
Western Pacific Zooplankton Community along Latitudinal and Equatorial Transects in Autumn 2017 (Northern Hemisphere)



Abstract
:1. Introduction
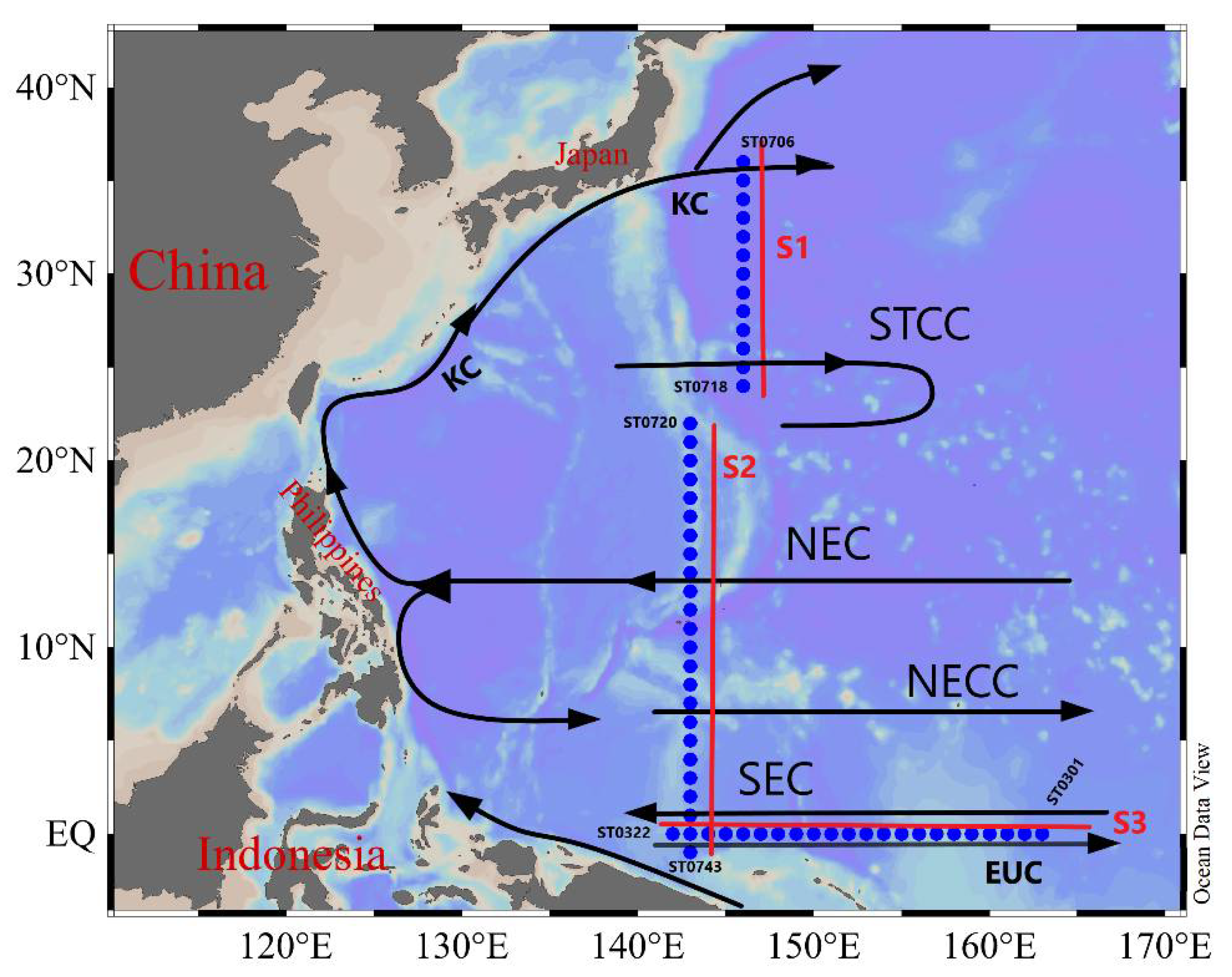
2. Materials and Methods
3. Results
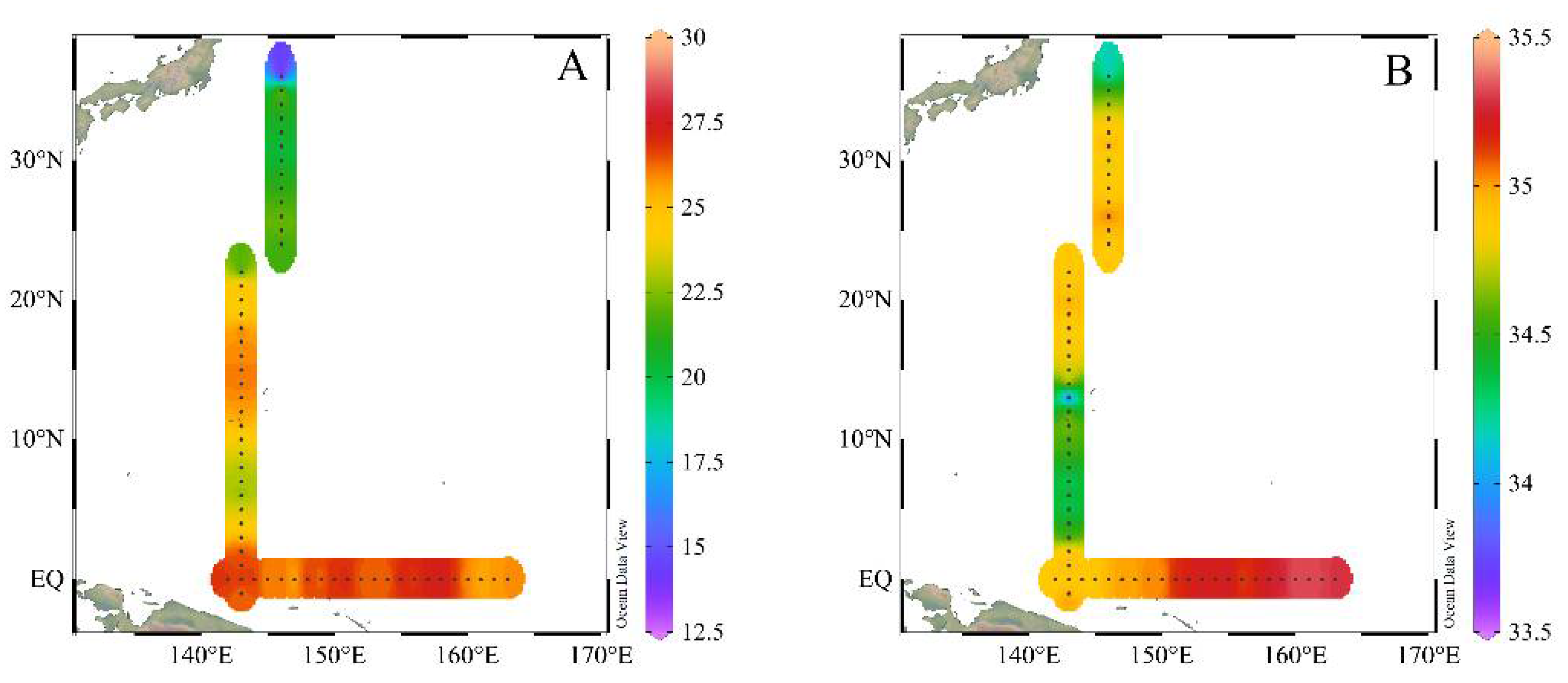
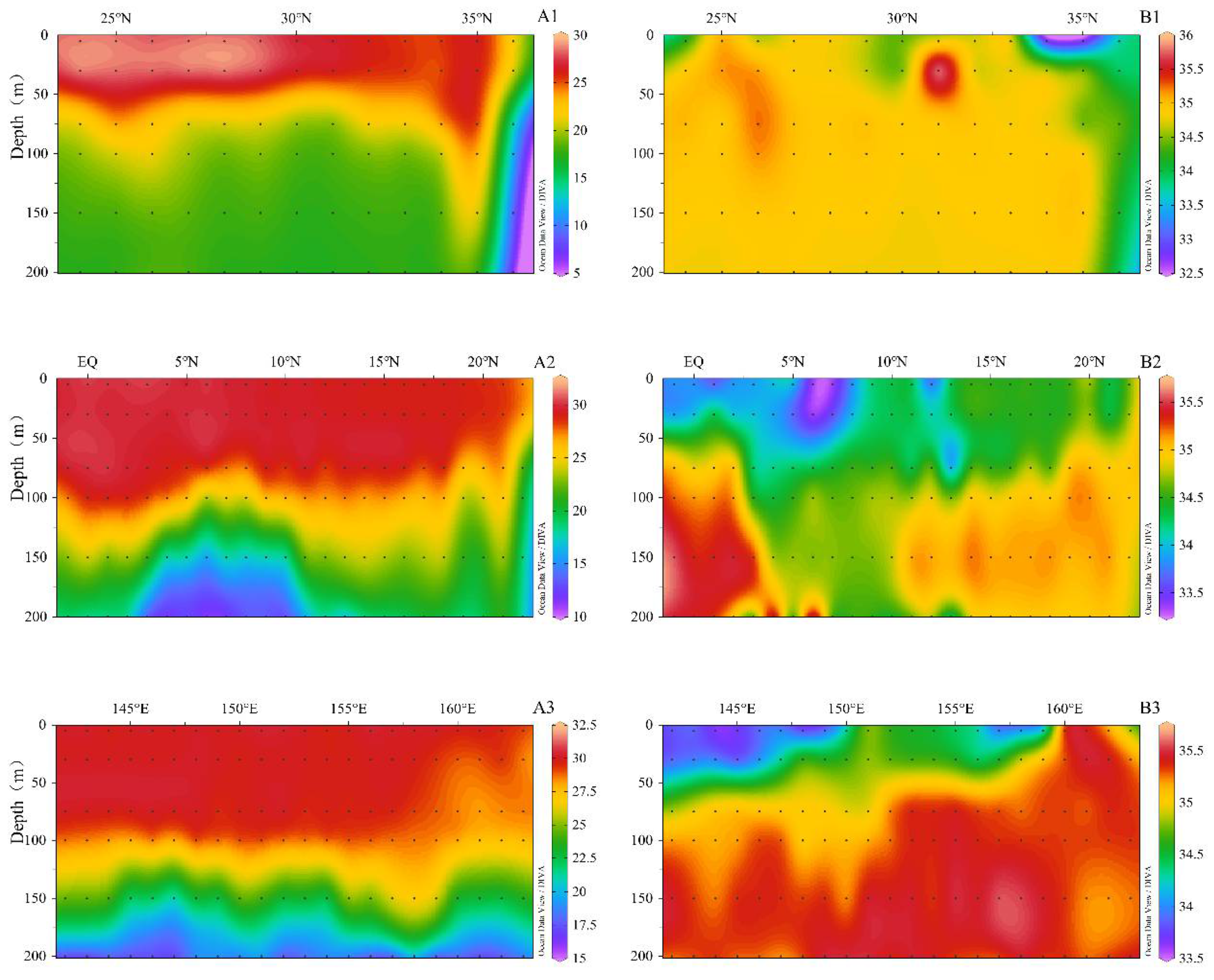
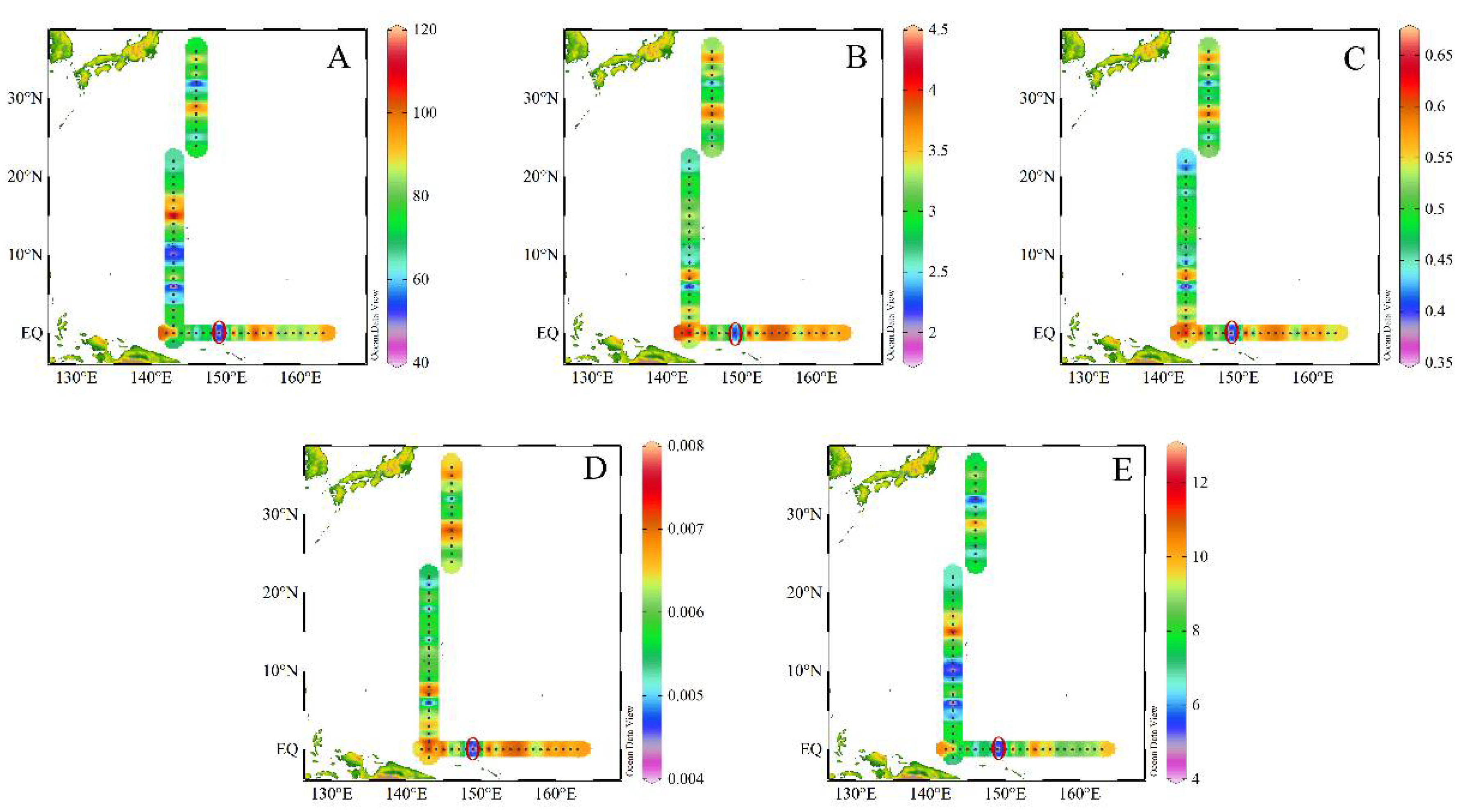
3.1. Hydrography of the Study Area
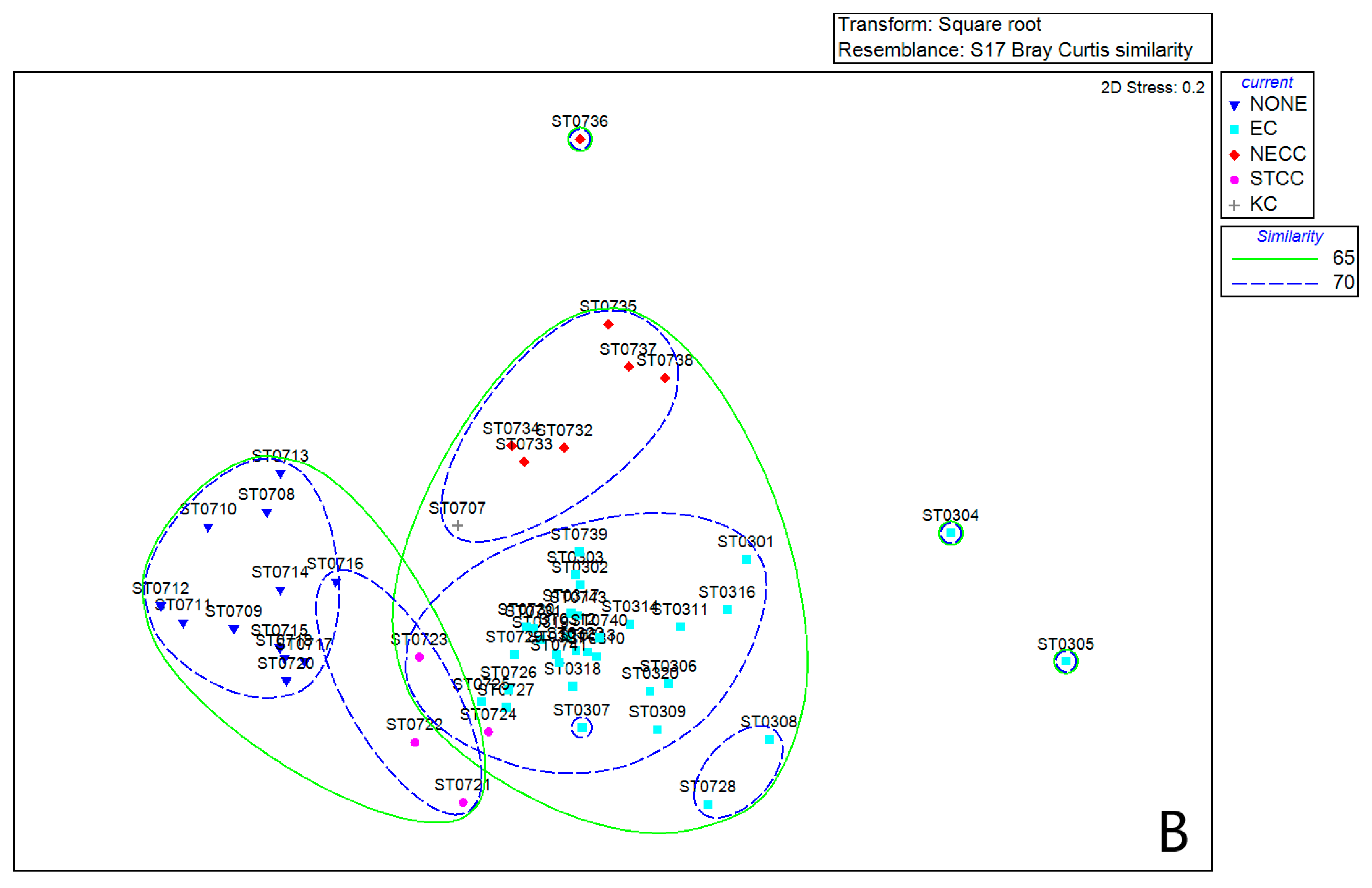
3.2. Zooplankton Species Composition
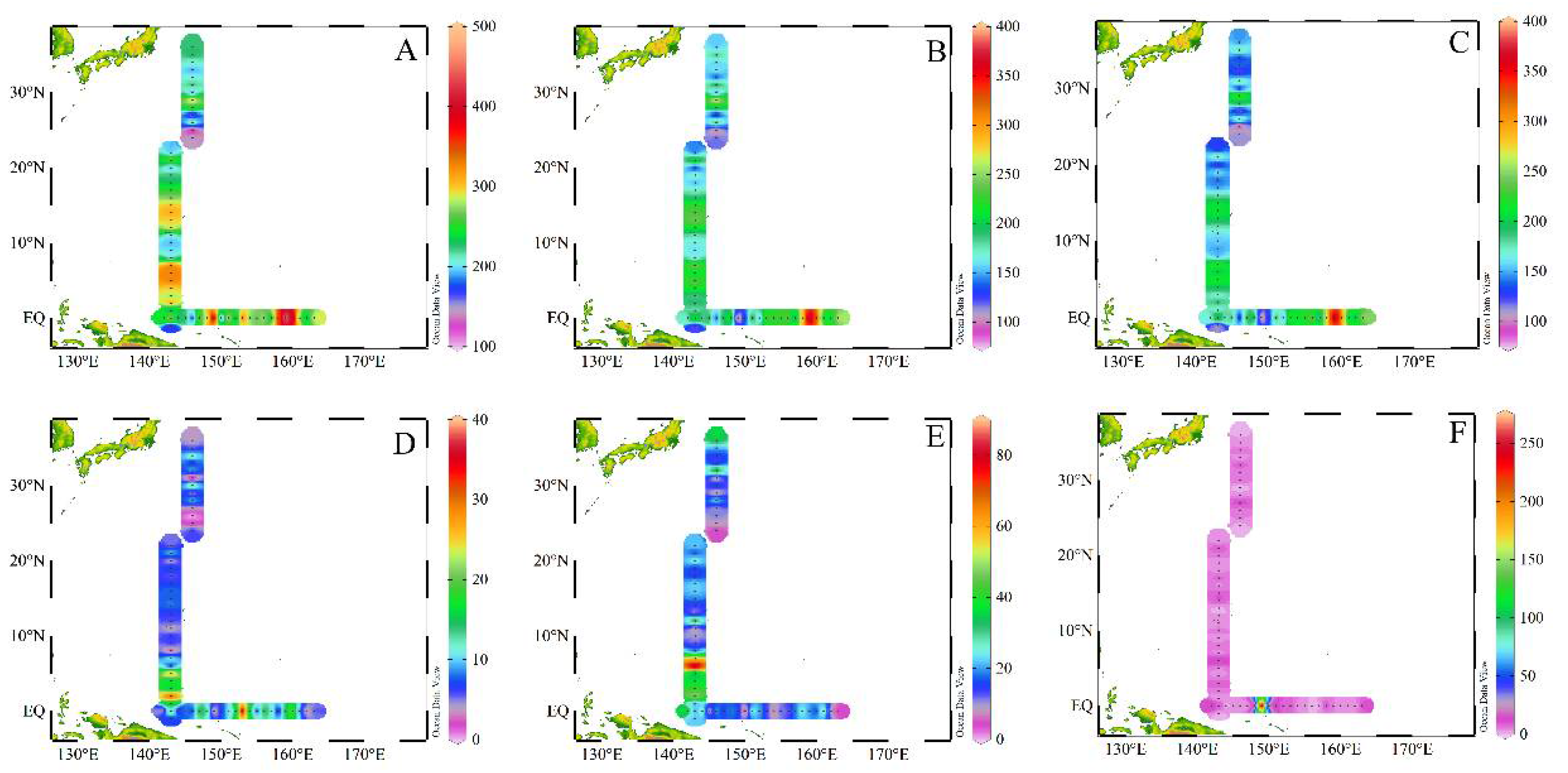
3.3. Zooplankton Abundance Distribution
3.4. Diversity Indices of the Zooplankton Community
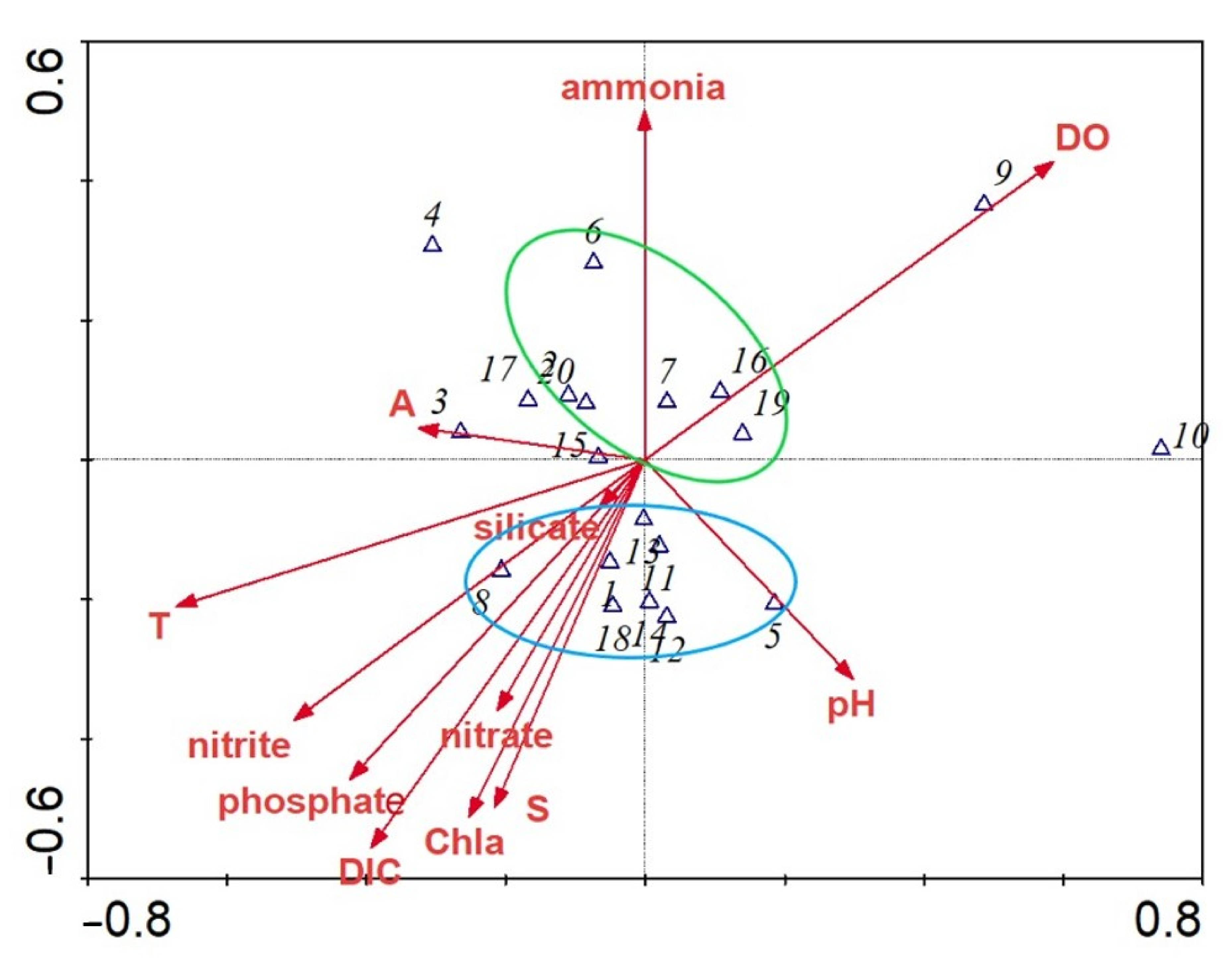
3.5. The Relationship between Zooplankton and the Environment
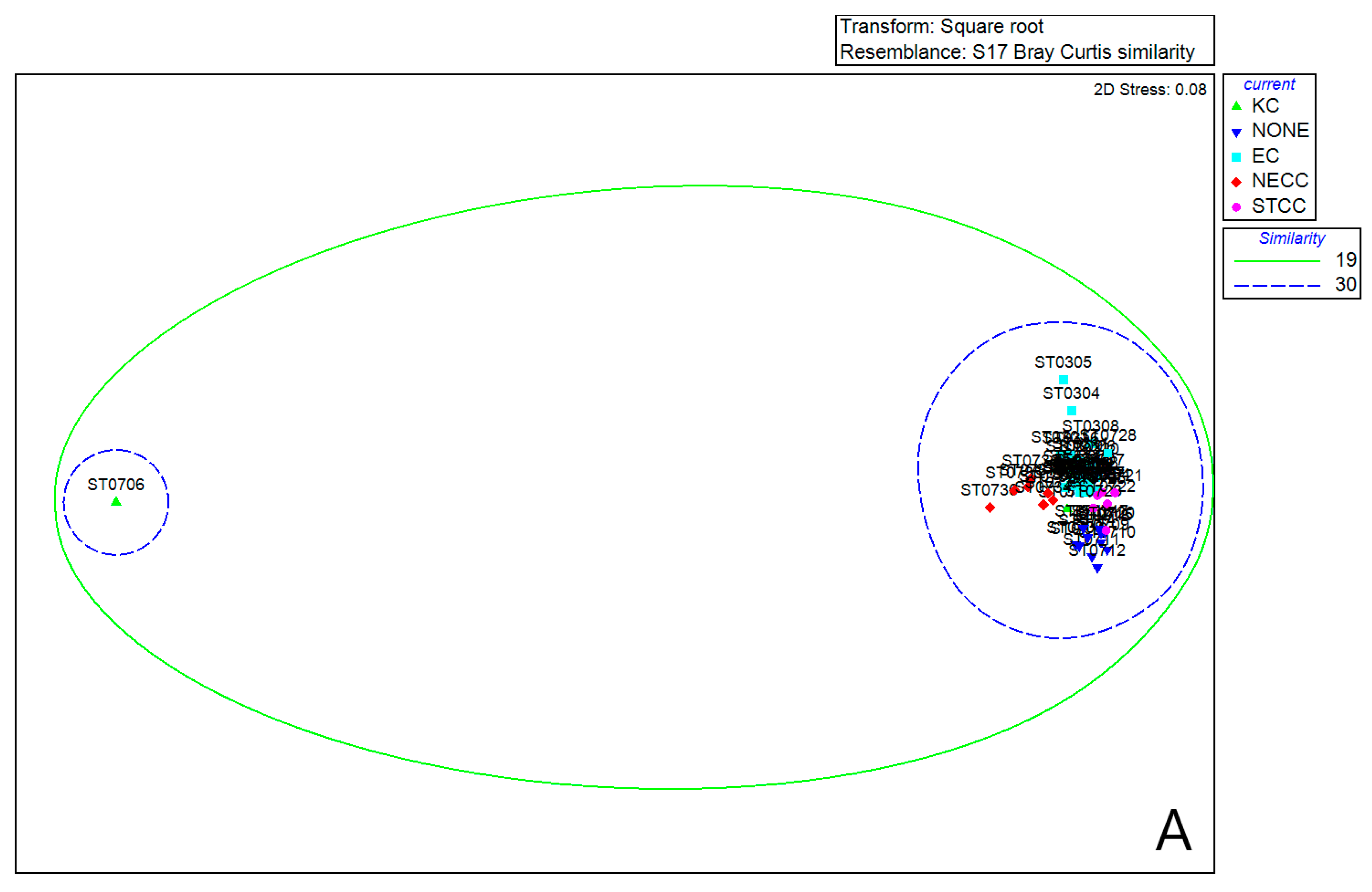
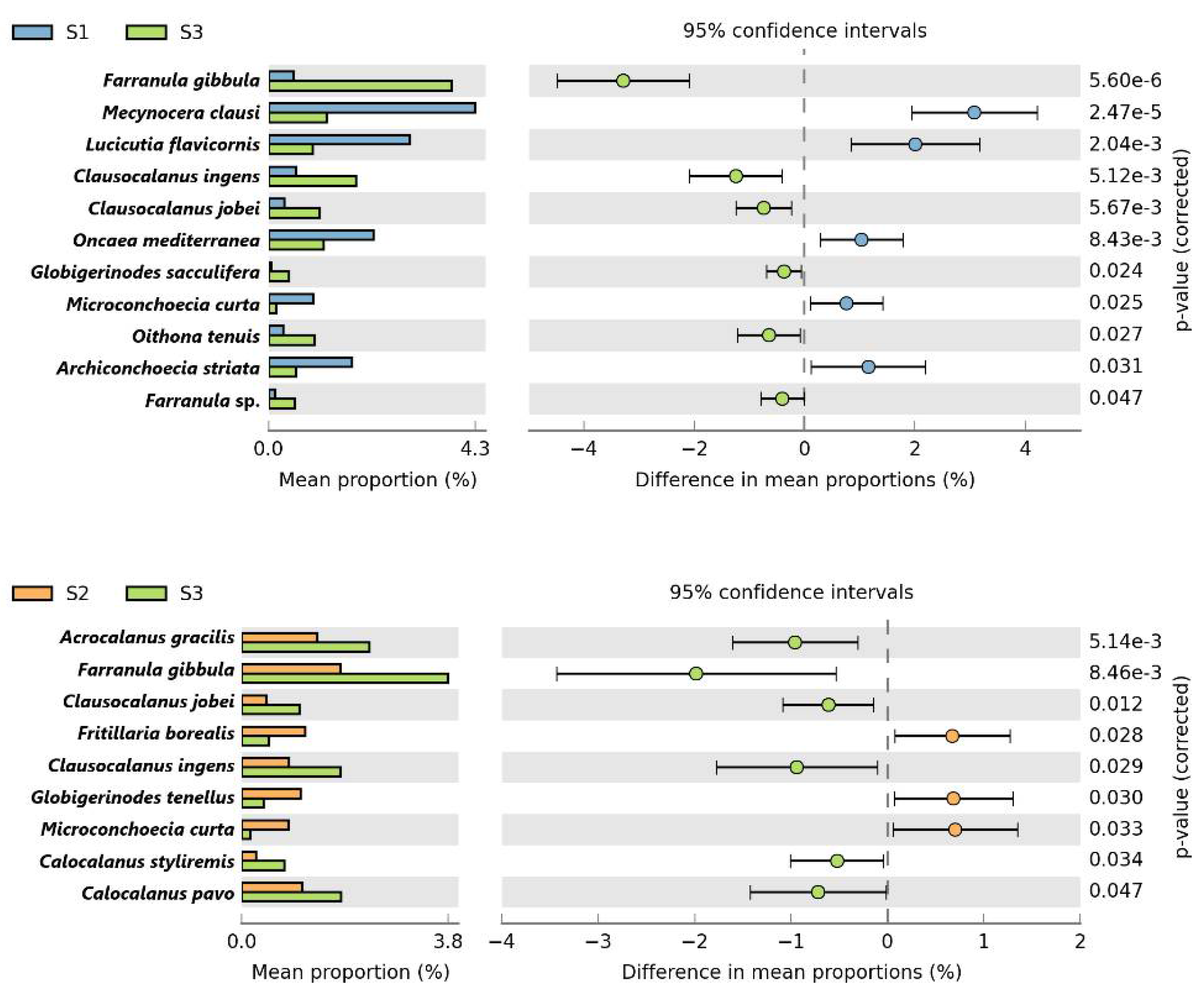
3.6. Comparison of Equatorial Zooplankton Communities with S1 and S2
4. Discussion
4.1. Comparison with Historical Data
4.2. Relationship between Ocean Current Factors and Zooplankton
4.3. Relationship between Other Factors and Zooplankton
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Allain, V.; Fernandez, E.; Hoyle, S.D.; Caillot, S.; Juradomolina, J.; Andrefouet, S.; Nicol, S.J. Interaction between coastal and oceanic ecosystems of the Western and Central Pacific Ocean through predator-prey relationship studies. PLoS ONE 2012, 7, 1–9. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Morato, T.; Hoyle, S.D.; Allain, V.; Nicol, S.J. Seamounts are hotspots of pelagic biodiversity in the open ocean. Proc. Natl. Acad. Sci. USA 2010, 107, 9707–9711. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Longhurst, A.R. Chapter 11—The pacific ocean. In Ecological Geography of the Sea, 2nd ed.; Longhurst, A.R., Ed.; Academic Press: Burlington, MA, USA, 2007; pp. 327–441. [Google Scholar]
- Hu, S.; Hu, D. Review on western pacific warm pool study. Stud. Mar. Sin. 2016, 51, 37–48. (In Chinese) [Google Scholar]
- Grenier, M.; Cravatte, S.; Blanke, B.; Menkes, C.E.; Kochlarrouy, A.; Durand, F.; Melet, A.; Jeandel, C. From the western boundary currents to the Pacific Equatorial Undercurrent: Modeled pathways and water mass evolutions. J. Geophys. Res. 2011, 116, C12044. [Google Scholar] [CrossRef] [Green Version]
- Lin, M.; Wang, C.; Wang, Y.; Xiang, P.; Wang, Y.; Lian, G.; Chen, R.; Chen, X.; Ye, Y.; Dai, Y.; et al. Zooplanktonic diversity in the western Pacific. Biodivers. Sci. 2011, 19, 646–654. (In Chinese) [Google Scholar]
- Tittensor, D.P.; Mora, C.; Jetz, W.; Lotze, H.K.; Ricard, D.; Berghe, E.V.; Worm, B. Global patterns and predictors of marine biodiversity across taxa. Nature 2010, 466, 1098–1101. [Google Scholar] [CrossRef]
- Allen, G.R. Conservation hotspots of biodiversity and endemism for Indo—Pacific coral reef fishes. Aquat. Conserv. Mar. Freshwat. Ecosyst. 2008, 18, 541–556. [Google Scholar] [CrossRef]
- Briggs, J.C. The marine East Indies: Diversity and speciation. J. Biogeogr. 2005, 32, 1517–1522. [Google Scholar] [CrossRef]
- Karl, D.M. A sea of change: Biogeochemical variability in the North Pacific Subtropical Gyre. Ecosystems 1999, 2, 181–214. [Google Scholar] [CrossRef]
- Taniguchi, A. Phytoplankton-Zooplankton relationships in the western Pacific Ocean and adjacent seas. Mar. Biol. 1973, 21, 115–121. [Google Scholar] [CrossRef]
- Sun, J.; Liu, D.; Wang, Z.; Zhu, M. The effects of zooplankton grazing on the development of red tides. Acta Ecol. Sin. 2004, 24, 1514–1522. [Google Scholar]
- Calbet, A.; Landry, M.R. Phytoplankton growth, microzooplankton grazing, and carbon cycling in marine systems. Limnol. Oceanogr. 2004, 49, 51–57. [Google Scholar] [CrossRef] [Green Version]
- Sun, J.; Li, X.; Chen, J.; Guo, S. Progress in oceanic biological pump. Acta Oceanol. Sin. 2016, 38, 1–21. [Google Scholar] [CrossRef]
- Menkes, C.E.; Allain, V.; Rodier, M.; Gallois, F.; Lebourges-Dhaussy, A.; Hunt, B.P.V.; Smeti, H.; Pagano, M.; Josse, E.; Daroux, A.; et al. Seasonal oceanography from physics to micronekton in the south-west Pacific. Deep Sea Res. II 2015, 113, 125–144. [Google Scholar] [CrossRef] [Green Version]
- Houssem, S.; Marc, P.; Christophe, M.; Anne, L.-D.; Hunt, B.P.V.; Valerie, A.; Martine, R.; de Boissieu, F.; Elodie, K.; Cherif, S. Spatial and temporal variability of zooplankton off New Caledonia (Southwestern Pacific) from acoustics and net measurements. J. Geophys. Res. Ocean. 2015, 120, 2676–2700. [Google Scholar]
- Dai, L.; Li, C.; Sun, X.; Ji, P.; Zhang, W. Abundance and biomass of zooplankton in Philippine Sea in winter 2012. Oceanol. Limnol. Sin. 2014, 45, 1225–1233. (In Chinese) [Google Scholar]
- Yang, G.; Li, C.; Wang, Y.; Wang, X.; Dai, L.; Tao, Z.; Ji, P. Spatial variation of the zooplankton community in the western tropical Pacific Ocean during the summer of 2014. Cont. Shelf Res. 2017, 135, 14–22. [Google Scholar] [CrossRef]
- Sun, D.; Wang, C. Latitudinal distribution of zooplankton communities in the Western Pacific along 160° E during summer 2014. J. Mar. Syst. 2017, 169, 52–60. [Google Scholar] [CrossRef]
- Le Borgne, R.; Rodier, M. Net zooplankton and the biological pump: A comparison between the oligotrophic and mesotrophic equatorial Pacific. Deep Sea Res. II 1997, 44, 2003–2023. [Google Scholar]
- Roman, M.R.; Dam, H.G.; Gauzens, A.L.; Urban-Rich, J.; Foley, D.G.; Dickey, T.D. Zooplankton variability on the equator at 140° W during the JGOFS EqPac study. Deep Sea Res. II 1995, 42, 673–693. [Google Scholar] [CrossRef]
- Ishizaka, J.; Harada, K.; Ishikawa, K.; Kiyosawa, H.; Furusawa, H.; Watanabe, Y.; Ishida, H.; Suzuki, K.; Handa, N.; Takahashi, M. Size and taxonomic plankton community structure and carbon flow at the equator, 175‡E during 1990–1994. Deep Sea Res. II 1997, 44, 1927–1949. [Google Scholar] [CrossRef]
- Hu, D.; Wu, L.; Cai, W.; Gupta, A.S.; Ganachaud, A.; Qiu, B.; Gordon, A.L.; Lin, X.; Chen, Z.; Hu, S.; et al. Pacific western boundary currents and their roles in climate. Nature 2015, 522, 299–308. [Google Scholar] [CrossRef] [PubMed]
- Grasshoff, K.; Kremling, K.; Ehrhardt, M. Methods of Seawater Analysis, 3rd ed.; Wiley: Hoboken, NJ, USA, 1999. [Google Scholar]
- Sun, J.; Liu, D. The application of diversity indices in marine phytoplankton studies. Acta Oceanol. Sin. 2004, 26, 62–75. (In Chinese) [Google Scholar]
- Ma, C.; Wang, Z.; Zhou, E. Characteristics of currents in the west Pacific Ocean. Adv. Mar. Sci. 1983, 1, 24–36. [Google Scholar]
- Ganachaud, A.; Cravatte, S.; Melet, A.; Schiller, A.; Holbrook, N.J.; Sloyan, B.M.; Widlansky, M.J.; Bowen, M.; Verron, J.; Wiles, P.; et al. The Southwest Pacific Ocean circulation and climate experiment (SPICE). J. Geophys. Res. 2014, 119, 7660–7686. [Google Scholar] [CrossRef]
- Graham, N.E.; Barnett, T.P. Sea surface temperature, surface wind divergence, and convection over tropical oceans. Science 1987, 238, 657–659. [Google Scholar] [CrossRef] [PubMed]
- Bai, X.; Yang, F.; Dong, M.; Wu, Y.; Chen, H. The vertical distribution of nutrients in the upper water of the equatorial western Pacific Ocean in autumn. Period. Ocean Univ. China 2019, 49, 64–71. (In Chinese) [Google Scholar]
- Lan, S. Summery basic characteristics of temperature and salinity at 155° E section. Stud. Mar. Sin. 1999, 00, 1–11. [Google Scholar]
- Kobashi, F.; Kawamura, H. Seasonal variation and instability nature of the North Pacific Subtropical Countercurrent and the Hawaiian Lee Countercurrent. J. Geophys. Res. Ocean. 2002, 107, 6-1–6-18. [Google Scholar] [CrossRef]
- Liu, H.; Huang, Y.; Zhai, W.; Guo, S.; Jin, H.; Sun, J. Phytoplankton communities and its controlling factors in summer and autumn in the southern Yellow Sea, China. Acta Oceanol. Sin. 2015, 34, 114–123. [Google Scholar] [CrossRef]
- Neumann-Leitão, S.; Sant’anna, E.M.E.; Gusmão, L.M.D.O.; Do Nascimento-Vieira, D.A.; Paranaguá, M.N.; Schwamborn, R. Diversity and distribution of the mesozooplankton in the tropical Southwestern Atlantic. J. Plankton Res. 2008, 30, 795–805. [Google Scholar] [CrossRef]
- Chiba, S.; Batten, S.D.; Yoshiki, T.; Sasaki, Y.; Sasaoka, K.; Sugisaki, H.; Ichikawa, T. Temperature and zooplankton size structure: Climate control and basin-scale comparison in the North Pacific. Ecol. Evol. 2015, 5, 968–978. [Google Scholar] [CrossRef] [PubMed]
- Pielou, E.C. Mathematical Ecology, 2nd ed.; Wiley & Sons: New York, NY, USA, 1977; p. 47. [Google Scholar]
- Zervoudaki, S.; Christou, E.D.; Nielsen, T.G.; Siokoufrangou, I.; Assimakopoulou, G.; Giannakourou, A.; Maar, M.; Pagou, K.; Krasakopoulou, E.; Christaki, U. The importance of small-sized copepods in a frontal area of the Aegean Sea. J. Plankton Res. 2007, 29, 317–338. [Google Scholar]
- Hernroth, L. Sampling and filtration efficiency of two commonly used plankton nets. A comparative study of the Nansen net and the Unesco WP 2 net. J. Plankton Res. 1987, 9, 719–728. [Google Scholar] [CrossRef]
- Evans, M.S.; Sell, D.W. Mesh size and collection characteristics of 50-cm diameter conical plankton nets. Hydrobiologia 1985, 122, 97–104. [Google Scholar] [CrossRef]
- Wen-Tseng, L.; Chang-Tai, S.; Jiang-Shiou, H. Diel vertical migration of the planktonic copepods at an upwelling station north of Taiwan, western North Pacific. J. Plankton Res. 2004, 26, 89–97. [Google Scholar]
- Li, K.; Yin, J.; Huang, L.; Tan, Y.; Lin, Q. A comparison of the zooplankton community in the Bay of Bengal and South China Sea during April–May, 2010. J. Ocean Univ. China 2017, 16, 1206–1212. [Google Scholar] [CrossRef]
- Sabatini, M.; Reta, R.; Matano, R. Circulation and zooplankton biomass distribution over the southern Patagonian shelf during late summer. Cont. Shelf Res. 2004, 24, 1359–1373. [Google Scholar] [CrossRef]
- Qiu, B. Seasonal Eddy Field Modulation of the North Pacific Subtropical Countercurrent: TOPEX/Poseidon Observations and Theory. J. Phys. Oceanogr. 1999, 29, 2471–2486. [Google Scholar] [CrossRef]
- Feng, S.; Li, F.; Li, S. Introduction to Ocean Science; Higher Education Press: Beijing, China, 1999; pp. 167–172. [Google Scholar]
- Hidaka, K.; Kawaguchi, K.; Tanabe, T.; Takahashi, M.; Kubodera, T. Biomass and taxonomic composition of micronekton in the western tropical-subtropical Pacific. Fish. Oceanogr. 2010, 12, 112–125. [Google Scholar] [CrossRef]
- Peterson, W.T.; Miller, C.B.; Hutchinson, A. Zonation and maintenance of copepod populations in the Oregon upwelling zone. Deep Sea Res. Part A Oceanogr. Res. Pap. 1979, 26, 467–494. [Google Scholar] [CrossRef]
- Sommer, U. Plankton ecology: Succession in plankton communities. In Brock/Springer Series in Contemporary Bioscience; Springer: Berlin/Heidelberg, Germany, 1989. [Google Scholar]
- Zheng, Z. Introduction to Planktonology; Science Press: Beijing, China, 1964; pp. 78–81. [Google Scholar]
- Lo, W.-T.; Dahms, H.-U.; Hwang, J.-S. Water mass transport through the northern Bashi Channel in the northeastern South China Sea affects copepod assemblages of the Luzon Strait. Zool. Stud. 2014, 53, 66. [Google Scholar] [CrossRef]
- Banse, K. Zooplankton: Pivotal role in the control of ocean production: I. Biomass and production. ICES J. Mar. Sci. 1995, 52, 265–277. [Google Scholar] [CrossRef]
- Quintana, X.D.; Arim, M.; Badosa, A.; Blanco, J.M.; Boix, D.; Brucet, S.; Compte, J.; Egozcue, J.J.; de Eyto, E.; Gaedke, U.; et al. Predation and competition effects on the size diversity of aquatic communities. Aquat. Sci. 2015, 77, 45–57. [Google Scholar] [CrossRef]
- Dagg, M. Ingestion of phytoplankton by the micro- and mesozooplankton communities in a productive subtropical estuary. J. Plankton Res. 1995, 17, 845–857. [Google Scholar] [CrossRef]
- White, J.R.; Zhang, X.; Welling, L.A.; Roman, M.R.; Dam, H.G. Latitudinal gradients in zooplankton biomass in the tropical Pacific at 140° W during the JGOFS EqPac study: Effects of El Niño. Deep Sea Res. Part II 1995, 42, 715–733. [Google Scholar] [CrossRef]
- Chen, Z.; Sun, J. Effects of ocean currents in the western Pacific Ocean on net-phytoplankton. manuscript in preparation.
- Peralba, U.; Mazzocchi, M.G.; Harris, R.P. Niche separation and reproduction of Clausocalanus species (Copepoda, Calanoida) in the Atlantic Ocean. Prog. Oceanogr. 2017, 158, 185–202. [Google Scholar] [CrossRef]
- Huang, B.; Wu, J.; Tang, J.; Hu, J.; Wang, J. The study on the correlation between zooplankton community and water mass in the Hangzhou Bay. Acta Oceanol. Sin. 2010, 32, 170–175. (In Chinese) [Google Scholar]










{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Section | |||
|---|---|---|---|
| S1 | S2 | S3 | |
| Location | 146° E, 24–36° N | 143° E, –1°–22° N | 0° N, 142–163° E |
| Temperature (°C) | 13.24–23.20 | 13.24–23.20 | 13.24–23.20 |
| Salinity | 34.12–35.06 | 33.65–35.00 | 34.88–35.34 |
| The top 10 dominant species (order of dominance from greatest to least) | Acrocalanus gibber | Acrocalanus gibber | Acrocalanus gracilis |
| Paracalanus aculeatus | Canthocalanus pauper | Paracalanus aculeatus | |
| Mecynocera clausi | Oithona similis | Farranula gibbula | |
| Oncaea venusta | Paracalanus aculeatus | Canthocalanus pauper | |
| Canthocalanus pauper | Oncaea venusta | Oncaea venusta | |
| Oithona similis | Nannocalanus minor | Oithona similis | |
| Lucicutia flavicornis | Clausocalanus furcatus | Acrocalanus gibber | |
| Oncaea mediterranea | Farranula concinna | Acartia negligens | |
| Parvocalanus crassirostris | Cosmocalanus darwinii | Cosmocalanus darwinii | |
| Acrocalanus gracili | Farranula rostrata | Oithona plumifera | |
| Species | DO | pH | Silicate | Phosphate | Nitrite | Nitrate | Ammonia | DIC | Temperature | Chl. a |
|---|---|---|---|---|---|---|---|---|---|---|
| H’ | −0.198 | 0.245 | −0.018 | 0.114 | 0.280 * | 0.062 | −0.195 | 0.050 | 0.222 | 0.314 * |
| J | −0.252 | 0.222 | 0.028 | 0.178 | 0.260 * | 0.131 | −0.180 | 0.104 | 0.200 | 0.319 * |
| Copepods | −0.348 ** | 0.057 | −0.001 | 0.249 | 0.495 ** | 0.127 | −0.058 | 0.217 | 0.325 * | 0.528 ** |
| Canthocalanus pauper | −0.246 | −0.148 | 0.047 | 0.168 | 0.233 | 0.119 | 0.053 | 0.132 | 0.238 | 0.244 |
| Cosmocalanus darwinii | −0.200 | −0.040 | 0.053 | 0.185 | 0.232 | 0.118 | −0.059 | 0.147 | 0.210 | 0.314 * |
| Calocalanus pavo | −0.118 | 0.269 * | −0.070 | 0.093 | 0.075 | 0.057 | −0.177 | 0.092 | 0.105 | 0.230 |
| Clausocalanus arcuicornis | −0.274 * | 0.094 | 0.000 | 0.213 | 0.418 ** | 0.101 | −0.021 | 0.285 * | 0.221 | 0.420 ** |
| Lucicutia flavicornis | 0.285 * | 0.088 | −0.145 | −0.289 * | −0.070 | −0.222 | 0.331 * | −0.263 * | −0.254 | −0.075 |
| Acrocalanus gibber | −0.337 ** | −0.011 | 0.109 | 0.309 * | 0.427 ** | 0.246 | −0.064 | 0.319 * | 0.133 | 0.486 |
| Paracalanus aculeatus | −0.275 * | 0.143 | 0.055 | 0.263 * | 0.369 ** | 0.188 | −0.083 | 0.332 * | 0.164 | 0.427 |
| Oithona similis | −0.130 | −0.069 | 0.043 | 0.131 | 0.413 ** | 0.068 | 0.041 | 0.084 | 0.169 | 0.295 |
| Farranula gibbula | −0.34** | 0.065 | −0.078 | 0.201 | 0.286 | 0.08 * | −0.143 | 0.207 | 0.487 ** | 0.249 |
| Clausocalanus ingens | −0.364** | 0.077 | −0.014 | 0.0252 | 0.465** | 0.162 | −0.044 | 0.268 * | 0.298 * | 0.465 ** |
| Clausocalanus jobei | −0.316* | 0.048 | 0.078 | 0.0242 | 0.349** | 0.159 | −0.17 | 0.278 * | 0.289 * | 0.27 * |
| Time | Research Area | References Method and Nesh Size | Depth | Number of Species and Larvae | Abundance (ind./m3) | Reference |
|---|---|---|---|---|---|---|
| Nov.–Dec. 2012 | 120–130° E, 0–20° N | ZooScan 500 um | 0–200 m | - | 11–116 | Dai et al. [17] |
| Aug.–Oct. 2014 | 120°–135° E, 0–20°N | Microscope 200 um | 0–300 m | 259 | Subregional average: 86.09–311.98 | Yang et al. [18] |
| June–July 2014 | 160° E 4°S–46° N | Microscope 200 um | 0–200 m | 498 | 45.11–439.84 | Sun et al. [19] |
| Oct.–Dec. 2017 | 142–164° E, –1–36° N | Microscope 200 um | 0–200 m | 405 | 118.33–452.22 | this study |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Long, Y.; Noman, M.A.; Chen, D.; Wang, S.; Yu, H.; Chen, H.; Wang, M.; Sun, J. Western Pacific Zooplankton Community along Latitudinal and Equatorial Transects in Autumn 2017 (Northern Hemisphere). Diversity 2021, 13, 58. https://doi.org/10.3390/d13020058
Long Y, Noman MA, Chen D, Wang S, Yu H, Chen H, Wang M, Sun J. Western Pacific Zooplankton Community along Latitudinal and Equatorial Transects in Autumn 2017 (Northern Hemisphere). Diversity. 2021; 13(2):58. https://doi.org/10.3390/d13020058
Chicago/Turabian StyleLong, Yi, Md Abu Noman, Dawei Chen, Shihao Wang, Hao Yu, Hongtao Chen, Min Wang, and Jun Sun. 2021. "Western Pacific Zooplankton Community along Latitudinal and Equatorial Transects in Autumn 2017 (Northern Hemisphere)" Diversity 13, no. 2: 58. https://doi.org/10.3390/d13020058
APA StyleLong, Y., Noman, M. A., Chen, D., Wang, S., Yu, H., Chen, H., Wang, M., & Sun, J. (2021). Western Pacific Zooplankton Community along Latitudinal and Equatorial Transects in Autumn 2017 (Northern Hemisphere). Diversity, 13(2), 58. https://doi.org/10.3390/d13020058