
How Can We Record Reliable Information on Animal Colouration in the Wild?




Abstract
:1. Introduction
2. Materials and Methods
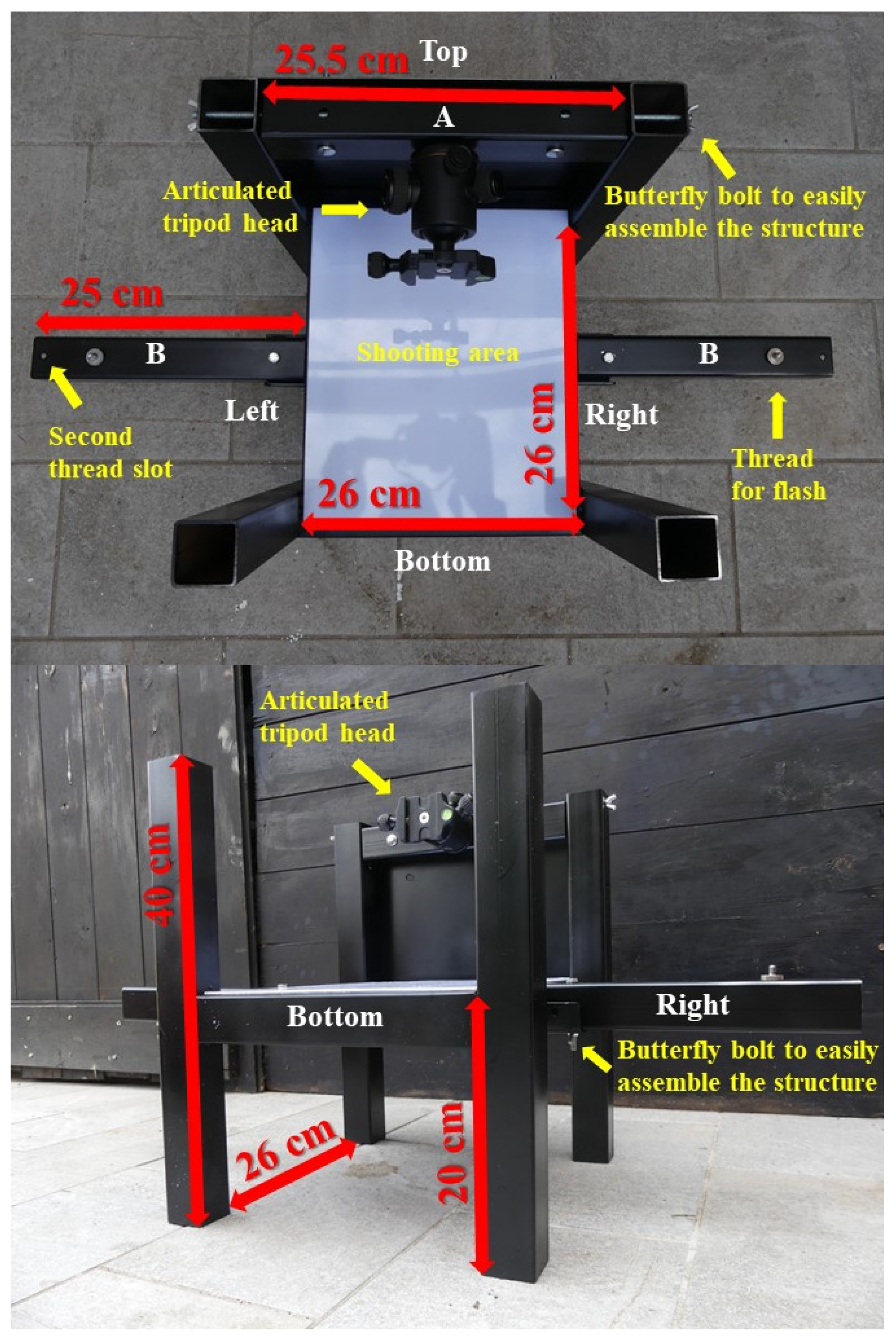
2.1. Description of the Structure
2.2. Testing the Reliability of the Method
3. Results
4. Discussion
Author Contributions
Funding
Institutional Review Board Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Laman, T.; Scholes, E. Birds of Paradise: Revealing the World’s Most Extraordinary Birds; Nationall Geographic Society: Washington, DC, USA, 2012; p. 227. [Google Scholar]
- Fenolio, D. LIfe in the Dark. Illuminating Biodiversity in the Shadowy Haunts of Planet Earth; Johns Hopkins University Press: Baltimore, MD, USA, 2016; p. 317. [Google Scholar]
- Cuthill, I.C.; Allen, W.L.; Arbuckle, K.; Caspers, B.; Chaplin, G.; Hauber, M.E.; Hill, G.E.; Jablonski, N.G.; Jiggins, C.D.; Kelber, A.; et al. The biology of color. Science 2017, 357, eaan0221. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Najbar, A.; Konowalik, A.; Najbar, B.; Ogielska, M. Yellow patterns polymorphism of the fire salamander Salamandra salamandra in Poland. Acta Herpetol. 2018, 13, 101–108. [Google Scholar] [CrossRef]
- Lunghi, E.; Giachello, S.; Zhao, Y.; Corti, C.; Ficetola, G.F.; Manenti, R. Photographic database of the European cave salamanders, genus Hydromantes. Sci. Data 2020, 7, 171. [Google Scholar] [CrossRef]
- Bereiter-Hahn, J.; Matoltsy, A.G.; Sylvia Richards, K. (Eds.) Biology of the Integument. 2 Vertebrales; Springer: Berlin, Germany, 1986. [Google Scholar]
- Gerl, E.J.; Morris, M.R. The causes and consequences of color vision. Evol. Educ. Outreach 2008, 1, 476–486. [Google Scholar] [CrossRef] [Green Version]
- Losey, G.S.J. Crypsis and communication functions of UV-visible coloration in two coral reef damselfish, Dascyllus aruanus and D. reticulatus. Anim. Behav. 2003, 66, 299–307. [Google Scholar] [CrossRef] [Green Version]
- Giurfa, M.; Lehrer, M. Honeybee vision and floral displays: From detection to close-up recognition. In Cognitive Ecology of Pollination. Animal Behavior and Floral Evolution; Chittka, L., Thomson, J.D., Eds.; Cambridge University Press: Cambridge, UK, 2004; pp. 61–82. [Google Scholar]
- Kodric-Brown, A. Female choice of multiple male criteria in guppies: Interacting effects of dominance, coloration and courtship. Behav. Ecol. Sociobiol. 1993, 32, 415–420. [Google Scholar] [CrossRef]
- de Lanuza, G.P.I.; Font, E.; Carretero, M.A. Colour assortative pairing in a colour polymorphic lizard is independent of population morph diversity. Sci. Nat. 2016, 103, 82. [Google Scholar] [CrossRef] [PubMed]
- Santos, J.C.; Coloma, L.A.; Cannatella, D.C. Multiple, recurring origins of aposematism and diet specialization in poison frogs. Proc. Natl. Acad. Sci. USA 2003, 100, 12792–12797. [Google Scholar] [CrossRef] [Green Version]
- Ruxton, G.D.; Sherratt, T.N.; Speed, M.P. Avoiding Attack. The Evolutionary Ecology of Crypsis, Warning Signals, and Mimicry; Oxford University Press: Oxford, UK, 2004; p. 249. [Google Scholar]
- Cott, H.B. Adaptive Coloration in Animals; Methuen & Co LTD: London, UK, 1940; p. 508. [Google Scholar]
- Lanza, B.; Caputo, V.; Nascetti, G.; Bullini, L. Morphologic and genetic studies of the European plethodontid salamanders: Taxonomic inferences (genus Hydromantes). Monogr. Del. Mus. Reg. Sci. Nat. Torino 1995, 16, 1–366. [Google Scholar]
- Spinner, M.; Kovalev, A.; Gorb, S.N.; Westhoff, G. Snake velvet black: Hierarchical microand nanostructure enhances dark colouration in Bitis rhinoceros. Sci. Rep. 2013, 3, 1846. [Google Scholar] [CrossRef] [Green Version]
- Lunghi, E.; Giachello, S.; Mulargia, M.; Dore, P.P.; Cogoni, R.; Corti, C. Variability in the dorsal pattern of the Sardinian grass snake (Natrix natrix cetti) with notes on its ecology. Acta Herpetol. 2019, 14, 141–145. [Google Scholar] [CrossRef]
- Berggren, Å.; Merilä, J. WWW design code–A new tool for colour estimation in animal studies. Front. Zool. 2004, 1, 2. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bergman, T.J.; Beehner, J.C. A simple method for measuring colour in wild animals: Validation and use on chest patch colour in geladas (Theropithecus gelada). Biol. J. Linn. Soc. 2008, 94, 231–240. [Google Scholar] [CrossRef] [Green Version]
- Thibaudeau, G.; Altig, R. Coloration of anuran tadpoles (Amphibia): Development, dynamics, function, and hypotheses. ISRN Zool. 2012, 2012, 725203. [Google Scholar] [CrossRef] [Green Version]
- Bierbach, D.; Penshorn, M.; Hamfler, S.; Herbert, D.B.; Appel, J.; Meyer, P.; Slattery, P.; Charaf, S.; Wolf, R.; Völker, J.; et al. Gradient evolution of body colouration in surfaceand cave-dwelling Poecilia mexicana and the role of phenotype-assortative female mate choice. BioMed Res. Int. 2013, 2013, 148348. [Google Scholar] [CrossRef] [PubMed]
- Brenes-Soto, A.; Dierenfeld, E.E.; Janssens, G.P.J. Colouration in amphibians as a reflection of nutritional status: The case of tree frogs in Costa Rica. PLoS ONE 2017, 12, e0182020. [Google Scholar] [CrossRef] [Green Version]
- Potash, A.D.; Greene, D.U.; Foursa, G.A.; Mathis, V.L.; Conner, L.M.; Mccleery, R.A. A comparison of animal color measurements using a commercially available digital color sensor and photograph analysis. Curr. Zool. 2020, 66, 601–606. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Stevens, M.; Párraga, C.A.; Cuthill, I.C.; Partridge, J.C.; Troscianko, T.S. Using digital photography to study animal coloration. Biol. J. Linn. Soc. 2007, 90, 211–237. [Google Scholar] [CrossRef] [Green Version]
- Lunghi, E.; Cianferoni, F.; Giachello, S.; Zhao, Y.; Manenti, R.; Corti, C.; Ficetola, G.F. Updating salamander datasets with phenotypic and stomach content information for two mainland Speleomantes. Sci. Data 2021, 8, 150. [Google Scholar] [CrossRef]
- Troscianko, J.; Stevens, M. Image calibration and analysis toolbox–A free software suite for objectivelymeasuring reflectance, colour and pattern. Methods Ecol. Evol. 2015, 6, 1320–1331. [Google Scholar] [CrossRef] [Green Version]
- Cooper, V.J.; Hosey, G.R. Sexual dichromatism and female preference in Eulemur fulvus subspecies. Int. J. Primatol. 2003, 24, 1177–1188. [Google Scholar] [CrossRef]
- Weller, H.I.; Westneat, M.W. Quantitative color profiling of digital images with earth mover’s distance using the R package colordistance. PeerJ 2019, 7, e6398. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Martel, A.; Blooi, M.; Adriaensen, C.; Van Rooij, P.; Beukema, W.; Fisher, M.C.; Farrer, R.A.; Schmidt, B.R.; Tobler, U.; Goka, K.; et al. Recent introduction of a chytrid fungus endangers Western Palearctic salamanders. Science 2014, 346, 630. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lunghi, E.; Manenti, R.; Canciani, G.; Scarì, G.; Pennati, R.; Ficetola, G.F. Thermal equilibrium and temperature differences among body regions in European plethodontid salamanders. J. Biol. 2016, 60, 79–85. [Google Scholar] [CrossRef] [PubMed]
- Luzuriaga, D.A.; Balaban, M.O.; Yeralan, S. Analysis of visual quality attributes of white shrimp by machine vision. J. Food Sci. 1997, 62, 113–118. [Google Scholar] [CrossRef]
- Flemons, P.; Berents, P. Image based Digitisation of Entomology Collections: Leveraging volunteers to increase digitization capacity. ZooKeys 2012, 2009, 203–217. [Google Scholar] [CrossRef] [Green Version]
- Mammola, S.; Lunghi, E.; Bilandzija, H.; Cardoso, P.; Grimm, V.; Schmidt, S.I.; Hesselberg, T.; Martínez, A. Collecting eco-evolutionary data in the dark: Impediments to subterranean research and how to overcome them. Ecol. Evol. 2021, 11, 5911–5926. [Google Scholar] [CrossRef]
- Schielzeth, H.; Dingemanse, N.J.; Nakagawa, S.; Westneat, D.F.; Allegue, H.; Teplitsky, C.; Réale, D.; Dochtermann, N.A.; Garamszegi, L.Z.; Araya-Ajoy, Y.G. Robustness of linear mixed-effects models to violations of distributional assumptions. Methods Ecol. Evol. 2020, 11, 1141–1152. [Google Scholar] [CrossRef]
- French, C.M.; Ingram, T.; Bolnick, D.I. Geographical variation in colour of female threespine stickleback (Gasterosteus aculeatus). PeerJ 2018, 6, peerj-e4807. [Google Scholar] [CrossRef]
- Corciolani, G.; Vichi, A. Repeatability of colour reading with a clinical and a laboratory spectrophotometer. Int. Dent. SA 2006, 8, 62–70. [Google Scholar]
- MacCoun, R.; Perlmutter, S. Hide results to seek the truth. Nature 2015, 526, 187–189. [Google Scholar] [CrossRef]
- Sannolo, M.; Gatti, F.; Mangiacotti, M.; Scali, S.; Sacchi, R. Photo-identification in amphibian studies: A test of I3S Pattern. Acta Herpetol. 2016, 11, 63–68. [Google Scholar] [CrossRef]
- Lunghi, E.; Romeo, D.; Mulargia, M.; Cogoni, R.; Manenti, R.; Corti, C.; Ficetola, G.F.; Veith, M. On the stability of the dorsal pattern of European cave salamanders (genus Hydromantes). Herpetozoa 2019, 32, 249–253. [Google Scholar] [CrossRef] [Green Version]
- Paulraj, M.P.; Hema, C.R.; Krishnan, R.P.; Mohd Radzi, S.S. Color recognition algorithm using a neural network model in determining the ripeness of a banana. In Proceedings of the International Conference on Man-Machine Systems (ICoMMS), Batu Ferringhi, Malaysia, 11–13 October 2009; pp. 2B7-1–2B7-4. [Google Scholar]
- Jablonski, D.; Szabolcs, M.; Simović, A.; Mizsei, E. Color and pattern variation of the Balkan whip snake, Hierophis gemonensis (Laurenti, 1768). Turk. J. Zool. 2017, 41, 363–369. [Google Scholar] [CrossRef]
- Vroonen, J.; Vervust, B.; Fulgione, D.; Maselli, V.; Van Damme, R. Physiological colour change in the Moorish gecko, Tarentola mauritanica (Squamata: Gekkonidae): Effects of background, light, and temperature. Biol. J. Linn. Soc. 2012, 107, 182–191. [Google Scholar] [CrossRef] [Green Version]
- Bell, A.M.; Hankison, S.J.; Laskowski, K.L. The repeatability of behaviour: A meta-analysis. Anim. Behav. 2009, 77, 771–783. [Google Scholar] [CrossRef] [Green Version]
- Raffard, A.; Santoul, F.; Cucherousset, J.; Blanchet, S. The community and ecosystem consequences of intraspecific diversity: A meta-analysis. Biol. Rev. 2019, 94, 648–661. [Google Scholar] [CrossRef]
- Tao, L.; Xu, G. Color in machine vision and its application. Chin. Sci. Bull. 2001, 46, 1411–1421. [Google Scholar] [CrossRef]
- Yagiz, Y.; Balaban, M.O.; Kristinsson, H.G.; Welt, B.A.; Marshall, M.R. Comparison of Minolta colorimeter and machine vision system inmeasuring colour of irradiated Atlantic salmon. J. Sci. Food Agric. 2009, 89, 728–730. [Google Scholar] [CrossRef]
- Rondinini, C.; Battistoni, A.; Peronace, V.; Teofili, C. Lista Rossa IUCN dei Vertebrati Italiani; Comitato Italiano IUCN e Ministero dell’Ambiente e della Tutela del Territorio e del Mare: Roma, Italy, 2013; p. 54. [Google Scholar]
- Drew, J.A.; Moreau, C.S.; Stiassny, M.L.J. Digitization of museum collections holds the potential to enhance researcher diversity. Nat. Ecol. Evol. 2017, 1, 1789–1790. [Google Scholar] [CrossRef] [PubMed]




{kind=link}
{kind=link}
{kind=link}
| Type of Equipment | Brand | Model | Settings | |||
|---|---|---|---|---|---|---|
| Camera | Panasonic | FZ300 | DiaphragmF8 | SensitivityISO 100 | Shutter speed 1/500 | |
| Flash (×2) | Neewer | Speedlite NW620 | Guide number 58 | Light intensity 1/128 | Zoom 18 mm | Colour temperature 5500 K |
| Flash (×1) | Yongnuo | YN14EX | Light intensity 1/64 | Colour temperature 5600 K | ||
| Light diffuser (×2) | TYCKA | EU-TK009 | Dimension 23 × 18 | |||
| Trigger (×3) | Neewer | 2.4 GHz trigger FC16 | ||||
| Remote control | Smartphone application | Image app | ||||
| Variable | Description | Simulated Event |
|---|---|---|
| Height of the flash | Flash supports (B, Figure 1) were positioned at the level of the shooting area (Normal) or 4 cm lower (Down) | This simulates the different height of light source. This problem happens if flashes do not have a fixed position, but it is adjusted every time according to the subject and/or the environment |
| Distance of the flash | Flash are both secured on the closest (19 cm) or the farthest (24 cm) thread | This simulates the different distance of light source. This problem happens if flashes do not have a fixed position, but it is adjusted every time according to the subject and/or the environment |
| Light condition | The light condition is disturbed by: nothing (A), a yellow parasite light (B), a hybrid parasite light (C), a different inclination of one flash (D) | This simulates multiple problems related to the managing of light source. The category A represents the most accurate condition, where no external light is introduced into the shooting area (e.g., complete darkness) and both flashes direct their light in the same way. The category B and C simulate an external source of light (e.g., sunlight or a pilot light) which can be used to set the camera focus. This usually happens when the photo shooting is not performed in darkness or when strong pilot lights are used to drive camera focus. The category D simulates a not uniform light direction. This problem can arise when flashes do not have fixed position or when sunlight is used. |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Lunghi, E.; Bacci, F.; Zhao, Y. How Can We Record Reliable Information on Animal Colouration in the Wild? Diversity 2021, 13, 356. https://doi.org/10.3390/d13080356
Lunghi E, Bacci F, Zhao Y. How Can We Record Reliable Information on Animal Colouration in the Wild? Diversity. 2021; 13(8):356. https://doi.org/10.3390/d13080356
Chicago/Turabian StyleLunghi, Enrico, Francesco Bacci, and Yahui Zhao. 2021. "How Can We Record Reliable Information on Animal Colouration in the Wild?" Diversity 13, no. 8: 356. https://doi.org/10.3390/d13080356
APA StyleLunghi, E., Bacci, F., & Zhao, Y. (2021). How Can We Record Reliable Information on Animal Colouration in the Wild? Diversity, 13(8), 356. https://doi.org/10.3390/d13080356