
Ojo Guareña: A Hotspot of Subterranean Biodiversity in Spain



Abstract
:1. Introduction
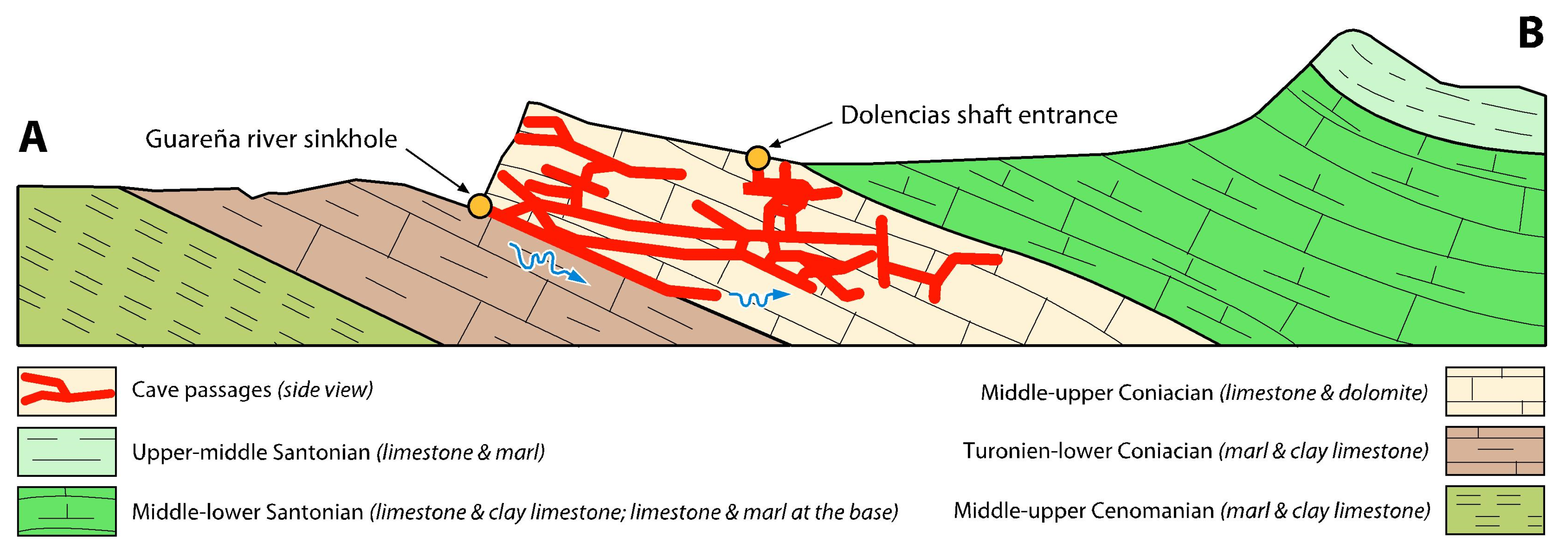
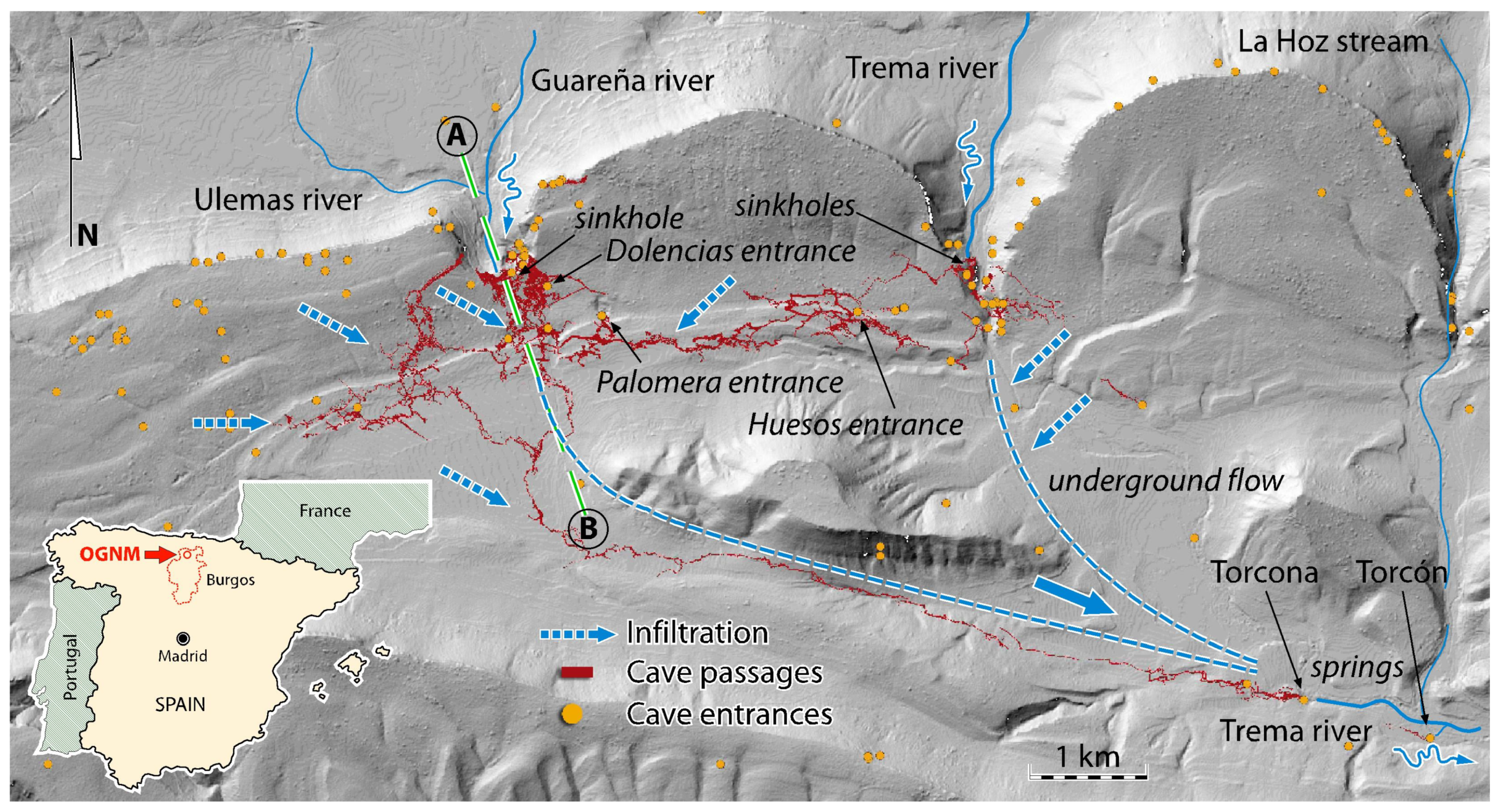
2. A Bit of History
3. Materials and Methods
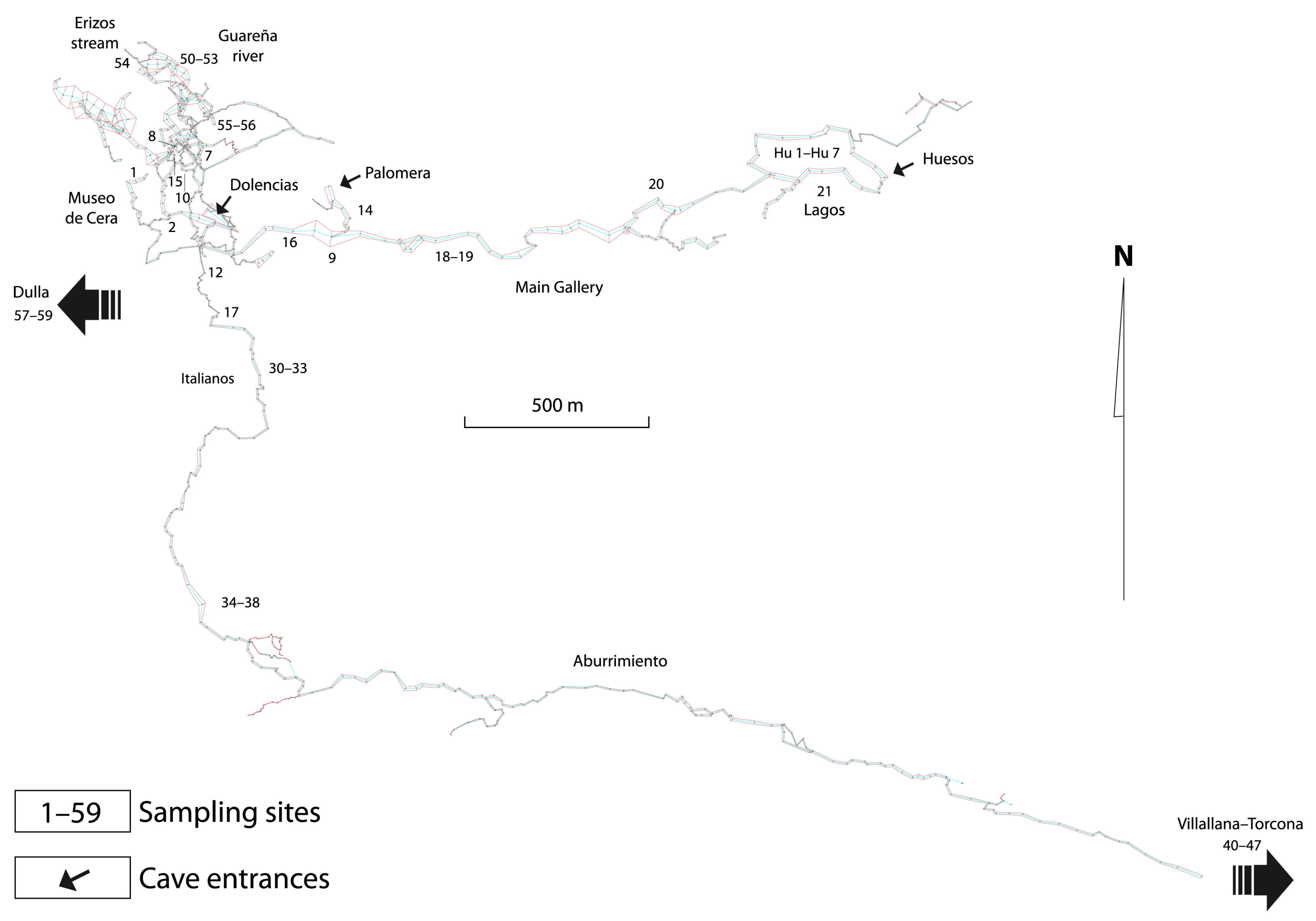
3.1. Sampling and Studies of the Fauna
3.1.1. Samples of Aquatic Fauna
3.1.2. Samples of Terrestrial Fauna
4. Results
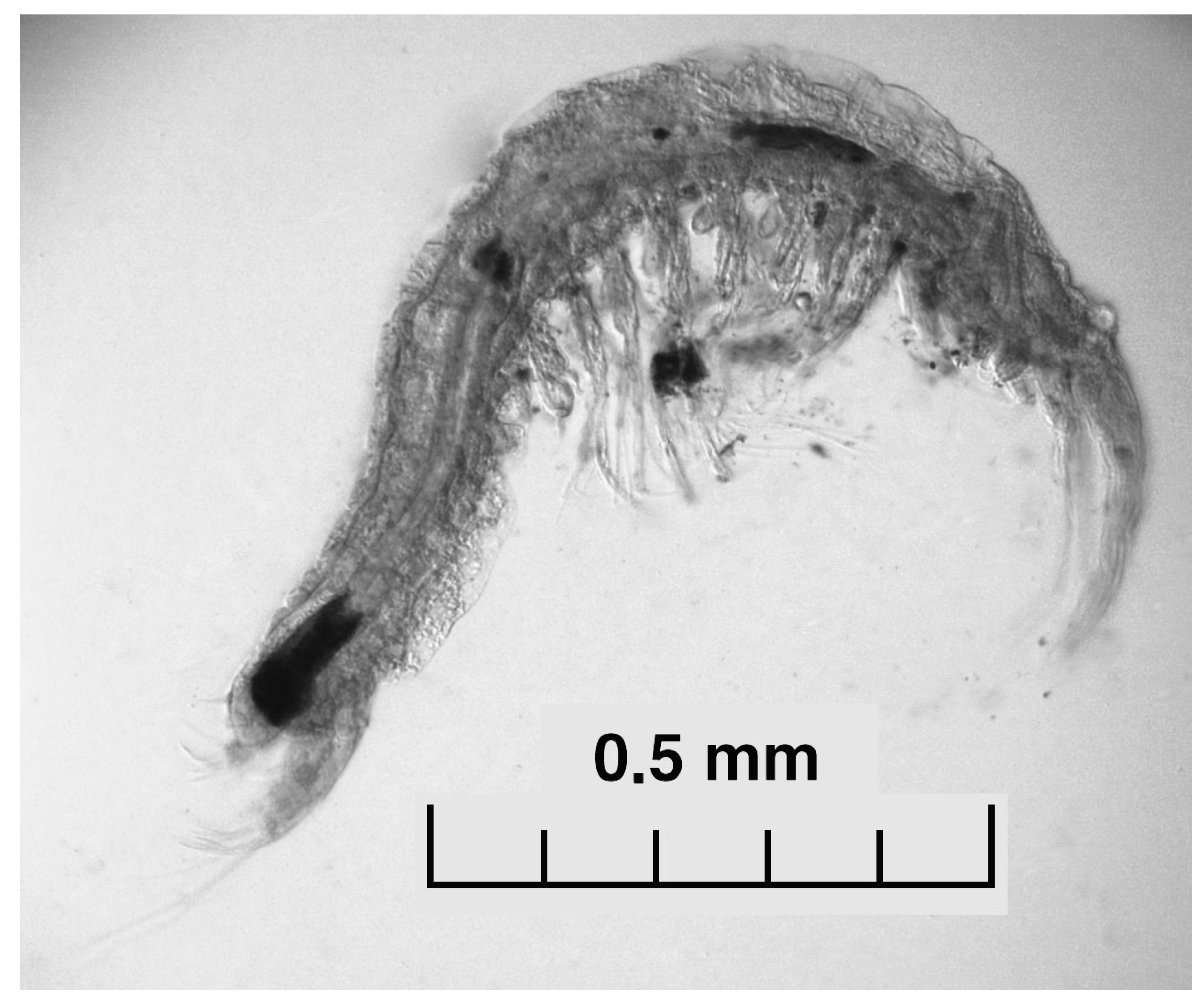
4.1. Aquatic Fauna
- Oligochaeta
- Gianius navarroi Rodriguez and Achurra, 2010. OG10 and OG14.
- Delaya navarrensis (Delay, 1973). OG21.
- Stylodrilus mariae Achurra, Rodriguez and Erséus, 2015. OG07 and OG08.
- Trichodrilus tenuis Habre, 1960. OG14.
- Trichodrilus sp. 1. OG09.
- Mollusca
- Palaospeum septentrionale (Rolan and Ramos, 1995). Sima Jaime Cave, De la Hoz Creek, and Torcona Spring.
- Spiralix (Burgosia) burgensis Boeters, 2003. Sima Jaime Cave, Trema River, and Trueba River.
- Hydrachnidia
- Axonopsis (Paraxonopsis) vietsi Motas and Tanasachi, 1947. Trueba River.
- Frontipodopsis reticulatifrons Szalay, 1945. OG08, OG10, and Engaña River.
- Frontipodopsis sp. Lasía River.
- Kongsbergia sp. Trueba and Lasía rivers.
- Neoacarus hibernicus Halbert, 1944. Nela River.
- Protzia sp. Trueba River.
- Pseudotorrenticola sp. Nela River.
- Sperchonopsis sp. Lasía River.
- Limnohalacarida (Halacaridae)
- Soldanellonyx chappuisi Walter, 1917. OG: 07–10, 12, 14, 15, 17, 19, 30, 31, 34, 36–38, and 50; caves: Rizuelos, García, San Miguel, Cornejo and Redonda; springs: Torcona and Calzada; rivers: Trema, Trueba, Engaña, Nela, and De la Hoz and Torcon Well.
- Copepoda, Cyclopoida
- Acanthocyclops cf. venustus (Norman and Scott, 1906). Engaña and Nela rivers.
- Acanthocyclops n. sp. OG: 10, 31, 34, 37, and 38.
- Graeteriella (G.) unisetigera (Graeter, 1910). OG02 and OG07.
- Speocyclops infernus (Kiefer, 1930). OG: 01, 08, 09, 14, 16–19, 21, 31, 38, and Huesos 2 and 4.
- Speocyclops sebastianus Kiefer, 1937. Trema River.
- Speocyclops spelaeus Kiefer, 1937. Caves: García, San Bernabé, Las Llanas, and Racino.
- Copepoda, Harpacticoida
- Bryocamptus (Rheocamptus) pyrenaicus (Chappuis, 1923). Caves: Kaite 2, García, Cornejo, Racino, and Redonda; Salce Spring.
- Paracamptus sp. SA. Nela River.
- Parastenocaris sp. OG: 07–09, 12, 14, 16, 17, 20, 21, 31–38, 41–43, 45, 46, 51–54, and Huesos 2; Villa Spring and De la Hoz and De la Cueva creeks.
- Ostracoda
- Candoninae cf. gen. sp. 1 “Rounded”. OG: 07, 09, 12, 17, 19, 21, 39; spring: Torcona, Cornejo and Calzada; Nela River.
- Candoninae “Trapezoid” 1. OG: 09, 19, 21; caves: Jaime, La Mina, Las Llanas; Guareña River.
- Candoninae cf. gen. sp. 2 “Trapezoid” 2. OG09.
- Candoninae cf. gen. sp. 2 “Trapezoid” 3. OG: 1, 18, 37, 38.
- Candoninae “Triangular” 1. OG: 15, 30–32 and Trema River.
- Candoninae “Triangular” 2. OG16 and Sima Jaime Cave.
- Candoninae “Triangular” 3. Torcona Spring and Trema River.
- Cypria sp. OG: 7, 9, 10, 35, 40, 42, 43, 51, 55–59, Huesos 2 and 3; San Miguel Cave; springs: Cornejo, Villa, Salce, Cubio, and Jordana; rivers: Guareña and Lasía.
- Amphipoda
- Haploginglymus sp. 1. OG38; Torcona Spring; river: Trema, Trueba, and Nela; Torcon well.
- Niphargus sp. OG37 and 38; Cubio Spring and Trueba River.
- Pseudoniphargus burgensis Notenboom, 1986. Torcona Spring.
- Pseudoniphargus n. sp. 1. Torcona Spring.
- Isopoda
- Cantabroniscus primitivus Vandel, 1965. OG21.
- Stenasellus virei buchneri (Stammer, 1936). OG: 02, 07, 09, 10, 14, 15, 17, 21, 39, 40, 44, 53, 55, and 57; caves: García, San Bernabé, San Miguel, Las Llanas, and Redonda and Salce Spring.
- Bathynellacea
- Iberobathynella burgalensis Camacho, 2005. OG53.
- Iberobathynella cornejoensis Camacho, 2005. Redonda and Racino caves and Trema River.
- Iberobathynella guarenensis Camacho, 2003. OG57 (“Erizos River”).
- Vejdovskybathynella edelweiss Camacho, 2007. OG01, 09, 16, 17, 38, 50, 57, and Huesos 3 and 4; Racino and Mina caves and Cubio Spring
- Vejdovskybathynella n. sp. 1 (cryptic species “edelweiss”). OG57 (“Erizos River”).
- Vejdovskybathynella n. sp. 2 (cryptic species “edelweiss”). Redonda and Sima Jaime caves.
- Bathynellidae n. gen. n. sp. OG09, 14, Torcona Spring and Prado Vargas Cave.
4.2. Terrestrial Fauna
- Oligochaeta:
- Aporrectodea rosea troglodyta (Álvarez, 1971).
- Orodrilus paradoxoides (Álvarez, 1971).
- Mollusca:
- Zospeum schaufussi Frauenfeld, 1862.
- Arachnida, Opiliones:
- Litocampa zaldivarae Sendra, Salgado and Monedero, 2003.
- Myriapoda, Chilopoda:
- Lithobius deroutae sexusbispiniger Demange and Serra, 1978.
- Collembola:
- Verhoeffiella cf hispanicus (Bonet, 1931).
- Coleoptera:
- Speocharis sharpi Escalera, 1898.
- Trechus ortizi Español, 1970.
5. Discussion and Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Puch, C. La Montaña subterránea. Pyrenaica 2009, 235, 331–333. [Google Scholar]
- Culver, D.; Pipan, T. Subterranean Ecosystems. In Encyclopedia of Biodiversity, 2nd ed.; Levin, S.A., Ed.; Academic Press: Waltham, MA, USA, 2013; Volume 7, pp. 49–62. [Google Scholar] [CrossRef]
- Martínez, A.; González, B.C. Volcanic Anchialine Habitats of Lanzarote. In Cave Ecology; Ecological Studies 235; Moldovan, O.T., Kovác, L., Halse, S., Eds.; Springer: Cham, Switzerland, 2018; pp. 195–228. [Google Scholar]
- Puch, C. Grandes Cuevas y Simas de España; Espelo Club de Grácia-Federación Española de Espeleología: Barcelona, Spain, 1998; p. 816. [Google Scholar]
- Culver, D.C.; Deharveng, L.; Bedos, A.; Lewis, J.J.; Madden, M.; Reddell, J.R.; Sket, B.; Trontelj, P.; White, D. The mid-latitude biodiversity ridge in terrestrial cave fauna. Ecography 2006, 29, 120–128. [Google Scholar] [CrossRef]
- Gulden, B. Official list of the National Speleological Society (USA). Available online: http://www.caverbob.com/wlong.htm (accessed on 2 February 2021).
- Camacho, A.I.; Temiño, C.; Cabeza, B.; Puch, C. El Monumento Natural de Ojo Guareña (Burgos, España): Un “hotspot” de biodiversidad acuática subterránea. In Cuevas: Patrimonio, Naturaleza, Cultura y Turismo; Durán, J.J., Carrasco, F., Eds.; Asociación de Cuevas Turísticas Españolas: Madrid, Spain, 2010; pp. 621–636, ISBN 13:978-84-614-4630-8. [Google Scholar]
- Wilson, E.O. Biodiversity research requires more boots on the ground. Nat. Ecol. Evol. 2017, 1, 1590–1591. [Google Scholar] [CrossRef]
- Ficetola, G.F.; Canedoli, C.; Stoch, F. The Racovitzan impediment and the hidden biodiversity of unexplored environments. Conser. Biol. 2018, 33, 214–216. [Google Scholar] [CrossRef]
- Camacho, A.I.; Dorda, B.A.; Rey, I. Undisclosed taxonomic diversity of Bathynellacea (Malacostraca: Syncarida) in the Iberian Peninsula revealed by molecular data. J. Crust. Biol. 2012, 32, 816–826. [Google Scholar] [CrossRef] [Green Version]
- Deharveng, L.; Stoch, F.; Gibert, J.; Bedos, A.; Galassi, D.; Zagmajster, M.; Brancelj, A.; Camacho, A.; Fiers, F.; Martin, P.; et al. Groundwater biodiversity in Europe. Freshw. Biol. 2009, 54, 709–726. [Google Scholar] [CrossRef]
- Camacho, A.I. Four new species of groundwater crustaceans (Syncarida, Bathynellacea, Parabathynellidae) endemic to the Iberian Peninsula. J. Nat. Hist. 2003, 37, 2885–2907. [Google Scholar] [CrossRef]
- Camacho, A.I. One more piece in the genus puzzle: A new species of Iberobathynella Schminke, 1973 (Syncarida, Bathynellacea, Parabathynellidae) from the Iberian Peninsula. Graellsia 2005, 61, 123–133. [Google Scholar] [CrossRef] [Green Version]
- Camacho, A.I. The first record of the genus Vejdovskybathynella Serban and Leclerc, 1984 (Syncarida, Bathynellacea, Bathynellidae) in the Iberian Peninsula: Three new species. J. Nat. Hist. 2007, 41, 2817–2841. [Google Scholar] [CrossRef]
- Camacho, A.I.; Torres, T.; Puch, C.J.; Ortiz, J.E.; Valdecasas, A.G. Small-scale biogeographical pattern in groundwater Crustacea (Syncarida, Parabathynellidae). Biodivers. Conserv. 2006, 15, 3527–3541. [Google Scholar] [CrossRef]
- Ortega, A.I.; Ruiz, F.; Martín, M.A.; Benito, A.; Vidal, M.; Bermejo, L.; Karampaglidis, T. Prehistoric human tracks in Ojo Guareña Cave System (Burgos, Spain): The Sala and Galerías de las Huellas. In Reading Prehistoric Human Tracks; Pastoor, A., Lenssen-Erz, T., Eds.; Springer: Berlin/Heidelberg, Germany, 2021; pp. 1–21. ISBN 10-3030604055; ISBN 13-978-3030604059. [Google Scholar]
- Sáenz, C. Notas acerca de la estratigrafía del Supracretáceo y del Numulítico en la cabecera del Nela y zonas próximas. Bol. R. Soc. Esp. Hist. Nat. 1933, XXXIII, 159–185. [Google Scholar]
- GEE (Grupo Espeleológico Edelweiss). Monografía sobre Ojo Guareña. Kaite 1986, 4–6, 1–415. [Google Scholar]
- Bellés, X. Ptinidos recogidos en cavidades subterráneas ibéricas (Col. Ptinidae). Speleon 1976, 22, 145–147. [Google Scholar]
- Bellés, X. Notas sobre Speocharis minus Jeannel, 1909 y otros catópidos recogidos en cuevas de la provincia de Burgos. Graellsia 1977, 31, 115–124. [Google Scholar]
- Bellés, X. Nuevos datos sobre la Fauna de Ojo Guareña. Ixiltasun Izkutuak, 1986; Unpublished. [Google Scholar]
- Bellés, X. Fauna Cavernícola i Intersticial de la Península Ibérica i les Illes Balears; Monografies Cientifiques, 4; CSIC: Mallorca, Spain, 1987; p. 207. [Google Scholar]
- Salgado, J.M. Origenes e distribuçao geográfica des Bathysciinae (Col. Catop.) cantábricos (Grupo Speocharis). Ciénc. Biol. 1976, 1, 105–130. [Google Scholar]
- Salgado, J.M. Nuevos datos sobre Entomofauna Cavernícola de la zona de Carranza (Vizcaya). Kobie 1977, 7, 127–138. [Google Scholar]
- Vives, E. Coleópteros cavernícolas nuevos o interesantes de la Península Ibérica y Baleares. Speleon 1976, 22, 159–169. [Google Scholar]
- Vives, M. Noves localitats de Trichoniscidae cavernicolas de la fauna espanyola (Crustacís: Isopodes: Oniscoides). Com. VI. Simp. Espeleol. Biospeleol. 1977, 97–101. [Google Scholar]
- Notenboom, J.; Meijers, I. Investigaciones sobre la fauna de las aguas subterráneas de España: Lista de estaciones y primeros resultados. In Verslagen en Technische Gegevens; Instituut voor Taxonomische Zoölogie (Zoologische Museum), Universiteit von Amsterdam: Amsterdam, The Netherlands, 1985; Volume 42, pp. 1–43. [Google Scholar]
- Notenboom, J. Introduction to the Iberian Groundwater amphipods. Limnetica 1990, 6, 165–176. [Google Scholar]
- Camacho, A.I. Informe Biológico: Ojo Guareña (Sotoscueva-Burgos), 1993; Unpublished. 29.
- Camacho, A.I.; Valdecasas, A.G.; Rodríguez, J.; Puch, C. Biodiversidad Faunística del Complejo Kárstico de Ojo Guareña: Evaluación de la Influencia de la Presión Humana en Algunas de Las Poblaciones de Invertebrados en el Monumento Natural de Ojo Guareña (Burgos), Informe Final, Marzo. 2005; Unpublished. 110.
- Camacho, A.I.; Puch, C. Colonización, éxito Evolutivo y Biodiversidad Faunística del Complejo Kárstico de Ojo Guareña en el Monumento Natural de Ojo Guareña (Burgos), Informe Final. 2010; Unpublished. 240.
- Olmo, P.d.; Ramírez del Pozo, J. Mapa Geológico 1:50.000 n° 84 (Espinosa de los Monteros); IGME: Madrid, Spain, 1978. [Google Scholar]
- Camacho, A.I. Sampling the subterranean biota. Cave (aquatic environment). In The Natural History of Biospeleology; Camacho, A.I., Ed.; Monografías del Museo Nacional de Ciencias Naturales, 7; CSIC: Madrid, Spain, 1992; pp. 135–168. ISBN 84-00-07280-4. [Google Scholar]
- Malard, F.; Dole-Olivier, M.J.; Mathieu, J.; Stoch, F.; Brancelj, A.; Camacho, A.I.; Fiers, F.; Galassi, D.; Gibert, J.; Lefebure, T.; et al. Sampling Manual for the Assessment of Regional Groundwater Biodiversity. PASCALIS Project (V Framework Programme. Key Action 2: Global Change, Climate and Biodiversity. 2.2.3 Assessing and Conserving Biodiversity); Malard, F., Ed.; Pascalis: Villeurbanne, France, Contract n°: N°EVK2-CT-2001-00121; 2002; p. 110. Available online: http//www.pascalis-project.org (accessed on 31 March 2021).
- Camacho, A.I.; Dorda, B.A.; Rey, I. Identifying cryptic speciation across groundwater populations: First COI sequences of Bathynellidae (Crustacea, Syncarida). Graellsia 2011, 67, 7–12. [Google Scholar] [CrossRef] [Green Version]
- Camacho, A.I.; Dorda, B.A.; Rey, I. Old and new taxonomic tools: Description of a new genus and two new species of Bathynellidae from Spain with morphological and molecular characters. J. Nat. Hist. 2013, 47, 1393–1420. [Google Scholar] [CrossRef]
- Deharveng, L.; Bedos, A. Diversity of terrestrial invertebrates in subterranean habitats. In Cave Ecology; Moldovan, O.T., Kovác, L., Halse, S., Eds.; Springer: Cham, Switzerland, 2018; pp. 107–172. [Google Scholar]
- Rodriguez, P.; Achurra, A. New species of aquatic oligochaetes (Annelida: Clitellata) from groundwaters in karstic areas of northern Spain, with taxonomic remarks on Lophochaeta ignota Stolc, 1886. Zootaxa 2010, 2331, 21–29. [Google Scholar] [CrossRef]
- Camacho, A.I. Expanding the taxonomic conundrum: Three new species of groundwater crustaceans (Syncarida, Bathynellacea, Parabathynellidae) endemic to the Iberian Peninsula. J. Nat. Hist. 2005, 39, 1819–1838. [Google Scholar] [CrossRef]
- Camacho, A.I.; Dorda, B.A.; Rey, I. Integrating DNA and morphological taxonomy to describe a new species of the family Bathynellidae (Crustacea, Syncarida) from Spain. Graellsia 2013, 69, 179–200. [Google Scholar] [CrossRef] [Green Version]
- Álvarez, J. Biospeleología de la cueva Ojo Guareña. Oligoquetos terrícolas. Bol. R. Soc. Esp. Hist. Nat. (Biol.) 1971, 69, 11–18. [Google Scholar]
- Pérez-Iñigo, C. Biospeleología de la cueva Ojo Guareña. Acaros oribátidos. Bol. R. Soc. Esp. Hist. Nat. (Biol.) 1969, 67, 143–160. [Google Scholar]
- Prieto, C.E.; Gómez, B.J. Primeros datos de Zospeum (Mollusca, Gastropoda, Ellobiidae) para la provincia de Burgos. Com. 2º Simp. Reg. Espeleol. Burgos 1984, 143–147. [Google Scholar]
- Demange, J.M.; Serra, A. Etude des rapports de longueur des articles des P.15 de quelques Lithobius cavernicoles de l’Espagne et des Pyrénées françaises. Description d’une espèce et une sous-espèce nouvelles (Chilopoda, Lithobiomorpha). Speleon 1978, 24, 39–54. [Google Scholar]
- Rambla, M. Contribución al estudio de los Opiliones de la Fauna Ibérica. Las especies del grupo nemastoma bacilliferum, Simon, 1879 en la Península Ibérica (Opiliones, Fam. Nemastomatidae). Publ. Inst. Biol. Aplic. 1968, 45, 33–56. [Google Scholar]
- Prieto, C.E.; Zubiaga, A. El género Ischyropsalis C.L. Koch (Ischyropsalididae, Opiliones) en la provincia de Burgos. Mem. 2º Simp. Reg. Espeleol. Burgos 1984, 15–19. [Google Scholar]
- Español, F. Un nuevo Trechus cavernícola del norte de Burgos (Col. Trechidae). Speleon 1970, XVII, 53–57. [Google Scholar]
- Mösslacher, F. Evolution, Adaption und Verbreitung. In Grundwasseroökologie; Griebler, C., Mösslacher, F., Eds.; UTB-Facultas Verlag: Wien, Austria, 2003; pp. 253–310. [Google Scholar]
- Culver, D.C.; Christman, M.C.; Sket, B.; Trontelj, P.P. Sampling adequacy in an extreme environment: Species richness patterns in Slovenian caves. Biodivers. Conserv. 2004, 13, 1209–1229. [Google Scholar] [CrossRef]
- Bregovic, P.; Zagmajster, M. Understanding hotspots within a global hotspot—Identifying the drivers of regional species richness patterns in terrestrial subterranean habitats. Insect Conserv. Biodivers. 2016, 9, 268–281. [Google Scholar] [CrossRef]
- Gibert, J.; Deharveng, L. Subterranean ecosystems: A truncated functional biodiversity. Bioscience 2002, 52, 473–481. [Google Scholar] [CrossRef]
- Griebler, C.; Mösslacher, F. Grundwasser—eine ökosystemare Betrachtung. In Grundwasserökologie; Griebler, C., Mösslacher, F., Eds.; UTB-Facultas Verlag: Wien, Austria, 2003; pp. 253–310. [Google Scholar]
- Hahn, H.J. The GW-Fauna-Index: A first approach to a quantitative ecological assessment of groundwater habitats. Limnologica 2006, 36, 119–137. [Google Scholar] [CrossRef] [Green Version]
- Stoch, F.; Galassi, D.M.P. Stygobiotic crustacean species richness: A question of numbers, a matter of scale. Hydrobiologia 2010, 653, 217–234. [Google Scholar] [CrossRef]
- Camacho, A.I.; Mas-Peinado, P.; Dorda, B.A.; Casado, A.; Brancelj, A.; Knight, L.R.F.D.; Hutchins, B.; Bou, C.; Perina, G.; Rey, I. Molecular tools unveil an underestimated diversity in a stygofauna family: A preliminary world phylogeny and updated morphology of Bathynellidae (Crustacea: Bathynellacea). Zool. J. Linn. Soc. 2018, 183, 70–96. [Google Scholar] [CrossRef]
- Zagmajster, M.; Malard, F.; Eme, D.; Culver, D.C. Subterranean biodiversity patterns from global to regional scales. In Cave Ecology, Ecological Studies 235; Moldovan, O.T., Kovác, L., Halse, S., Eds.; Springer: Cham, Switzerland, 2018; pp. 195–228. [Google Scholar]
- Deharveng, L.; Bedos, A. The cave fauna of southeast Asia. Origin, evolution, and ecology. In Subterranean Ecosystems; Wilken, H., Culver, D.C., Humphreys, W.F., Eds.; Elsevier: Amsterdam, The Netherlands, 2000; pp. 603–632. [Google Scholar]
- Pipan, T.; Deharveng, L.; Culver, D.C. Hotspots of Subterranean Biodiversity. Diversity 2020, 12, 209. [Google Scholar] [CrossRef]
- Camacho, A.I. Diversity, morphological homogeneity and genetic divergence in a taxonomically complex group of groundwater crustaceans: The little known Bathynellacea (Malacostraca). Bull. Soc. d’Hist. Nat. Toulouse 2019, 154, 105–160. [Google Scholar]








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
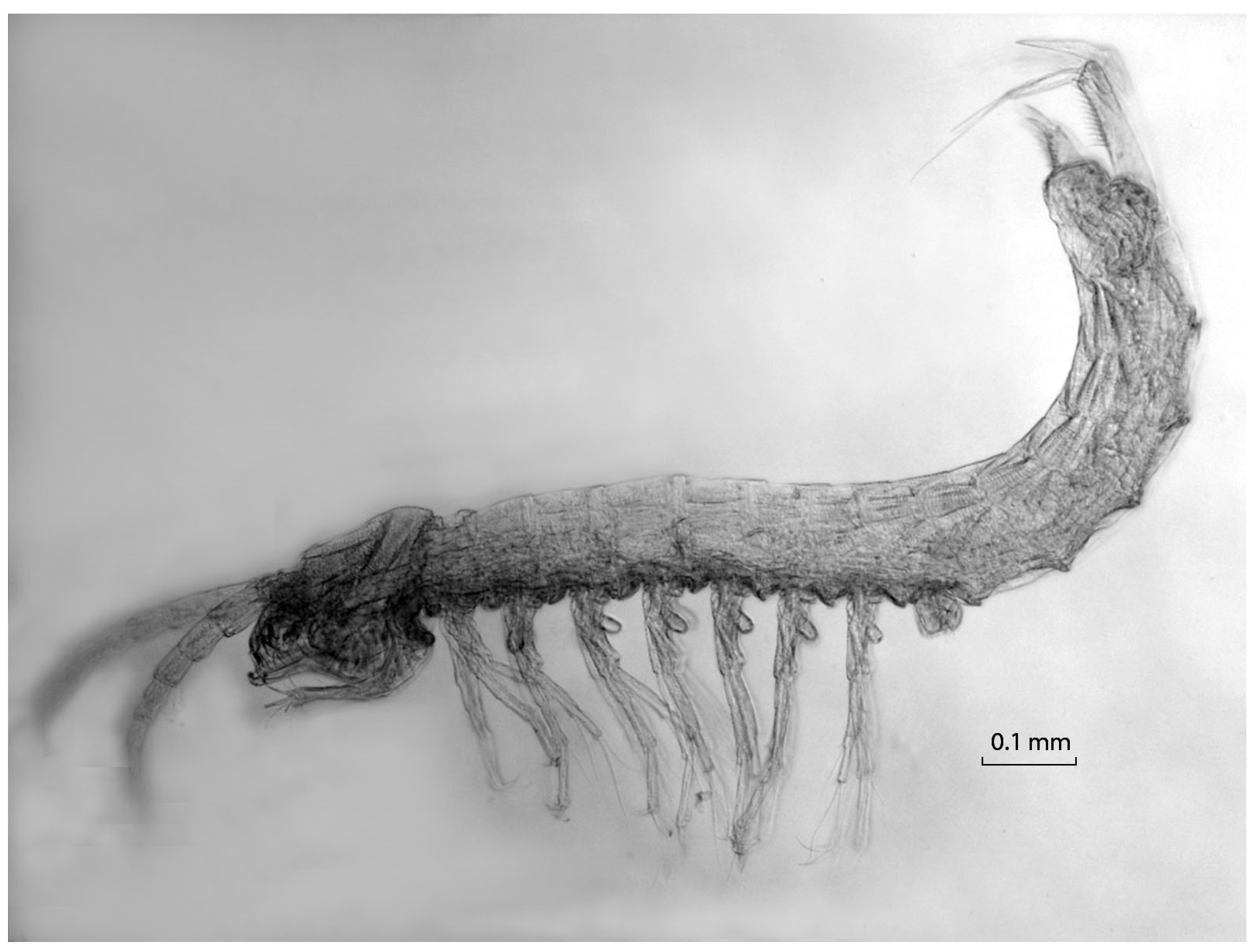{kind=link}
| TAXA | Total Species | Stygobiotic | Troglobiotic | Stygophilic | Troglophilic |
|---|---|---|---|---|---|
| Aquatic/Terrestrial | Species | Species | Species | Species | |
| Rotifera | 1/--- | 0 | --- | 1 | --- |
| Cnidaria | 1/--- | 0 | --- | 0 | --- |
| Nematoda | 1/? | --- | 0 | --- | 0 |
| Turbellaria | 5/--- | 0 | --- | 0 | --- |
| Hirudinea | 3/--- | 0 | --- | 1 | --- |
| Tardigrada | 13/3 | 0 | 0 | 0 | 0 |
| Oligochaeta | 49/9 | 5 | 2 | 8 | 0 |
| Mollusca | |||||
| Gastropoda | 11/4 | 2 | 1 | 1 | 0 |
| Bivalvia | 5/0 | 0 | --- | 0 | --- |
| Crustacea | |||||
| Cladocera | 1/0 | 0 | --- | 0 | --- |
| Ostracoda | 22/0 | 8 | --- | 9 | --- |
| Copepoda | 37/0 | 9 | --- | 12 | --- |
| Amphipoda | 8/0 | 4 | --- | 0 | --- |
| Isopoda | 3/1 | 2 | 0 | 1 | 1 |
| Bathynellacea | 7/0 | 7 | --- | --- | --- |
| Limnohalacarida | 1/--- | 1 | --- | 0 | --- |
| Hydrachnidia | 19/--- | 8 | --- | 1 | --- |
| Oribatida | ---/54 | --- | 0 | --- | 5 |
| Araneae | ---/1 | --- | 0 | --- | 1 |
| Opiliones | ---/2 | --- | 0 | --- | 1 |
| Pseudoscorpionida | ---/1 | --- | 0 | --- | 1 |
| Myriapoda | ---/3 | --- | 1 Chilopoda | --- | 2 Diplopoda |
| Collembola | ---/1 | --- | 1 | --- | 0 |
| Diplura | ---/2 | --- | 1 | --- | 0 |
| Zygentoma | ---/1 | --- | 0 | --- | 0 |
| Coleoptera | ---/13 | --- | 2 | --- | 3 |
| Diptera | ---/12 (Families) | --- | 0 | --- | 0 |
| Hemiptera | ---/1 (Order level) | --- | 0 | --- | 0 |
| Lepidoptera | ---/1 (Order level) | --- | 0 | --- | 0 |
| Orthoptera | ---/1 (Order level) | --- | 0 | --- | 0 |
| Psocoptera | ---/1 (Order level) | --- | 0 | --- | 0 |
| Psiphonaptera | ---/1 (Order level) | --- | 0 | --- | 0 |
| TOTAL: 299 | 187/112 | 46 | 8 | 34 | 14 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Camacho, A.I.; Puch, C. Ojo Guareña: A Hotspot of Subterranean Biodiversity in Spain. Diversity 2021, 13, 199. https://doi.org/10.3390/d13050199
Camacho AI, Puch C. Ojo Guareña: A Hotspot of Subterranean Biodiversity in Spain. Diversity. 2021; 13(5):199. https://doi.org/10.3390/d13050199
Chicago/Turabian StyleCamacho, Ana Isabel, and Carlos Puch. 2021. "Ojo Guareña: A Hotspot of Subterranean Biodiversity in Spain" Diversity 13, no. 5: 199. https://doi.org/10.3390/d13050199
APA StyleCamacho, A. I., & Puch, C. (2021). Ojo Guareña: A Hotspot of Subterranean Biodiversity in Spain. Diversity, 13(5), 199. https://doi.org/10.3390/d13050199