
Integrative Descriptions of Two New Mesobiotus Species (Tardigrada, Eutardigrada, Macrobiotidae) from Vietnam †


Abstract
:1. Introduction
2. Material and Methods
2.1. Sample Processing
2.2. Microscopy and Imaging
2.3. Morphometrics and Morphological Nomenclature
2.4. DNA Sequencing
2.5. Phylogenetic Analysis and Genetic Comparisons
3. Results
3.1. Taxonomic Account of the New Species
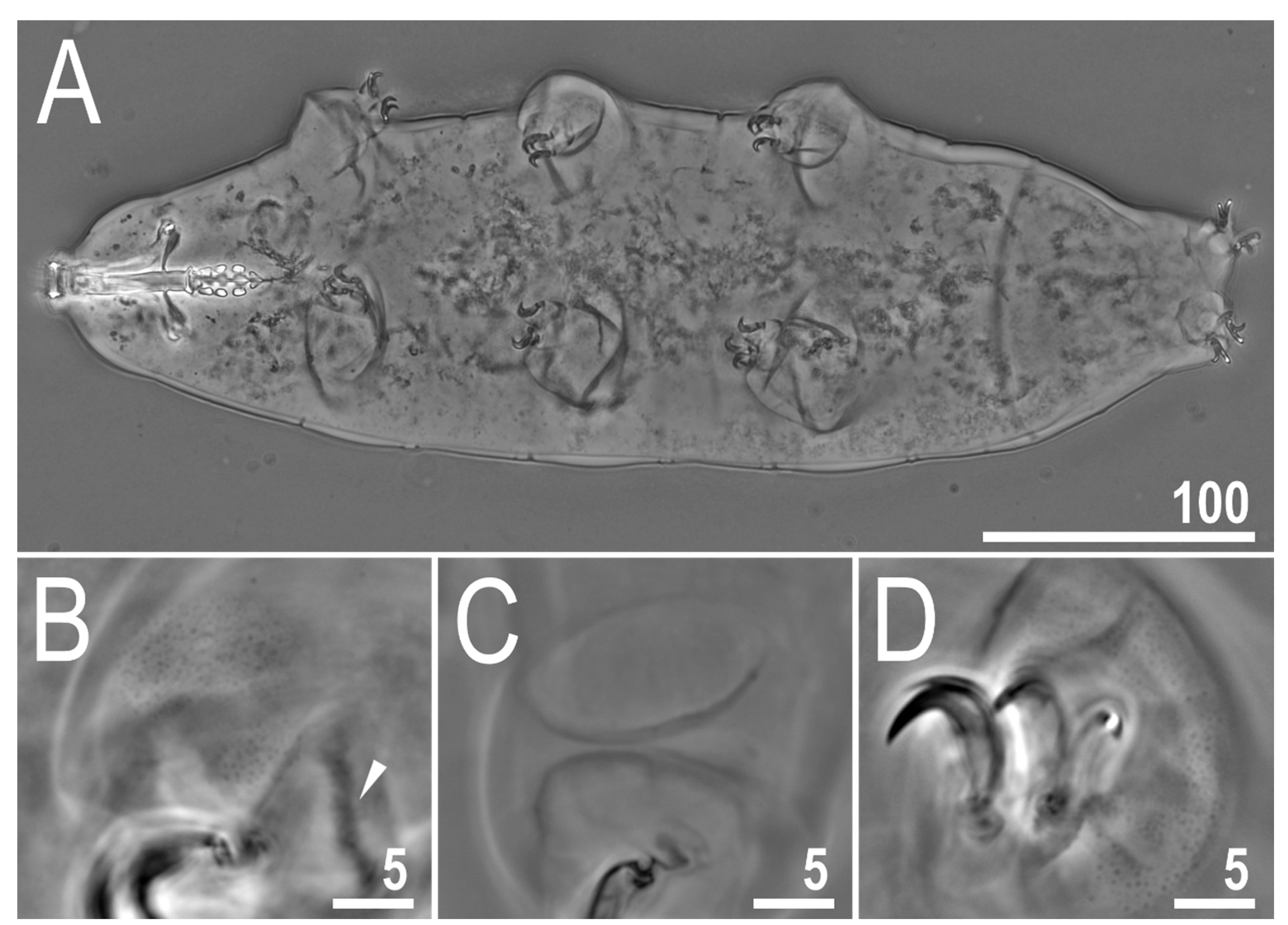
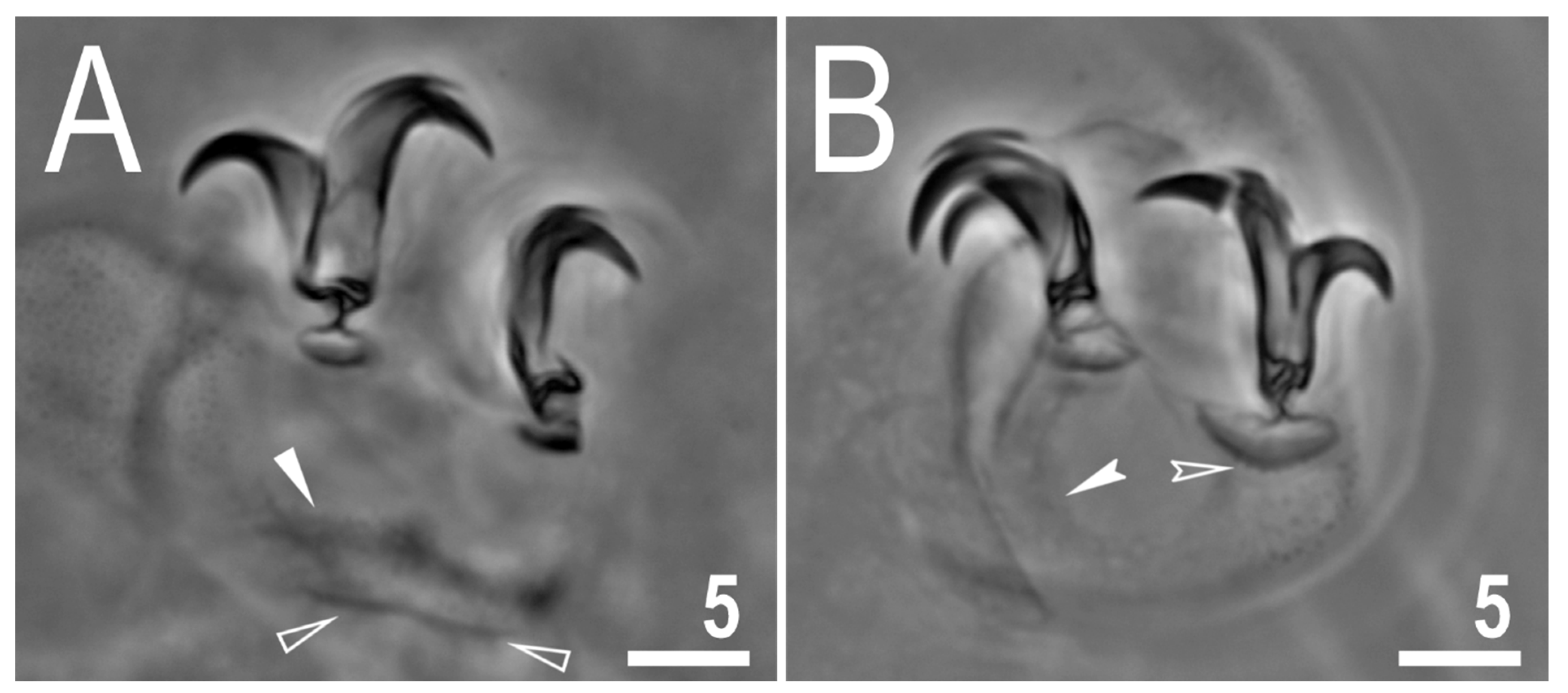
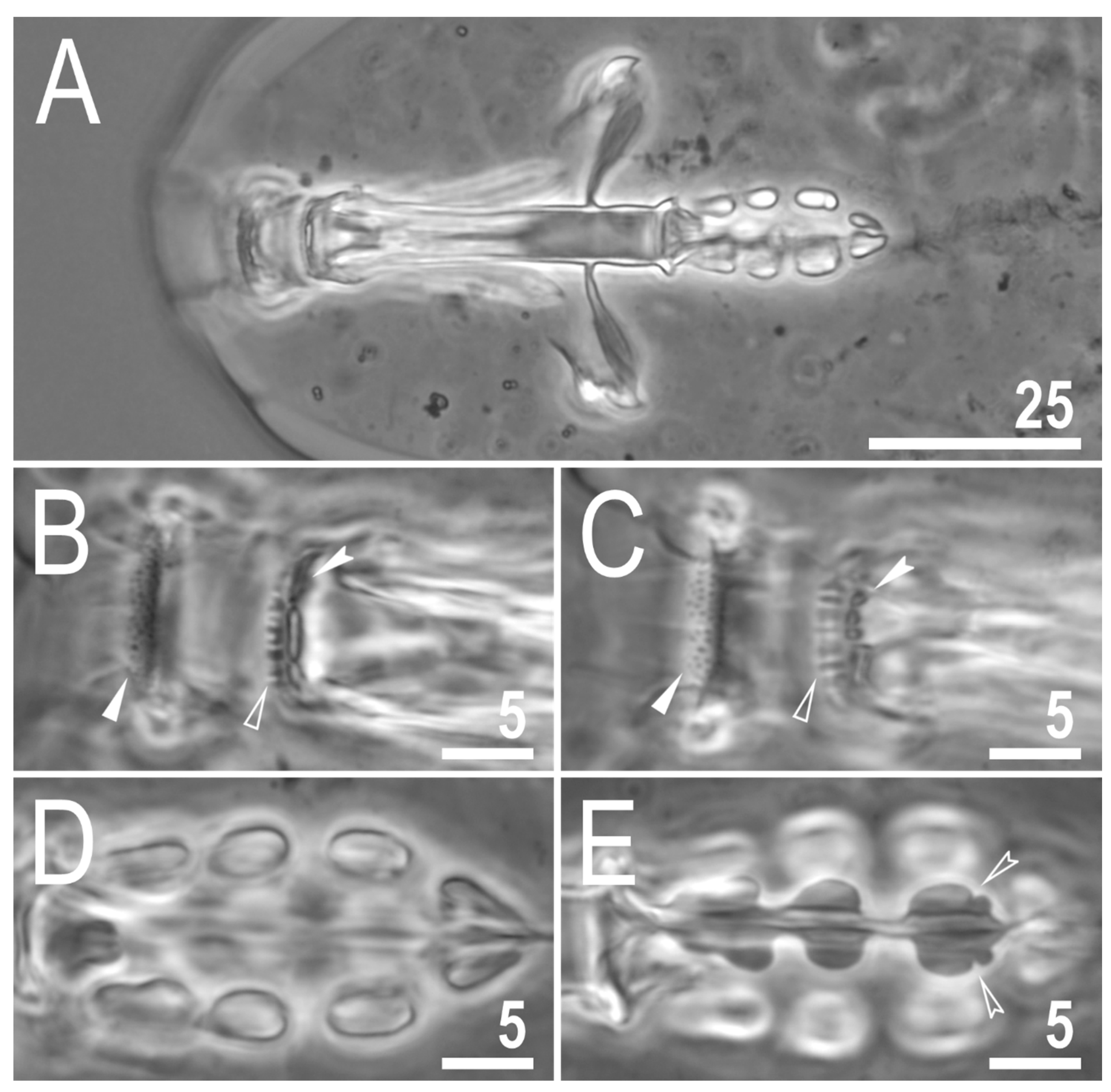
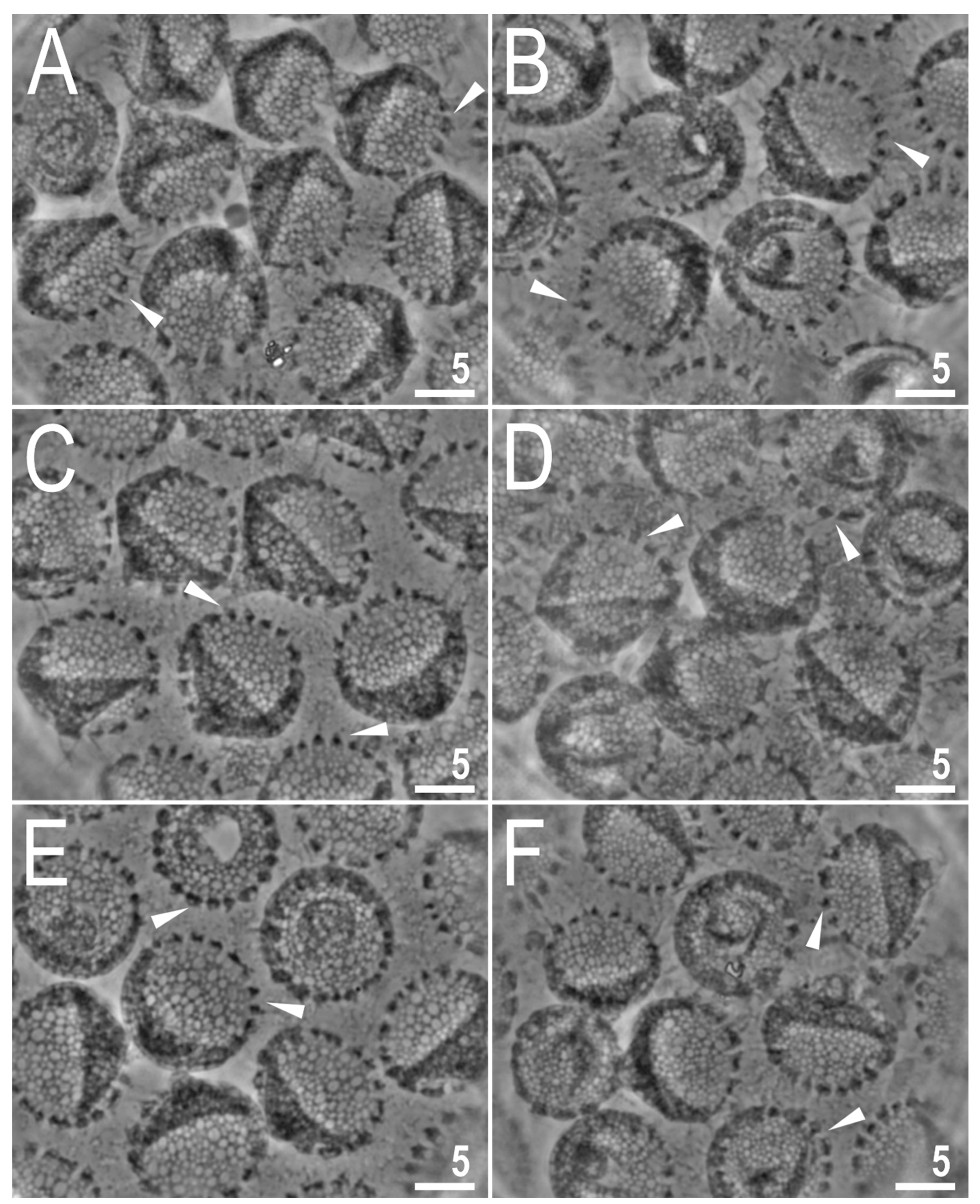
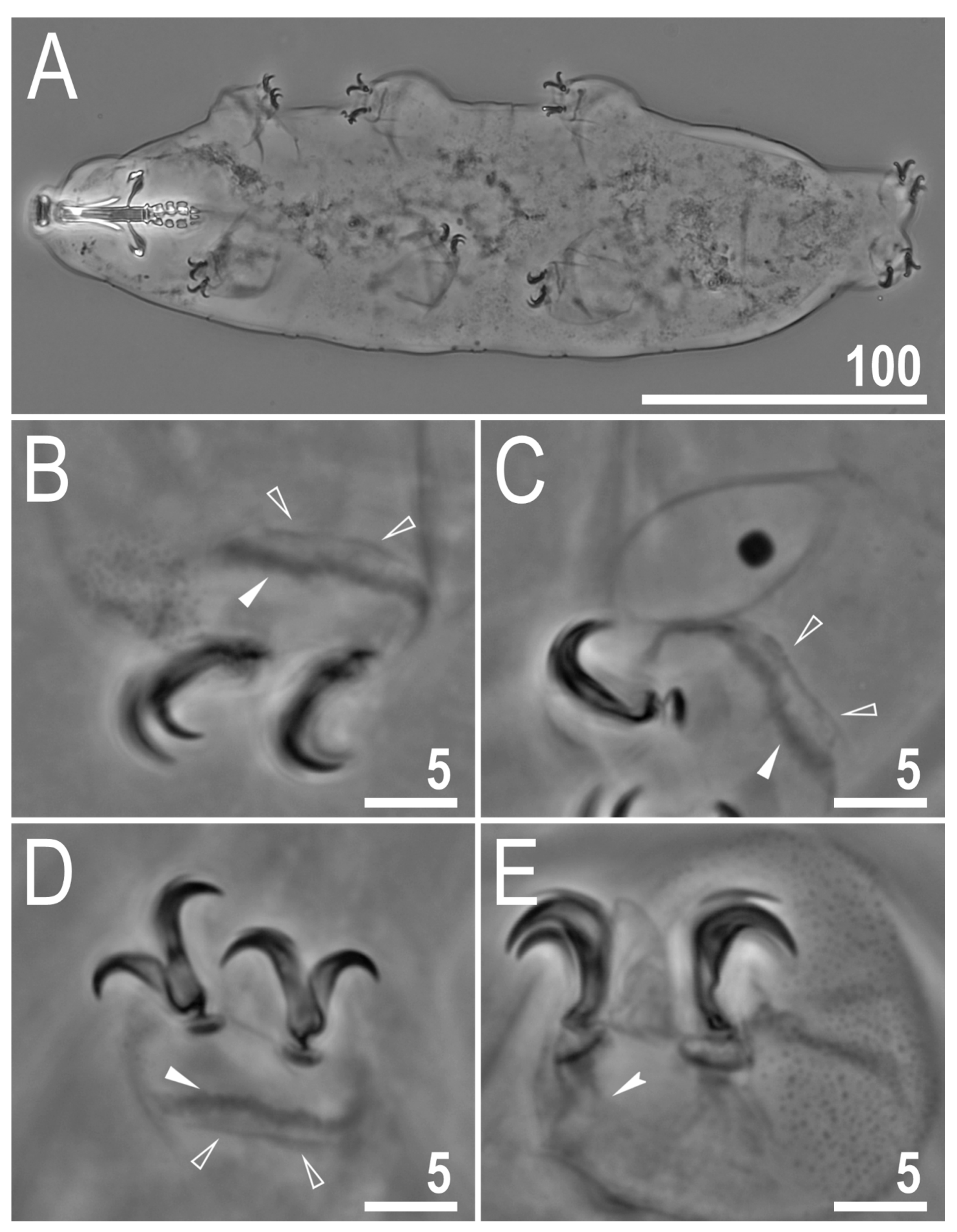
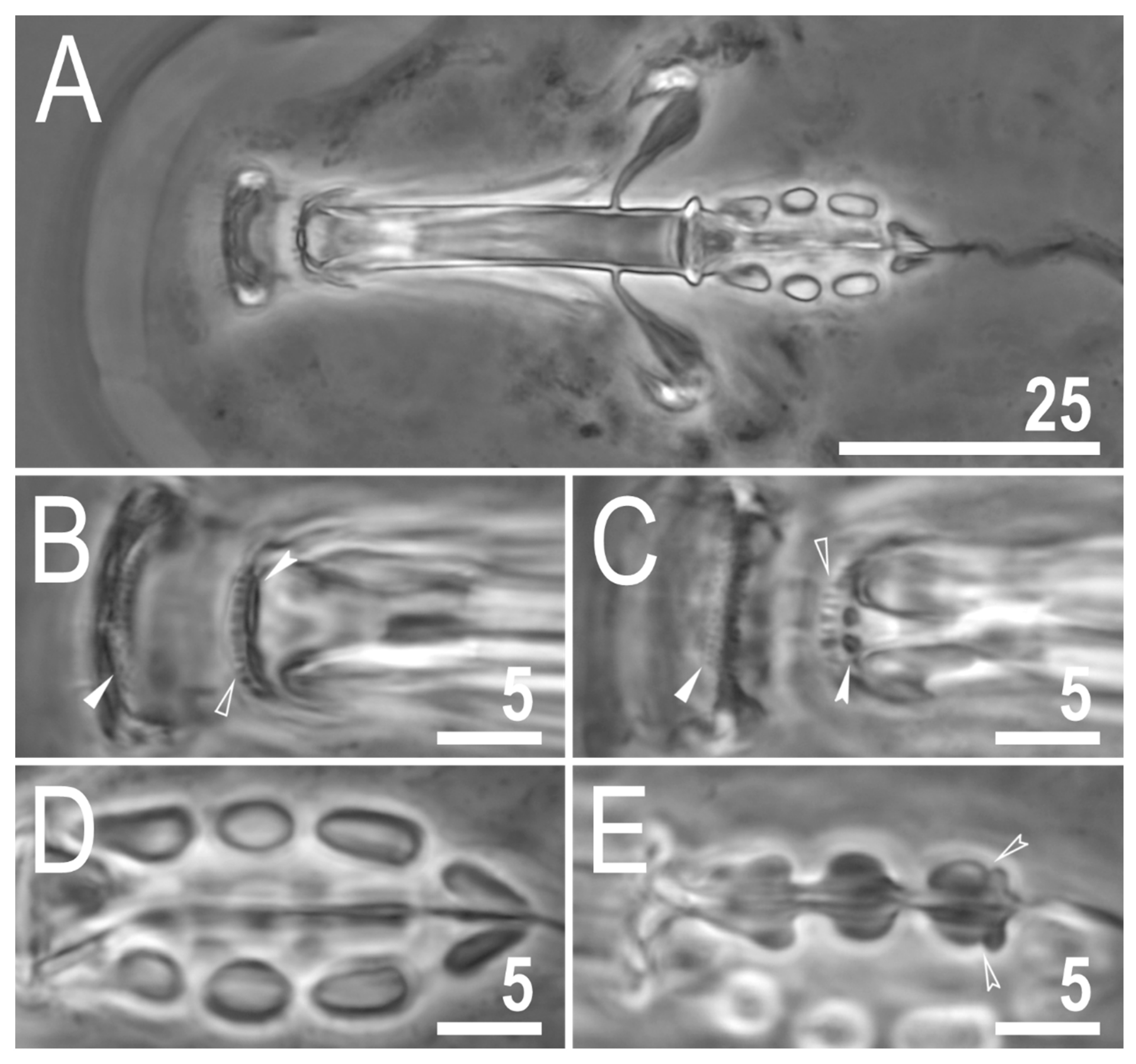
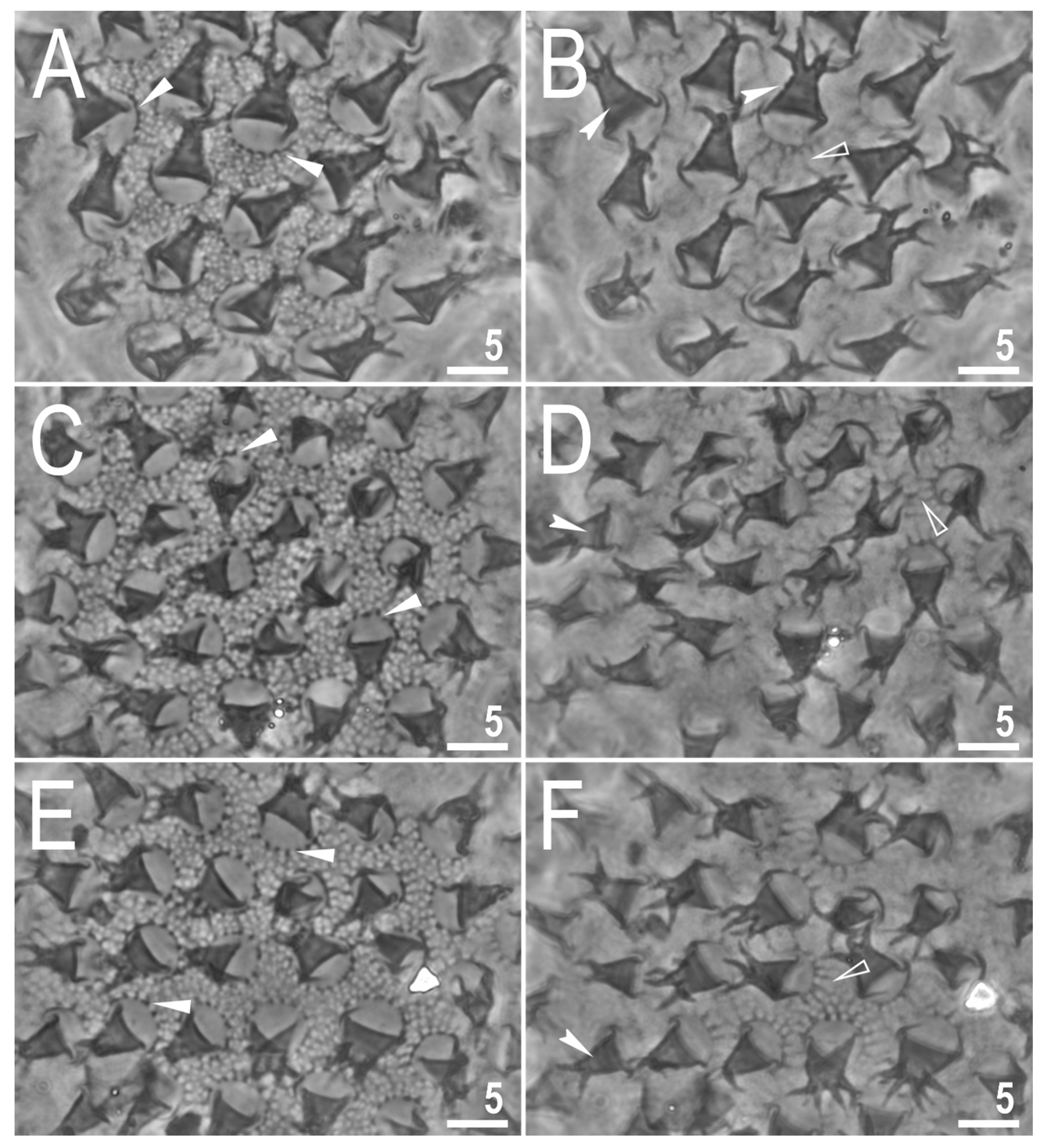
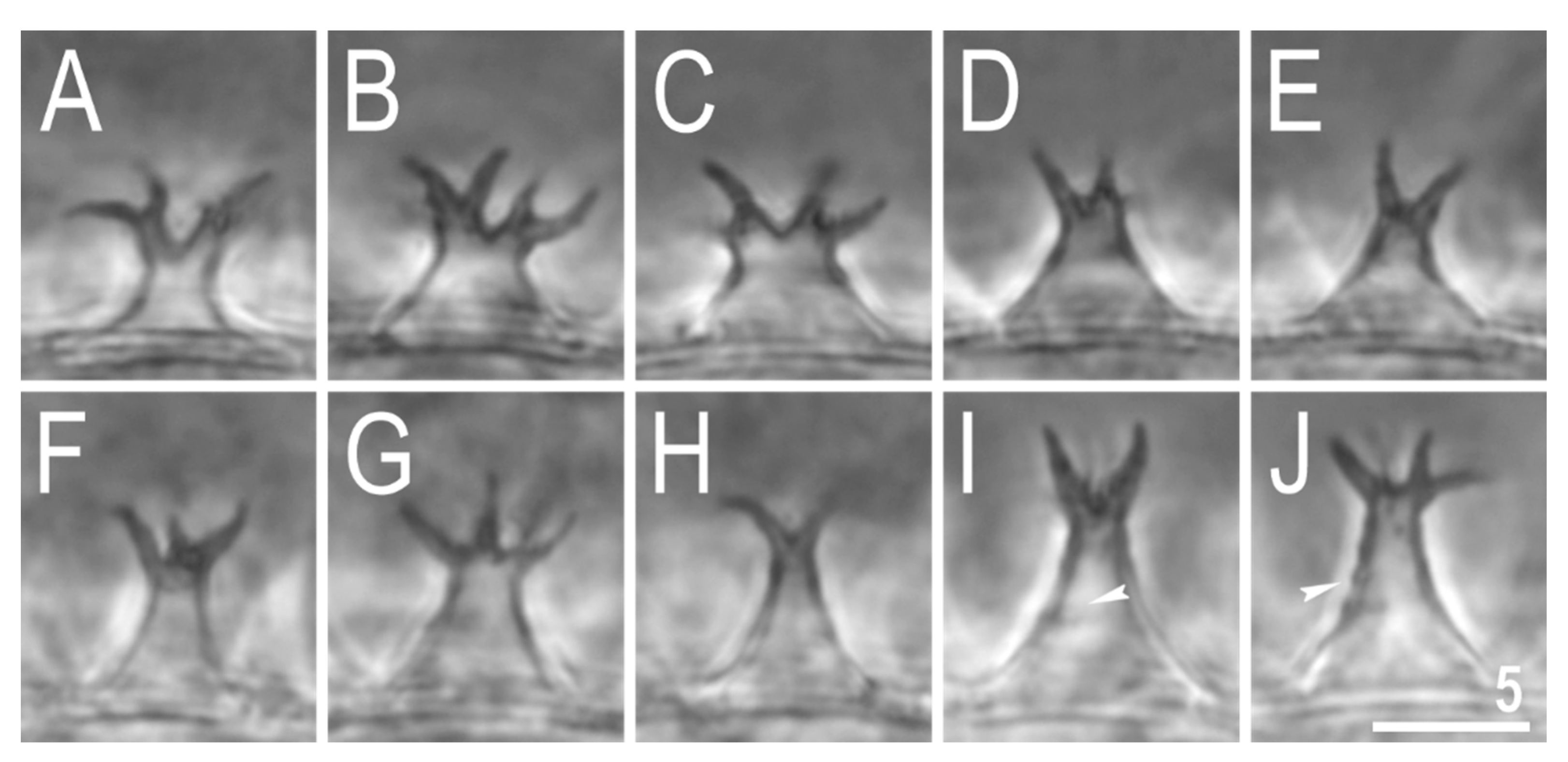
3.2. Description of the New Species
3.2.1. Material Examined
3.2.2. Type Locality
3.2.3. Etymology
3.2.4. Type Depositories
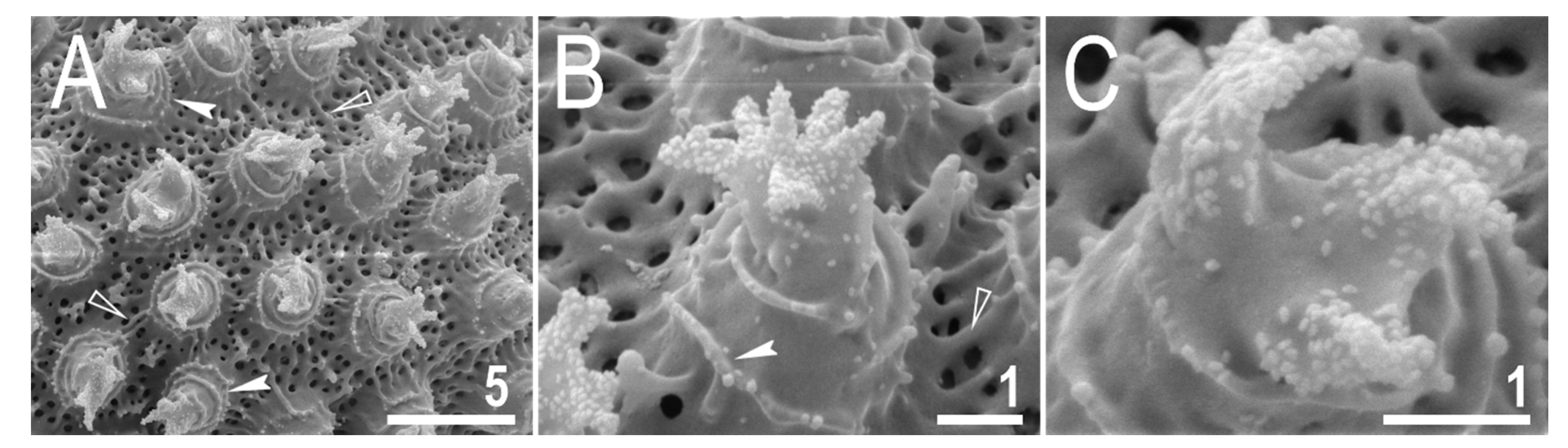
3.2.5. Animals
3.2.6. Eggs
3.2.7. Reproduction
3.2.8. DNA sequences
3.3. Description of the New Species
3.3.1. Material Examined
3.3.2. Type Locality
3.3.3. Etymology
3.3.4. Type Depositories
3.3.5. Animals
3.3.6. Eggs
3.3.7. Reproduction
3.3.8. DNA Sequences
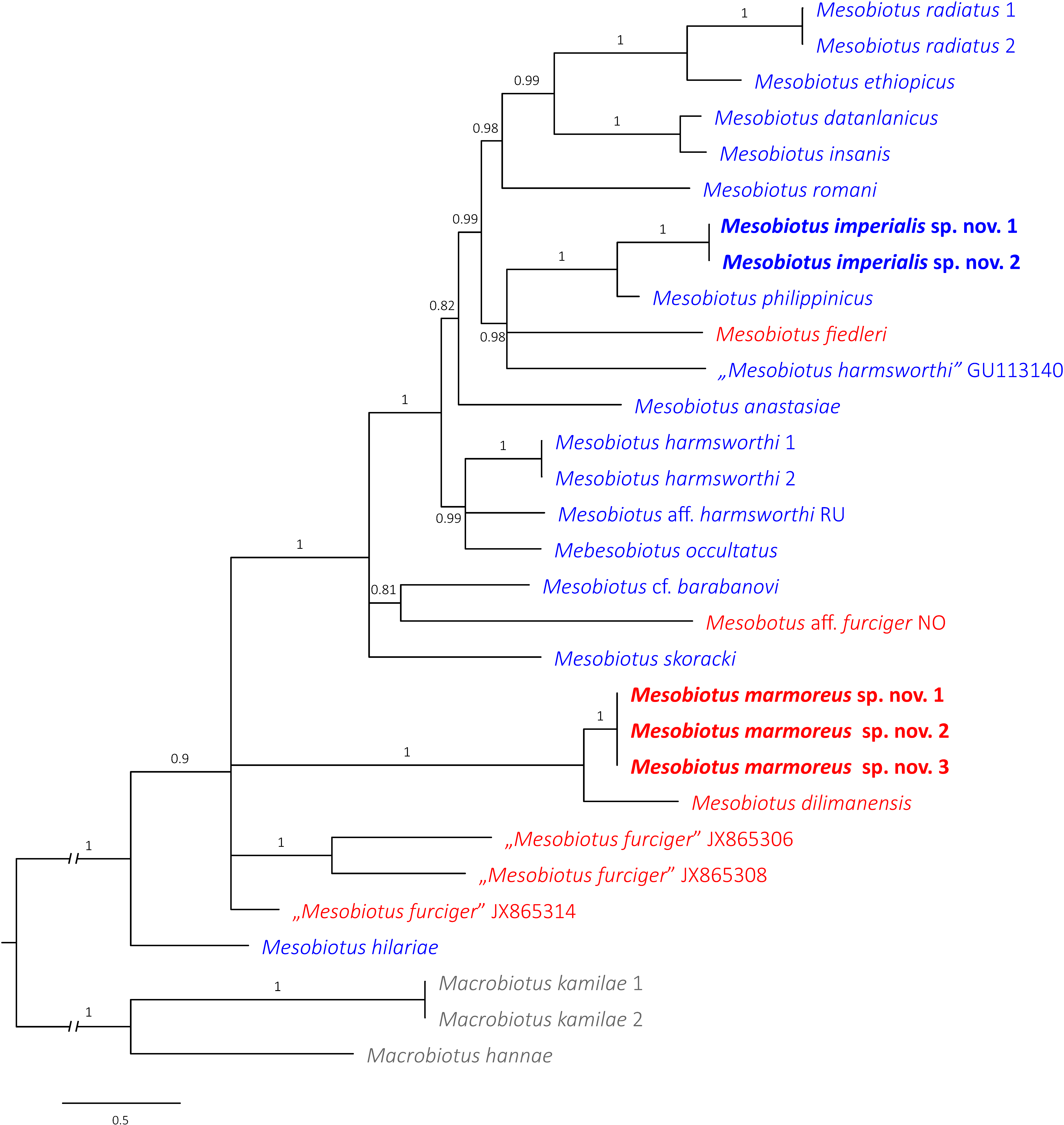
3.4. Phylogenetic Position of the New Taxa
4. Discussion
4.1. Differential Diagnosis of Mesobiotus imperialis sp. nov.
4.2. Differential Diagnosis of Mesobiotus marmoreus sp. nov.
4.3. Conclusions
Supplementary Materials
Funding
Institutional Review Board Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Nelson, D.R.; Guidetti, R.; Rebecchi, L. Phylum Tardigrada. In Ecology and General Biology, 4th ed.; Thorp, J., Rogers, D.C., Eds.; Thorp and Covich’s Freshwater Invertebrates; Academic Press Inc.: Cambridge, MA, USA, 2015; Volume 1, pp. 347–380. [Google Scholar] [CrossRef]
- Guidetti, R.G.; Bertolani, R.B. Tardigrade taxonomy: An updated check list of the taxa and a list of characters for their identification. Zootaxa 2005, 845, 1–46. [Google Scholar] [CrossRef]
- Degma, P.; Guidetti, R. Notes to the current checklist of Tardigrada. Zootaxa 2007, 1579, 41–53. [Google Scholar] [CrossRef]
- Degma, P.; Bertolani, R.; Guidetti, R. Actual Checklist of Tardigrada Species. Available online: https://doi.org/10.25431/11380_1178608 (accessed on 26 April 2021).
- Vecchi, M.; Cesari, M.; Bertolani, R.; Jönsson, K.I.; Rebecchi, L.; Guidetti, R. Integrative systematic studies on tardigrades from Antarctica identify new genera and new species within Macrobiotoidea and Echiniscoidea. Invertebr. Syst. 2016, 30, 303–322. [Google Scholar] [CrossRef]
- Stec, D.; Vecchi, M.; Calhim, S.; Michalczyk, Ł. New multilocus phylogeny reorganises the family Macrobiotidae (Eutardigrada) and unveils complex morphological evolution of the Macrobiotus hufelandi group. Mol. Phylogenetics Evol. 2020, 160, 106987. [Google Scholar] [CrossRef] [PubMed]
- Kaczmarek, Ł.; Bartylak, T.; Stec, D.; Kulpa, A.; Kepel, M.; Kepel, A.; Roszkowska, M. Revisiting the genus Mesobiotus Vecchi et al. 2016 (Eutardigrada, Macrobiotidae)—remarks, updated dichotomous key and an integrative description of new species from Madagascar. Zoöl. Anz. A J. Comp. Zoöl. 2020, 287, 121–146. [Google Scholar] [CrossRef]
- Tumanov, D.V. Integrative description of Mesobiotus anastasiae sp. nov. (Eutardigrada, Macrobiotoidea) and first record of Lobohalacarus (Chelicerata, Trombidiformes) from the Republic of South Africa. Eur. J. Taxon. 2020, 726, 102–131. [Google Scholar] [CrossRef]
- Kaczmarek, Ł.; Zawierucha, K.; Buda, J.; Stec, D.; Gawlak, M.; Michalczyk, Ł.; Roszkowska, M. An integrative redescription of the nominal taxon for the Mesobiotus harmsworthi group (Tardigrada: Macrobiotidae) leads to descriptions of two new Mesobiotus species from Arctic. PLoS ONE 2018, 13, e0204756. [Google Scholar] [CrossRef]
- Węglarska, B. Die Tardigraden Vietnams. Acta Soc. Zool. Bohemoslov. 1962, 26, 300–307. [Google Scholar]
- Iharos, G. Einige Angaben zur Tardigradenfauna Vietnams. Opusc. Zool. Bp. 1969, 9, 273–277. [Google Scholar]
- Pilato, G.; Binda, M.G. Isohypsibius barbarae, a new species of eutardigrade from Vietnam. Bolletino Delle Sedute Dell’accademia Gioenia Di Sci. Nat. Catania 2002, 35, 637–642. [Google Scholar]
- Beasley, C.W.; Kaczmarek, Ł.; Michalczyk, Ł. Redescription of Doryphoribius vietnamensis (Iharos, 1969) comb. nov. on the basis of the holotype and additional material from China. Acta Zool. Acad. Sci. Hung. 2006, 52, 367–372. [Google Scholar] [CrossRef]
- Tchesunov, A.V. Marine tardigrade Halechiniscus jejuensis Chang et Rho, 2002 (Arthrotardigrada: Halechiniscidae) found in Vietnam. Invertzool 2011, 8, 79–85. [Google Scholar] [CrossRef]
- Stec, D. Mesobiotus datanlanicus sp. nov., a new tardigrade species (Macrobiotidae: Mesobiotus harmsworthi group) from Lâm Đồng Province in Vietnam. Zootaxa 2019, 4679, 164–180. [Google Scholar] [CrossRef] [PubMed]
- Gąsiorek, P.; Vončina, K.; Nelson, D.R.; Michalczyk, Ł. The importance of being integrative: A remarkable case of synonymy in the genus Viridiscus (Heterotardigrada: Echiniscidae). Zool. Lett. 2021, in press. [Google Scholar] [CrossRef]
- Murray, J. XXV.—Arctic Tardigrada, collected by Wm. S. Bruce. Trans. R. Soc. Edinb. 1907, 45, 669–681. [Google Scholar] [CrossRef] [Green Version]
- Stec, D.; Smolak, R.; Kaczmarek, Ł.; Michalczyk, Ł. An integrative description of Macrobiotus paulinae sp. nov. (Tardigrada: Eutardigrada: Macrobiotidae: Hufelandi group) from Kenya. Zootaxa 2015, 4052, 501–526. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Morek, W.; Stec, D.; Gąsiorek, P.; Schill, R.O.; Kaczmarek, Ł.; Michalczyk, Ł. An experimental test of eutardigrade preparation methods for light microscopy. Zoöl. J. Linn. Soc. 2016, 178, 785–793. [Google Scholar] [CrossRef] [Green Version]
- Coughlan, K.; Stec, D. Two new species of the Macrobiotus hufelandi complex (Tardigrada: Eutardigrada: Macrobiotidae) from Australia and India, with notes on their phylogenetic position. Eur. J. Taxon. 2019, 573, 1–38. [Google Scholar] [CrossRef]
- Coughlan, K.; Michalczyk, Ł.; Stec, D. Macrobiotus caelestis sp. nov., a New Tardigrade Species (Macrobiotidae: Hufelandi Group) from the Tien Shan Mountains (Kyrgyzstan). Ann. Zoöl. 2019, 69, 499. [Google Scholar] [CrossRef]
- Stec, D.; Gąsiorek, P.; Morek, W.; Kosztyła, P.; Zawierucha, K.; Michno, K.; Kaczmarek, Ł.; Prokop, Z.M.; Michalczyk, Ł. Estimating optimal sample size for tardigrade morphometry. Zoöl. J. Linn. Soc. 2016, 178, 776–784. [Google Scholar] [CrossRef] [Green Version]
- Pilato, G.; Binda, M.G. Definition of families, subfamilies, genera and subgenera of the Eutardigrada, and keys to their identification. Zootaxa 2010, 2404, 1–54. [Google Scholar] [CrossRef] [Green Version]
- Michalczyk, Ł.; Kaczmarek, Ł. A description of the new tardigrade Macrobiotus reinhardti (Eutardigrada: Macrobiotidae, harmsworthi group) with some remarks on the oral cavity armature within the genus Macrobiotus Schultze. Zootaxa 2003, 331, 1–24. [Google Scholar] [CrossRef] [Green Version]
- Kaczmarek, Ł.; Michalczyk, Ł. The Macrobiotus hufelandi group (Tardigrada) revisited. Zootaxa 2017, 4363, 101–123. [Google Scholar] [CrossRef]
- Kaczmarek, Ł.; Cytan, J.; Zawierucha, K.; Diduszko, D.; Michalczyk, Ł. Tardigrades from Peru (South America), with descriptions of three new species of Parachela. Zootaxa 2014, 3790, 357–379. [Google Scholar] [CrossRef] [Green Version]
- Kiosya, Y.; Pogwizd, J.; Matsko, Y.; Vecchi, M.; Stec, D. Phylogenetic position of two Macrobiotus species with a revisional note on Macrobiotus sottilei Pilato, Kiosya, Lisi & Sabella, 2012 (Tardigrada: Eutardigrada: Macrobiotidae). Zootaxa 2021, 4933, 113–135. [Google Scholar] [CrossRef]
- Pilato, G. Analisi di nuovi caratteri nello studio degli Eutardigradi. Animalia 1981, 8, 51–57. [Google Scholar]
- Michalczyk, Ł.; Kaczmarek, Ł. The Tardigrada Register: A comprehensive online data repository for tardigrade taxonomy. J. Limnol. 2013, 72, e22. [Google Scholar] [CrossRef]
- Mapalo, M.A.; Stec, D.; Mirano-Bascos, D.; Michalczyk, Ł. Mesobiotus philippinicus sp. nov., the first limnoterrestrial tardigrade from the Philippines. Zootaxa 2016, 4126, 411–426. [Google Scholar] [CrossRef]
- R Core Team. R: A Language and Environment for Statistical Computing; R Foundation for Statistical Computing: Vienna, Austria, 2021; Available online: http://www.R-project.org/ (accessed on 26 April 2021).
- Benjamini, Y.; Hochberg, Y. Controlling the False Discovery Rate: A Practical and Powerful Approach to Multiple Testing. J. R. Stat. Soc. Ser. B Methodol. 1995, 57, 289–300. [Google Scholar] [CrossRef]
- Casquet, J.; Thébaud, C.; Gillespie, R.G. Chelex without boiling, a rapid and easy technique to obtain stable amplifiable DNA from small amounts of ethanol-stored spiders. Mol. Ecol. Resour. 2011, 12, 136–141. [Google Scholar] [CrossRef] [PubMed]
- Stec, D.; Kristensen, R.M.; Michalczyk, Ł. An integrative description of Minibiotus ioculator sp. nov. from the Republic of South Africa with notes on Minibiotus pentannulatus Londoño et al., 2017 (Tardigrada: Macrobiotidae). Zoöl. Anz. A J. Comp. Zoöl. 2020, 286, 117–134. [Google Scholar] [CrossRef]
- Hall, T.A. BioEdit: A user-friendly biological sequence alignment editor and analysis program for Windows 95/98/NT. Nucleic Acids Symp. Ser. 1999, 41, 95–98. [Google Scholar]
- Kumar, S.; Stecher, G.; Tamura, K. MEGA7: Molecular Evolutionary Genetics Analysis Version 7.0 for Bigger Datasets. Mol. Biol. Evol. 2016, 33, 1870–1874. [Google Scholar] [CrossRef] [Green Version]
- Stec, D.; Zawierucha, K.; Michalczyk, Ł. An integrative description of Ramazzottius subanomalus (Biserov, 1985 (Tardigrada) from Poland. Zootaxa 2017, 4300, 403–420. [Google Scholar] [CrossRef]
- Gąsiorek, P.; Stec, D.; Zawierucha, K.; Kristensen, R.M.; Michalczyk, Ł. Revision of Testechiniscus Kristensen, 1987 (Heterotardigrada: Echiniscidae) refutes the polar-temperate distribution of the genus. Zootaxa 2018, 4472, 261–297. [Google Scholar] [CrossRef]
- Mironov, S.V.; Dabert, J.; Dabert, M. A new feather mite species of the genus Proctophyllodes Robin, 1877 (Astigmata: Proctophyllodidae) from the Long-tailed Tit Aegithalos caudatus (Passeriformes: Aegithalidae)—morphological description with DNA barcode data. Zootaxa 2012, 3253, 54–61. [Google Scholar] [CrossRef] [Green Version]
- Stec, D.; Morek, W.; Gąsiorek, P.; Michalczyk, Ł. Unmasking hidden species diversity within the Ramazzottius oberhaeuseri complex, with an integrative redescription of the nominal species for the family Ramazzottiidae (Tardigrada: Eutardigrada: Parachela). Syst. Biodivers. 2018, 16, 357–376. [Google Scholar] [CrossRef]
- Astrin, J.J.; Stüben, P.E. Phylogeny in cryptic weevils: Molecules, morphology and new genera of western Palaearctic Cryptorhynchinae (Coleoptera:Curculionidae). Invertebr. Syst. 2008, 22, 503–522. [Google Scholar] [CrossRef] [Green Version]
- Nowak, B. An integrative description of Macrobiotus hannae sp. nov. (Tardigrada: Eutardigrada: Macrobiotidae: Hufelandi group) from Poland. Turk. J. Zoöl. 2018, 42, 269–286. [Google Scholar] [CrossRef]
- Katoh, K.; Misawa, K.; Kuma, K.; Miyata, T. MAFFT: A novel method for rapid multiple sequence alignment based on fast Fourier transform. Nucleic Acids Res. 2002, 30, 3059–3066. [Google Scholar] [CrossRef] [Green Version]
- Katoh, K.; Toh, H. Recent developments in the MAFFT multiple sequence alignment program. Briefings Bioinform. 2008, 9, 286–298. [Google Scholar] [CrossRef] [Green Version]
- Vaidya, G.; Lohman, D.J.; Meier, R. SequenceMatrix: Concatenation software for the fast assembly of multi-gene datasets with character set and codon information. Cladistics 2011, 27, 171–180. [Google Scholar] [CrossRef]
- Lanfear, R.; Frandsen, P.B.; Wright, A.M.; Senfeld, T.; Calcott, B. PartitionFinder 2: New Methods for Selecting Partitioned Models of Evolution for Molecular and Morphological Phylogenetic Analyses. Mol. Biol. Evol. 2017, 34, 772–773. [Google Scholar] [CrossRef] [Green Version]
- Ronquist, F.; Huelsenbeck, J.P. MrBayes 3: Bayesian phylogenetic inference under mixed models. Bioinformatics 2003, 19, 1572–1574. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rambaut, A.; Drummond, A.J. Tracer v1.6. 2014. Available online: http://beast.bio.ed.ac.uk/Tracer (accessed on 8 September 2021).
- Stec, D.; Kristensen, R.M. An integrative description of Mesobiotus ethiopicus sp. nov. (Tardigrada: Eutardigrada: Parachela: Macrobiotidae: Harmsworthi group) from the northern Afrotropic region. Turk. J. Zoöl. 2017, 41, 800–811. [Google Scholar] [CrossRef]
- Itang, L.A.M.; Stec, D.; Mapalo, M.A.; Mirano-Bascos, D.; Michalczyk, Ł. An integrative description of Mesobiotus dilimanensis, a new tardigrade species from the Philippines (Eutardigrada: Macrobiotidae: Furciger group). Raffles Bull. Zool. 2020, 68, 19–31. [Google Scholar]
- Mapalo, M.; Stec, D.; Mirano-Bascos, D.; Michalczyk, Ł. An integrative description of a limnoterrestrial tardigrade from the Philippines, Mesobiotus insanis, new species (Eutardigrada: Macrobiotidae: Harmsworthi group). Raffles Bull. Zool. 2017, 65, 440–454. [Google Scholar]
- Pilato, G.; Binda, M.G.; Catanzaro, R. Remarks on some tardigrades of the African fauna with the description of three new species of Macrobiotus Schultze 1834. Trop. Zoöl. 1991, 4, 167–178. [Google Scholar] [CrossRef] [Green Version]
- Stec, D.; Roszkowska, M.; Kaczmarek, Ł.; Michalczyk, Ł. An integrative description of a population of Mesobiotus radiatus (Pilato, Binda & Catanzaro, 1991) from Kenya. Turk. J. Zoöl. 2018, 42, 523–540. [Google Scholar] [CrossRef] [Green Version]
- Roszkowska, M.; Stec, D.; Gawlak, M.; Kaczmarek, Ł. An integrative description of a new tardigrade species Mesobiotus romani sp. nov. (Macrobiotidae: Harmsworthi group) from the Ecuadorian Pacific coast. Zootaxa 2018, 4450, 550–564. [Google Scholar] [CrossRef]
- Murray, J. Encystment of Tardigrada. Trans. R. Soc. Edinb. 1908, 45, 837–854. [Google Scholar] [CrossRef] [Green Version]
- Czechowski, P.; Sands, C.J.; Adams, B.J.; D’Haese, C.A.; Gibson, J.A.E.; McInnes, S.J.; Stevens, M.I. Antarctic Tardigrada: A first step in understanding molecular operational taxonomic units (MOTUs) and biogeography of cryptic meiofauna. Invertebr. Syst. 2012, 26, 526–538. [Google Scholar] [CrossRef]
- Kayastha, P.; Roszkowska, M.; Mioduchowska, M.; Gawlak, M.; Kaczmarek, Ł. Integrative Descriptions of Two New Tardigrade Species along with the New Record of Mesobiotus skorackii Kaczmarek et al., 2018 from Canada. Diversity 2021, 13, 394. [Google Scholar] [CrossRef]
- Doyère, P.L.N. Memoire sur les Tardigrades. Ann. Des Sci. Nat. 1840, 14, 269–362. [Google Scholar]
- Richters, F. Tardigrada. In Handbuch der Zoologie; Kükenthal, W., Krumbach, T., Eds.; Walter de Gruyter & Co.: Berlin/Heidelberg, Germany, 1926; Volume 3, pp. 58–61. [Google Scholar]
- Schuster, R.O.; Nelson, D.R.; Grigarick, A.A.; Christenberry, D. Systematic Criteria of the Eutardigrada. Trans. Am. Microsc. Soc. 1980, 99, 284–303. [Google Scholar] [CrossRef]
- Thulin, G. Über die phylogenie und das system der tardigraden. Hereditas 2010, 11, 207–266. [Google Scholar] [CrossRef]
- Marley, N.; McInnes, S.J.; Sands, C. Phylum Tardigrada: A re-evaluation of the Parachela. Zootaxa 2011, 2819, 51–64. [Google Scholar] [CrossRef]
- Pilato, G.; Lisi, O.P.V. Tardigrades of the Seychelles Islands, with the description of three new species. Zootaxa 2009, 2124, 1–20. [Google Scholar] [CrossRef] [Green Version]
- Binda, M.G.; Pilato, G.; Lisi, O.P.V. Remarks on Macrobiotus furciger Murray, 1906 and description of three new species of the furciger group (Eutardigrada, Macrobiotidae). Zootaxa 2005, 1075, 55–68. [Google Scholar] [CrossRef]
- Tumanov, D.V. Macrobiotus kovalevi, a new species of Tardigrada from New Zealand (Eutadigrada, Macrobiotidae). Zootaxa 2004, 406, 1–8. [Google Scholar] [CrossRef]
- Tumanov, D.V. Macrobiotus siamensis sp. n. (Eutardigrada, Macrobiotidae) from Thailand (Asia). Zootaxa 2006, 1202, 53–59. [Google Scholar] [CrossRef]













{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| DNA Marker | Primer Name | Primer Direction | Primer Sequence (5′-3′) | Primer Source |
|---|---|---|---|---|
| 18S rRNA | 18S_Tar_Ff1 | forward | AGGCGAAACCGCGAATGGCTC | [37] |
| 18S_Tar_Rr1 | reverse | GCCGCAGGCTCCACTCCTGG | ||
| 28S rRNA | 28S_Eutar_F | forward | ACCCGCTGAACTTAAGCATAT | [38] [39] |
| 28SR0990 | reverse | CCTTGGTCCGTGTTTCAAGAC | ||
| ITS-2 | ITS2_Eutar_Ff | forward | CGTAACGTGAATTGCAGGAC | [40] |
| ITS2_Eutar_Rr | reverse | TCCTCCGCTTATTGATATGC | ||
| COI | LCO1490-JJ | forward | CHACWAAYCATAAAGATATYGG | [41] |
| HCO2198-JJ | reverse | AWACTTCVGGRTGVCCAAARAATCA |
| Species | 18S rRNA | 28S rRNA | ITS-2 | COI | Source |
|---|---|---|---|---|---|
| M. ethiopicus Stec & Kristensen, 2017 [49] | MF678793 | MF678792 | MN122776 | MF678794 | [15,49] |
| M. datanlanicus Stec, 2019 [15] | MK584659 | MK584658 | MK584657 | MK578905 | [15] |
| M. dilimanensis Itang et al., 2020 [50] | MN257048 | MN257049 | MN257050 | MN257047 | [50] |
| M. philippinicus Mapalo et al., 2016 [30] | KX129793 | KX129794 | KX129795 | KX129796 | [30] |
| M. insanis Mapalo et al., 2017 [51] | MF441488 | MF441489 | MF441490 | MF441491 | [51] |
| M. hilariae Vecchi et al., 2016 [5] | KT226070 | KT226108 | [5] | ||
| M. radiatus (Pilato et al., 1991) [52] | MH197153 | MH197152 | MH197267 | MH195147 | [53] |
| MH197268 | MH195148 | ||||
| M. romani Roszkowska et al., 2018 [54] | MH197158 | MH197151 | MH197150 | MH195149 | [54] |
| M. harmsworthi (Murray, 1907) [17] | MH197146 | MH197264 | MH197154 | MH195150 | [9] |
| MH195151 | |||||
| M. occultatus Kaczmarek et al., 2018 [9] | MH197147 | MH197155 | MH195152 | [9] | |
| M. furciger group species NO | MH197148 | MH197265 | MH197156 | MH195153 | [9] |
| M. harmsworthi group species RU | MH197149 | MH197266 | MH197157 | MH195154 | [9] |
| M. furciger (Murray, 1907) [55] | JX865306 | [56] | |||
| JX865308 | |||||
| JX865314 | |||||
| M. fiedleri Kaczmarek et al., 2020 [7] | MH681585 | MH681693 | MH681724 | MH676056 | [7] |
| “M. harmsworthi” | GU113140 | Li and Xiao (unpublished) | |||
| M. anastasiae Tumanov, 2020 [8] | MT903468 | MT903612 | MT903470 | MT904513 | [8] |
| M. skoracki Kaczmarek et al., 2018 [9] | MW680636 | MW656257 | [57] | ||
| M. imperialissp. nov. | OL257854 | OL257866 | OL311514 | this study | |
| OL257855 | OL257867 | OL311515 | this study | ||
| M. marmoreussp. nov. | OL257856 | OL257868 | OL257861 | OL311516 | this study |
| OL257857 | OL257869 | OL257862 | OL311517 | this study | |
| OL257858 | OL257870 | OL257863 | OL311518 | this study | |
| M. cf. barabanovi | MN310392 | MN310388 | MN310390 | MN313170 | [7] |
| Macrobiotus kamilae Coughlan & Stec, 2019 [20] | MK737070 | MK737064 | MK737067 | MK737920 | [20] |
| MK737921 | |||||
| Macrobiotus hannae Nowak & Stec, 2018 [42] | MH063922 | MH063924 | MH063923 | MH057764 | [42] |
| CHARACTER | N | RANGE | Mean | SD | Holotype | ||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| µm | pt | µm | pt | µm | pt | µm | pt | ||||||
| Body length | 20 | 313 | – | 539 | 895 | – | 1219 | 389 | 1012 | 51 | 75 | 436 | 1053 |
| Buccal tube | |||||||||||||
| Buccal tube length | 20 | 30.5 | – | 44.2 | – | 38.4 | – | 3.4 | – | 41.4 | – | ||
| Stylet support insertion point | 20 | 22.9 | – | 34.1 | 75.0 | – | 77.3 | 29.2 | 76.0 | 2.7 | 0.8 | 31.2 | 75.4 |
| Buccal tube external width | 20 | 5.2 | – | 7.6 | 15.7 | – | 18.1 | 6.5 | 16.8 | 0.6 | 0.6 | 7.0 | 16.9 |
| Buccal tube internal width | 20 | 4.0 | – | 5.7 | 10.8 | – | 13.8 | 4.9 | 12.6 | 0.5 | 0.7 | 5.7 | 13.8 |
| Ventral lamina length | 20 | 19.9 | – | 27.0 | 56.2 | – | 65.2 | 23.2 | 60.4 | 1.8 | 2.2 | 23.6 | 57.0 |
| Placoid lengths | |||||||||||||
| Macroplacoid 1 | 20 | 3.8 | – | 6.0 | 11.7 | – | 14.4 | 5.1 | 13.4 | 0.6 | 0.9 | 5.6 | 13.5 |
| Macroplacoid 2 | 20 | 2.9 | – | 4.5 | 7.7 | – | 11.3 | 3.7 | 9.7 | 0.5 | 0.9 | 4.2 | 10.1 |
| Macroplacoid 3 | 20 | 3.1 | – | 5.8 | 10.2 | – | 13.1 | 4.5 | 11.6 | 0.6 | 0.8 | 4.7 | 11.4 |
| Microplacoid | 20 | 3.2 | – | 5.2 | 8.3 | – | 12.7 | 4.1 | 10.7 | 0.5 | 1.1 | 4.2 | 10.1 |
| Macroplacoid row | 20 | 11.9 | – | 19.4 | 38.6 | – | 43.9 | 16.0 | 41.5 | 1.8 | 1.6 | 17.9 | 43.2 |
| Placoid row | 20 | 16.0 | – | 25.7 | 52.0 | – | 58.5 | 21.3 | 55.4 | 2.4 | 2.2 | 23.8 | 57.5 |
| Claw I heights | |||||||||||||
| External primary branch | 20 | 7.5 | – | 10.6 | 20.7 | – | 25.6 | 8.7 | 22.8 | 0.8 | 1.4 | 9.6 | 23.2 |
| External secondary branch | 18 | 6.4 | – | 8.5 | 17.2 | – | 22.3 | 7.3 | 19.0 | 0.7 | 1.5 | 7.9 | 19.1 |
| Internal primary branch | 20 | 6.7 | – | 10.0 | 19.7 | – | 22.6 | 8.2 | 21.2 | 0.9 | 0.9 | 9.2 | 22.2 |
| Internal secondary branch | 18 | 5.1 | – | 8.0 | 15.8 | – | 19.2 | 6.7 | 17.3 | 0.7 | 1.1 | 7.4 | 17.9 |
| Claw II heights | |||||||||||||
| External primary branch | 20 | 7.3 | – | 11.1 | 22.3 | – | 26.0 | 9.2 | 24.0 | 0.9 | 1.0 | 10.3 | 24.9 |
| External secondary branch | 18 | 6.6 | – | 8.8 | 17.6 | – | 22.9 | 7.6 | 19.9 | 0.7 | 1.5 | 7.3 | 17.6 |
| Internal primary branch | 20 | 7.2 | – | 9.6 | 19.2 | – | 23.6 | 8.1 | 21.2 | 0.7 | 1.2 | 8.6 | 20.8 |
| Internal secondary branch | 13 | 6.1 | – | 8.0 | 16.4 | – | 20.3 | 7.1 | 18.4 | 0.6 | 1.4 | 7.1 | 17.1 |
| Claw III heights | |||||||||||||
| External primary branch | 20 | 7.8 | – | 11.1 | 21.4 | – | 27.9 | 9.3 | 24.2 | 0.9 | 1.6 | 10.4 | 25.1 |
| External secondary branch | 18 | 6.4 | – | 8.8 | 17.2 | – | 23.1 | 7.6 | 19.8 | 0.7 | 1.6 | 8.8 | 21.3 |
| Internal primary branch | 19 | 6.0 | – | 9.7 | 16.2 | – | 23.3 | 8.0 | 20.9 | 1.0 | 1.6 | 8.5 | 20.5 |
| Internal secondary branch | 15 | 5.9 | – | 8.1 | 16.9 | – | 20.8 | 7.1 | 18.3 | 0.7 | 1.1 | 8.0 | 19.3 |
| Claw IV heights | |||||||||||||
| Anterior primary branch | 20 | 8.0 | – | 11.2 | 23.4 | – | 28.2 | 9.7 | 25.3 | 0.9 | 1.2 | 10.8 | 26.1 |
| Anterior secondary branch | 18 | 6.5 | – | 8.8 | 17.9 | – | 22.1 | 7.8 | 20.3 | 0.7 | 1.1 | 8.5 | 20.5 |
| Posterior primary branch | 20 | 8.5 | – | 12.1 | 25.0 | – | 30.1 | 10.3 | 27.0 | 0.9 | 1.2 | 11.0 | 26.6 |
| Posterior secondary branch | 9 | 7.7 | – | 9.3 | 20.2 | – | 22.1 | 8.3 | 21.2 | 0.5 | 0.7 | ? | ? |
| Character | N | RANGE | Mean | SD | ||
|---|---|---|---|---|---|---|
| Egg bare diameter | 20 | 53.9 | – | 70.2 | 62.8 | 4.0 |
| Egg full diameter | 20 | 74.7 | – | 94.6 | 85.7 | 4.9 |
| Process height | 60 | 8.3 | – | 15.3 | 11.9 | 1.3 |
| Process base width | 60 | 6.9 | – | 12.5 | 10.0 | 1.1 |
| Process base/height ratio | 60 | 65% | – | 116% | 85% | 11% |
| Inter-process distance | 60 | 1.7 | – | 3.9 | 2.7 | 0.5 |
| Number of processes on the egg circumference | 20 | 15 | – | 18 | 16.2 | 0.8 |
| Character | N | RANGE | Mean | SD | Holotype | ||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| µm | pt | µm | pt | µm | pt | µm | pt | ||||||
| Body length | 20 | 234 | – | 372 | 883 | – | 1042 | 320 | 970 | 32 | 48 | 308 | 982 |
| Buccal tube | |||||||||||||
| Buccal tube length | 20 | 26.5 | – | 37.1 | – | 33.0 | – | 2.7 | – | 31.3 | – | ||
| Stylet support insertion point | 20 | 20.2 | – | 28.7 | 76.2 | – | 77.8 | 25.5 | 77.1 | 2.2 | 0.5 | 24.0 | 76.7 |
| Buccal tube external width | 20 | 4.5 | – | 6.4 | 16.2 | – | 18.4 | 5.6 | 17.0 | 0.5 | 0.6 | 5.3 | 16.9 |
| Buccal tube internal width | 20 | 3.2 | – | 4.9 | 12.0 | – | 14.6 | 4.2 | 12.7 | 0.4 | 0.6 | 3.9 | 12.5 |
| Ventral lamina length | 20 | 15.9 | – | 22.5 | 57.9 | – | 62.5 | 19.9 | 60.2 | 1.6 | 1.1 | 19.2 | 61.3 |
| Placoid lengths | |||||||||||||
| Macroplacoid 1 | 20 | 3.6 | – | 6.1 | 12.9 | – | 16.4 | 4.8 | 14.6 | 0.6 | 0.9 | 4.2 | 13.4 |
| Macroplacoid 2 | 20 | 2.3 | – | 3.8 | 8.5 | – | 10.8 | 3.2 | 9.6 | 0.4 | 0.6 | 3.0 | 9.6 |
| Macroplacoid 3 | 20 | 3.2 | – | 5.0 | 10.6 | – | 13.6 | 4.0 | 12.0 | 0.6 | 1.1 | 3.4 | 10.9 |
| Microplacoid | 20 | 3.0 | – | 4.7 | 9.6 | – | 12.7 | 3.6 | 10.9 | 0.4 | 0.8 | 3.5 | 11.2 |
| Macroplacoid row | 20 | 11.1 | – | 16.9 | 38.9 | – | 45.6 | 14.0 | 42.4 | 1.5 | 1.6 | 12.6 | 40.3 |
| Placoid row | 20 | 15.0 | – | 22.7 | 53.7 | – | 61.2 | 18.8 | 56.7 | 1.9 | 1.8 | 17.3 | 55.3 |
| Claw I heights | |||||||||||||
| External primary branch | 20 | 6.4 | – | 8.6 | 19.5 | – | 25.4 | 7.6 | 23.0 | 0.6 | 1.5 | 7.7 | 24.6 |
| External secondary branch | 16 | 5.1 | – | 7.6 | 15.5 | – | 20.9 | 6.3 | 18.9 | 0.6 | 1.4 | 6.2 | 19.8 |
| Internal primary branch | 20 | 6.2 | – | 8.4 | 18.9 | – | 24.2 | 7.3 | 22.1 | 0.6 | 1.4 | 6.8 | 21.7 |
| Internal secondary branch | 15 | 5.0 | – | 7.1 | 14.3 | – | 19.6 | 6.1 | 18.0 | 0.6 | 1.3 | 5.6 | 17.9 |
| Claw II heights | |||||||||||||
| External primary branch | 20 | 7.0 | – | 8.8 | 21.0 | – | 26.6 | 8.0 | 24.3 | 0.5 | 1.5 | 7.9 | 25.2 |
| External secondary branch | 18 | 5.3 | – | 7.6 | 16.2 | – | 21.7 | 6.5 | 19.5 | 0.5 | 1.3 | 6.8 | 21.7 |
| Internal primary branch | 20 | 6.2 | – | 8.9 | 19.2 | – | 24.5 | 7.3 | 22.1 | 0.6 | 1.5 | 7.1 | 22.7 |
| Internal secondary branch | 19 | 5.0 | – | 7.0 | 16.3 | – | 20.1 | 6.1 | 18.6 | 0.5 | 1.2 | 6.3 | 20.1 |
| Claw III heights | |||||||||||||
| External primary branch | 20 | 7.5 | – | 9.8 | 21.5 | – | 28.3 | 8.2 | 24.9 | 0.6 | 1.7 | 8.2 | 26.2 |
| External secondary branch | 15 | 5.8 | – | 7.6 | 16.6 | – | 22.6 | 6.6 | 20.1 | 0.6 | 1.4 | 6.8 | 21.7 |
| Internal primary branch | 20 | 6.4 | – | 8.8 | 19.8 | – | 25.7 | 7.5 | 22.6 | 0.6 | 1.6 | 7.0 | 22.4 |
| Internal secondary branch | 17 | 5.3 | – | 7.4 | 16.2 | – | 20.9 | 6.1 | 18.7 | 0.6 | 1.3 | 6.2 | 19.8 |
| Claw IV heights | |||||||||||||
| Anterior primary branch | 19 | 7.5 | – | 9.5 | 21.5 | – | 29.1 | 8.3 | 25.1 | 0.6 | 1.8 | 8.1 | 25.9 |
| Anterior secondary branch | 17 | 5.9 | – | 7.5 | 17.5 | – | 22.5 | 6.7 | 20.5 | 0.5 | 1.2 | 6.1 | 19.5 |
| Posterior primary branch | 18 | 8.0 | – | 10.1 | 22.6 | – | 30.6 | 9.0 | 27.2 | 0.6 | 2.0 | 8.5 | 27.2 |
| Posterior secondary branch | 12 | 6.2 | – | 7.9 | 19.2 | – | 22.8 | 7.2 | 21.4 | 0.5 | 1.0 | 7.0 | 22.4 |
| Character | N | RANGE | Mean | SD | ||
|---|---|---|---|---|---|---|
| Egg bare diameter | 7 | 63.4 | – | 69.6 | 67.0 | 2.4 |
| Egg full diameter | 7 | 77.9 | – | 82.2 | 80.0 | 1.4 |
| Process height | 27 | 5.6 | – | 8.8 | 6.9 | 0.9 |
| Process base width | 27 | 3.4 | – | 6.5 | 5.0 | 0.7 |
| Process base/height ratio | 27 | 59% | – | 94% | 73% | 10% |
| Inter-process distance | 27 | 1.5 | – | 3.3 | 2.3 | 0.4 |
| Number of processes on the egg circumference | 7 | 26 | – | 30 | 28.3 | 1.5 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the author. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Stec, D. Integrative Descriptions of Two New Mesobiotus Species (Tardigrada, Eutardigrada, Macrobiotidae) from Vietnam. Diversity 2021, 13, 605. https://doi.org/10.3390/d13110605
Stec D. Integrative Descriptions of Two New Mesobiotus Species (Tardigrada, Eutardigrada, Macrobiotidae) from Vietnam. Diversity. 2021; 13(11):605. https://doi.org/10.3390/d13110605
Chicago/Turabian StyleStec, Daniel. 2021. "Integrative Descriptions of Two New Mesobiotus Species (Tardigrada, Eutardigrada, Macrobiotidae) from Vietnam" Diversity 13, no. 11: 605. https://doi.org/10.3390/d13110605
APA StyleStec, D. (2021). Integrative Descriptions of Two New Mesobiotus Species (Tardigrada, Eutardigrada, Macrobiotidae) from Vietnam. Diversity, 13(11), 605. https://doi.org/10.3390/d13110605