typographus (L.) in Poland. Insect Conserv. Divers. 2015, 8, 448–455. [Google Scholar] [CrossRef]
- Hofstetter, R.W.; Moser, J.C.; Blomquist, S.R. Mites associated with bark beetles and their hyperphoretic ophiostomatoid fungi. In The Ophiostomatoid Fungi: Expanding Frontiers, 1st ed.; Seifert, K.A., de Beer, Z.W., Wingfield, M.J., Eds.; CBS-KNAW Biodiversity Centre: Utrecht, The Netherlands, 2014; pp. 165–176. [Google Scholar]
- Chaires-Grijalva, M.P.; Estrada-Venegas, E.G.; Equihua-Martínez, A.; Moser, J.C.; Blomquist, S.R. Trophic habits of mesostigmatid mites associated with bark beetles in Mexico. J. Acarol. Soc. Jpn. 2016, 25, 161–167. [Google Scholar] [CrossRef]
- Khaustov, A.A.; Trach, V.; Bobylev, A.N. Mites (Acari) phoretic on six-toothed spruce bark beetle, Pityogenes chalcographus Linnaeus (Coleoptera: Curculionidae: Scolytinae), in Western Siberia, Russia. Acarina 2016, 24, 137–151. [Google Scholar] [CrossRef] [Green Version]
- Zach, P.; Kršiak, B.; Kulfan, J.; Parák, M.; Kontschán, J. Mites Trichouropoda and Uroobovella spp. (Uropodoidea) phoretic on bark beetles (Scolytinae): A comparison from a declining mountain spruce forest in Central Europe. Int. J. Acarol. 2016, 42, 212–217. [Google Scholar] [CrossRef]
- Khaustov, A.A.; Klimov, P.B.; Trach, V.; Bobylev, A.N.; Salavatulin, V.M.; Khaustov, V.A.; Tolstikov, A.V. Review of mites (Acari) associated with the European spruce bark beetle, Ips typographus (Coleoptera: Curculionidae: Scolytinae) in Asian Russia. Acarina 2018, 26, 3–79. [Google Scholar] [CrossRef]
- Paraschiv, M.; Martínez-Ruiz, C.; Fernández, M.M. Dynamic associations between Ips sexdentatus (Coleoptera: Scolytinae) and its phoretic mites in a Pinus pinaster forest in northwest Spain. Exp. Appl. Acarol. 2018, 75, 369–381. [Google Scholar] [CrossRef] [PubMed]
- Kontschán, J.; Szőcs, G.; Kiss, B.; Khaustov, A.A. Bark beetle associated trematurid mites (Acari: Uropodina: Trematuridae) from Asian Russia with description of a new species. Syst. Appl. Acarol. 2019, 24, 1592–1603. [Google Scholar] [CrossRef] [Green Version]
- Vissa, S.; Hofstetter, R.W.; Bonifácio, L.; Khaustov, A.; Knee, W.; Uhey, D.A. Phoretic mite communities associated with bark beetles in the maritime and stone pine forests of Setúbal, Portugal. Exp. Appl. Acarol. 2019, 77, 117–131. [Google Scholar] [CrossRef]
- Hofstetter, R.W.; Vissa, S.; Moser, J.; Bonifácio, L. Interactions among mites, host trees, fungi and beetles. In Interactions behind the Pine Wilt Disease (FCT NEMATRANSFER Project—PTDC/AGR-FOR/4391/2014), Proceedings of the Interactions behind the Pine Wilt Disease, Oeiras, Portugal, 21 June 2018; Bonifácio, L., Henriques, J., Eds.; Instituto Nacional de Investigação Agrária e Veterinária: Oeiras, Portugal, 2018; pp. 31–37. [Google Scholar]
- Gwiazdowicz, D.J.; Gutowski, J. Records of phoretic mesostigmatid mites (Acari: Mesostigmata) on beetles (Coleoptera: Carabidae, Cerambycidae, Elateridae, Erotylidae, Scolytinae) in the Białowieża Primeval Forest. Pol. J. Èntomol. 2012, 81, 305–310. [Google Scholar] [CrossRef]
- Błoszyk, J.; Gutowski, J.; Gwiazdowicz, D.J.; Mądra-Bielewicz, A. Phoresy of Trichouropoda shcherbakae Hirschmann, 1972 (Acari: Mesostigmata) on beetles of the genus Tetropium Kirby, 1837 (Coleoptera: Cerambycidae) in Białowieża Forest, Poland. Open Life Sci. 2013, 8, 986–992. [Google Scholar] [CrossRef]
- Gwiazdowicz, D.J.; Gutowski, J.M.; Kamczyc, J.; Teodorowicz, E. Phoretic relationships between Plagionotus detritus (Coleoptera: Cerambycidae) and Trichouropoda sociata (Acari: Mesostigmata). Èntomol. Fenn. 2013, 24, 59–64. [Google Scholar] [CrossRef] [Green Version]
- Błoszyk, J.; Gutowski, J.M.; Gwiazdowicz, D.J.; Mądra-Bielewicz, A.; Konwerski, S.; Książkiewicz, Z. Monochamus sartor (Coleoptera: Cerambycidae) contributes to alpha diversity of Uropodina mites (Acari: Mesostigmata) in first stage of wood decay in Białowieża Primeval Forest. Int. J. Acarol. 2016, 42, 218–223. [Google Scholar] [CrossRef]
- Konwerski, S.; Gutowski, J.M.; Książkiewicz-Parulska, Z.; Błoszyk, J. Repeatability of the phoretic relationships between mites of the genus Trichouropoda Berlese (Parasitiformes: Uropodina) and longhorn beetles of the genus Tetropium Kirby (Coleoptera: Cerambycidae) in Białowieża Primeval Forest, Central Europe. Int. J. Acarol. 2017, 43, 612–621. [Google Scholar] [CrossRef]
- Konwerski, S.; Gutowski, J.M.; Błoszyk, J. Analysis of the phoretic relationships between mites of the genus Trichouropoda Berlese (Parasitiformes: Uropodina) and the longhorn beetle Plagionotus detritus (Linnaeus) (Coleoptera: Cerambycidae) based on multiannual observations in Białowieża Primeval Forest, Central Europe. Int. J. Acarol. 2018, 45, 29–40. [Google Scholar] [CrossRef]
- Faliński, J.B. Vegetation Dynamics in Temperate Lowland Primeval Forest: Ecological Studies in Białowieża Forest, 1st ed.; Dr W. Junk Publishers: Dordrecht, The Netherlands, 1986; pp. 1–537. [Google Scholar]
- Bobiec, A.; Van Der Burgt, H.; Meijer, K.; Zuyderduyn, C.; Haga, J.; Vlaanderen, B. Rich deciduous forests in Białowieża as a dynamic mosaic of developmental phases: Premises for nature conservation and restoration management. For. Ecol. Manag. 2000, 130, 159–175. [Google Scholar] [CrossRef]
- Kujawa, A.; Orczewska, A.; Falkowski, M.; Blicharska, M.; Bohdan, A.; Buchholz, L.; Chylarecki, P.; Gutowski, J.M.; Latałowa, M.; Mysłajek, R.W.; et al. The Białowieża Forest—A UNESCO Natural Heritage Site—Protection priorities. For. Res. Pap. 2016, 77, 302–323. [Google Scholar] [CrossRef] [Green Version]
- Jaroszewicz, B.; Cholewińska, O.; Gutowski, J.M.; Samojlik, T.; Zimny, M.; Latałowa, M. Białowieża Forest —A relic of the high naturalness of European forests. Forests 2019, 10, 849. [Google Scholar] [CrossRef] [Green Version]
- Eckelt, A.; Müller, J.; Bense, U.; Brustel, H.; Bußler, H.; Chittaro, Y.; Cizek, L.; Frei, A.; Holzer, E.; Kadej, M.; et al. “Primeval forest relict beetles” of Central Europe: A set of 168 umbrella species for the protection of primeval forest remnants. J. Insect Conserv. 2017, 22, 15–28. [Google Scholar] [CrossRef]
- Gutowski, J.M.; Królik, R.; Ługowoj, J.; Sućko, K.; Sweeney, J. New data on the occurrence of buprestid beetles (Coleoptera: Buprestidae) in the Białowieża Primeval Forest. For. Res. Pap. 2019, 80, 167–176. [Google Scholar] [CrossRef] [Green Version]
- Gutowski, J.M.; Kurzawa, J. New data on the distribution, biology and morphology of Asemum tenuicorne Kraatz, 1879 (Coleoptera: Cerambycidae), with new records from Poland. Èntomol. Fenn. 2019, 30, 57–71. [Google Scholar] [CrossRef] [Green Version]
- Plewa, R.; Jaworski, T.; Tarwacki, G.; Sućko, K.; Konwerski, S.; Królik, R.; Lasoń, A.; Melke, A.; Przewoźny, M.; Ruta, R.; et al. Beetles (Coleoptera) new for the fauna of the Białowieża Forest including a species new for Poland. Èntomol. Fenn. 2019, 30, 114–125. [Google Scholar] [CrossRef] [Green Version]
- Gutowski, J.M.; Sućko, K.; Borowski, J.; Kubisz, D.; Mazur, M.A.; Melke, A.; Mokrzycki, T.; Plewa, R.; Żmihorski, M. Post-fire beetle succession in a biodiversity hotspot: Białowieża Primeval Forest. For. Ecol. Manag. 2020, 461, 117893. [Google Scholar] [CrossRef]
- Plewa, R.; Jaworski, T.; Tarwacki, G.; Sućko, K.; Konwerski, S.; Królik, R.; Lasoń, A.; Melke, A.; Przewoźny, M.; Ruta, R.; et al. New records of beetle species (Coleoptera) from the Polish part of Białowieża Forest with special emphasis on the genus Episernus, C.G. Thomson, 1863 (Ptinidae) in Central Europe. Pol. J. Èntomol. 2020, 1, 26–42. [Google Scholar] [CrossRef]
- Lachat, T.; Müller, J. Importance of Primary Forests for the Conservation of Saproxylic Insects. In Parasitic Crustacea; Springer Science and Business Media LLC.: Berlin/Heidelberg, Germany, 2018; pp. 581–605. [Google Scholar]
- Löbl, I.; Smetana, A. Catalogue of Palaearctic Coleoptera, Vol. 6—Chrysomeloidea; Apollo Books: Stenstrup, Denmark, 2010; pp. 1–924. [Google Scholar]
- Plewa, R.; Sikora, K.; Gutowski, J.M.; Jaworski, T.; Tarwacki, G.; Tkaczyk, M.; Rossa, R.; Hilszczański, J.; Magoga, G.; Kajtoch, Ł. Morphology, genetics and Wolbachia endosymbionts support distinctiveness of Monochamus sartor sartor and M. s. urussovii (Coleoptera: Cerambycidae). Arthropod Syst. Phylo. 2018, 76, 123–135. [Google Scholar]
- Błoszyk, J. Uropodina. In Fauna of Poland. Characteristics and Checklist of Species, 1st ed.; Bogdanowicz, W., Chudzicka, E., Pilipiuk, I., Skibińska, E., Eds.; Muzeum i Instytut Zoologii PAN: Warszawa, Poland, 2008; Volume 3, pp. 61–64, 76–78. [Google Scholar]
- Lawrence, J.F.; Ślipiński, A.; Seago, A.E.; Thayer, M.K.; Newton, A.F.; Marvaldi, A.E. Phylogeny of the Coleoptera Based on Morphological Characters of Adults and Larvae. In Annales zoologici; Museum and Institute of Zoology, Polish Academy of Sciences: Warszawa, Poland, 2011; Volume 61, pp. 1–217. [Google Scholar] [CrossRef]
- Hirschmann, W.; Wiśniewski, J. Gangsystematik der Parasitiformes Teil 507. Weltweite Revision der Ganggatung Trichouropoda Berlese 1916—Geschichte, Bestimmungstabellen, Artenverzeichnisse (Trichouropodini, Uropodinae). Acarologia 1989, 36, 1–65. [Google Scholar]
- Karg, W. Acari (Acarina), Milben Unterordnung Parasitiformes (Anactinochaeta). Uropodina Kramer, Schildkrötenmilben; VEB Gustav Fischer Verlag: Jena, Germany, 1989; Volume 67, pp. 1–203. [Google Scholar]
- Kinn, D.; Linit, M. A Key to Phoretic Mites Commonly Found on Long-Horned Beetles Emerging from Southern Pines; Southern Forest Experiment Station: Asheville, NC, USA, 1989; Volume 357, pp. 1–8. [Google Scholar] [CrossRef]
- Mašán, P. Mites of the kohort Uropodina (Acarina, Mesostigmata) in Slovakia. Annot. Zool. Bot. 2001, 223, 1–320. [Google Scholar]
- StatSoft Inc. Statistica (Data Analysis Software System), version 10; StatSoft Polska Sp. z o.o.: Kraków, Poland, 2011. [Google Scholar]
- Judd, W.W. Nymphal Mites of the Genus Uropoda Attached to a Cerambycid Beetle. Ann. Èntomol. Soc. Am. 1950, 43, 608–609. [Google Scholar] [CrossRef]
- Knee, W.; Hartzenberg, T.; Forbes, M.R.; Beaulieu, F. The natural history of mites (Acari: Mesostigmata) associated with the white-spotted sawyer beetle (Coleoptera: Cerambycidae): Diversity, phenology, host attachment, and sex bias. Can. Èntomol. 2012, 144, 711–719. [Google Scholar] [CrossRef] [Green Version]
- Bajerlein, D.; Adamski, Z.; Kacalak, W.; Tandecka, K.; Wiesner, M.; Jurga, S. To attach or not to attach? The effect of carrier surface morphology and topography on attachment of phoretic deutonymphs of Uropoda orbicularis (Acari). Naturwissenschaften 2016, 103, 61. [Google Scholar] [CrossRef] [Green Version]
- Hunter, P.E. Mites associated with New World passalid beetles (Coleoptera: Passalidae). Acta Zool. Mex. 1993, 58, 1–37. [Google Scholar] [CrossRef]
- Barrios-Torres, P.L.; Villegas-Guzmán, G.A. Ácaros asociados a dos species de Odontotaenius (Coleopetra: Passalidae) en México. Rev. Biol. Trop. 2015, 63, 659–671. [Google Scholar] [CrossRef]
- Hawes, C. Hitching a lift: Stag beetles as carriers of mites. White Admiral 2010, 77, 16–17. [Google Scholar]
- Mazza, G.; Cini, A.; Cervo, R.; Longo, S. Just phoresy? Reduced lifespan in red palm weevils Rhynchophorus ferrugineus (Coleoptera: Curculionidae) infested by the mite Centrouropoda almerodai (Uroactiniinae: Uropodina). Ital. J. Zool. 2011, 78, 101–105. [Google Scholar] [CrossRef] [Green Version]
- Bloszyk, J.; Klimczak, J.; Lesniewska, M. Phoretic relationships between Uropodina (Acari: Mesostigmata) and centipedes (Chilopoda) as an example of evolutionary adaptation of mites to temporary microhabitats. Eur. J. Èntomol. 2006, 103, 699–707. [Google Scholar] [CrossRef] [Green Version]
- Napierała, A.; Książkiewicz, Z.; Lesniewska, M.; Gwiazdowicz, D.J.; Mądra-Bielewicz, A.; Błoszyk, J. Phoretic relationships between uropodid mites (Acari: Mesostigmata) and centipedes (Chilopoda) in urban agglomeration areas. Int. J. Acarol. 2015, 41, 250–258. [Google Scholar] [CrossRef]
- Leśniewska, M.; (Adam Mickiewicz University, Poznań, Poland). Personal communication, 2020.
- Moser, J.C. Phoretic carrying capacity of flying southern pine beetles (Coleoptera: Scolytidae). Can. Èntomol. 1976, 108, 807–808. [Google Scholar] [CrossRef] [Green Version]
- Moser, J.C. Surveying mites (Acarina) phoretic on the southern pine beetle (Coleoptera: Scolytidae) with sticky traps. Can. Èntomol. 1976, 108, 809–813. [Google Scholar] [CrossRef] [Green Version]
- Knee, W.; Forbes, M.R.; Beaulieu, F. Diversity and Host Use of Mites (Acari: Mesostigmata, Oribatida) Phoretic on Bark Beetles (Coleoptera: Scolytinae): Global Generalists, Local Specialists? Ann. Èntomol. Soc. Am. 2013, 106, 339–350. [Google Scholar] [CrossRef]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Beetle Species | L1 | L2 | L3 | H | Sc | Es | Eu | W | Pn | Ps | Me | Mt | St | T |
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| M. s. urussovii | 0 | 0 | 0 | 0 | 0 | 141 | 0 | 0 | 930 | 0 | 13 | 0 | 4 | 0 |
| P. detritus | 37 | 688 | 1645 | 41 | 9 | 745 | 839 | 64 | 261 | 266 | 681 | 251 | 5989 | 7545 |
| Te. Castaneum | 214 | 265 | 231 | 5 | 0 | 21 | 0 | 0 | 40 | 13 | 12 | 16 | 21 | 1 |
| Te. Fuscum | 2262 | 1270 | 591 | 21 | 3 | 49 | 0 | 0 | 213 | 32 | 21 | 20 | 33 | 0 |
| Beetle Species | Co1 | Tr1 | Fe1 | Ti1 | Ta1 | Co2 | Tr2 | Fe2 | Ti2 | Ta2 | Co3 | Tr3 | Fe3 | Ti3 | Ta3 |
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Plagionotus detritus | 7 | 0 | 30 | 0 | 0 | 25 | 0 | 661 | 2 | 0 | 791 | 5 | 843 | 6 | 0 |
| Tetropium castaneum | 8 | 0 | 204 | 2 | 0 | 5 | 1 | 251 | 8 | 0 | 3 | 1 | 222 | 4 | 1 |
| Tetropium fuscum | 50 | 0 | 2209 | 3 | 0 | 4 | 0 | 1265 | 1 | 0 | 10 | 0 | 580 | 1 | 0 |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Konwerski, S.; Gutowski, J.M.; Błoszyk, J. Patterns of Distribution of Phoretic Deutonymphs of Uropodina on Longhorn Beetles in Białowieża Primeval Forest, Central Europe. Diversity 2020, 12, 239. https://doi.org/10.3390/d12060239
Konwerski S, Gutowski JM, Błoszyk J. Patterns of Distribution of Phoretic Deutonymphs of Uropodina on Longhorn Beetles in Białowieża Primeval Forest, Central Europe. Diversity. 2020; 12(6):239. https://doi.org/10.3390/d12060239
Chicago/Turabian StyleKonwerski, Szymon, Jerzy M. Gutowski, and Jerzy Błoszyk. 2020. "Patterns of Distribution of Phoretic Deutonymphs of Uropodina on Longhorn Beetles in Białowieża Primeval Forest, Central Europe" Diversity 12, no. 6: 239. https://doi.org/10.3390/d12060239