Abstract
Understanding the mechanisms of diversity–productivity relationships is a central question in community ecology. Grazing is the main driving force affecting biodiversity, function, and stability of grassland ecosystems, and thus should play an important role in mediating diversity-productivity relationships. In this study, we examined the effect of grazing intensity on both aboveground biomass and biodiversity and explored the relationship between them in alpine meadow ecosystems in Sanjiangyuan, which is the source of the Yangtze, Yellow, and Lancang rivers. The results showed that the aboveground biomass and species richness decreased significantly due to multi-state succession in alpine meadows caused by long-term grazing, while the Shannon–Wiener index and Pielou evenness index decreased and then increased with increasing grazing intensity. The relationship between the aboveground biomass and biodiversity was U-shaped. Our results highlighted the opposite pattern of the diversity–productivity relationship under low and medium grazing intensity versus an extremely high grazing intensity; evenness contributed largely to this pattern. This study provided a new perspective on grassland management and the relationship between productivity and biodiversity. Attention should be paid to rational grazing to restore biodiversity and ecosystem functions and services in alpine meadows.
1. Introduction
Grasslands are widely distributed globally, contribute significantly to the balance between pasture and livestock production [1], and play an important role in the global carbon cycle [2]. However, the increasing intensity of human activities is changing grassland ecosystems, and overgrazing is a great threat [3] that may affect both biodiversity and productivity, as well as their relationship.
Understanding grassland ecosystem functions such as productivity in the face of anthropogenic environmental change is an important issue in sustainable ecosystem management. The relationship between biodiversity and biomass or productivity has been a central focus in community ecology [4]. Numerous experimental and theoretical works have shown that biodiversity increases community productivity [5,6,7]. Two sets of mechanisms have been proposed to explain the positive effects of biodiversity on ecosystem functioning (such as productivity), namely complementarity effects (such as differences in resource use) and selection effects (higher productivity advantage) [8]. In addition to a positive association [9], the relationship between species diversity and biomass has been manifested in four other forms [4,10,11]: a negative association, a unimodal relationship, a U-shaped relationship, and no clear association. However, due to the spatiotemporal differences among the research objects, the views and interpretations of these five relationship patterns do not form a unified theory or mechanism.
Overgrazing is considered to be one of the most important disturbance factors that contributes to grassland degradation and the declining grassland productivity worldwide [12,13]. In previous studies, grazing was shown to affect the structure and dynamics of vegetation communities. Overgrazing can increase or decrease species diversity and thus affect productivity depending on the type and intensity of grazing and resource use [14,15,16,17]. This particular response of species diversity to grazing may be one of the reasons for the effects of overgrazing on productivity.
The Qinghai–Tibet Plateau accounts for about 25% of the total rural area in China [18]; 40% of the plateau is alpine meadow [19]. This area is very sensitive to human interference. Due to long-term overgrazing, nearly 30% of alpine meadows on the Qinghai–Tibet Plateau have been seriously degraded. Studies have shown that ecosystem change is usually a slow process under grazing and other disturbances, but rapid and dramatic changes sometimes occur. The reason for such abrupt changes is often due to the existence of multi-stable ecosystems [20]. “Multi-stable ecosystem” refers to the existence of stable states with completely different structures and functions under the same conditions [21]. The occurrence of multi-stable ecosystems is often caused by overexploitation, and the restoration of such ecosystems often requires much external resource input. Affected by varying degrees by grazing activities, the alpine meadow ecosystem can contain four types of stable communities: the Gramineae grass–Kobresia humilis community, the K. humilis community, the K. pygmaea community, and Forbs-Black soil [19,22,23].
Most studies have shown that grazing significantly reduces grassland productivity and biodiversity on the Tibetan Plateau [24]. Previous studies focused on the relationship between productivity and biodiversity in one or different ecosystems and attempted to reveal the underlying mechanisms of this relationship. There are few studies on the relationship between biodiversity and biomass in multi-steady-state ecosystems formed by a long-term disturbance such as grazing.
In this study, we investigated the relationship between the aboveground biomass and biodiversity in four types of alpine meadows after grassland degradation due to long-term grazing in Sanjiangyuan, the source of the Yangtze, Yellow, and Lancang rivers. We aimed to reveal the biodiversity–productivity relationships in multi-stable ecosystem communities in the alpine meadows under the background of overgrazing. It is of great significance to improve the ecological service of the alpine grassland on the Qinghai–Tibet Plateau to rationally graze grassland according to the carrying capacity and to maintain the ecological security of the Qinghai–Tibet Plateau and its middle and lower reaches.
2. Materials and Methods
2.1. Study Area
The study area was selected from the Yellow River source area in Sanjiangyuan, Qinghai Province, China (Figure 1). Located in the southeastern part of Qinghai Province, the area is an important ecological shelter on the Tibetan Plateau and an important place for herdsmen and nomadic people. The area exhibits an alpine and semi-humid climate; there are no obvious four seasons and no absolute frost-free period. The cold monsoon brings a large amount of snow. It lasts for 7–8 months, causing many snow disasters. The warm season has a humid climate lasting for 4–5 months. The highest temperature is 23.2 °C and the lowest temperature −34 °C, with an annual average of −0.5 °C. The diurnal range of temperature is 15–25 °C. The average annual precipitation is 595 mm. The vegetation types are mainly alpine shrub meadows dominated by Potentilla fruticosa, Salix cupularis, and alpine meadow vegetation with a dominant species of Kobresia humilis.
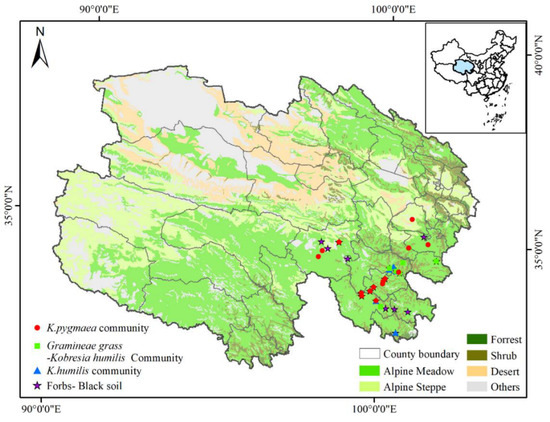
Figure 1.
Locations of research sites in the multi-stable alpine meadows in southeastern Qinghai Province.
2.2. Experimental Design
We chose 49 alpine meadow sites in the research area, each of which contained around 3 plots; the area of each plot was 50 cm × 50 cm. There were a total of 143 plots. All the plots showed a similar species composition initially before the experiment. Different intensities of grazing have been conducted since 2007. Grazing was conducted from October to April in the following year. The grazing intensities can be found in Table 1. Due to the different intensities of grazing, the meadow communities were divided into four types of typical and well-developed communities: the Kobresia humilis community (GK), the K. humilis community (KH), the K. pygmaea community (KP), and the Forbs-Black soil (FB) (Table 1). From GK to KH, KP, and FB, the communities suffered from an increasing grazing intensity. The plants in each plot were sampled in 2015 using harvesting methods around the peak season of plant growth (late July). We recorded the species, density, height, and coverage. The aboveground biomass was measured in dry weight using plants dried to a constant weight in a 70 °C incubator. The list of all species in each type of communities can be found in Table S1.

Table 1.
Feature of four types of communities.
2.3. Measures of Biodiversity
Plant species diversity was measured for each plot and estimated from three biodiversity indices [25]:
- (i).
- Richness index (R), the number of vascular species recorded in each plot;
- (ii).
- Shannon-Wiener Diversity index(H), calculated as:
- (iii).
- Pielou evenness index (J), calculated as:where Pi = Ni/N, Ni is the importance value of species i (which is calculated based on plant coverage), and N is the total importance value of all vascular species present.
2.4. Statistical Analysis
Data from all plots of each type of community were pooled to test for differences in plant richness, evenness, diversity, and total aboveground biomass across different levels of grazing intensity. A one-way ANOVA was conducted and Duncan’s post hoc test was used to compare differences among different groups. Before the ANOVA, the Shapiro–Wilk test was used to check the normality of the data. Depending on the pattern of the scatterplot, quadratic regression or simple linear regression was used to analyze the relationship between the plant diversity and the aboveground biomass. The calculation of the biodiversity index was performed using the vegan package in R version 3.6.0; all statistical analyses were performed using SPSS 17.0.
3. Results
3.1. Soil Properties of the Multi-Stable Alpine Meadows
The soil properties of the four types of communities at different depths can be found in Figure 2, which shows that soil organic matter (SOM) had nonsignificant differences across the four types of communities in general (except in the surface soil, in which KH and KP showed a significantly higher SOM than GK and FB). For soil bulk density (BD), it had nonsignificant differences across the four types of communities in general (except in the surface soil, in which KH and KP showed a significantly lower SOM than GK and FB). For soil compaction (SC), in general KP and FB showed a significant lower value than GK and KH. The four types of communities showed nonsignificant differences in soil total porosity (TP), soil capillary porosity (CP), and noncapillary porosity (NCP) at a shallow depth.
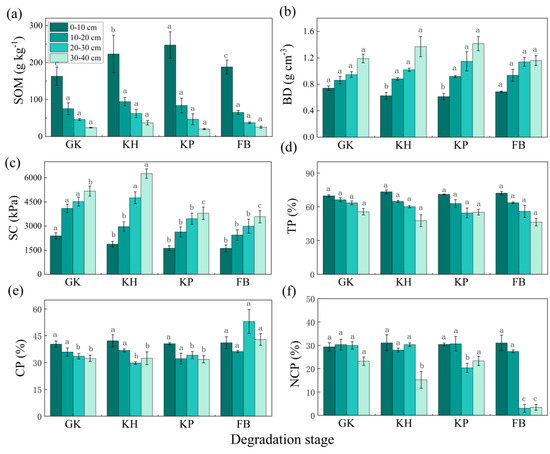
Figure 2.
Soil properties of multi-stable alpine meadows at different depths: (a) soil organic matter (SOM); (b) soil bulk density (BD); (c) soil compaction (SC); (d) soil total porosity (TP); (e) soil capillary porosity (CP); (f) noncapillary porosity (NCP). The error bar represents the standard error. Letters above bars mean a statistical significance was detected using ANOVA and Duncan’s post hoc test (α = 0.05) for each depth across different types of communities. GK, Gramineae grass–Kobresia humilis community; KH, K. humilis community; KP, K. pygmaea community; FB, Forbs-Black soil.
3.2. Aboveground Biomass of the Multi-Stable Alpine Meadows
The aboveground biomass in the alpine meadows decreased significantly with increasing degrees of degradation: from 287.99 ± 3.96 g·m2 to 84.13 ± 2.53 g·m2 (p < 0.05; Figure 3). Among them, KH decreased by 18.8% compared with GK, KP decreased by 22.3% compared with KH, and FB decreased by 53.6% compared with KP. FB had a 70.7% reduction compared with GK.
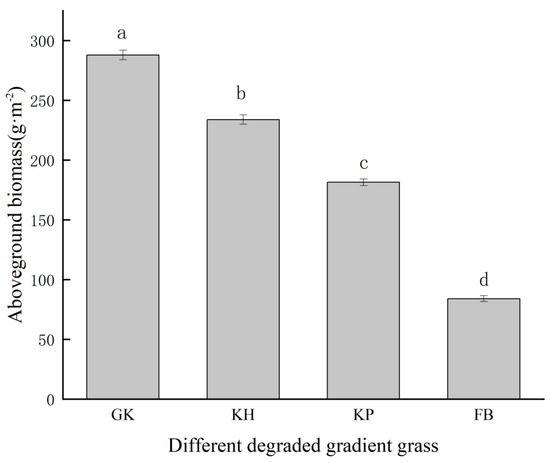
Figure 3.
Aboveground biomasses of multi−stable alpine meadows. The error bar represents the standard error. Letters above bars mean a statistical significance was detected using ANOVA and Duncan’s post hoc test (α = 0.05). GK, Gramineae grass–Kobresia humilis community; KH, K. humilis community; KP, K. pygmaea community; FB, Forbs-Black soil.
3.3. Biodiversity of the Multi-Stable Alpine Meadows
As the degradation increased, the species richness of the alpine meadows declined (Figure 4a). From GK to FB, the number of species was reduced by 50%. The Pielou evenness index and the Shannon–Wiener diversity index showed different patterns of species richness. These two indices showed a trend of decreasing firstly and then increasing. The lowest value was in KP rather than the most degraded FB (Figure 4b,c). Of particular note was that in the Pielou evenness index, the highest value was found for the most degraded FB, which showed a nonsignificant difference with the least degraded GK (Figure 4b).
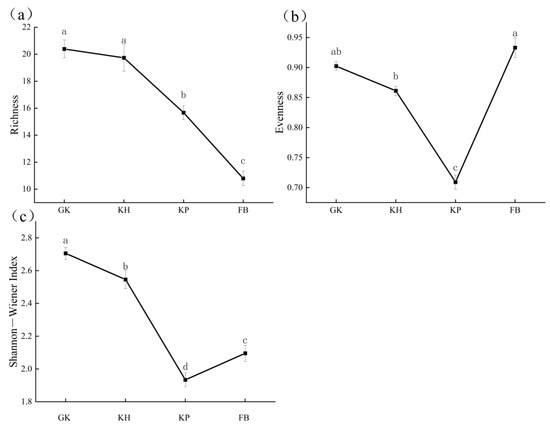
Figure 4.
Different biodiversity measures of the multi-stable alpine meadows: (a) species richness; (b) Pielou evenness; (c) Shannon–Wiener index. The error bar represents the standard error. Letters above bars mean a statistical significance was detected using ANOVA and Duncan’s post hoc test (α = 0.05). GK, Gramineae grass–Kobresia humilis community; KH, K. humilis community; KP, K. pygmaea community; FB, Forbs−Black soil.
3.4. Relationship between Biodiversity and Biomass
Scatter plots of the aboveground biomass and biodiversity indices across 143 plots in the four types of degraded alpine meadows showed quadratic relationships between the aboveground biomass and biodiversity (p < 0.05; Figure 5). It was clear that two clusters appeared in each panel. When the biomass was low, the relationships were negative or unclear. When the biomass was high, the relationships were clearly positive. The two clusters were caused by different levels of grazing intensity; that is, the left cluster consisted of points for a very high grazing intensity (FB), while the right cluster consisted of points for low or medium grazing intensities (GK, KH, and KP).
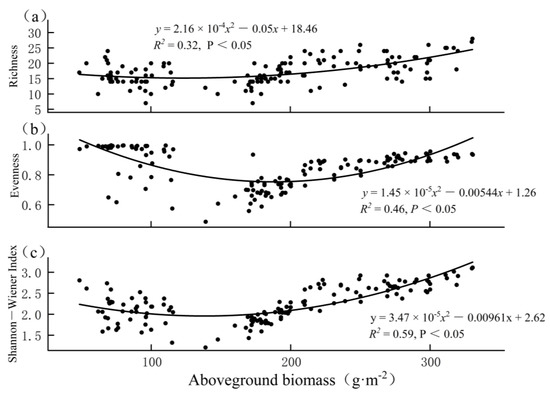
Figure 5.
Relationships between aboveground biomass and (a) richness, (b) evenness, and (c) Shannon−Wiener index of alpine meadows across all 143 plots.
Separate analyses of the relationships between the aboveground biomass and biodiversity across plots in GK, KH, and KP showed significant positive correlations between the biomass and diversity with Pearson correlation coefficients from 0.85 to 0.88 (p < 0.05; Figure 6). That is, both richness and evenness contributed to the positive relationship between the Shannon–Wiener biodiversity index and the aboveground biomass while considering biodiversity consisting of two components: richness and evenness.
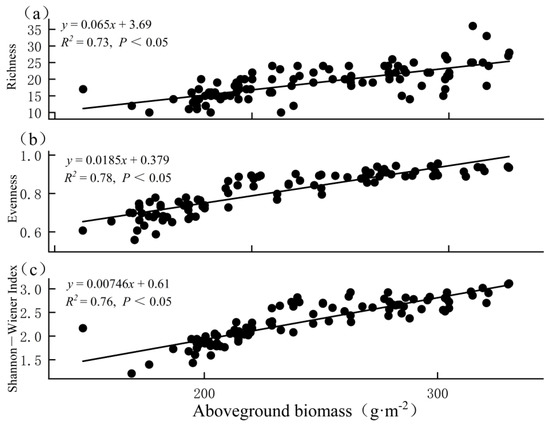
Figure 6.
Relationships between aboveground biomass and (a) richness, (b) evenness, and (c) Shannon−Wiener index across 99 plots in GK, KH, and KP.
Separate analyses of the plots in FB showed different relationships between the aboveground biomass and biodiversity (Figure 7). For richness, its relationship with the aboveground biomass was nonsignificant. For evenness, the correlation coefficient with the aboveground biomass was −0.57, indicating a significant negative correlation. As a result, the Shannon–Wiener biodiversity index and the aboveground biomass showed a correlation of −0.6. This meant that evenness, but not richness, contributed mostly to the relationship between the biodiversity and the aboveground biomass.
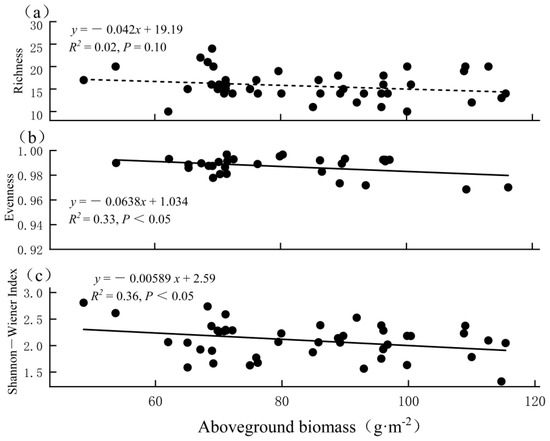
Figure 7.
Relationships between aboveground biomass and (a) richness, (b) evenness, and (c) Shannon−Wiener index across 44 plots in FB.
4. Discussion
The responses of the relationship between biodiversity and productivity to different grazing intensities were investigated by fitting the aboveground biomass and biodiversity across 143 plots in four types of degraded alpine meadows in Sanjiangyuan on the Qinghai–Tibet Plateau. We obtained three main results: (1) the aboveground biomass and species richness of the meadow communities decreased with an increase in the grazing intensity; (2) with the increase in grazing intensity, the evenness index and the Shannon–Wiener index firstly decreased but then increased; (3) the relationship between the aboveground biomass and the biodiversity of the alpine meadows with different degradation degrees was basically U-shaped.
Decades of experimental studies have reported the strong positive effects of biodiversity on ecosystem functioning; e.g., productivity [6,26,27]. Theoretically, this biodiversity effect can be explained by two sets of mechanisms, namely complementarity effects (CEs) and selection effects (SEs) [8]. CEs play a role if the average performance is higher in mixed cultures than in monocultures due to the niche differentiation or facilitation. Recent studies have highlighted the importance of complementary effects in determining productivity [28,29]. Meanwhile, SEs emerge if species with a higher monoculture productivity is dominant in mixed cultures. These mechanisms can be used to explain the positive diversity–productivity relationship under low or medium grazing pressure (Figure 6); that is, grazing-induced biodiversity loss impairs ecosystem productivity through CEs, SEs, or both. For example, changes in complementation effects among populations caused by grazing can affect community productivity. In addition, grazing may lead to a change in the dominant species, which would lead to a change in the selection effects and would affect community productivity. Usually, competition among species would also alter the diversity–stability relationships [30]. However, this may not be an issue when considering the poor living conditions on the Tibetan Plateau, where species interactions usually are symbiotic and competition is not strong enough, so communities with a high biomass could also have a high species richness. A recent study reported large changes in productivity of the dominant plant functional groups after 6 years of grazing [31]. found that grazing reduced the biodiversity and community productivity in the Xilin Gol grassland Nature Reserve, Inner Mongolia. This finding was consistent with our findings. Ref. [32] reported the impairment of biodiversity due to overgrazing. Our results of slight or medium grazing intensity showed a consistent pattern with these studies.
However, under a high grazing pressure, the biodiversity showed a negative correlation with the aboveground biomass that was mainly due to the negative correlation between the evenness and the biomass (Figure 7). Biodiversity consists of two aspects: species richness and evenness of abundance distribution among species. Prior studies generally focused on how species richness affected ecosystem productivity [27,33], and rare attention has been paid to the role of evenness (but see [24,34]. In our system, the Forbs-Black soil community was characterized by a low species richness but a high evenness. This is mainly caused by the loss in the large dominant species; e.g., Gramineae and Cyperaceae plants, and the communities consisted of different small weeds and had no clear dominant species. This on one hand increased the evenness of the abundance distribution among species and on other hand decreased the community biomass due to the lack of large species. Therefore, we observed a negative correlation between the aboveground biomass and the evenness (and thus the biodiversity). This negative evenness–biomass relationship was consistent with the findings of [24]. It is worth mentioning that although biodiversity and aboveground biomass are both indicators of community productivity [35], they must be considered comprehensively [36] because sometimes they showed different trends, especially in harsh environments (e.g., under a high grazing intensity in our case). Our results highlighted that the relationships between biodiversity and productivity may be different in good environments and in harsh environments. Moreover, the different relationships may be mediated by different mechanisms, and the contributions of richness and evenness may be altered.
The relationship between plant species diversity and biomass, as well as the interaction mechanism between plants and topography, soil, climatic factors, and human activities, is extremely complex; further research is needed on multi-stable alpine meadows to further reveal the underlying mechanisms therein. Our study could inspire further research on different aspects. First, our current study could not calculate the complementarity effects and selection effects because we did not conduct monoculture experiments [8]. So, future monoculture and polyculture studies can be conducted under different degrees of grazing intensity to quantitatively measure the complementarity effects and selection effects and explore how grazing alters these two types of effects and thus alters diversity–productivity relationships. Second, interactions between species and interaction networks can be further explored to determine how the degree of interactions [30] and topological properties of interaction networks [37] affect the diversity–stability relationships. Third, as shown by our result that evenness was more important than richness under a high grazing intensity in mediating diversity–productivity relationships, we could further check if this conclusion still holds for other ecosystem functions; e.g., stability. It has been well documented that species richness promotes community stability, mainly by promoting asynchrony among species [38,39]. It would be of great significance to determine if evenness plays an important role in mediating diversity–stability relationships, especially in harsh environments.
5. Conclusions
This study reported the effects of grazing on the relationship between the biodiversity and the biomass production in four types of alpine meadow plant communities in the Sanjiangyuan region of Qinghai Province. The results showed that the relationship between the aboveground biomass and the biodiversity in alpine meadows with different degrees of degradation was basically U-shaped. The productivity of alpine meadows and the biodiversity responded differently to degradation. Grazing impaired communities in terms of productivity and species richness. However, an extremely high grazing intensity promoted evenness and thus the biodiversity index, which contributed to the U shape of the relationship between the productivity and the biodiversity. Our study highlighted the opposite pattern of the diversity–productivity relationship under low and medium grazing intensities vs. an extremely high grazing intensity. We also emphasized the important role of evenness in mediating this opposite pattern.
Supplementary Materials
The following supporting information can be downloaded at: https://www.mdpi.com/article/10.3390/d14111002/s1, Table S1: List of all species in four types of communities.
Author Contributions
Conceptualization, D.Q. and K.S.; methodology, K.S. and L.Z.; software, K.S. and D.Q.; validation, X.G. and L.D.; formal analysis, L.Z. and K.S.; investigation, X.G. and Q.L.; data curation, K.S. and X.G.; writing—original draft preparation, K.S. and X.G.; writing—review and editing, L.Z. and D.Q.; visualization, K.S. and D.Q.; project administration, L.D.; funding acquisition, L.Z. and D.Q. All authors have read and agreed to the published version of the manuscript.
Funding
We acknowledge funding from the National Natural Science Foundation of China (Nos. 31901108 and 32001241), the Beijing Natural Science Foundation (No. 5222014), the National Key Research Development Program of China (No. 2021YFD190090307), the Hainan Provincial Joint Project of Sanya Yazhou Bay Science and Technology City (No: 2021JJLH0005), Sanya Yazhou Bay Science and Technology City Hainan Special Doctoral Science Research Fund Project (No:HSPHDSRF-2022-05-010) and the 2115 Talent Development Program of China Agricultural University.
Institutional Review Board Statement
Not applicable.
Data Availability Statement
The data and material that support the findings of this study are available from the corresponding author upon reasonable request.
Acknowledgments
We thank Xiaowei Guo for his help in visualization and the editor and reviewers for their valuable comments.
Conflicts of Interest
The authors declare that they have no conflict of interest.
References
- Feng, R.; Long, R.; Shang, Z.; Ma, Y.; Dong, S.; Wang, Y. Establishment of Elymus natans improves soil quality of a heavily degraded alpine meadow in Qinghai-Tibetan Plateau, China. Plant Soil 2010, 327, 403–411. [Google Scholar] [CrossRef]
- Yang, Y.; Fang, J.; Ji, C.; Han, W. Above- and belowground biomass allocation in Tibetan grasslands. J. Veg. Sci. 2009, 20, 177–184. [Google Scholar] [CrossRef]
- Hobbs, R.J.; Huenneke, L.F. Disturbance, Diversity, and Invasion: Implications for Conservation. Conserv. Biol. 1992, 6, 324–337. [Google Scholar] [CrossRef]
- Akatov, V.V.; Akatova, T.V.; Chefranov, S.G. Relationship between Degree of Dominance and Species Richness in Grass Communities with Different Productivities. Contemp. Probl. Ecol. 2018, 11, 355–365. [Google Scholar] [CrossRef]
- Cardinale, B.J.; Gross, K.; Fritschie, K.; Flombaum, P.; Fox, J.W.; Rixen, C.; Ruijven, J.V.; Reich, P.B.; Scherer-Lorenzen, M.; Wilsey, B.J. Biodiversity simultaneously enhances the production and stability of community biomass, but the effects are Independent. Ecology 2013, 94, 1697–1707. [Google Scholar] [CrossRef]
- Isbell, F.; Craven, D.; Connolly, J.; Loreau, M.; Schmid, B.; Beierkuhnlein, C.; Bezemer, T.M.; Bonin, C.; Bruelheide, H.; De Luca, E.; et al. Biodiversity increases the resistance of ecosystem productivity to climate extremes. Nature 2015, 526, 574–577. [Google Scholar] [CrossRef]
- Tilman, D.; Reich, P.B.; Knops, J.M. Biodiversity and ecosystem stability in a decade-long grassland experiment. Nature 2006, 441, 629–632. [Google Scholar]
- Loreau, M.; Hector, A. Partitioning selection and complementarity in biodiversity experiments. Nature 2001, 412, 72–76. [Google Scholar]
- Liu, Y.X.; Jian, W.H.; Qin, Y.; Xue-Fei, X.U.; Zhang, Y.Z. The relationship between biomass and biodiversity in the forest of Larix principis-rupprechtii. Hebei J. For. Orchard. Res. 2010, 25, 225–227. [Google Scholar]
- Bhattarai, K.R.; Vetaas, O.R.; Grytnes, J.A. Relationship between plant species richness and biomass in an arid sub-alpine grassland of the central Himalayas, Nepal. Folia Geobot. 2004, 39, 57–71. [Google Scholar] [CrossRef]
- Liira, J.; Zobel, K. The Species Richness-Biomass Relationship in Herbaceous Plant Communities: What Difference Does the Incorporation of Root Biomass Data Make? Oikos 2010, 91, 109–114. [Google Scholar] [CrossRef]
- Collins, S.L.; Glenn, J.M.; Briggs, S.M. Effect of local and regional processes on plant species richness in tallgrass prairie. Oikos 2002, 99, 571–579. [Google Scholar] [CrossRef]
- Sala, O.E.; Parton, W.J.; Joyce, L.A.; Lauenroth, W.K. Primary Production of the Central Grassland Region of the United States. Ecology 1988, 69, 40–45. [Google Scholar] [CrossRef]
- Cingolani, A.M.; Noy-Meir, I.; Díaz, S. Grazing Effects on Rangeland Diversity: A Synthesis of Contemporary Models. Ecol. Appl. 2005, 15, 757–773. [Google Scholar] [CrossRef]
- Collins, S.L.; Knapp, A.K.; Briggs, J.M.; Blair, J.M.; Steinauer, E.M. Modulation of diversity by grazing and mowing in native tallgrass prairie. Science 1998, 280, 745–747. [Google Scholar] [CrossRef] [PubMed]
- Filazzola, A.; Brown, C.; Dettlaff, M.A.; Batbaatar, A.; Grenke, J.; Bao, T.; Peetoom Heida, I.; Cahill, J.F., Jr. The effects of livestock grazing on biodiversity are multi-trophic: A meta-analysis. Ecol. Lett. 2020, 23, 1298–1309. [Google Scholar] [CrossRef]
- Wang, L.; Delgado-Baquerizo, M.; Wang, D.; Isbell, F.; Liu, J.; Feng, C.; Liu, J.; Zhong, Z.; Zhu, H.; Yuan, X.; et al. Diversifying livestock promotes multidiversity and multifunctionality in managed grasslands. Proc. Natl. Acad. Sci. USA 2019, 116, 6187–6192. [Google Scholar] [CrossRef]
- Wen, L.; Dong, S.; Li, Y.; Li, X.; Shi, J.; Wang, Y.; Liu, D.; Ma, Y. Effect of Degradation Intensity on Grassland Ecosystem Services in the Alpine Region of Qinghai-Tibetan Plateau, China. PLoS ONE 2013, 8, e58432. [Google Scholar] [CrossRef]
- Miehe, G.; Schleuss, P.M.; Seeber, E.; Babel, W.; Biermann, T.; Braendle, M.; Chen, F.; Coners, H.; Foken, T.; Gerken, T.; et al. The Kobresia pygmaea ecosystem of the Tibetan highlands—Origin, functioning and degradation of the world’s largest pastoral alpine ecosystem: Kobresia pastures of Tibet. Sci. Total Environ. 2019, 648, 754–771. [Google Scholar] [CrossRef]
- Kefi, S.; Rietkerk, M.; van Baalen, M.; Loreau, M. Local facilitation, bistability and transitions in arid ecosystems. Theor. Popul. Biol. 2007, 71, 367–379. [Google Scholar] [CrossRef]
- Scheffer, M.; Carpenter, S.R. Catastrophic regime shifts in ecosystems: Linking theory to observation. Trends Ecol. Evol. 2003, 18, 648–656. [Google Scholar] [CrossRef]
- Ganjurjav, H.; Gornish, E.S.; Hu, G.; Wan, Y.; Li, Y.; Danjiu, L.; Gao, Q. Temperature leads to annual changes of plant community composition in alpine grasslands on the Qinghai-Tibetan Plateau. Environ. Monit. Assess 2018, 190, 585. [Google Scholar] [CrossRef] [PubMed]
- Shang, Z.; Long, R. Formation causes and recovery of the “Black Soil Type” degraded alpine grassland in Qinghai-Tibetan Plateau. Front. Agric. China 2007, 1, 197–202. [Google Scholar] [CrossRef]
- Zhang, Z.; Zhou, H.; Zhao, X.; Yao, B.; Ma, Z.; Dong, Q.; Zhang, Z.; Wang, W.; Yang, Y. Relationship between biodiversity and ecosystem functioning in alpine meadows of the Qinghai-Tibet Plateau. Biodivers. Sci. 2018, 26, 111–129. [Google Scholar] [CrossRef]
- Zobel, K.J. A scale-independent approach to the richness vs biomass relationship in ground-layer plant communities. Oikos 1997, 80, 325–332. [Google Scholar] [CrossRef]
- Tilman, D. The Influence of Functional Diversity and Composition on Ecosystem Processes. Science 1997, 277, 1300–1302. [Google Scholar] [CrossRef]
- Tilman, D.; Reich, P.B.; Knops, J.; Wedin, D.; Mielke, T.; Lehman, C. Diversity and productivity in a long-term grassland experiment. Science 2001, 294, 843–845. [Google Scholar] [CrossRef]
- Wang, C.T.; Long, R.J. The effects of differences in functional group diversity and composition on plant community productivity in four types of alpine meadow communities. Chin. Biodivers. 2004, 12, 403–409. [Google Scholar]
- Xu, Q.; Yang, X.; Yan, Y.; Wang, S.; Loreau, M.; Jiang, L. Consistently positive effect of species diversity on ecosystem, but not population, temporal stability. Ecol. Lett. 2021, 24, 2256–2266. [Google Scholar] [CrossRef]
- Abrams, P.A. Resource Productivity-Consumer Species Diversity: Simple Models of Competition in Spatially Heterogeneous Environments. Ecology 1988, 69, 1433. [Google Scholar] [CrossRef]
- Liang, M.; Liang, C.; Hautier, Y.; Wilcox, K.R.; Wang, S. Grazing-induced biodiversity loss impairs grassland ecosystem stability at multiple scales. Ecol. Lett. 2021, 24, 2054–2064. [Google Scholar] [CrossRef] [PubMed]
- Gross, K.; Cardinale, B.J.; Fox, J.W.; Gonzalez, A.; Loreau, M.; Wayne Polley, H.; Reich, P.B.; van Ruijven, J. Species richness and the temporal stability of biomass production: A new analysis of recent biodiversity experiments. Am. Nat. 2014, 183, 1–12. [Google Scholar] [CrossRef] [PubMed]
- Huang, Y.; Chen, Y.; Castro-Izaguirre, N.; Baruffol, M.; Brezzi, M.; Lang, A.; Li, Y.; Härdtle, W.; Von Oheimb, G.; Yang, X.; et al. Impacts of species richness on productivity in a large-scale subtropical forest experiment. Science 2018, 362, 80–83. [Google Scholar] [CrossRef] [PubMed]
- Mulder, C.P.; Bazeley-White, E.; Dimitrakopoulos, P.G.; Hector, A.; Scherer-Lorenzen, M.; Schmid, B. Species evenness and productivity in experimental plant communities. Oikos 2004, 107, 50–63. [Google Scholar] [CrossRef]
- Briske, D.D.; Fuhlendorf, S.D.; Smeins, F.E. State-and-Transition Models, Thresholds, and Rangeland Health: A Synthesis of Ecological Concepts and Perspectives. Rangel. Ecol. Manag. 2005, 58, 1–10. [Google Scholar] [CrossRef]
- Folke, C.; Carpenter, S.; Walker, B.; Scheffer, M.; Elmqvist, T.; Gunderson, L.; Holling, C.S. Regime Shifts, Resilience, and Biodiversity in Ecosystem Management. Annu. Rev. Ecol. Evol. Syst. 2004, 35, 557–581. [Google Scholar] [CrossRef]
- Gong, Y.; Yu, L.; Zhao, L. Ecological Networks in Agroecosystems: Approaches and Applications. Front. Agric. Sci. Eng. 2022, 9, 523–535. [Google Scholar]
- Ma, Z.; Liu, H.; Mi, Z.; Zhang, Z.; Wang, Y.; Xu, W.; Jiang, L.; He, J.S. Climate warming reduces the temporal stability of plant community biomass production. Nat. Commun. 2017, 8, 15378. [Google Scholar] [CrossRef]
- Schnabel, F.; Liu, X.; Kunz, M.; Barry, K.E.; Bongers, F.J.; Bruelheide, H.; Fichtner, A.; Härdtle, W.; Li, S.; Pfaff, C.T.; et al. Species richness stabilizes productivity via asynchrony and drought-tolerance diversity in a large-scale tree biodiversity experiment. Sci. Adv. 2021, 7, eabk1643. [Google Scholar] [CrossRef]







Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).