
Impacts of Cd Pollution on the Vitality, Anatomy and Physiology of Two Morphologically Different Lichen Species of the Genera Parmotrema and Usnea, Evaluated under Experimental Conditions

 ,
, {kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Materials and Methods
2.1. Lichen Material and Experimental Conditions
2.2. Assessment of Algal Cell Viability and Tolerance Index
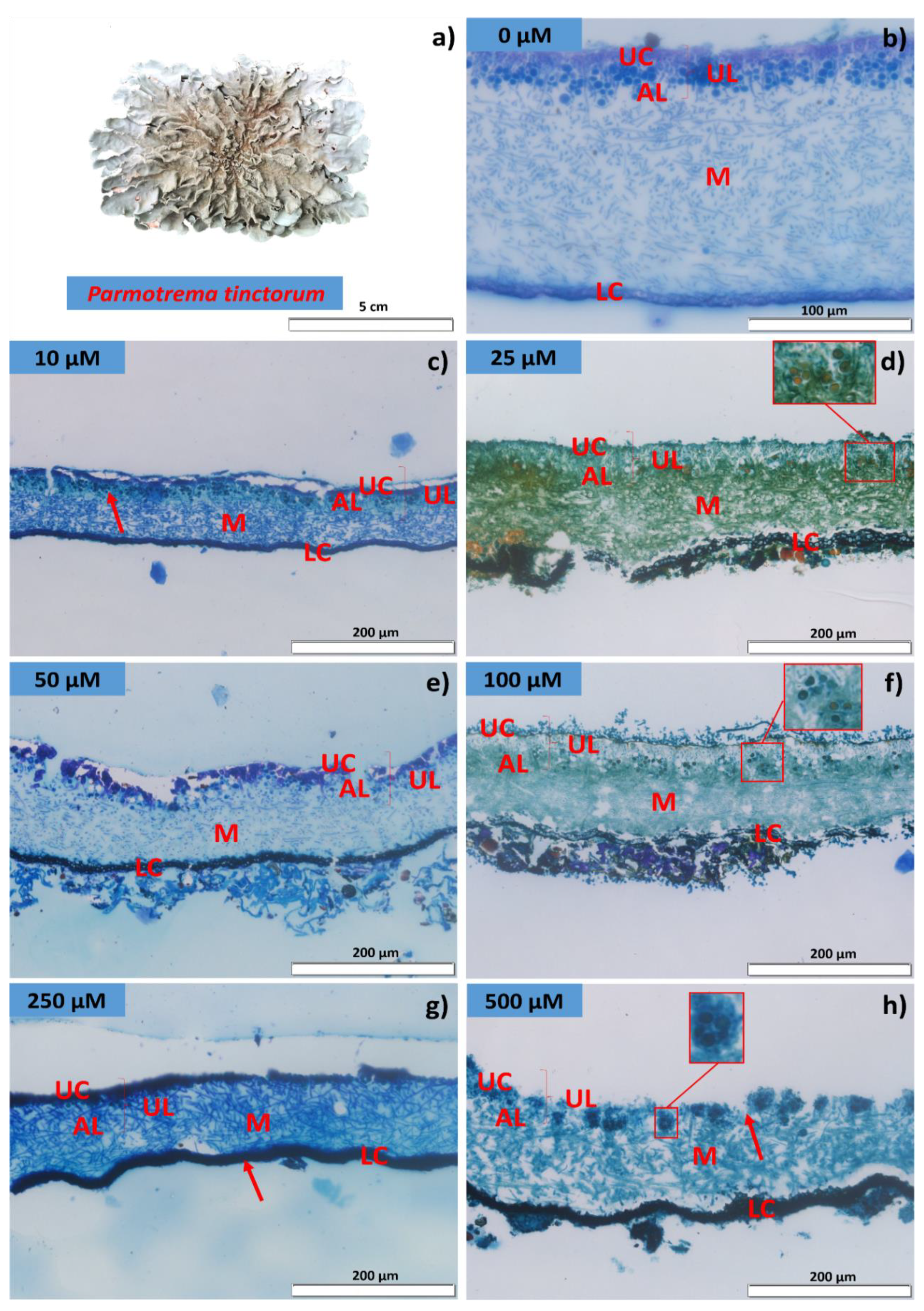
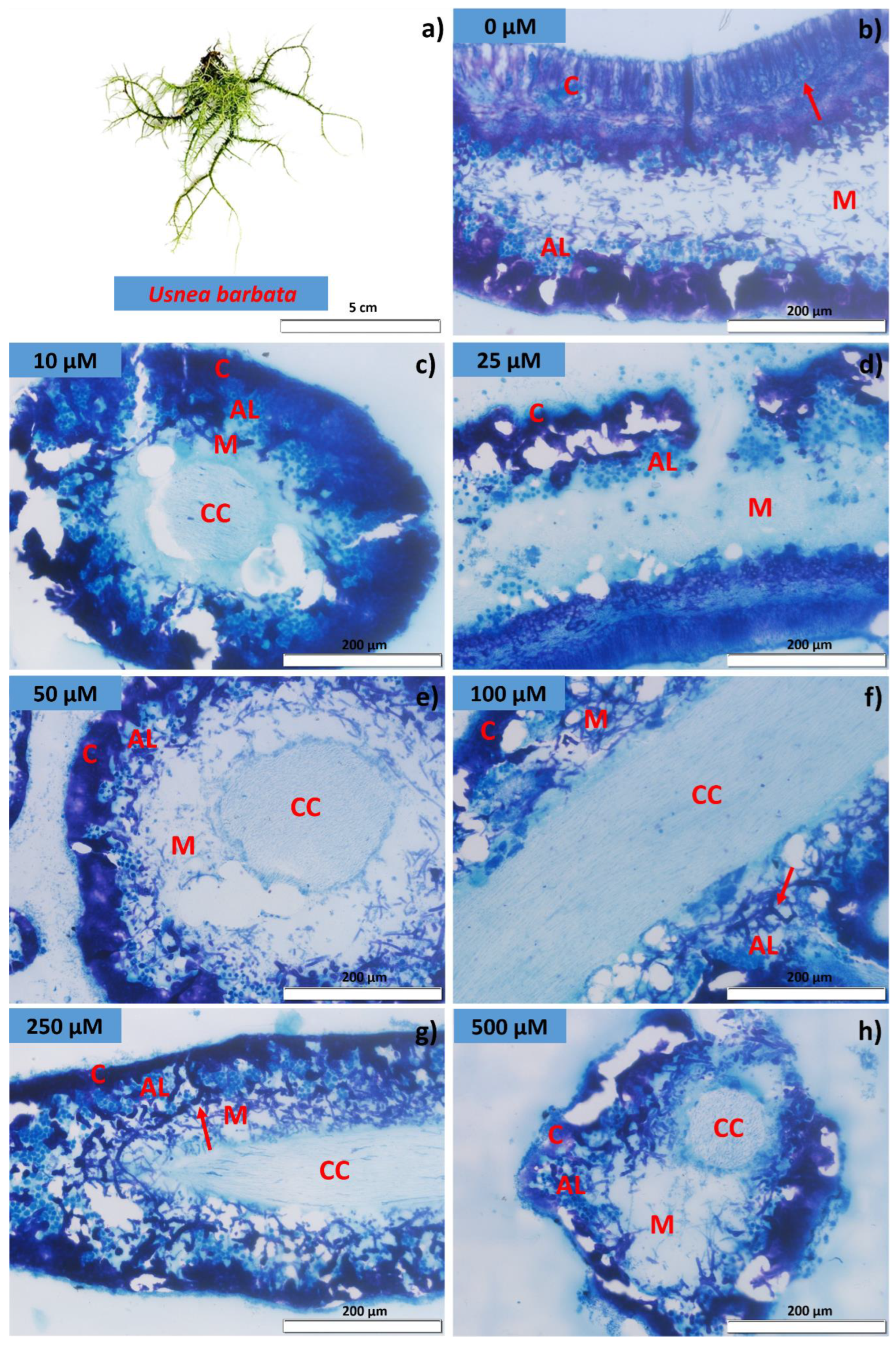
2.3. Membrane Integrity and Lichen Anatomy
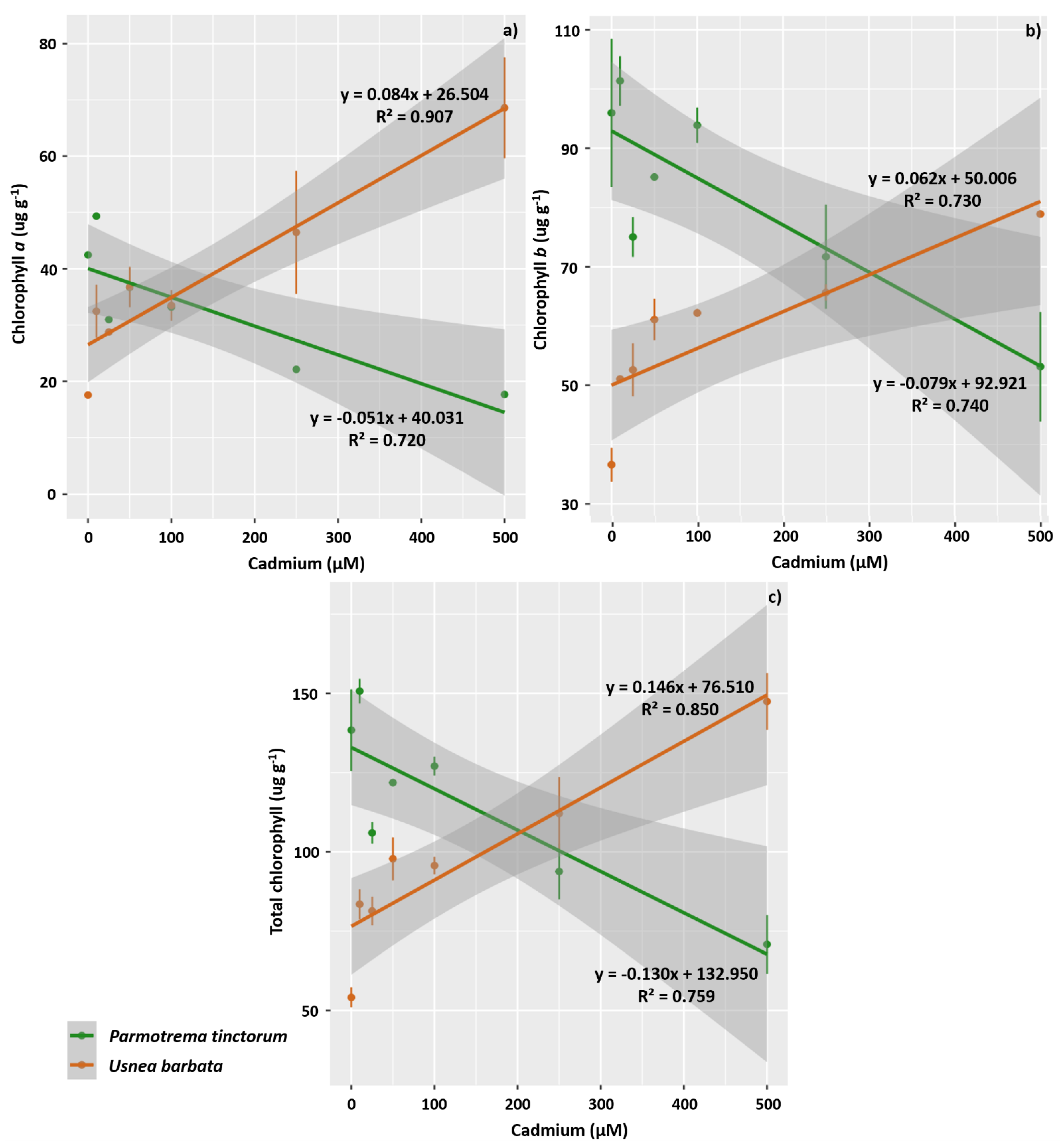
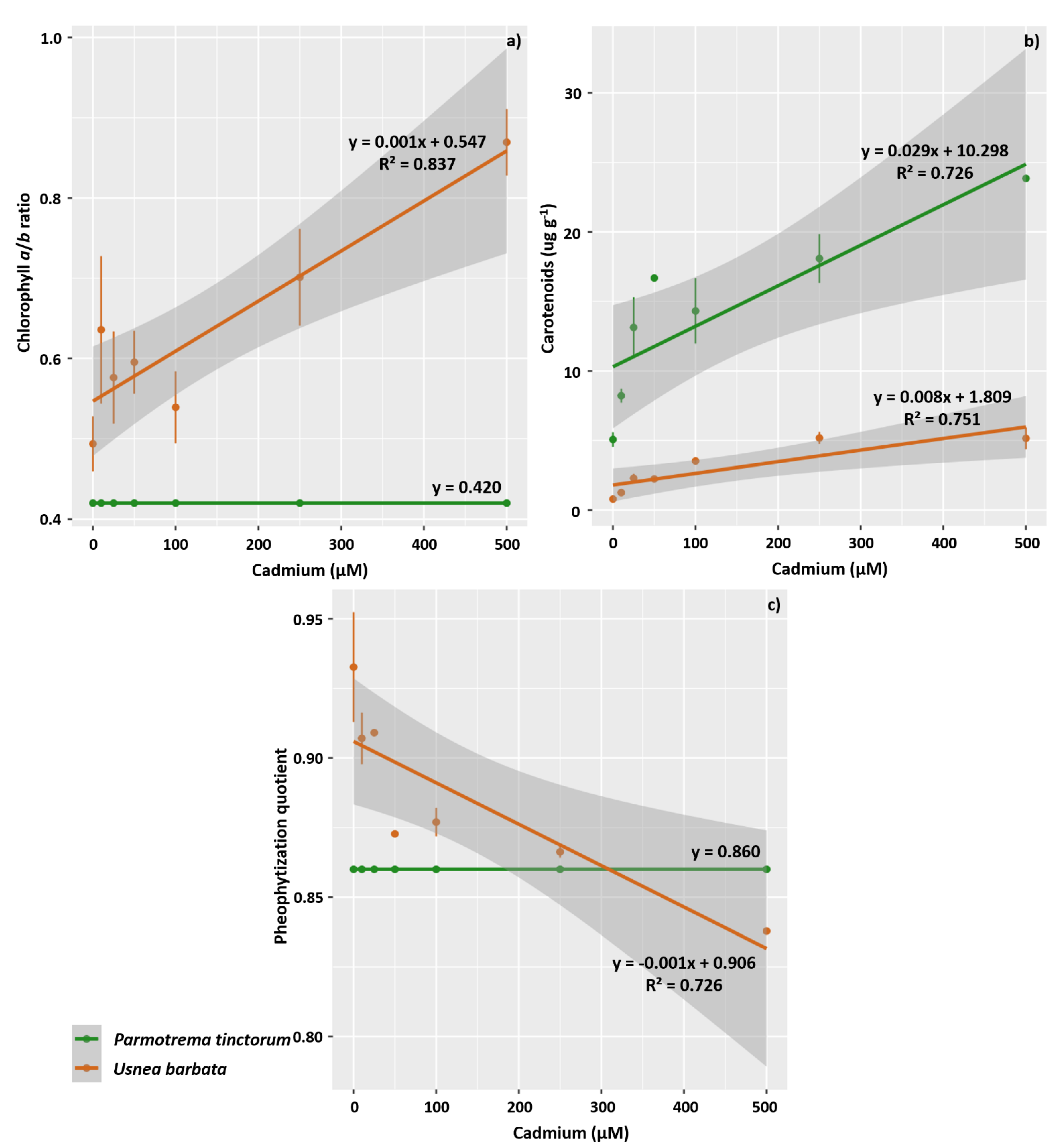
2.4. Evaluation of Photosynthetic Pigments
2.5. Chlorophyll a Fluorescence Parameters
2.6. Enzyme Activity of Antioxidant Metabolism, Hydrogen Peroxide, and Malondialdehyde
2.7. Experimental Design and Statistical Analyses
3. Results
3.1. Algal Cell Viability, Electrical Conductivity, and Lichen Anatomy
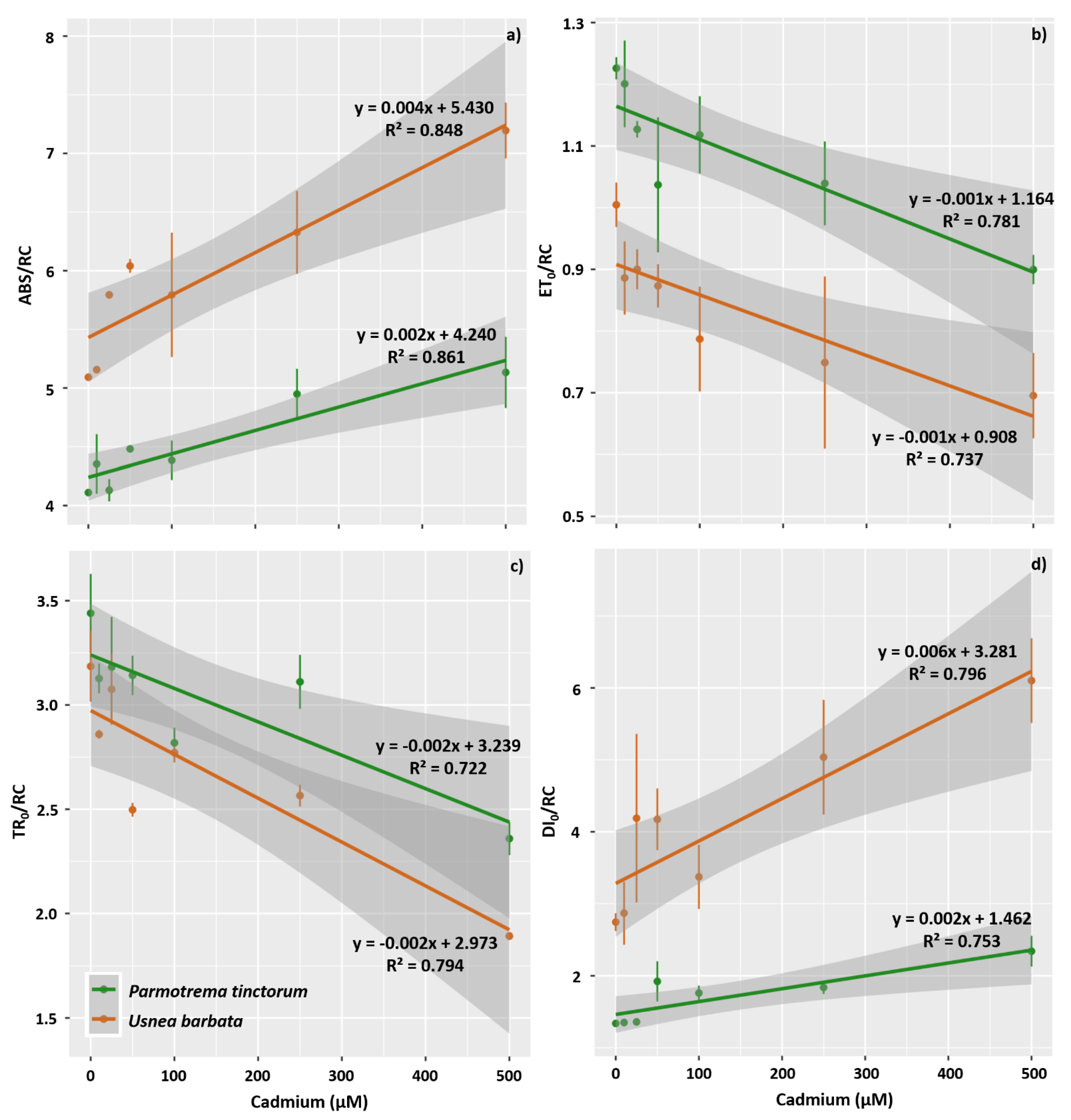
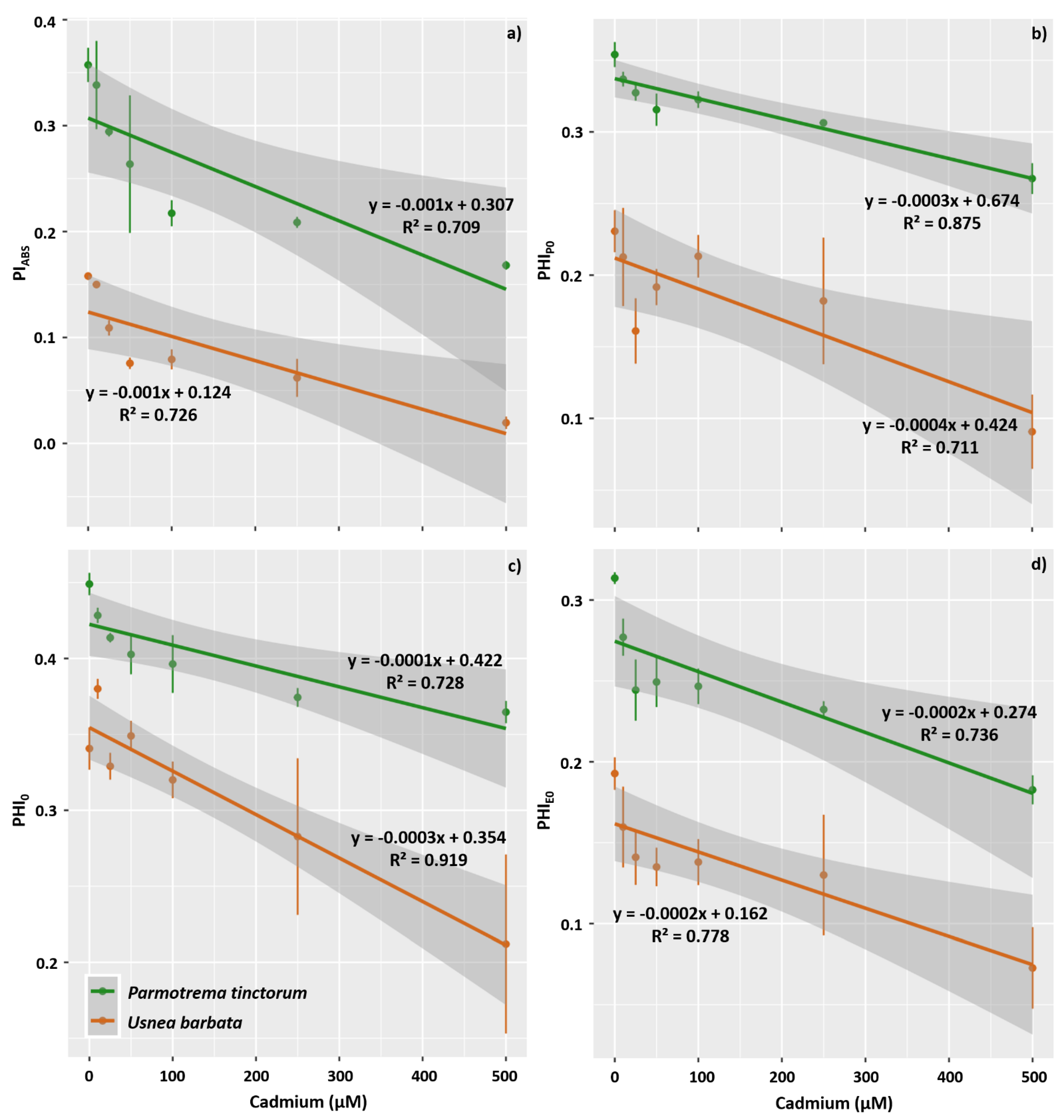
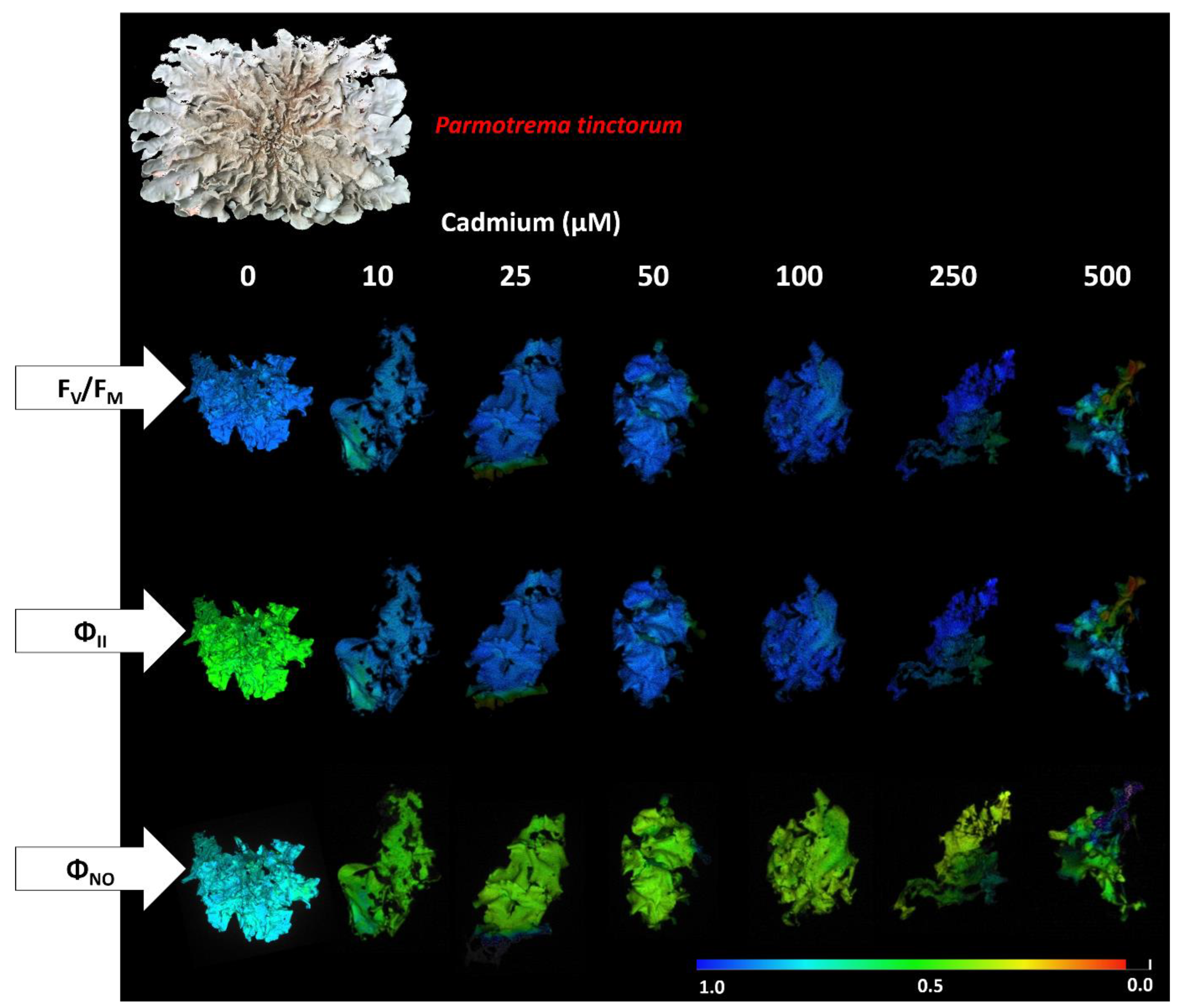
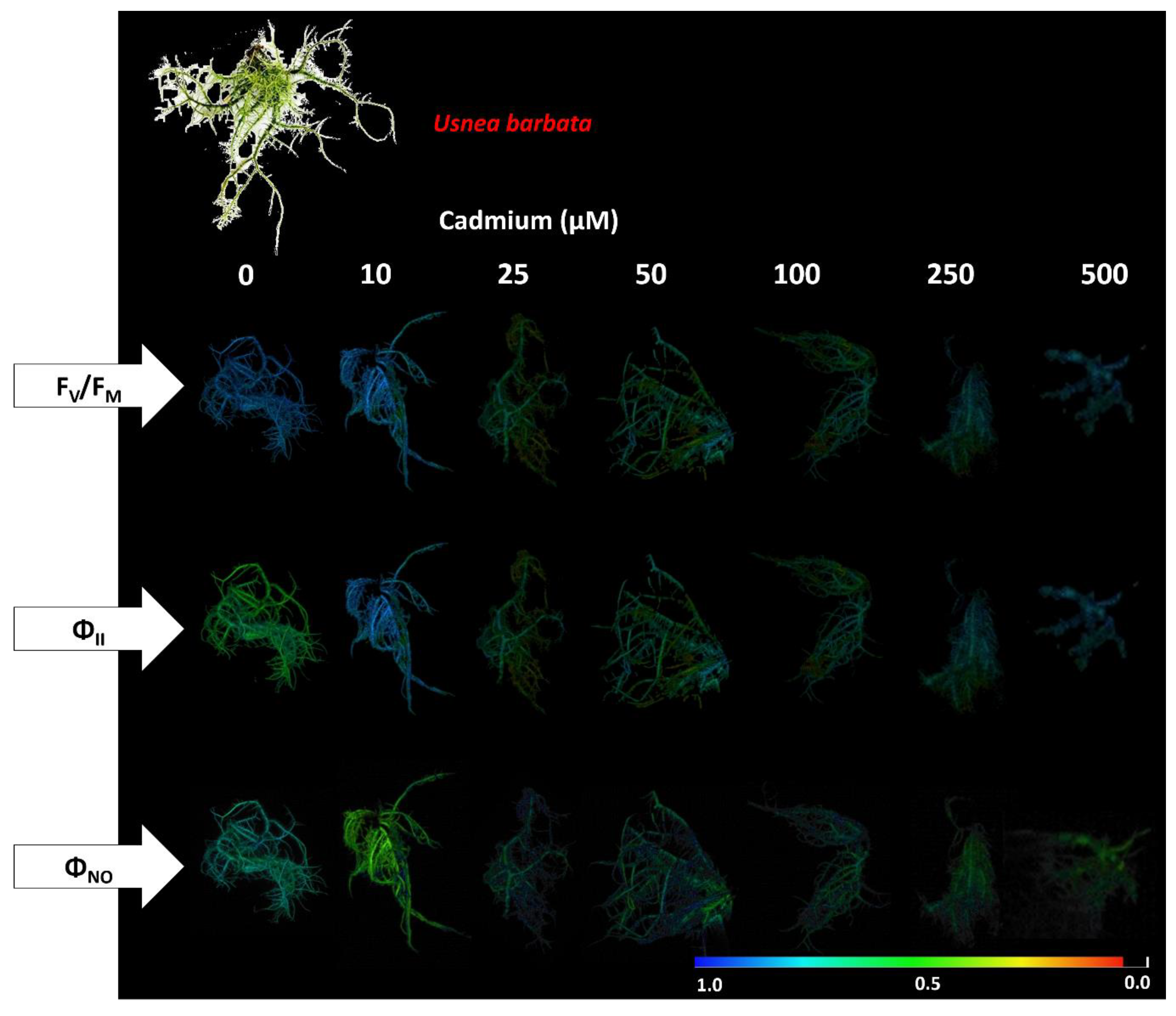
3.2. Photosynthetic Pigment Concentration and Chlorophyll a Fluorescence in the Photobiont
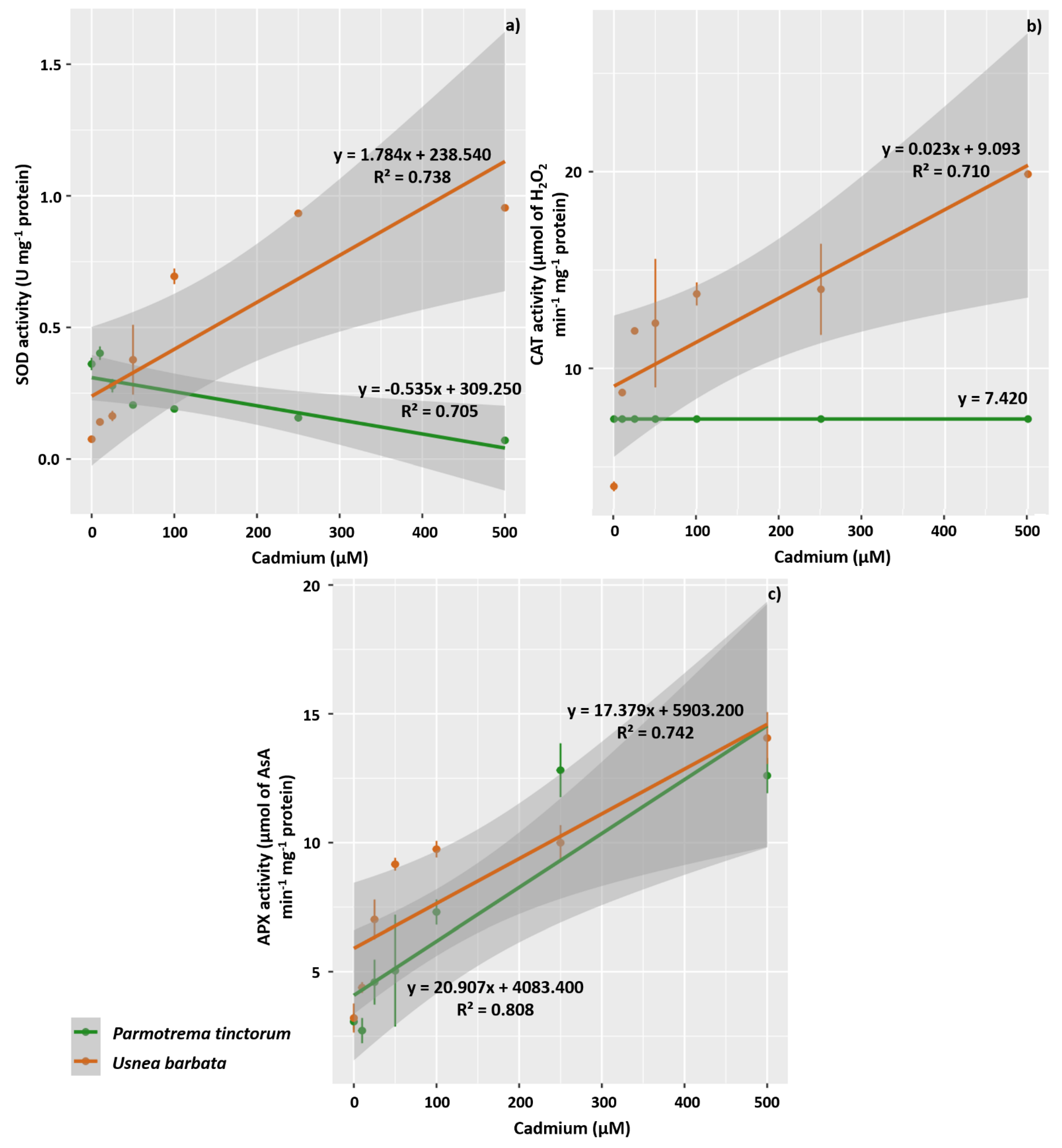
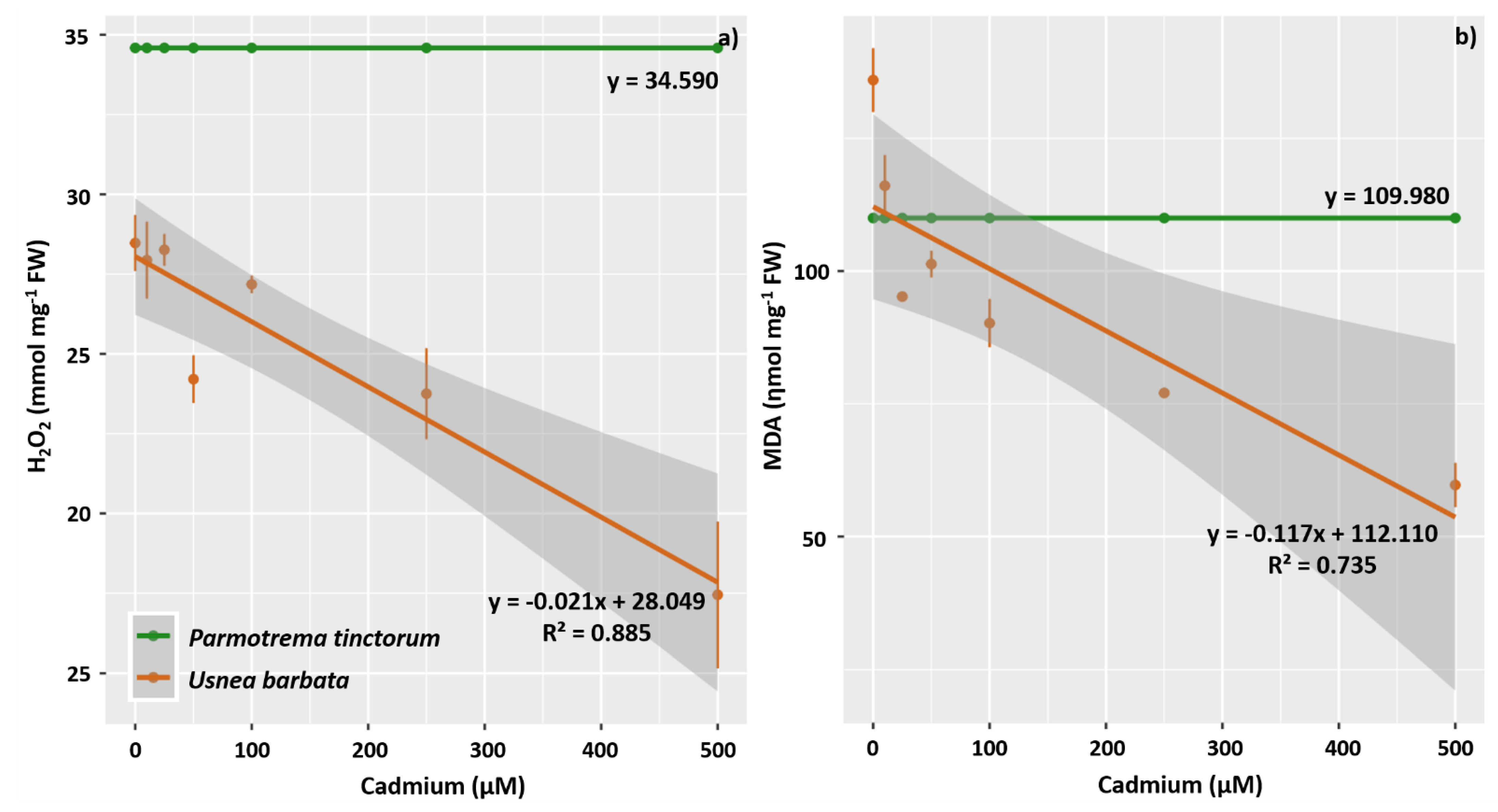
3.3. Enzyme Activity of Antioxidant Metabolism, H2O2, and MDA
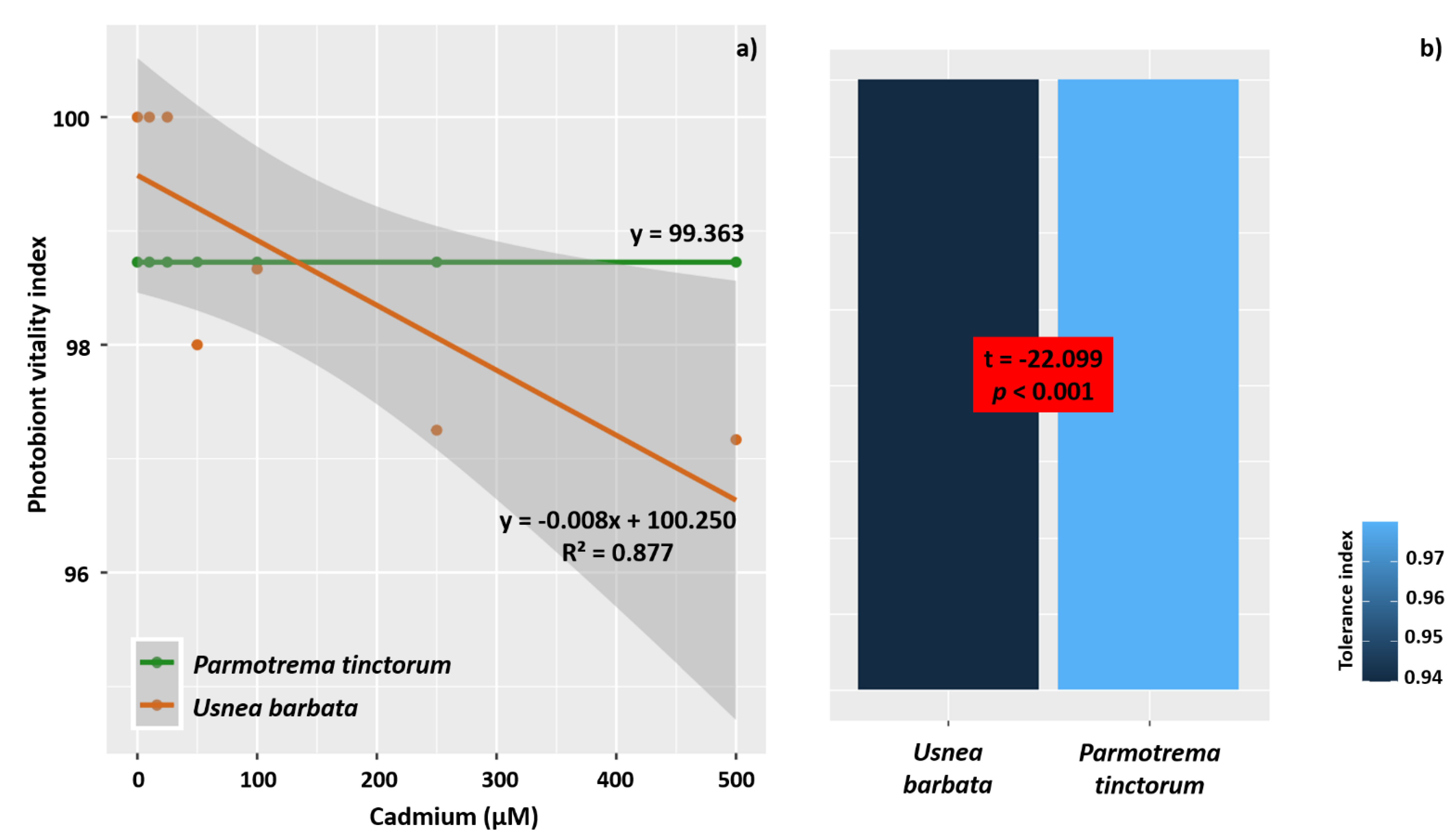
3.4. PVI and Tolerance Index
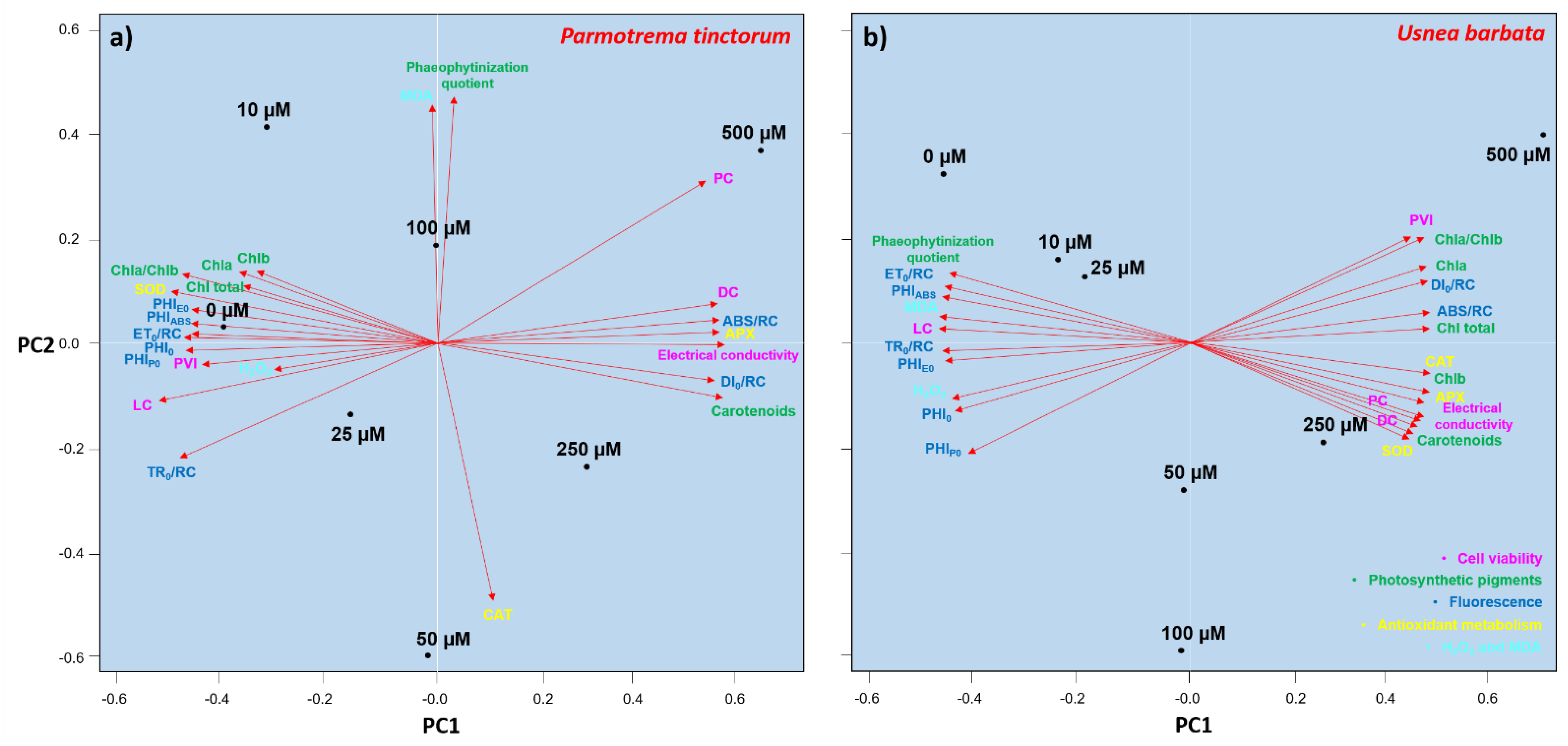
3.5. Analysis of Variables as Biomarkers of the Effect of Cd
4. Discussion
4.1. Cd Exposure Affects Photobiont Viability and Membrane Integrity, and Promotes Damage and Morphoanatomical Changes in P. tinctorum and U. barbata
4.2. Cd Exposure Negatively Affects the Concentration of Chlorophyll a, Chlorophyll b, and Total Chlorophyll in the Photobionts of P. tinctorum and Increases the Synthesis of Carotenoids
4.3. Cd Exposure Affects Primary Photochemistry in Photobionts of P. tinctorum and U. barbata
4.4. The Activity of Antioxidant Enzymes Increases in U. barbata Thalli Exposed to Increasing Concentrations of Cd
4.5. Parmotrema tinctorum Is More Sensitive to Cd Stress Than Usnea barbata
4.6. Biomarkers for the Effect of Cd-Induced Stress in P. tinctorum and U. barbata
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Melo, E.E.C.; Costa, E.T.S.; Guilherme, L.R.G.; Faquin, V.; Nascimento, C.W.A. Accumulation of arsenic and nutrients by castor bean plants grown on an As-enriched nutrient solution. J. Hazard. Mater. 2009, 168, 479–483. [Google Scholar] [CrossRef] [PubMed]
- Gall, J.E.; Boyd, R.S.; Rajakaruna, N. Transfer of heavy metals through terrestrial food webs: A review. Environ. Monit Assess. 2015, 187, 201. [Google Scholar] [CrossRef] [Green Version]
- Suhani, I.; Sahab, S.; Srivastava, V.; Singh, R.P. Impact of cadmium pollution on food safety and human health. Curr. Opin. Toxicol. 2021, 27, 1–7. [Google Scholar] [CrossRef]
- Yuan, Z.; Luo, T.; Liu, X.; Hua, H.; Zhuang, Y.; Zhang, X.; Ren, J. Tracing anthropogenic cadmium emissions: From sources to pollution. Sci. Total Environ. 2019, 676, 87–96. [Google Scholar] [CrossRef] [PubMed]
- Niño-Savala, A.G.; Zhuang, Z.; Ma, X.; Fangmeier, A.; Li, H.; Tang, A.; Liu, X. Cadmium pollution from phosphate fertilizers in arable soils and crops: An overview. Front. Agric. Sci. Eng. 2019, 6, 419–430. [Google Scholar] [CrossRef] [Green Version]
- Bačkor, M.; Váczi, P.; Barták, M.; Buďová, J.; Dzubaj, A. Uptake, photosynthetic characteristics and membrane lipid peroxidation levels in the lichen photobiont Trebouxia erici exposed to copper and cadmium. Bryologist 2007, 110, 100–107. [Google Scholar] [CrossRef]
- Huybrechts, M.; Cuypers, A.; Deckers, J.; Iven, V.; Vandionant, S.; Jozefczak, M.; Hendrix, S. Cadmium and plant development: An agony from seed to seed. Int. J. Mol. Sci. 2019, 20, 3971. [Google Scholar] [CrossRef] [Green Version]
- Jamers, A.; Blust, R.; De Coen, W.; Griffin, J.L.; Jones, O.A. An omics based assessment of cadmium toxicity in the green alga Chlamydomonas reinhardtii. Aquat. Toxicol. 2013, 126, 355–364. [Google Scholar] [CrossRef]
- Mishra, V.K.; Tripathi, B.D. Concurrent removal and accumulation of heavy metals by the three aquatic macrophytes. Bioresour. Technol. 2008, 99, 7091–7097. [Google Scholar] [CrossRef]
- Metwally, A.; Finkemeier, I.; Georgi, M.; Diezt, K.-J. Salicylic acid alleviates the cadmium toxicity in barley seedlings. Plant Physiol. 2003, 132, 272–281. [Google Scholar] [CrossRef] [PubMed]
- Kováčik, J.; Dresler, S.; Babula, P.; Hladký, J.; Sowa, I. Calcium has protective impact on cadmium-induced toxicity in lichens. Plant Physiol. Biochem. 2020, 156, 591–599. [Google Scholar] [CrossRef]
- Hassan, M.; Mansoor, S. Oxidative stress and antioxidant defense mechanism in mung bean seedlings after lead and cadmium treatments. Turk. J. Agric. For. 2014, 38, 55–61. [Google Scholar] [CrossRef]
- Wang, S.; Wufuer, R.; Duo, J.; Li, W.; Pan, X. Cadmium caused different toxicity to photosystem i and photosystem ii of freshwater unicellular algae Chlorella pyrenoidosa (Chlorophyta). Toxics 2022, 10, 352. [Google Scholar] [CrossRef]
- Conti, M.E.; Cecchetti, G. Biological monitoring: Lichens as bioindicators of air pollution assessment—A review. Environ. Pollut. 2001, 114, 471–492. [Google Scholar] [CrossRef]
- Raimundo-Costa, W.; Ferreira, D.C.; Anhê, A.C.B.M.; Senhuk, A.P.M.D.S. The use of Parmotrema tinctorum (Parmeliaceae) as a bioindicator of air pollution. Rodriguésia 2021, 72, e01872019. [Google Scholar] [CrossRef]
- Viana, C.O.; Vaz, R.P.; Cano, A.; Santos, A.P.; Cançado, L.G.; Ladeira, L.O.; Junior, A.C. Physiological changes of the lichen Parmotrema tinctorum as result of carbon nanotubes exposition. Ecotoxicol. Environ. Saf. 2015, 120, 110–116. [Google Scholar] [CrossRef] [PubMed]
- Conti, M.E.; Pino, A.; Botrè, F.; Bocca, B.; Alimonti, A. Lichen Usnea barbata as biomonitor of airborne elements deposition in the Province of Tierra del Fuego (southern Patagonia, Argentina). Ecotoxicol. Environ. Saf. 2009, 72, 1082–1089. [Google Scholar] [CrossRef] [PubMed]
- Palharini, K.M.Z.; Vitorino, L.C.; Bessa, L.A.; Vasconcelos Filho, S.C.; Silva, F.G. Parmotrema tinctorum as an indicator of edge effect and air quality in forested areas bordered by intensive agriculture. Environ. Sci. Pollut. Res. 2021, 28, 68997–69011. [Google Scholar] [CrossRef] [PubMed]
- Bizarro, V.G.; Meurer, E.J.; Tatsch, F.R.P. Cadmium contents of phosphate fertilizers marketed in Brazil. Ciênc Rural. 2008, 38, 247–250. [Google Scholar] [CrossRef] [Green Version]
- Gomes, L.; Simões, S.J.; Dalla Nora, E.L.; Sousa-Neto, E.R.; Forti, M.C.; Ometto, J.P.H. Agricultural expansion in the Brazilian Cerrado: Increased soil and nutrient losses and decreased agricultural productivity. Land 2019, 8, 12. [Google Scholar] [CrossRef]
- Palharini, K.M.Z.; Vitorino, L.C.; Menino, G.C.D.O.; Bessa, L.A. Edge effects reflect the impact of the agricultural matrix on the corticolous lichens found in fragments of cerrado savanna in central Brazil. Sustainability 2020, 12, 7149. [Google Scholar] [CrossRef]
- MAPA. Ministério da Agricultura, Pecuária e Abastecimento. SDA Normative Instruction: Limits of Phytotoxic Agents, Pathogens to Humans, Animals and Plants, Toxic Heavy Metals, Pests and Weeds Admitted in Fertilizers, Correctives, Inoculants and Biofertilizers. 2006. Available online: http://extranet.agricultura.gov.br/sislegisconsulta/consultarLegislacao.do?operacao=visualizar&id=16951 (accessed on 23 October 2022).
- Rola, K. Insight into the pattern of heavy-metal accumulation in lichen thalli. J. Trace Elem. Med. Biol. 2020, 61, 126512. [Google Scholar] [CrossRef]
- Vannini, A.; Tedesco, R.; Loppi, S.; Di Cecco, V.; Di Martino, L.; Nascimbene, J.; Barbante, C. Lichens as monitors of the atmospheric deposition of potentially toxic elements in high elevation Mediterranean ecosystems. Sci. Total Environ. 2021, 798, 149369. [Google Scholar] [CrossRef] [PubMed]
- Dzubaj, A.; Bačkor, M.; Tomko, J.; Peli, E.; Tuba, Z. Tolerance of the lichen Xanthoria parietina (L.) Th. Fr. to metal stress. Ecotoxicol. Environ. Saf. 2008, 70, 319–326. [Google Scholar] [CrossRef] [PubMed]
- Galun, M.; Ronen, R. Interaction of lichens and pollutants. In Handbook of Lichenology; CRC Press: Boca Raton, FL, USA, 2019; pp. 55–74. [Google Scholar]
- Monnet, F.; Bordas, F.; Deluchat, V.; Baudu, M. Toxicity of copper excess on the lichen Dermatocarpon luridum: Antioxidant enzyme activities. Chemosphere 2006, 65, 1806–1813. [Google Scholar] [CrossRef] [PubMed]
- Ohmura, Y.; Takeshita, S.; Kawachi, M. Photobiont diversity within populations of a vegetatively reproducing lichen, Parmotrema tinctorum, can be generated by photobiont switching. Symbiosis 2019, 77, 59–72. [Google Scholar] [CrossRef]
- Ohmura, Y.; Kawachi, M.; Kasai, F.; Watanabe, M.M.; Takeshita, S. Genetic combinations of symbionts in a vegetatively reproducing lichen, Parmotrema tinctorum, based on ITS rDNA sequences. Bryologist 2006, 109, 43–59. [Google Scholar] [CrossRef]
- Balarinová, K.; Váczi, P.; Barták, M.; Hazdrová, J.; Forbelská, M. Temperature-dependent growth rate and photosynthetic performance of Antarctic symbiotic alga Trebouxia sp. cultivated in a bioreactor. Czech Polar Rep. 2013, 3, 19–27. [Google Scholar] [CrossRef] [Green Version]
- Mohamed, E.; Mohamed, L. Using calcicolous and corticolous lichens to assess lead and cadmium air pollution of the Moroccan Atlantic Coast Safi-Essaouira. Pol. J. Environ. Stud. 2019, 29, 779–787. [Google Scholar] [CrossRef]
- Vingiani, G.M.; Gasulla, F.; Barón-Sola, Á.; Sobrino-Plata, J.; Henández, L.E.; Casano, L.M. Physiological and molecular alterations of phycobionts of genus Trebouxia and Coccomyxa exposed to cadmium. Microb. Ecol. 2021, 82, 334–343. [Google Scholar] [CrossRef] [PubMed]
- Xavier, W.D.; Castoldi, G.; Cavalcante, T.J.; Rodrigues, C.R.; Trindade, P.R.; Luiz, I.A.; Damin, V. Portable chlorophyll meter for indirect evaluation of photosynthetic pigments and nitrogen content in sweet Sorghum. Sugar Tech. 2021, 23, 560–570. [Google Scholar] [CrossRef]
- Ruiz-Espinoza, F.H.; Murillo-Amador, B.; Garcia-Hernandez, J.L.; Fenech-Larios, L.; Rueda-Puente, E.O.; Troyo-Dieguez, E.; Beltran-Morales, A. Field evaluation of the relationship between chlorophyll content in basil leaves and a portable chlorophyll meter (SPAD-502) readings. J. Plant Nutr. 2010, 33, 423–438. [Google Scholar] [CrossRef]
- Rascher, U.; Liebig, M.; Lüttge, U. Evaluation of instant light-response curves of chlorophyll fluorescence parameters obtained with a portable chlorophyll fluorometer on site in the field. Plant Cell Environ. 2000, 23, 1397–1405. [Google Scholar] [CrossRef]
- Akyiğit, F.; Demir, M.; Öztürk, Ş.; Oran, S. Changes of photosynthetic pigment content in lichens collected from urban and rural localities in Bursa province. J. Biol. Environ. Sci. 2019, 13, 65–69. [Google Scholar]
- Beckett, P.J. Lichens: Sensitive indicators of improving air quality. In Restoration and Recovery of an Industrial Region; Gunn, J.M., Ed.; Springer: New York, NY, USA, 1995; pp. 81–91. [Google Scholar] [CrossRef]
- Ferreira, R.L.; Olivati, F.N. A utilização de bioindicadores como instrumento de perícia ambiental. Meio Ambiente Sustent. 2014, 5, 3. [Google Scholar]
- Rodrigues, D.A.; de Fátima Sales, J.; Vasconcelos Filho, S.C.; Rodrigues, A.A.; Teles, E.M.G.; Costa, A.C.; Reis, E.L.; Andrade de Carvalho Silva, T.; Müller, C. Bioindicator potential of Ricinus communis to simulated rainfall containing potassium fluoride. PeerJ 2020, 8, e9445. [Google Scholar] [CrossRef]
- Zetsche, E.M.; Meysman, F.J. Dead or alive? Viability assessment of micro-and mesoplankton. J. Plankton Res. 2012, 34, 493–509. [Google Scholar] [CrossRef]
- Port, R.K.; Käffer, M.I.; Schmitt, J.L. Morphophysiological variation and metal concentration in the thallus of Parmotrema tinctorum (Despr. ex Nyl.) Hale between urban and forest areas in the subtropical region of Brazil. Environ. Sci. Pollut. Res. 2018, 25, 33667–33677. [Google Scholar] [CrossRef] [PubMed]
- Käffer, M.I.; Dantas, R.V.; Martins, S.M.A. Characterization of the epiphytic lichen vegetation in a riparian forest in southern Brazil. Plant Ecol. Evol. 2016, 149, 92–100. [Google Scholar] [CrossRef]
- Souza, T.C.; Castro, E.M.; Magalhães, C.P.; Lino, L.O.; Alves, E.T.; Albuquerque, P.E.P. Morphophysiology, morphoanatomy, and grain yield under field conditions for two maize hybrids with contrasting response to drought stress. Acta Physiol. Plant. 2013, 35, 1007–1738. [Google Scholar] [CrossRef] [Green Version]
- Brandão, J.R.; Ribeiro, D.S.; Bernardino Filho, D.C.A.; Vieira, J.R. Adequação do teste de condutividade elétrica para determinar a qualidade fisiológica de sementes. Inf. Abrates. 1997, 7, 184. [Google Scholar]
- O’brien, T.P.; Mccully, M.E. The Study of Plant Structure: Principles and Selected Methods; Termarcarphy Pry Ltd.: Melbourne, Australia, 1981. [Google Scholar]
- Wellburn, A.R. The spectral determination of chlorophylls a and b, as well as total carotenoids, using various solvents with spectrophotometers of different resolutions. J. Plant Physiol. 1994, 144, 307–313. [Google Scholar] [CrossRef]
- Ronen, R.; Galun, M. Pigment extraction from lichens with dimethyl sulphoxide (DMSO) and estimation of chlorophyll degradation. Environ. Exp. Bot. 1984, 24, 239–245. [Google Scholar] [CrossRef]
- Strasser, R.J.; Srivastava, A.; Tsimilli-Michael, M. The fluorescence transient as a tool to characterize and screen photosynthetic samples. In Probing Photosynthesis: Mechanism, Regulation and Adaptation; Yunus, M., Pathre, U., Mohanty, P., Eds.; Taylor and Francis: New York, NY, USA, 2000; pp. 445–483. [Google Scholar]
- Biemelt, S.; Keetman, U.; Albrecht, G. Re-aeration following hypoxia or anoxia leads to activation of the antioxidative defense system in roots of wheat seedlings. Plant Physiol. 1998, 116, 651–658. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Havir, E.A.; Mchale, N.A. Biochemical and developmental characterization of multiple forms of catalase in tobacco leaves. Plant Physiol. 1987, 84, 450–455. [Google Scholar] [CrossRef] [Green Version]
- Nakano, Y.; Asada, K. Hydrogen peroxide is scavenged by ascorbate-specific peroxidase in spinach chloroplasts. Plant Cell Physiol. 1981, 22, 867–880. [Google Scholar] [CrossRef]
- Giannopolitis, C.N.; Ries, S.K. Superoxide dismutases: I. Occurrence in higher plants. Plant Physiol. 1977, 59, 309–314. [Google Scholar] [CrossRef] [PubMed]
- Velikova, V.; Yordanov, I.; Edreva, A. Oxidative stress and some antioxidant systems in acid rain-treated bean plants: Protective role of exogenous polyamines. Plant Sci. 2000, 151, 56–66. [Google Scholar] [CrossRef]
- Buege, J.A.; Aust, S.D. Microsomal lipid peroxidation. In Methods in enzymology; Qin, P.Z., Ed.; Academic Press: New York, NY, USA, 1978. [Google Scholar] [CrossRef]
- R Core Team. R: A Language and Environment for Statistical Computing. R Foundation for Statistical Computing, Vienna, Austria. 2021. Available online: https://www.R-project.org/ (accessed on 20 February 2022).
- La Rocca, N.; Andreoli, C.; Giacometti, G.Á.; Rascio, N.; Moro, I. Responses of the Antarctic microalga Koliella antarctica (Trebouxiophyceae, Chlorophyta) to cadmium contamination. Photosynthetica 2009, 47, 471–479. [Google Scholar] [CrossRef]
- Di Toppi, L.S.; Pawlik-Skowrońska, B.; Vurro, E.; Vattuone, Z.; Kalinowska, R.; Restivo, F.M.; Skowroński, T. First and second line mechanisms of cadmium detoxification in the lichen photobiont Trebouxia impressa (Chlorophyta). Environ. Pollut. 2008, 151, 280–286. [Google Scholar] [CrossRef]
- Rihab, B.A.; Sabrine, B.O.; Lina, C.; Imed, M.; Hatem, B.O.; Ali, O. Cadmium effect on physiological responses of the tolerant Chlorophyta specie Picocystis sp. isolated from Tunisian wastewaters. Environ. Sci. Pollut. Res. 2017, 24, 1803–1810. [Google Scholar] [CrossRef] [PubMed]
- El Rasafi, T.; Oukarroum, A.; Haddioui, A.; Song, H.; Kwon, E.E.; Bolan, N.; Rinklebe, J. Cadmium stress in plants: A critical review of the effects, mechanisms, and tolerance strategies. Crit. Rev. Environ. Sci. Technol. 2022, 52, 675–726. [Google Scholar] [CrossRef]
- Anjum, S.A.; Tanveer, M.; Hussain, S.; Bao, M.; Wang, L.; Khan, I.; Shahzad, B. Cadmium toxicity in Maize (Zea mays L.): Consequences on antioxidative systems, reactive oxygen species and cadmium accumulation. Environ. Sci. Pollut. Res. 2015, 22, 17022–17030. [Google Scholar] [CrossRef]
- Skórzyńska-Polit, E.; Drążkiewicz, M.; Krupa, Z. Lipid peroxidation and antioxidative response in Arabidopsis thaliana exposed to cadmium and copper. Acta Physiol. Plant. 2010, 32, 169–175. [Google Scholar] [CrossRef]
- Sujetovienė, G.; Česynaitė, J. Assessment of air pollution at the indoor environment of a shooting range using lichens as biomonitors. J. Toxicol. Environ. Health Part A 2021, 84, 273–278. [Google Scholar] [CrossRef]
- Bora, M.S.; Sarma, K.P. Anatomical and ultrastructural alterations in Ceratopteris pteridoides under cadmium stress: A mechanism of cadmium tolerance. Ecotoxicol. Environ. Saf. 2021, 218, 112285. [Google Scholar] [CrossRef]
- Pawlik-Skowrońska, B.; Di Toppi, L.S.; Favali, M.A.; Fossati, F.; Pirszel, J.; Skowroński, T. Lichens respond to heavy metals by phytochelatin synthesis. New Phytol. 2002, 156, 95–102. [Google Scholar] [CrossRef]
- Daimari, R.; Bhuyan, P.; Hussain, S.; Nayaka, S.; Mazumder, M.A.; Hoque, R.R. Anatomical, physiological, and chemical alterations in lichen (Parmotrema tinctorum (Nyl.) Hale) transplants due to air pollution in two cities of Brahmaputra Valley, India. Environ. Monit. Assess. 2021, 193, 101. [Google Scholar] [CrossRef] [PubMed]
- Tyler, G. Uptake, retention and toxicity of heavy metals in lichens. A brief review. Water Air. Soil. Poll. 1989, 47, 321–333. [Google Scholar] [CrossRef]
- Aprile, G.G.; Catalano, I.; Migliozzi, A.; Mingo, A. Monitoring epiphytic lichen biodiversity to detect environmental quality and air pollution: The case study of Roccamonfina Park (Campania Region—Italy). In Air Pollution--New Developments; Moldoveanu, A.M., Ed.; In Tech Publisher: Rijeka, Croatia, 2011; pp. 227–244. [Google Scholar] [CrossRef] [Green Version]
- Solhaug, K.A.; Gauslaa, Y. Photosynthates stimulate the UV-B induced fungal anthraquinone synthesis in the foliose lichen Xanthoria parietina. Plant Cell Environ. 2004, 27, 167–176. [Google Scholar] [CrossRef]
- Solhaug, K.A.; Gauslaa, Y.N.; Line, B.W. UV-induction of sun-screening pigments in lichens. New Phytol. 2003, 158, 91–100. [Google Scholar] [CrossRef]
- Bačkor, M.; Fahselt, D.; Davidson, R.D.; Wu, C.T. Effects of copper on wild and tolerant strains of the lichen photobiont Trebouxia erici (Chlorophyta) and possible tolerance mechanisms. Arch. Environ. Contam. Toxicol. 2003, 45, 159–167. [Google Scholar] [CrossRef] [PubMed]
- Kalinowska, R.; Bačkor, M.; Pawlik-Skowrońska, B. Parietin in the tolerant lichen Xanthoria parietina (L.) Th. Fr. increases protection of Trebouxia photobionts from cadmium excess. Ecol. Indic. 2015, 58, 132–138. [Google Scholar] [CrossRef]
- Sujetovienė, G.; Šliumpaitė, I. Effects of cadmium on physiological parameters of the lichen Evernia prunastri and Ramalina fastigiata. E3S Web Conf. 2013, 1, 29007. [Google Scholar] [CrossRef] [Green Version]
- Rustichelli, C.; Visioli, G.; Kostecka, D.; Vurro, E.; di Toppi, L.S.; Marmiroli, N. Proteomic analysis in the lichen Physcia adscendens exposed to cadmium stress. Environ. Pollut. 2008, 156, 1121–1127. [Google Scholar] [CrossRef]
- Bačkor, M.; Kováčik, J.; Dzubaj, A.; Bačkorová, M. Physiological comparison of copper toxicity in the lichens Peltigera rufescens (Weis) Humb. and Cladina arbuscula subsp. mitis (Sandst.) Ruoss. J. Plant Growth Regul. 2009, 58, 279–286. [Google Scholar] [CrossRef] [Green Version]
- Vantová, I.; Bačkor, M.; Klejdus, B.; Bačkorová, M.; Kováčik, J. Copper uptake and copper-induced physiological changes in the epiphytic lichen Evernia prunastri. Plant Growth Regul. 2013, 6, 1–9. [Google Scholar] [CrossRef]
- Rola, K.; Latkowska, E.; Myśliwa-Kurdziel, B.; Osyczka, P. Heavy-metal tolerance of photobiont in pioneer lichens inhabiting heavily polluted sites. Sci. Total Environ. 2019, 679, 260–269. [Google Scholar] [CrossRef]
- Garty, J.; Weissman, L.; Tamir, O.; Beer, S.; Cohen, Y.; Karnieli, A.; Orlovsky, L. Comparison of five physiological parameters to assess the vitality of the lichen Ramalina lacera exposed to air pollution. Physiol. Plant. 2000, 109, 410–418. [Google Scholar] [CrossRef] [Green Version]
- Dobroviczká, T.; Piršelová, B.; Meszaros, P.; Blehová, A.; Libantova, J.; Moravčiková, J.; Matušíková, I. Effects of cadmium and arsenic ions on content of photosynthetic pigments in the leaves of Glycine max (L.) Merrill. Pak. J. Bot. 2013, 45, 105–110. [Google Scholar]
- Young, A.J.; Britton, G. Carotenoids and oxidative stress. In Current Research in Photosynthesis; Baltscheffsky, M., Ed.; Kluwer: Dordrecht, The Netherlands, 1990; pp. 587–590. [Google Scholar]
- Bačkor, M.; Kováčik, J.; Piovár, J.; Pisani, T.; Loppi, S. Physiological aspects of cadmium and nickel toxicity in the lichens Peltigera rufescens and Cladina arbuscula subsp. mitis. Water Air Soil Pollut. 2010, 207, 253–262. [Google Scholar] [CrossRef]
- Carmo Araújo, S.A.; Deminicis, B.B. Fotoinibição da fotossíntese. Rev. Bras. Biociênc. 2009, 7, 4. [Google Scholar]
- Öquist, G.; Chow, W.S.; Anderson, J.M. Photoinhibition of photosynthesis represents a mechanism for the long-term regulation of photosystem II. Planta 1992, 186, 450–460. [Google Scholar] [CrossRef] [PubMed]
- Sunil, B.; Saini, D.; Bapatla, R.B.; Aswani, V.; Raghavendra, A.S. Photorespiration is complemented by cyclic electron flow and the alternative oxidase pathway to optimize photosynthesis and protect against abiotic stress. Photosynth. Res. 2019, 139, 67–79. [Google Scholar] [CrossRef]
- Sharma, S.; Joshi, J.; Kataria, S.; Verma, S.K.; Chatterjee, S.; Jain, M.; Brestic, M. Regulation of the Calvin cycle under abiotic stresses: An overview. Plant Life Chang. Environ. 2020, 1, 681–717. [Google Scholar] [CrossRef]
- Schroeter, B. In situ photosynthetic differentiation of the green algal and the cyanobacterial photobiont in the crustose lichen Placopsis contortuplicata. Oecologia 1994, 98, 212–220. [Google Scholar] [CrossRef]
- Maslać, A.; Maslać, M.; Tkalec, M. The impact of cadmium on photosynthetic performance and secondary metabolites in the lichens Parmelia sulcata, Flavoparmelia caperata and Evernia prunastri. Acta Bot. Croat. 2016, 75, 186–193. [Google Scholar] [CrossRef] [Green Version]
- Cuny, D.; Van Haluwyn, C.; Shirali, P.; Zerimech, F.; Jérôme, L.; Haguenoer, J.M. Cellular impact of metal trace elements in terricolous lichen Diploschistes muscorum (Scop.) R. Sant.–identification of oxidative stress biomarkers. Water Air Soil Pollut. 2004, 152, 55–69. [Google Scholar] [CrossRef]
- Deltoro, V.I.; Gimeno, C.; Calatayud, A.; Barreno, E. Effects of SO2 fumigations on photosynthetic CO2 gas exchange, chlorophyll a fluorescence emission and antioxidant enzymes in the lichens Evernia prunastri and Ramalina farinacea. Physiol. Plant. 1999, 105, 648–654. [Google Scholar] [CrossRef]
- Silberstein, L.; Siegel, B.Z.; Siegel, S.M.; Mukhtar, A.; Galun, M. Comparative studies on Xanthoria parietina, a pollution-resistant lichen, and Ramalina duriaei, a sensitive species. II. Effects of possible air pollution protection mechanisms. Lichenologist 1996, 28, 367–383. [Google Scholar] [CrossRef]
- Weissman, L.; Fraiberg, M.; Shine, L.; Garty, J.; Hochman, A. Responses of antioxidants in the lichen Ramalina lacera may serve as an early-warning bioindicator system for the detection of air pollution stress. FEMS Microbiol. Ecol. 2006, 58, 41–53. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Panchal, F.H.; Ray, S.; Munshi, R.P.; Bhalerao, S.S.; Nayak, C.S. Alterations in lipid metabolism and antioxidant status in lichen planus. Indian J. Dermatol. 2015, 60, 439. [Google Scholar] [PubMed]
- Caverzan, A.; Passaia, G.; Rosa, S.B.; Ribeiro, C.W.; Lazzarotto, F.; Margis-Pinheiro, M. Plant responses to stresses: Role of ascorbate peroxidase in the antioxidant protection. Genet. Mol. Biol. 2012, 35, 1011–1019. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Devasagayam, T.P.A.; Boloor, K.K.; Ramasarma, T. Methods for estimating lipid peroxidation: An analysis of merits and demerits. Indian J. Biochem. Biophys. 2003, 40, 300–308. [Google Scholar]
- Araújo, A.A.S.; Melo, M.G.D.; Rabelo, T.K.; Nunes, P.S.; Santos, S.L.; Serafini, M.R.; Gelain, D.P. Review of the biological properties and toxicity of usnic acid. Nat. Prod. Res. 2015, 29, 2167–2180. [Google Scholar] [CrossRef]
- Oran, S.; Sahin, S.; Sahinturk, P.; Ozturk, S.; Demir, C. Antioxidant and antimicrobial potential, and HPLC analysis of stictic and usnic acids of three Usnea species from Uludag mountain (Bursa, Turkey). Iran. J. Pharm. Res. 2016, 15, 527. [Google Scholar]
- Cakmak, K.C.; Gülçin, İ. Anticholinergic and antioxidant activities of usnic acid-an activity-structure insight. Toxicol. Rep. 2019, 6, 1273–1280. [Google Scholar] [CrossRef]
- Santos, R.K.; Gomes, N.C.; Assis, O.G.; Silva, J.J.R.; Alvarenga, C.A.; Belardi, R.M. Lichens used as bioindicator of air quality in mining town of Itabira. Res. Soc. Dev. 2018, 7, e4712480. [Google Scholar] [CrossRef]
- Raven, P.H.; Evert, R.F.; Eichhorn, S.E. Biologia Vegetal; Editora Guanabara Koogan: Rio de Janeiro, BR, Brazil, 2007. [Google Scholar]
- Martins-Mazzitelli, S.M.A.; Mota Filho, F.O.; Pereira, E.C.; Figueira, R. Utilização de liquens no biomonitoramento da qualidade do ar. In Biologia de Liquens; Xavier Filho, L., Legaz, M.E., Córdoba, C.V., Pereira, E.C., Eds.; Âmbito Cultural: Rio de Janeiro, BR, Brazil, 2006; pp. 101–133. [Google Scholar]
- Le Blanc, F.S.C. Possibilities and methods for mapping air pollution on the basis of lichen sensitivity. Vienna Forstl Bundes Vers. Mariabrunn Mitt 1971, 92, 103–126. [Google Scholar]
- Barnes, J.D.; Balaguer, L.; Manrique, E.; Elvira, S.; Davison, A.W. A reappraisal of the use of DMSO for the extraction and determination of chlorophylls a and b in lichens and higher plants. Environ. Exp. Bot. 1992, 32, 85–100. [Google Scholar] [CrossRef]
- Sepahvand, A.; Studzińska-Sroka, E.; Ramak, P.; Karimian, V. Usnea sp.: Antimicrobial potential, bioactive compounds, ethnopharmacological uses and other pharmacological properties; a review article. J. Ethnopharmacol. 2021, 268, 113656. [Google Scholar] [CrossRef] [PubMed]
- Grill, E.; Löffler, S.; Winnacker, E.L.; Zenk, M.H. Phytochelatins, the heavy-metal-binding peptides of plants, are synthesized from glutathione by a specific γ-glutamylcysteine dipeptidyl transpeptidase (phytochelatin synthase). Proc. Natl. Acad. Sci. USA 1989, 86, 6838–6842. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ingolfsdottir, K. Usnic acid. Phytochemistry 2002, 61, 729–736. [Google Scholar] [CrossRef]
- Popovici, V.; Bucur, L.; Popescu, A.; Caraiane, A.; Badea, V. Determination of the content in usnic acid and polyphenols from the extracts of Usnea barbata L. and the evaluation of their antioxidant activity. Farmacia 2018, 66, 337–341. [Google Scholar]
- Rodriguez, J.H.; Carreras, H.A.; Pignata, M.L.; Gonzalez, C.M. Nickel exposure enhances the susceptibility of lichens Usnea amblyoclada and Ramalina celastri to urban atmospheric pollutants. Arch. Environ. Contam. Toxicol. 2007, 53, 533–540. [Google Scholar] [CrossRef]
- Abeyrathne, E.D.N.S.; Nam, K.; Ahn, D.U. Analytical methods for lipid oxidation and antioxidant capacity in food systems. Antioxidants 2021, 10, 1587. [Google Scholar] [CrossRef]
- Streit, N.M.; Canterle, L.P.; Canto, M.W.D.; Hecktheuer, L.H.H. The chlorophylls. Cienc. Rural. 2005, 35, 748–755. [Google Scholar] [CrossRef]
- Karakaş, V.E.; Öztürk, Ş.; Oran, S. Comparison of photosynthetic pigment contents in lichen samples were collected from different localities in Bursa. J. Biol. Environ. Sci. 2017, 11, 121–127. [Google Scholar]
- Lucadamo, L.; Corapi, A.; Loppi, S.; Paoli, L.; Gallo, L. Spatial variation of eco-physiological parameters in the lichen Pseudevernia furfuracea transplanted in an area surrounding a cement plant (S Italy). Environ. Monit. Assess. 2015, 187, 500. [Google Scholar] [CrossRef]













Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Santos, A.M.d.; Vitorino, L.C.; Cruvinel, B.G.; Ávila, R.G.; Vasconcelos Filho, S.d.C.; Batista, P.F.; Bessa, L.A. Impacts of Cd Pollution on the Vitality, Anatomy and Physiology of Two Morphologically Different Lichen Species of the Genera Parmotrema and Usnea, Evaluated under Experimental Conditions. Diversity 2022, 14, 926. https://doi.org/10.3390/d14110926
Santos AMd, Vitorino LC, Cruvinel BG, Ávila RG, Vasconcelos Filho SdC, Batista PF, Bessa LA. Impacts of Cd Pollution on the Vitality, Anatomy and Physiology of Two Morphologically Different Lichen Species of the Genera Parmotrema and Usnea, Evaluated under Experimental Conditions. Diversity. 2022; 14(11):926. https://doi.org/10.3390/d14110926
Chicago/Turabian StyleSantos, Alex Marcelino dos, Luciana Cristina Vitorino, Bárbara Gonçalves Cruvinel, Roniel Geraldo Ávila, Sebastião de Carvalho Vasconcelos Filho, Priscila Ferreira Batista, and Layara Alexandre Bessa. 2022. "Impacts of Cd Pollution on the Vitality, Anatomy and Physiology of Two Morphologically Different Lichen Species of the Genera Parmotrema and Usnea, Evaluated under Experimental Conditions" Diversity 14, no. 11: 926. https://doi.org/10.3390/d14110926
APA StyleSantos, A. M. d., Vitorino, L. C., Cruvinel, B. G., Ávila, R. G., Vasconcelos Filho, S. d. C., Batista, P. F., & Bessa, L. A. (2022). Impacts of Cd Pollution on the Vitality, Anatomy and Physiology of Two Morphologically Different Lichen Species of the Genera Parmotrema and Usnea, Evaluated under Experimental Conditions. Diversity, 14(11), 926. https://doi.org/10.3390/d14110926