1. Introduction
The order Psittaciformes contains some of the most charismatic and recognizable bird species in the world [
1]. However, of the order’s approximately 352 species, 26% face some degree of extinction risk [
2]. For example, out of the 22 Psittacidae species recorded in Mexico [
1,
3], 20 are at risk according to Mexican law [
4], and at the international level, the Red List of the International Union for the Conservation of Nature (IUCN) places eight of those species in some risk category [
5]. The Military Macaw (
Ara militaris) is one of the endangered psittacid species in Mexico, and faces two main threats: (1) habitat transformation (loss, fragmentation and degradation) [
6,
7], and (2) illegal collection for the national and international illegal pet trade [
7,
8,
9,
10,
11,
12,
13,
14,
15]. Indeed, illegal trafficking has led to the extirpation of populations from conserved areas [
11,
12,
16].
In Mexico, the illegal wildlife trade has threatened 19 out of the 22 Psittacidae species [
7]. The capture of any wild Psittacidae species was outlawed in Mexico in 2003, and the current General Wildlife Law (LGVS) prohibits the extraction of psittacid species, only granting permits for conservation or scientific research purposes [
17]. One of the objectives of the LGVS was to guide management efforts, including the recovery, reproduction, research, release, and/or relocation of individuals [
18]. One of the problems faced by reintroduction and recovery efforts is that in most cases, the geographic origin of animals recovered from the illegal pet trade is unknown. Information on the geographic origin of rehabilitated individuals is crucial in order to avoid mixing individuals from genetically distinct populations, which can lead to genetic problems (e.g., local maladaptation and outbreeding depression) [
19,
20,
21,
22].
Molecular tools make it possible to answer questions concerning evolutionary history, define taxonomic uncertainties, and identify release locations using molecular markers (e.g., microsatellites) and statistical approaches [
23,
24,
25,
26]. However, these techniques are not often used for the identification of release locations for rehabilitated birds illegally taken from the wild [
21,
27]. The use of molecular tools to establish the origin of individuals for conservation purposes is increasing in reintroduction plans and for identifying illegal trade sites, as demonstrated by studies of several species, such as the Hyacinth Macaw (
Anodorhynchus hyacinthinus) [
28], the Blue-and-Yellow Macaw (
Ara ararauna) [
21] and the European Pond Turtle (
Emys orbicularis) [
29].
The purpose of this study was to determine the probable geographic origin of a captive Military Macaw of unknown origin using different molecular statistical analyses and test the accuracy of these techniques using individuals of known origin, in order to generate a protocol that can be used for reintroduction programs, for management and conservation.
The Military Macaw is one of the most charismatic species in the New World. Its distribution is fragmented, ranging from northern Mexico to northwestern Argentina [
1,
6,
30]. In Mexico, the Military Macaw is distributed in apparently isolated colonies in two separate areas. One includes the Sierra Madre Occidental and the Sierra Madre del Sur (from southern Sonora to Chiapas); the other is in the Sierra Madre Oriental, where the macaws are reported in Tamaulipas, San Luis Potosi, Guanajuato, and Querétaro [
31,
32]. The geographic distribution of the Military Macaw in Mexico declined by 43% over 16 years (2000–2016) [
10,
13,
14]. It is endangered under Mexican law [
4,
13], vulnerable on the IUCN Red List [
5], and listed in Appendix I of the Convention on International Trade in Species [
33].
A study conducted by Rivera-Ortíz et al. [
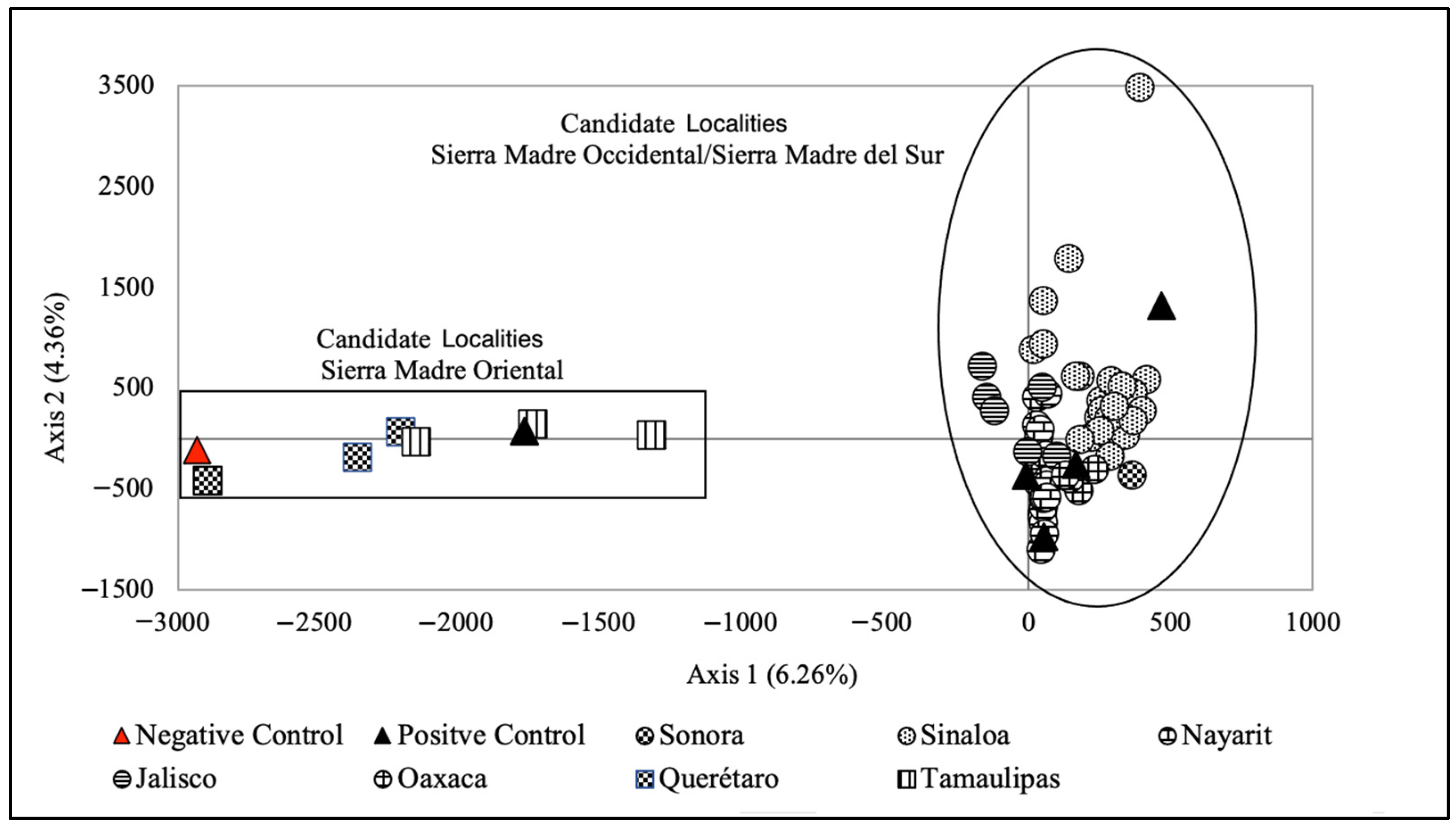
34] on the genetics of the Military Macaw used microsatellites from samples collected in seven Mexican locations and found strong genetic structuring, showing two groups presenting geographic concordance. Group one corresponded to the locations found in the Sierra Madre Occidental and the Sierra Madre del Sur (Pacific slope), and Group two corresponded to the locations found in the Sierra Madre Oriental (Gulf of Mexico slope). Given these results, the authors proposed the protection of the two genetic groups found in the three physiographic regions as independent conservation units. In this study, we used both classification (correspondence analysis) and genetic assignment methods to evaluate whether the Military Macaw individual of unknown origin belonged to any of those previously identified genetic groups, and if possible, assign it to a particular location.
4. Discussion
Identification of an individual’s geographic origin by means of genetic analysis depends on the ability to assign it to a particular location, which in turn depends on the level of genetic structuring among reference locations [
58]. Here, we tested the ability of molecular genetic assignment to identify the likely location of origin of one individual of the Military Macaw of unknown origin and five individuals of known origin, in order to evaluate the method’s utility in future conservation efforts for the species. In this study, the results showed that the methods tested were useful in identifying the geographic areas from which individuals likely originated, for both the unknown individual and the five positive controls.
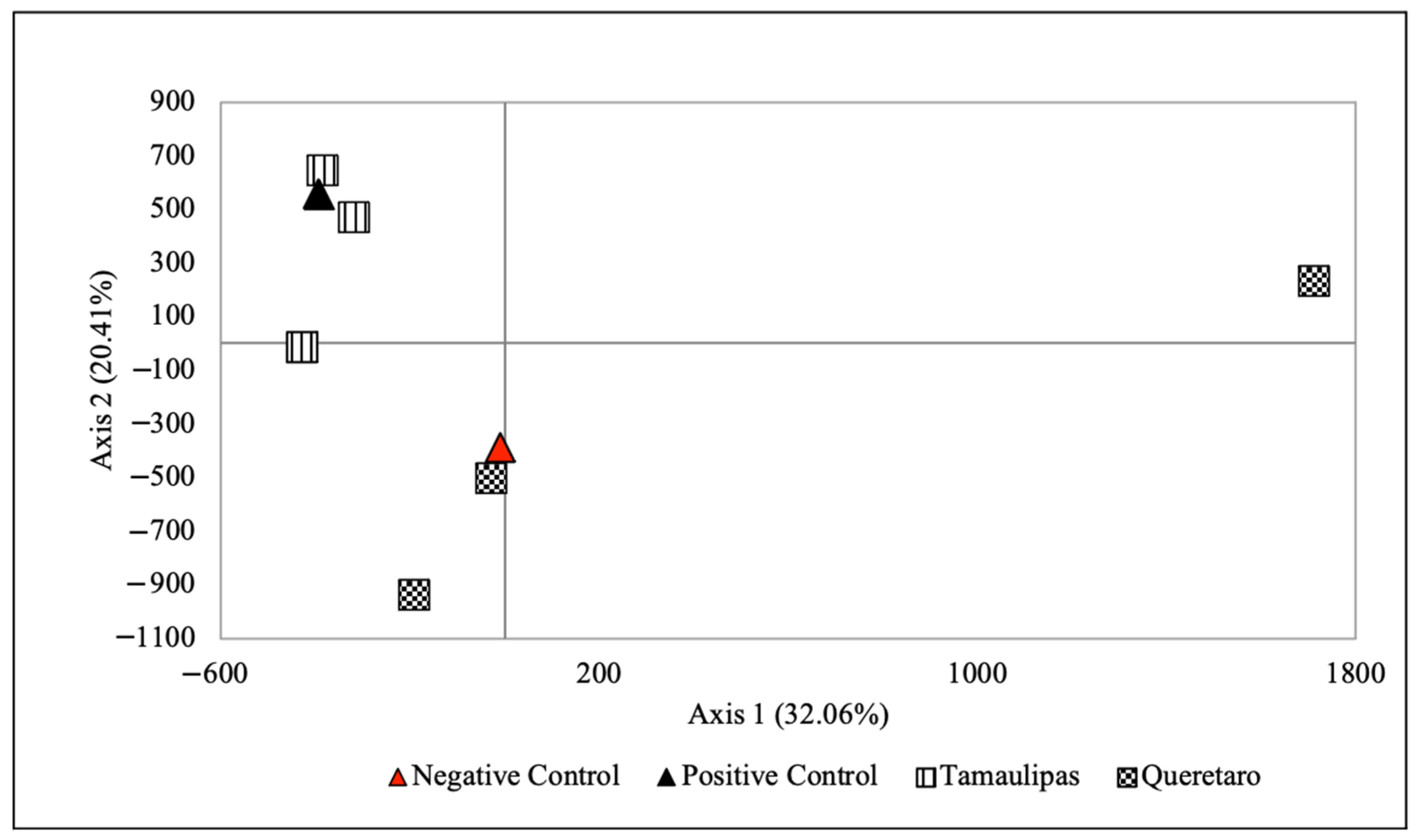
The results of the FCA, STRUCTURE and DAPC tests grouped the unknown individual with the Sierra Madre Oriental genetic group with high confidence. However, it was impossible to assign it to a specific geographic location because there is no differentiation between individuals from different reference locations in this genetic group, indicating gene flow. These FCA, STRUCTURE and DAPC results are reliable because the reference sample of individuals used in the study and provided by Rivera-Ortíz et al. [
34] presents a marked genetic structure and differentiation across the distribution range of the Military Macaw in Mexico, showing a pattern that was also found by Eberhard et al. [
59] with mitochondrial markers. These previously documented patterns of genetic structure are important in the context of the present study because structure and differentiation among the reference locations must be high if there is to be reasonable success in geographic allocation using grouping methods (with 80–100% correct allocation) [
26,
60,
61].
The allocation/exclusion analyses carried out using GENCLASS suggest that the likely origin of the unknown individual is the Sierra Madre Oriental metapopulation, as determined by the grouping analyses. Three of the four criteria used for the allocation/exclusion analyses show some probability that the unknown individual belongs to the Querétaro location, although with relatively low certainty (30–46% probability). These low probability values should be interpreted with caution, since they may be affected by small sample sizes in some of the reference locations. Some authors suggest that a sample of 30–50 individuals per reference location is necessary to allow accurate estimates [
26,
55,
58]. Unfortunately, obtaining large sample sizes in studies of endangered species is extremely difficult due to small population sizes, restricted areas, and difficulty accessing their distributional areas [
13,
62,
63], as in the case of the Military Macaw.
The different methods used to identify the probable location of origin of the individual Military Macaw of unknown origin proved to be effective and complementary, as demonstrated in this study. When carrying out this type of analysis, we recommend graphically showing the genetic similarity of the individuals as a first step that reveals if the samples of unknown origin are grouped in the reference localities [
64]. Then, consider a Bayesian approach to determine the probability that the individuals of unknown origin originate from a population, considering all reference localities together [
26]. Finally, use the tests to exclude or identify individuals of unknown origin in the reference localities, to determine the probability that individuals of unknown origin are rejected or belong to the reference localities [
65].
Our study shows that given the degree of population genetic structure in Military Macaw locations in Mexico, it is possible to use microsatellite data to identify the probable location of origin of an individual of unknown provenance. This, in turn, makes it possible to make a more informed selection of locations at which the individual could be released. The captive specimen was geographically assigned to the Sierra Madre Oriental, and according to our results, is a candidate for release in that zone. It is essential that the programs for reintroducing and releasing Military Macaw individuals into the wild make proper use of this kind of molecular tools [
42,
66,
67], given that for an endangered species, such as the Military Macaw, the strong genetic structuring of wild locations may reflect local adaptations that would be lost if they were to be managed as a single group [
34].
To improve the accuracy of assigning individuals of unknown origin to their correct populations, it is crucial to continue genetic studies of wild locations and increase the number of molecular markers used in genetic analyses. Relatively low numbers of microsatellites were used in this study, but microsatellites have provided sufficient power for geographic assignment of a variety of wild species due to their high level of polymorphism and genetic structure of the populations [
68,
69,
70]. The use of other markers such as mitochondrial DNA would be very informative and complementary since it might allow us to distinguish lineages that correspond to particular geographic areas [
21]. Identification of single nucleotide polymorphisms (SNPs) from genomic data also have a significant advantage for geographic assignment, since information from hundreds or thousands of SNPs could potentially provide improved resolution of patterns of genetic structure, and thus, the more precise assignment of an individual’s geographic origin [
61].
Our study demonstrates that in combination with the reference samples analyzed by Rivera-Ortiz et al. [
34], currently available molecular markers and statistical assignment and exclusion software can help identify the geographic origin of captive individuals or specimens confiscated from illegal trade [
50]. No studies have been conducted to analyze the number of Military Macaw individuals trapped each year, but Cantú et al. [
16] estimated that 65,000 to 78,000 psittacid individuals are poached for illegal trade and suffer a mortality rate of 77%. Only about 2% of poached individuals are seized by the Mexican Federal Environmental Protection Agency (PROFEPA) [
16], but given how many are poached, this small percentage still represents several hundred individuals. In this sense, identifying the geographic origin of captive individuals or specimens confiscated from illegal trade helps biodiversity managers to detect locations with intense poaching, and thus, focus efforts and resources on these sites to prevent poaching. It will also support and guide restoration or demographic translocation programs if they are deemed necessary to increase genetic variability [
23,
28].
A crucial component of this study was the availability of the set of reference samples of known geographic origin [
34]. We recommend the establishment of large DNA reference collections and large public databases containing allele frequencies from many populations, and the use of museum collections, which can play an essential role since DNA can be extracted from museum skins. Any genetic analysis that attempts to identify geographic origin of an individual/sample depends on having good data on georeferenced genetic variation. These databases would be extremely valuable in efforts to conserve endangered species [
26], by helping to detect and reduce illegal trade and informing conservation management plans.


 ,
, 



{kind=link}
{kind=link}
{kind=link}
{kind=link}