
Influence of Habitat on Presence of Striped Skunks in Midwestern North America


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Materials and Methods
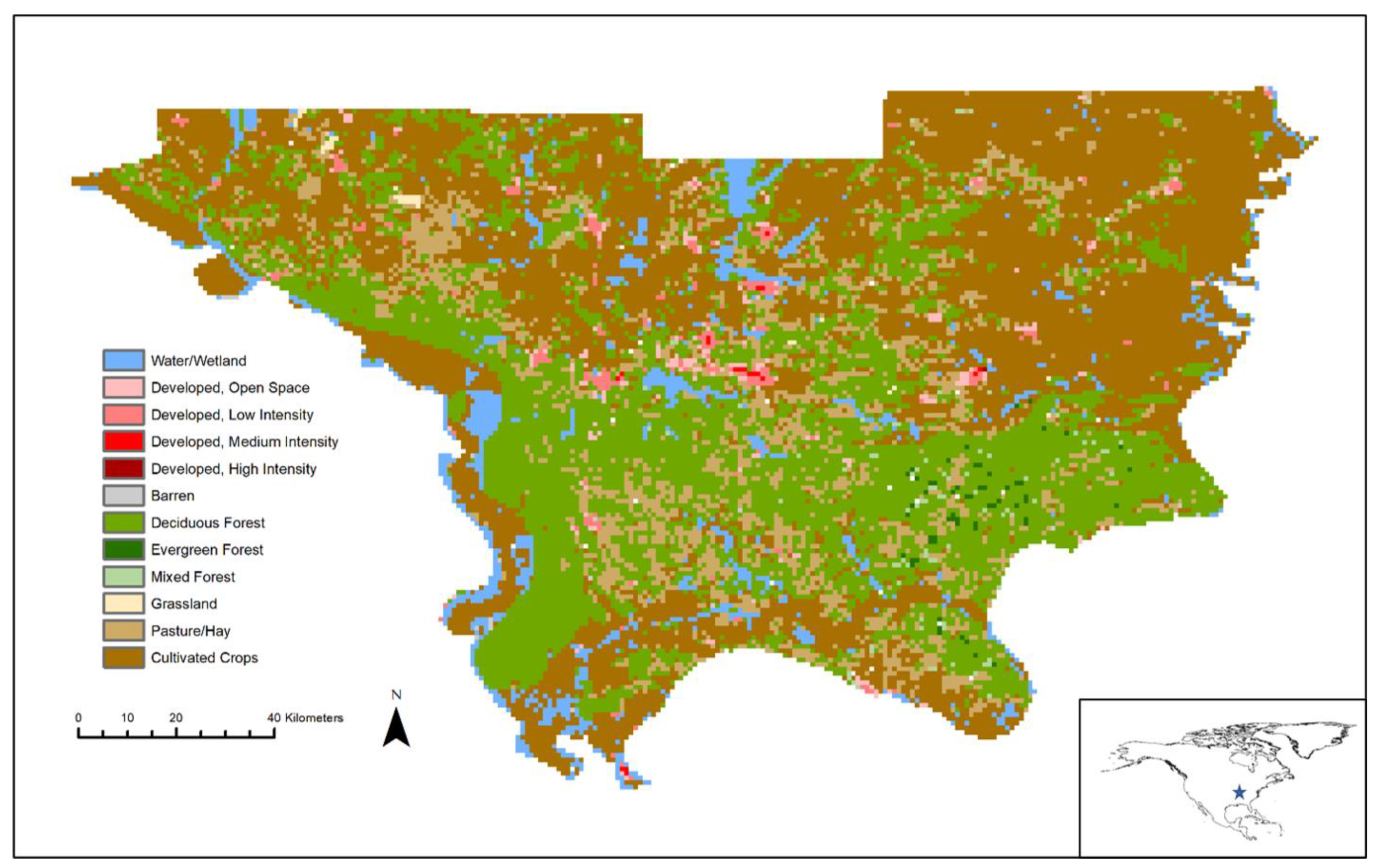
2.1. Study Area
2.2. Striped Skunk Presence
2.3. Environmental Variables
2.4. Species Distribution Modeling
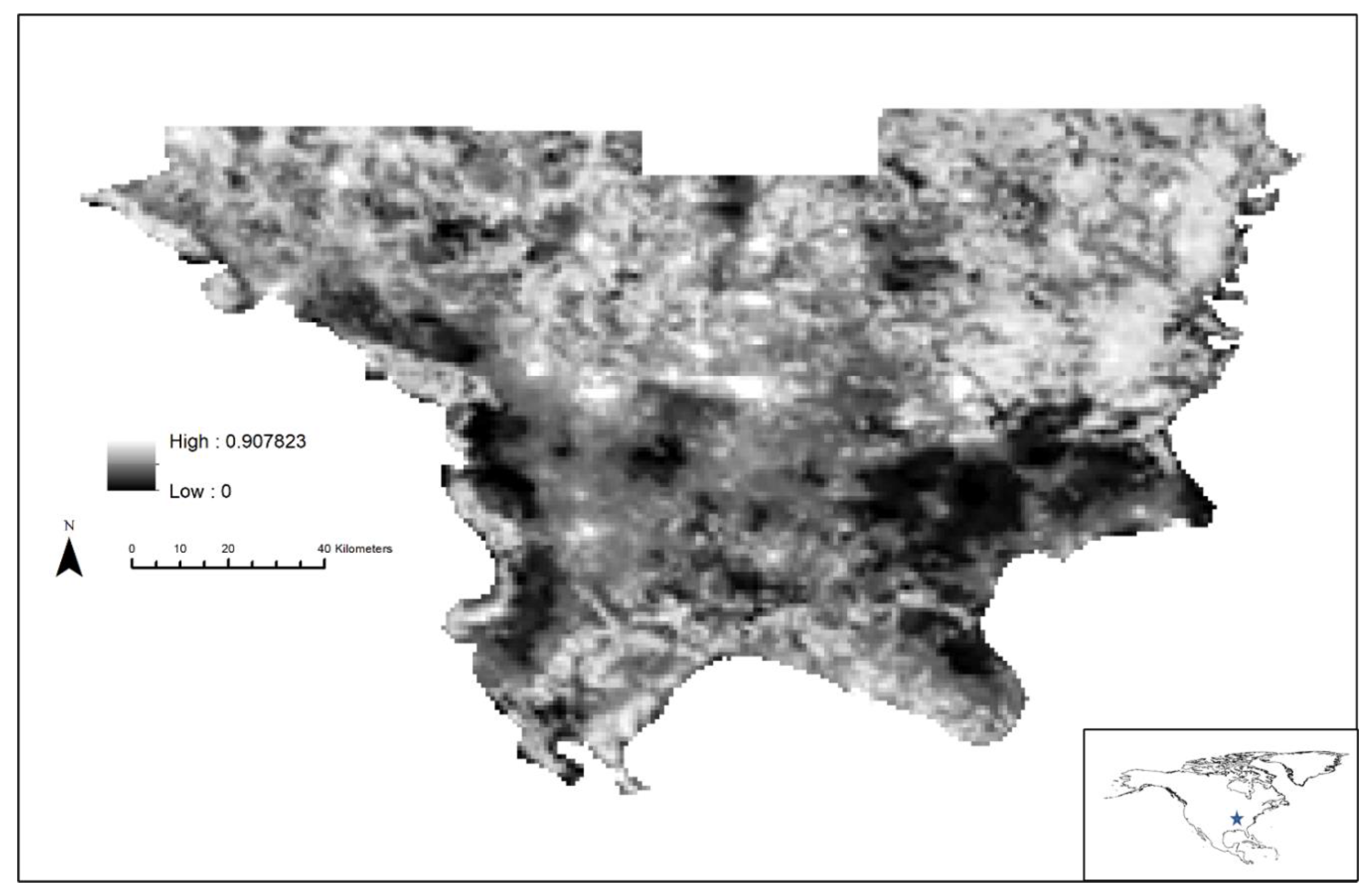
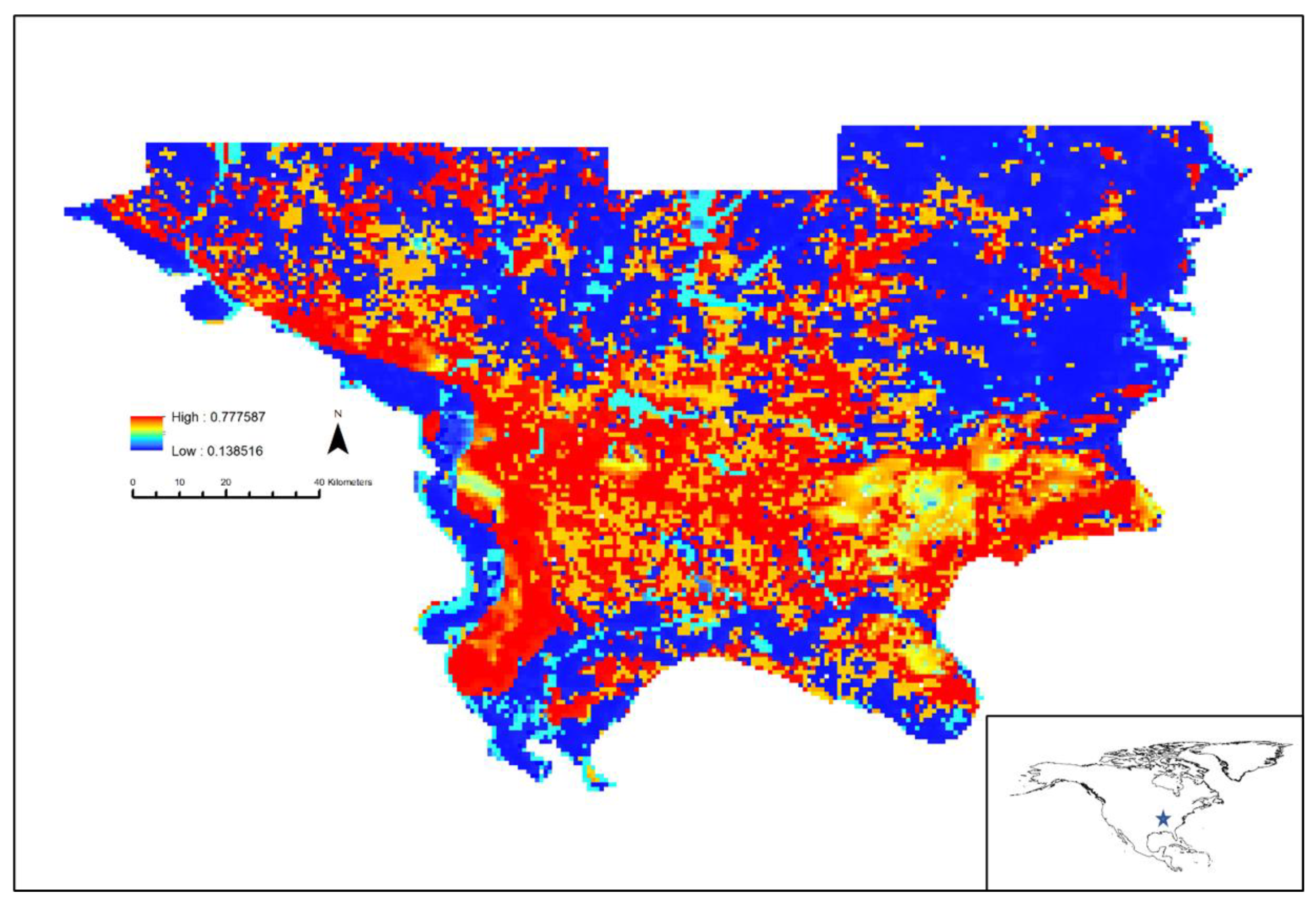
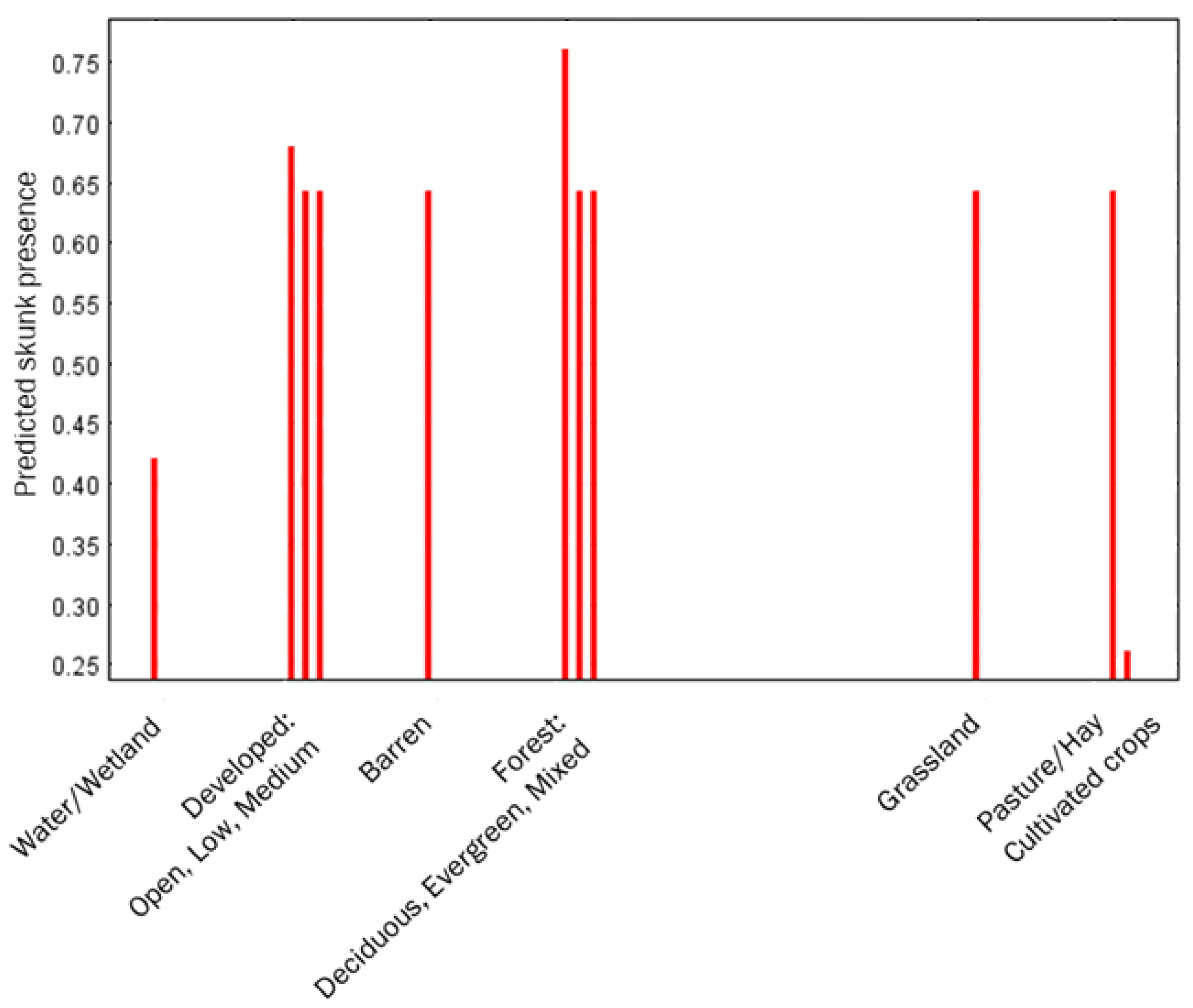
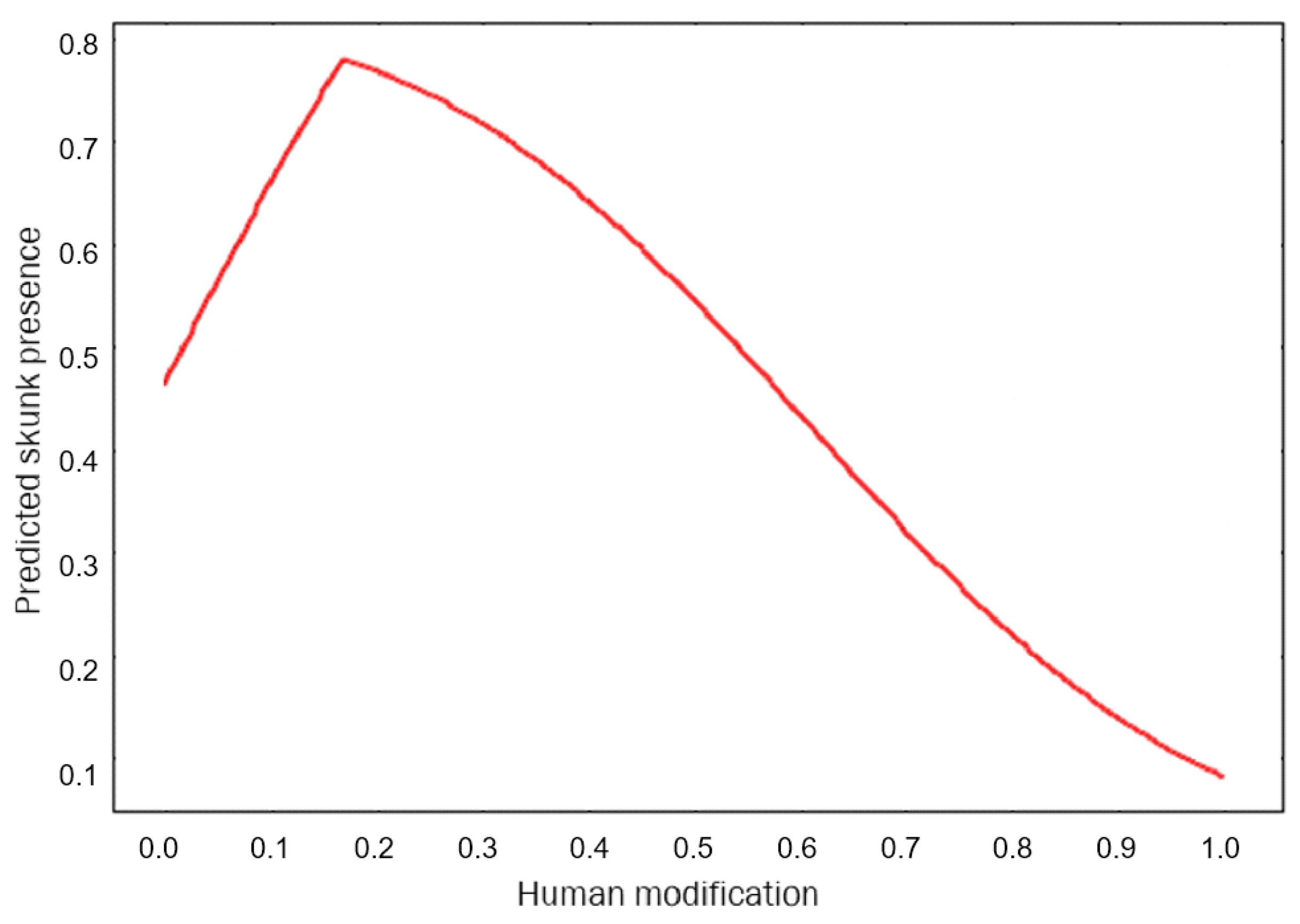
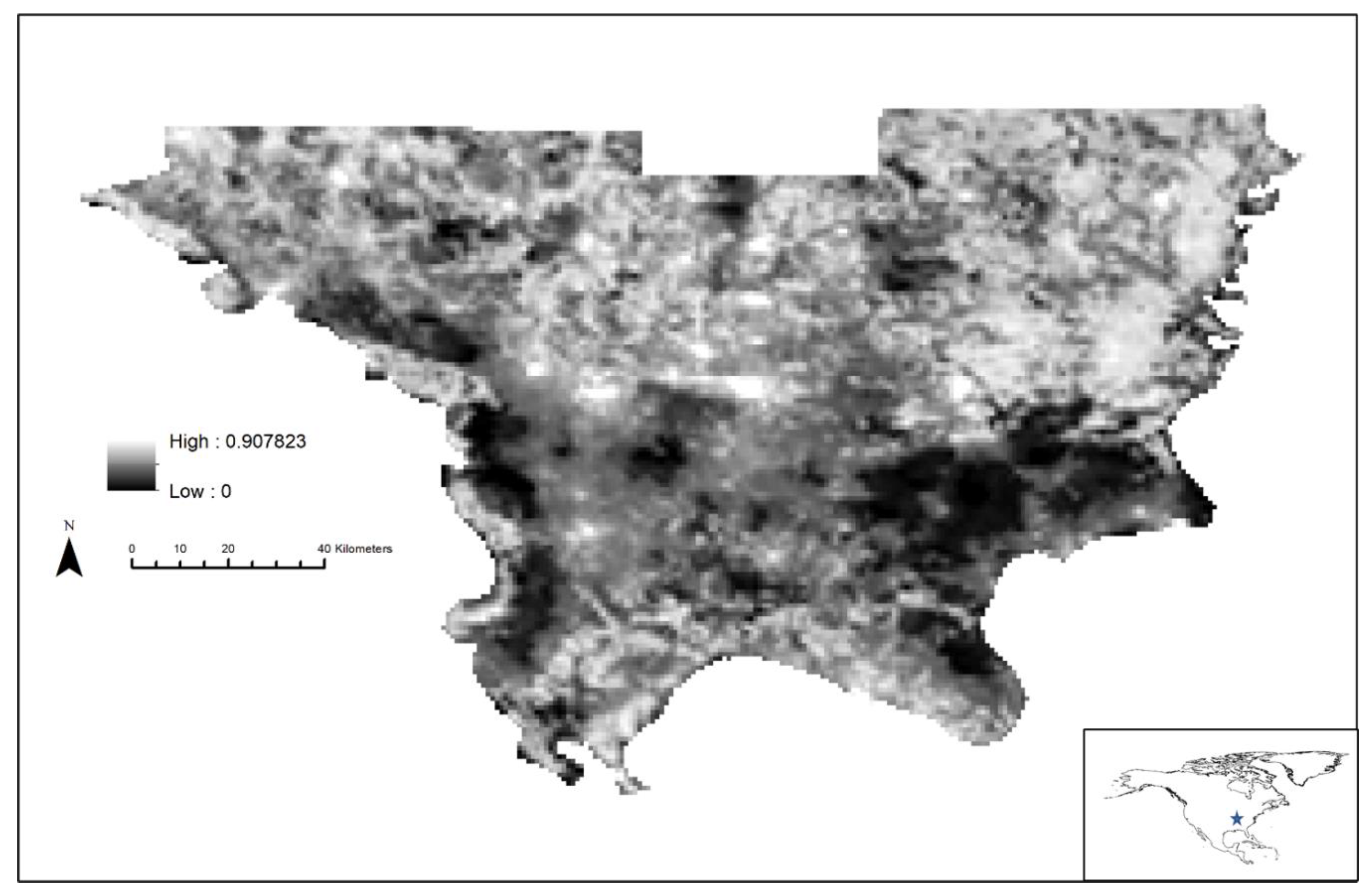
3. Results
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Ritchie, E.G.; Johnson, C.N. Predator interactions, mesopredator release and biodiversity conservation. Ecol. Lett. 2009, 12, 982–998. [Google Scholar] [CrossRef] [PubMed]
- Berger, J.; Stacey, P.B.; Bellis, L.; Johnson, M.P. A mammalian predator-prey imbalance: Grizzly bear and wolf extinction affect avian neotropical migrants. Ecol. Appl. 2001, 11, 947–960. [Google Scholar] [CrossRef]
- Laliberte, A.S.; Ripple, W.J. Range contractions of North American carnivores and ungulates. Bioscience 2004, 54, 123–138. [Google Scholar] [CrossRef] [Green Version]
- Roemer, G.W.; Gompper, M.E.; Van Valkenburgh, B. The ecological role of the mammalian mesocarnivore. Bioscience 2009, 59, 165–173. [Google Scholar] [CrossRef]
- Litvaitis, J.A.; Villafuerte, R. Predation, mesopredator release, and prey stability. Conserv. Biol. 1996, 10, 676–677. [Google Scholar] [CrossRef]
- Oehler, J.D.; Litvaitis, J.A. The role of spatial scale in understanding responses of medium-sized carnivores to forest fragmentation. Can. J. Zool. 1996, 74, 2070–2079. [Google Scholar] [CrossRef]
- Houle, M.; Fortin, D.; Mainguy, J.; Canac-Marquis, P. Landscape composition and structure influence the abundance of mesopredators: Implications for the control of the raccoon (Procyon lotor) variant of rabies. Can. J. Zool. 2011, 89, 1107–1116. [Google Scholar] [CrossRef]
- Terraube, J.; Arroyo, B. Factors influencing diet variation in a generalist predator across its range distribution. Biodivers. Conserv. 2011, 20, 2111–2131. [Google Scholar] [CrossRef] [Green Version]
- Sanz-Aguilar, A.; Jovani, R.; Melián, C.J.; Pradel, R.; Tella, J.L. Multi-event capture-recapture analysis reveals individual foraging specialization in a generalist species. Ecology 2015, 96, 1650–1660. [Google Scholar] [CrossRef] [Green Version]
- Nielsen, C.K.; Woolf, A. Survival of unexploited bobcats in southern Illinois. J. Wildl. Manag. 2002, 66, 833–838. [Google Scholar] [CrossRef]
- Wilson, S.E.; Nielsen, C.K. Habitat characteristics of raccoon daytime resting sites in southern Illinois. Am. Midl. Nat. 2007, 157, 175–186. [Google Scholar] [CrossRef]
- Hanrahan, A.T.; Rutter, A.U.; Nielsen, C.K.; Schauber, E.M. Spatial ecology of river otters in a human-modified landscape. J. Mammal. 2019, 100, 1327–1339. [Google Scholar] [CrossRef]
- Beasley, J.C.; Olson, Z.H.; Dharmarajan, G.; Eagan, T.S.; Rhodes, O.E. Spatio-temporal variation in the demographic attributes of a generalist mesopredator. Landsc. Ecol. 2011, 26, 937–950. [Google Scholar] [CrossRef]
- Moore, J.E.; Swihart, R.K. Modeling patch occupancy by forest rodents: Incorporating detectability and spatial autocorrelation with hierarchically structured data. J. Wildl. Manag. 2005, 69, 933–949. [Google Scholar] [CrossRef]
- Homer, C.; Dewitz, J.; Jin, S.; Xian, G.; Costello, C.; Danielson, P.; Gass, L.; Funk, M.; Wickham, J.; Stehman, S.; et al. Conterminous United States land cover change patterns 2001–2016 from the 2016 National Land Cover Database. ISPRS J. Photogramm. Remote Sens. 2020, 162, 184–199. [Google Scholar] [CrossRef]
- Lesmeister, D.B.; Nielsen, C.K.; Schauber, E.M.; Hellgren, E.C. Spatial and temporal structure of a mesocarnivore guild in midwestern North America. Wildl. Monogr. 2015, 191, 1–61. [Google Scholar] [CrossRef] [Green Version]
- Beatty, W.S.; Beasley, J.C.; Rhodes, O.E. Habitat selection by a generalist mesopredator near its historical range boundary. Can. J. Zool. 2014, 92, 41–48. [Google Scholar] [CrossRef]
- Verts, B.J. The Biology of the Striped Skunk; University of Illinois Press: Urbana, IL, USA, 1967; p. 218. [Google Scholar]
- Bailey, T.N. Biology of striped skunks on a southwestern Lake Erie marsh. Am. Midl. Nat. 1971, 85, 196–207. [Google Scholar] [CrossRef]
- Storm, G.L. Daytime retreats and movements of skunks on farmlands in Illinois. J. Wildl. Manag. 1972, 36, 31–45. [Google Scholar] [CrossRef]
- Greenwood, R.J.; Sargeant, A.B.; Johnson, D.H. Evaluation of mark-recapture for estimating striped skunk abundance. J. Wildl. Manag. 1985, 49, 332–340. [Google Scholar] [CrossRef] [Green Version]
- Rosatte, R.C.; Power, M.J.; MacInnes, C.D. Ecology of urban skunks, raccoons, and foxes in metropolitan Toronto. In Wildlife Conservation in Metropolitan Environments; Adams, L.W., Leedy, D.L., Eds.; National Institute for Urban Wildlife: Columbia, MD, USA, 1991; pp. 31–38. ISBN 09-420-15037. [Google Scholar]
- Broadfoot, J.D.; Rosatte, R.C.; O’Leary, D.T. Raccoon and skunk population models for urban disease control planning in Ontario, Canada. Ecol. Appl. 2001, 11, 295–303. [Google Scholar] [CrossRef]
- Bixler, A.; Gittleman, J.L. Variation in home range and use of habitat in the striped skunk (Mephitis mephitis). J. Zool. Lond. 2000, 251, 525–533. [Google Scholar] [CrossRef]
- McKinney, M.L. Urbanization, biodiversity, and conservation. Bioscience 2002, 52, 883–890. [Google Scholar] [CrossRef]
- Prange, S.; Gehrt, S.D. Changes in mesopredator-community structure in response to urbanization. Can. J. Zool. 2004, 82, 1804–1817. [Google Scholar] [CrossRef]
- Gehrt, S.D. Ecology and management of striped skunks, raccoons, and coyotes in urban landscapes. In People and Predators: From Conflict to Coexistence; Fascione, N., Delach, A., Smith, M.E., Eds.; Island Press: Washington, DC, USA, 2004; pp. 81–104. ISBN 1-55963-084-1. [Google Scholar]
- Brown, L.G.; Yeager, L.E. Survey of the Illinois fur resource. Illinois Nat. Hist. Surv. Bull. 1943, 22, 435–504. [Google Scholar] [CrossRef]
- Mohr, C.O. Illinois furbearer distribution and income. Illinois Nat. Hist. Surv. Bull. 1943, 22, 505–537. [Google Scholar] [CrossRef]
- Greenspan, E.; Nielsen, C.K.; Cassel, K.W. Potential distribution of coyotes (Canis latrans), Virginia opossums (Didelphis virginiana), striped skunks (Mephitis mephitis), and raccoons (Procyon lotor) in the Chicago Metropolitan Area. Urban Ecosyst. 2018, 16, 983–997. [Google Scholar] [CrossRef]
- Hiestand, S.J.; Nielsen, C.K.; Jiménez, F.A. Modeling potential occurrence of metazoan endoparasites of bobcats (Lynx rufus) using verified records. Folia Parasitol. 2014, 61, 401–410. [Google Scholar] [CrossRef] [Green Version]
- Phillips, S.J.; Anderson, R.P.; Schapire, R.E. Maximum entropy modeling of species geographic distributions. Ecol. Model. 2006, 190, 231–259. [Google Scholar] [CrossRef] [Green Version]
- Peterson, A.T.; Soberón, J.; Pearson, R.G.; Anderson, R.P.; Martínez-Meyer, E.; Nakamura, M.; Araujó, M.B. Ecological Niches and Geographic Distributions: Monographs in Population Biology, 49; Princeton University Press: Princeton, NJ, USA, 2011; p. 328. [Google Scholar]
- Schwegman, J.E. Comprehensive Plan for the Illinois Nature Preserves System, Part 2: The Natural Divisions of Illinois; Illinois Nature Preserves Commission: Rockford, IL, USA, 1973; p. 32.
- County Population by Characteristics: 2010–2018. Available online: https://www.census.gov/data/datasets/time-series/demo/popest/2010s-counties-detail.html?# (accessed on 25 June 2020).
- Streams and Shorelines for Illinois. Available online: https://clearinghouse.isgs.illinois.edu/data/hydrology/streams-and-shorelines (accessed on 25 June 2020).
- Illinois Highway and Street Mileage Statistics. Available online: http://www.idot.illinois.gov/Assets/uploads/files/Transportation-System/Reports/OP&P/Travel-Stats/Illinois%20Highway%20and%20Street%20Mileage%20Statistics%202013.pdf (accessed on 25 June 2020).
- Global Summary of the Year (GSOY), Version 1. Available online: https://www.ncdc.noaa.gov/cdo-web/ (accessed on 25 June 2020).
- Wilting, A.; Cord, A.; Hearn, A.J.; Hesse, D.; Mohamed, A.; Traeholdt, C.; Cheyne, S.M.; Sunarto, S.; Jayasilan, M.A.; Ross, J.; et al. Modelling the species distribution of flat-headed cats (Prionailurus planiceps), an endangered South-East Asian small felid. PLoS ONE 2010, 5, e9612. [Google Scholar] [CrossRef] [Green Version]
- Wang, Y.; Casajus, N.; Buddle, C.; Berteaux, D.; Larrivée, M. Predicting the distribution of poorly-documented species, Northern black widow (Latrodectus variolus) and Black purse-web spider (Sphodros Niger), using museum specimens and citizen science data. PLoS ONE 2018, 13, e0201094. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kennedy, C.M.; Oakleaf, J.R.; Theobald, D.M.; Baruch-Mordo, S.; Kiesecker, J. Managing the middle: A shift in conservation priorities based on the global human modification gradient. Glob. Chang. Biol. 2019, 25, 811–826. [Google Scholar] [CrossRef] [PubMed]
- Brown, J.L.; Bennett, J.R.; French, C.M. SDMtoolbox 2.0: The next generation python-based GIS toolkit for landscape genetic, biogeographic and species distribution model analyses. PeerJ 2017, 5, 1–12. [Google Scholar] [CrossRef] [Green Version]
- Phillips, S.J.; Dudík, M. Modeling of species distributions with Maxent: New extensions and a comprehensive evaluation. Ecography 2008, 31, 161–175. [Google Scholar] [CrossRef]
- Anderson, R.P.; Gonzalez, J.I. Species-specific tuning increases robustness to sampling bias in models of species distributions: An implementation with MaxEnt. Ecol. Model. 2011, 222, 2796–2811. [Google Scholar] [CrossRef]
- Wade-Smith, J.; Verts, B.J. Mephitis mephitis. Mamm. Spec. 1982, 173, 1–7. [Google Scholar] [CrossRef]
- Rosatte, R.; Kelly, P.; Power, M. Home range, movements, and habitat utilization of striped skunk (Mephitis mephitis) in Scarborough, Ontario, Canada: Disease management implications. Can. Field Nat. 2011, 125, 27–33. [Google Scholar] [CrossRef] [Green Version]
- Theimer, T.C.; Clayton, A.C.; Martinez, A.; Peterson, D.L.; Bergman, D.L. Visitation rate and behavior of urban mesocarnivores differs in the presence of two common anthropogenic food sources. Urban Ecosyst. 2015, 18, 895–906. [Google Scholar] [CrossRef] [Green Version]
- Iverson, L.R. Land-use changes in Illinois, USA: The influence of landscape attributes on current and historic land use. Landsc. Ecol. 1988, 2, 45–61. [Google Scholar] [CrossRef]
- Bennitt, R.; Nagel, W.O. A survey of the resident game and furbearers of Missouri. Univ. Missouri Stud. 1937, 12, 1–215. [Google Scholar]
- Houseknecht, C.R. Denning habits of the striped skunk and the exposure potential for disease. Bull. Wildl. Dis. Assoc. 1969, 5, 302–306. [Google Scholar] [CrossRef]




Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Amspacher, K.; Jiménez, F.A.; Nielsen, C. Influence of Habitat on Presence of Striped Skunks in Midwestern North America. Diversity 2021, 13, 83. https://doi.org/10.3390/d13020083
Amspacher K, Jiménez FA, Nielsen C. Influence of Habitat on Presence of Striped Skunks in Midwestern North America. Diversity. 2021; 13(2):83. https://doi.org/10.3390/d13020083
Chicago/Turabian StyleAmspacher, Katelyn, F. Agustín Jiménez, and Clayton Nielsen. 2021. "Influence of Habitat on Presence of Striped Skunks in Midwestern North America" Diversity 13, no. 2: 83. https://doi.org/10.3390/d13020083
APA StyleAmspacher, K., Jiménez, F. A., & Nielsen, C. (2021). Influence of Habitat on Presence of Striped Skunks in Midwestern North America. Diversity, 13(2), 83. https://doi.org/10.3390/d13020083