
The Effects of Sampling Depth on Benthic Testate Amoeba Assemblages in Freshwater Lakes: A Case Study in Lake Valdayskoe (the East European Plain)



Abstract
1. Introduction
2. Materials and Methods
2.1. Study Site
2.2. Field Sampling
2.3. Testatea Amoeba Analysis
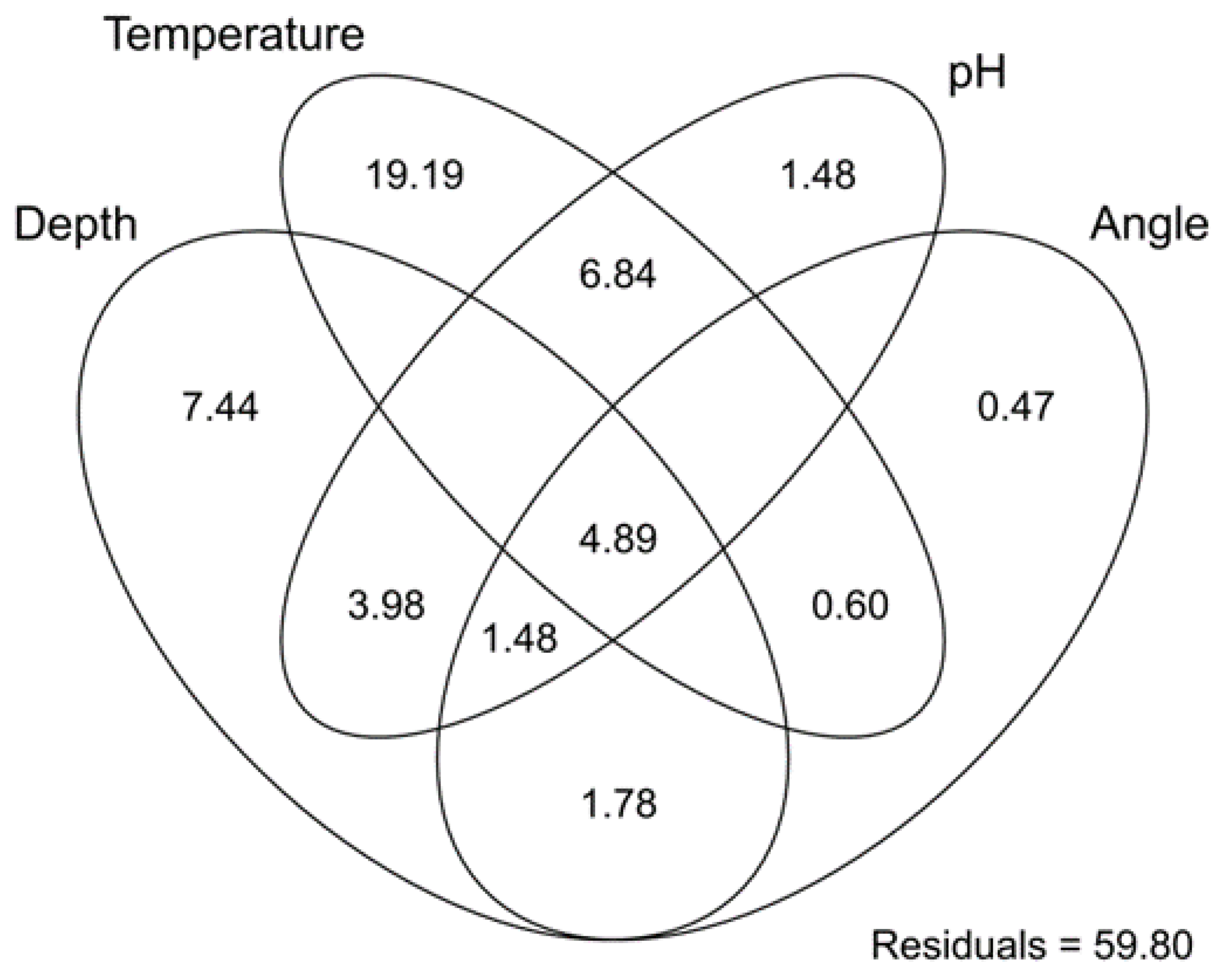
2.4. Data Analysis
3. Results
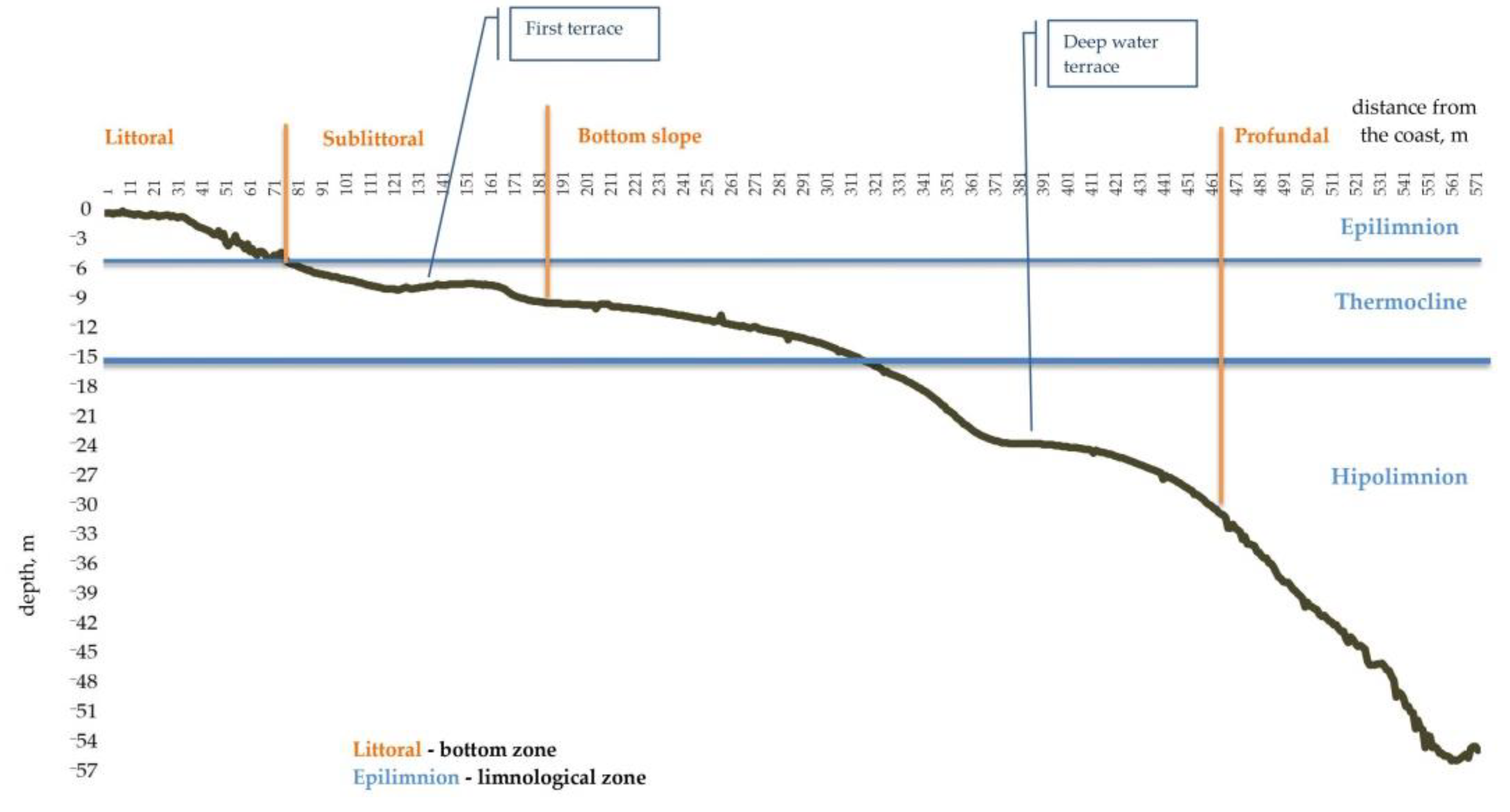
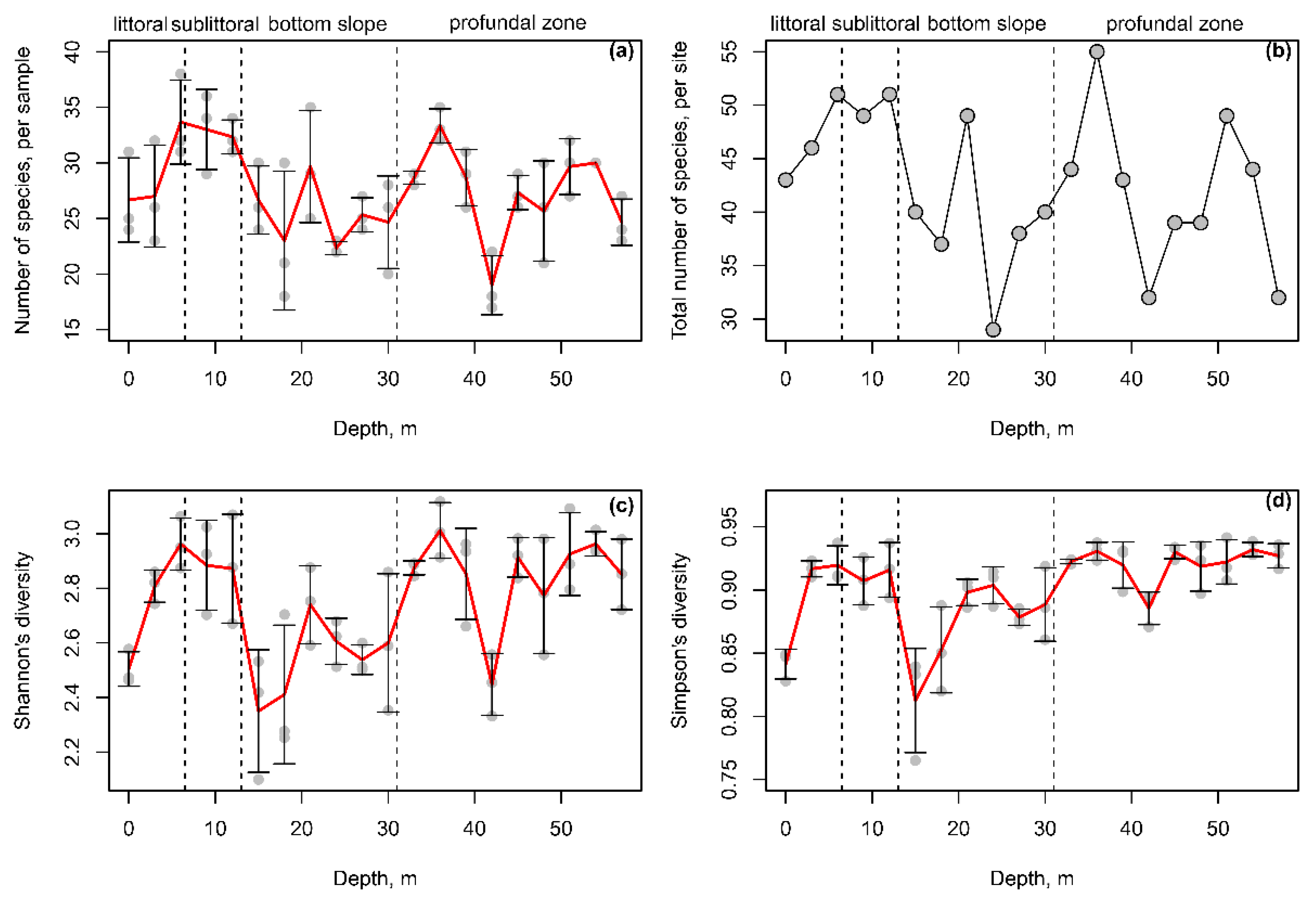
3.1. Characteristics of the Transect
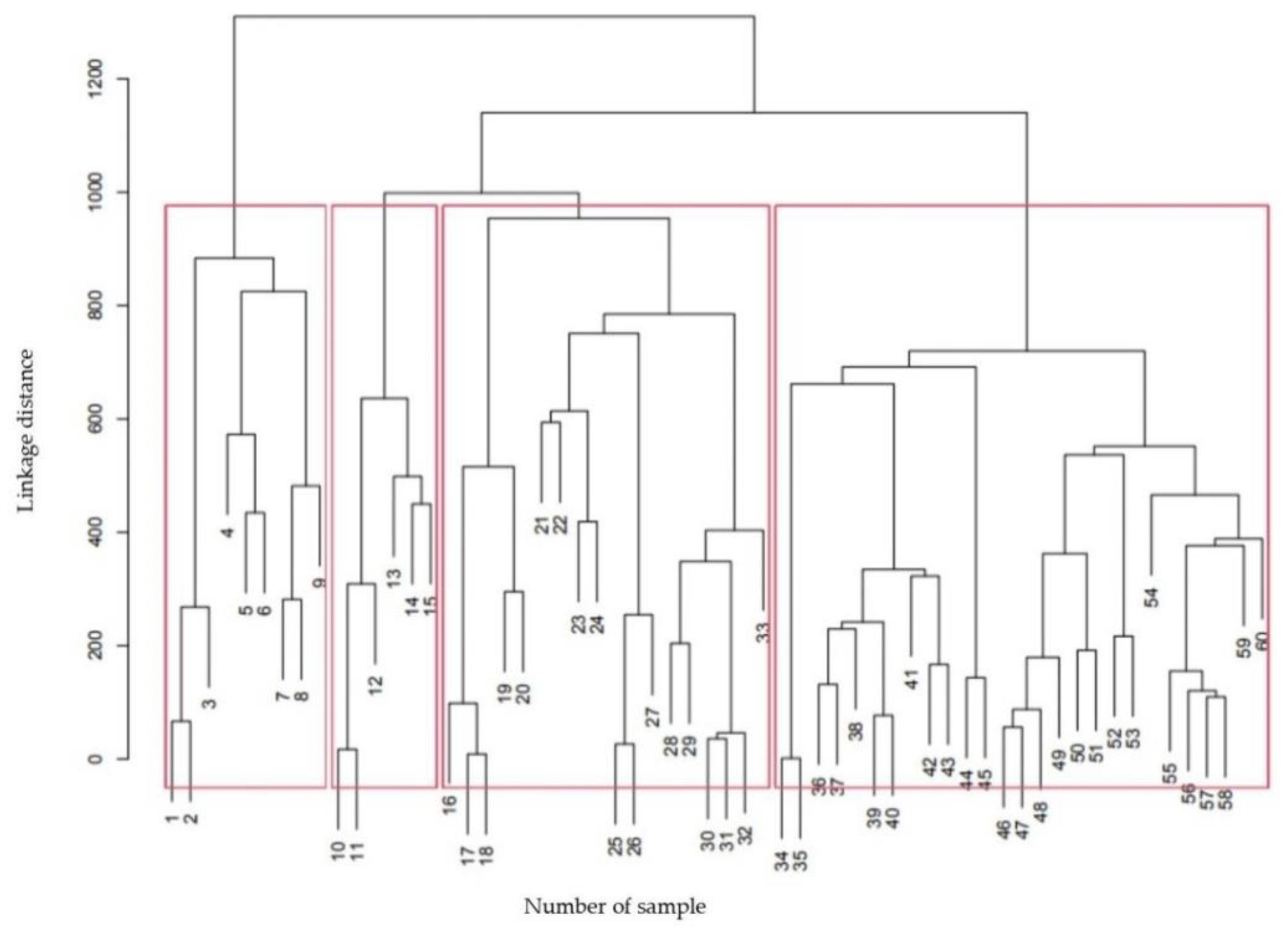
3.2. General Characteristics of Testate Amoeba Assemblages
3.3. Variation in Species Structure of Testate Amoeba Assemblages along the Depth Gradient
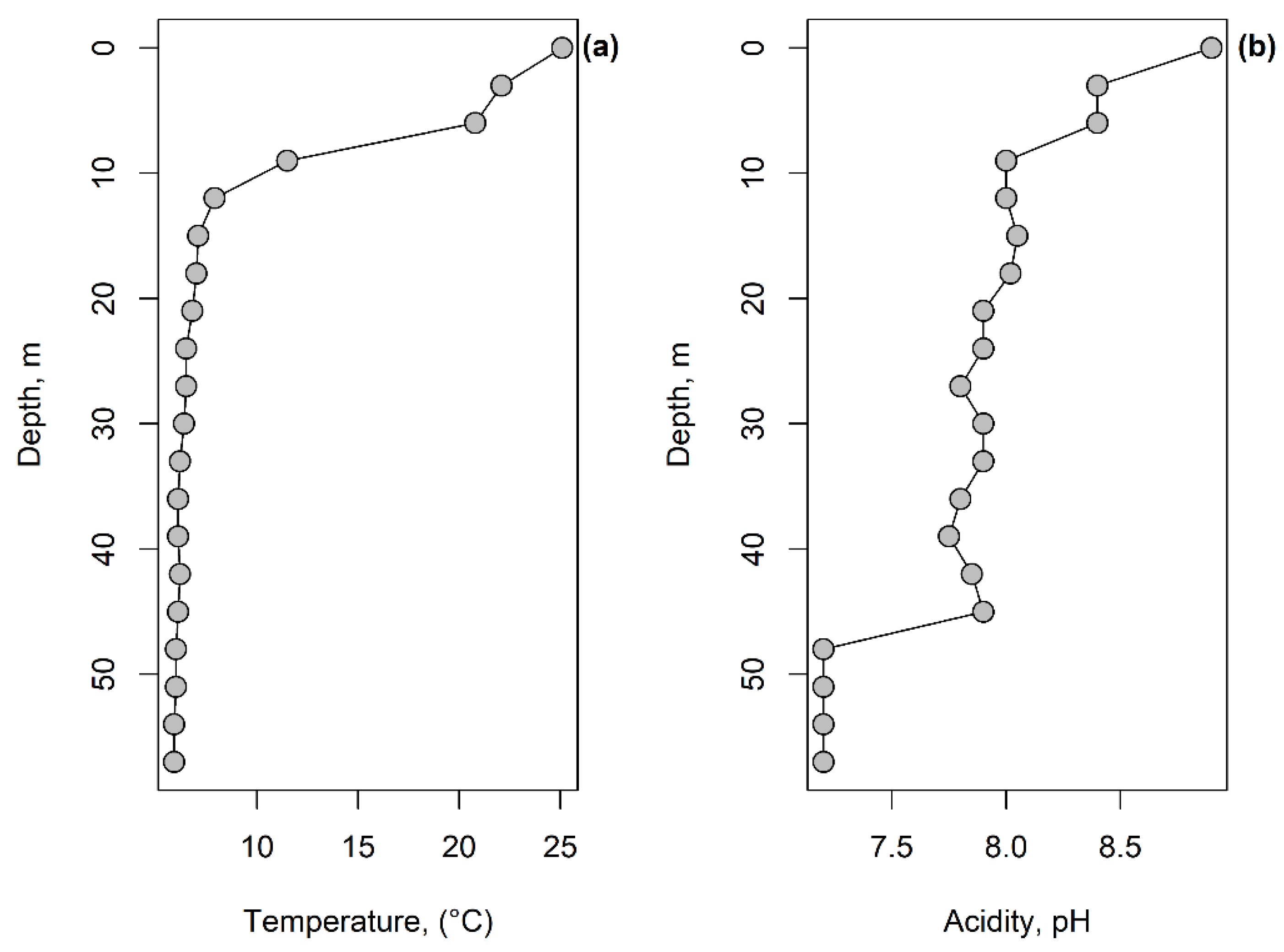
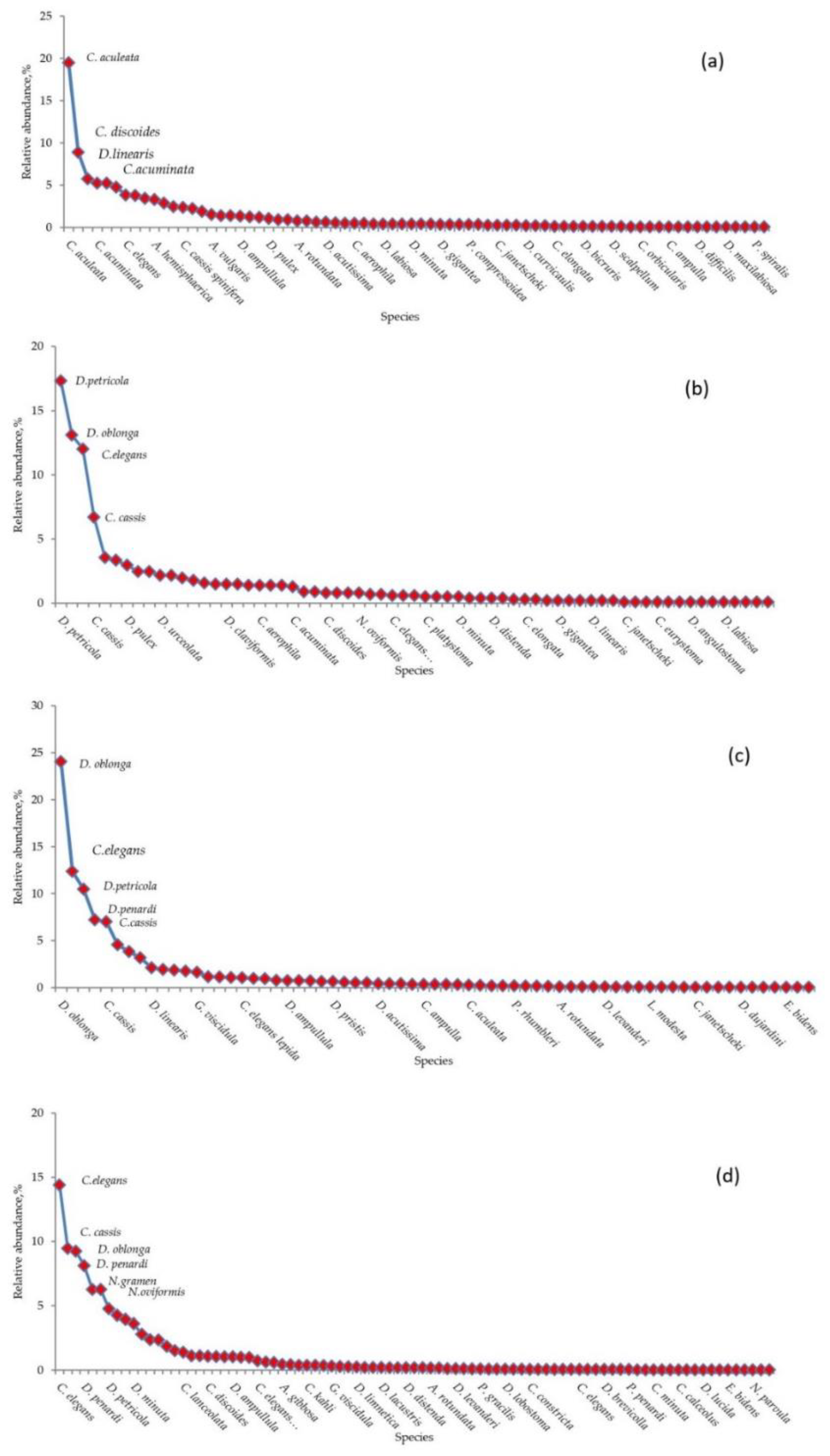
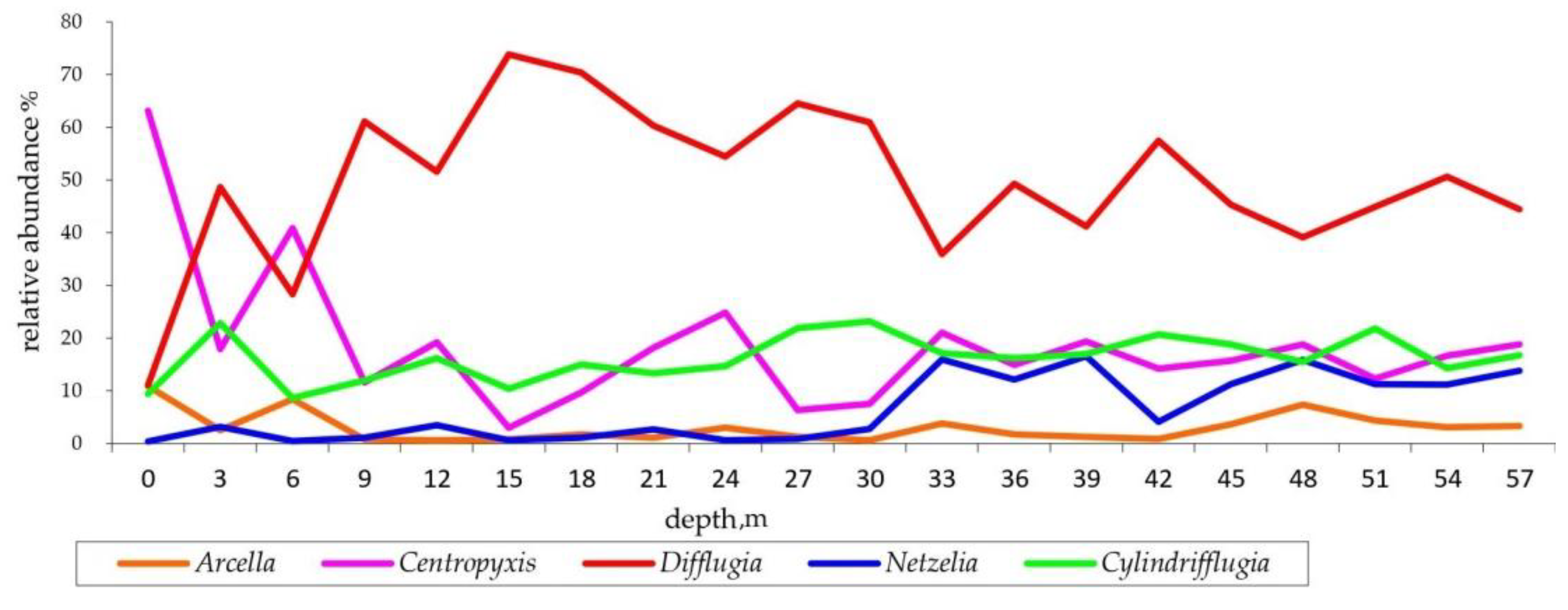
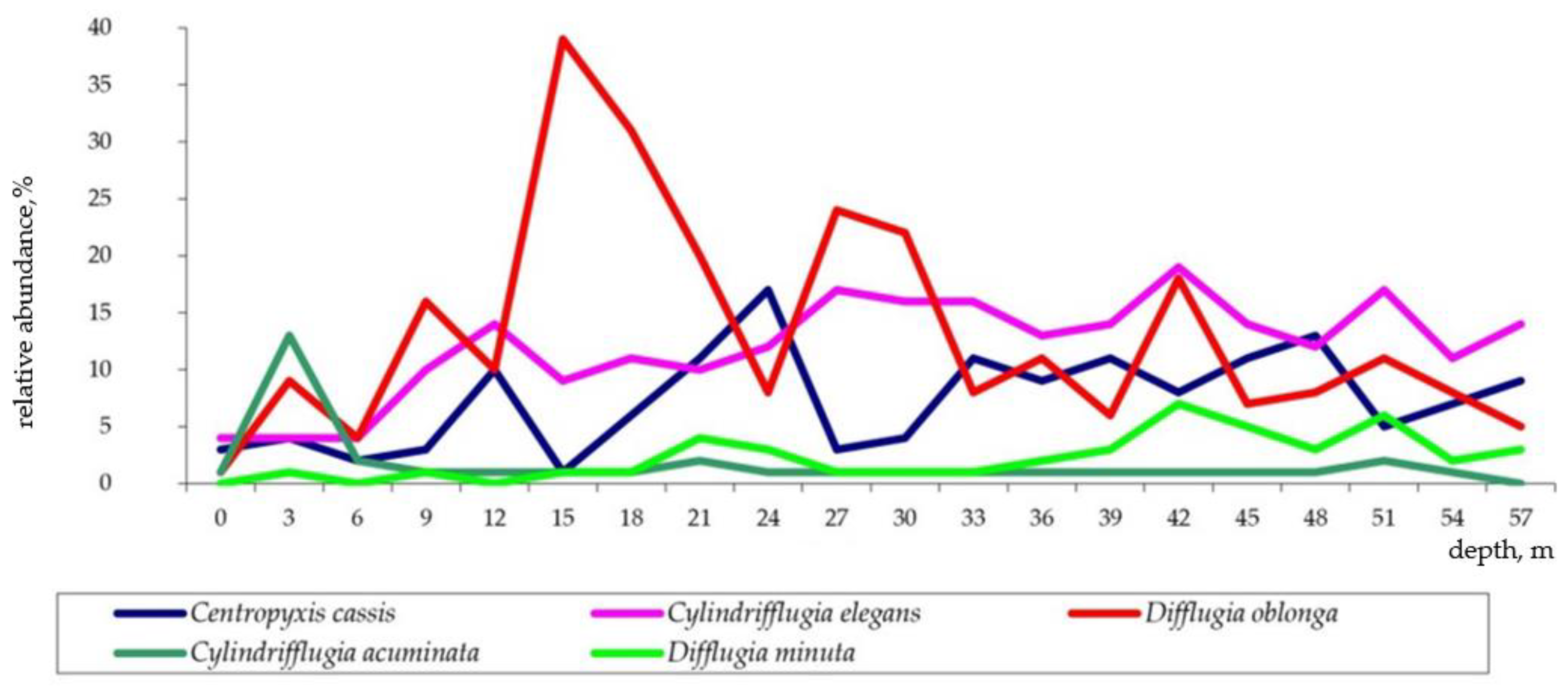
- Littoral assemblage (LA) (depth 0–6 m). Environment conditions: temperature 25.1–20.8 °C, pH 8.9–8.4, macrophytes and mollusk shells on the bottom, sand and plant residues in the surface sediments (Table S1). At these depths, 74 species belonging to 14 genera were found (Table S2). The most abundant genera were Centropyxis (41% herein and after the total shells counts in the zone), Difflugia (29%), Cylindrifflugia (13.6%), Arcella (7.4%) and Galeripora [41] (4.5%) (Table S4). The species with the maximum indicator value for the assemblage were Centropixys discoides, C. aculeata, Galeripora discoides, Cylindrifflugia acuminata, and Difflugia lacustris (Table 2). An analysis of the structure of the LA showed the presence of one dominant species Centropyxis aculeata (19%, hereafter the relative abundance in the community), and three subdominants: C. discoides (9%), Difflugia linearis (6%), and Cylindrifflugia acuminata (5.3%); the proportion of the other species did not exceed 5% (Figure 5a). Shannon’s diversity index was 3.24.
- Sublittoral assemblage (SA) (depth 9–12 m). Environmental conditions: temperature 7.9 °C, pH 8, presence of plant residues in the surface sediments. The SA consists of 65 species belonging to 11 genera (Table S2). The SA was placed in the thermocline. Genera Difflugia (56.5%), Centropyxis (15.3%), and Cylindrifflugia (14%) were characterized by the greatest relative abundance (Table S4). The species with the maximum indicator value in the SA were Pontigulasia rhumbleri, Difflugia claviformis, D. urceolata, Golemanskia viscidula [41], D. petricola, and Zivcovicia spectabilis (Table 2. The SA was dominated by species Difflugia petricola (17.3%) with three subdominant species D. oblonga (13.1%), Cylindrifflugia elegans (12%), and Centropyxis cassis (6.7%). The proportion of other species did not exceed 4% (Figure 5b). Shannon’s diversity index was 3.26.
- Bottom slope assemblage (BA) (depth 15–30 m). Environmental conditions–temperature 7.1–6.4 °C, pH 7.9–8.0. The BA was located under the thermocline. In this zone, 67 species belonging to 17 genera were found (Table S2). Genera Difflugia (64.2%), Cylindrifflugia (16.3%), and Centropyxis (11.6%) had the highest relative abundance (Table S4). The species with the maximum indicator value in the BA were Difflugia oblonga and D. lithophila (Table 2). The community structure of the BA is characterized by one dominant D. oblonga (24%) and five subdominats Cylindrifflugia elegans (12%), D. petricola (10%), D. penardi (7%), Centropyxis cassis (7%), and D. lithophila (4.6%). The proportion of other species was 4% or less (Figure 5c). Shannon’s diversity index was 2.98.
- Profundal assemblages (PA) (depth 33–57 m). Environmental conditions: temperature 5.9–6.2 °C, pH 7.9–7.2. 87 species belonging to 20 genera were found at this depth (Table S2). Genera Difflugia (45.2%), Cylindrifflugia (17.6%), Centropyxis (16.9%), and Netzelia (12.5%) had the highest relative abundance (Table S4). Species with the maximum indicator value in the PA were Netzelia oviformis, N. gramen, Difflugia minuta, and D. pristis (Table 2). The community structure of the PA was characterized by three dominant species: Cylindrifflugia elegans (14.4%), Centropyxis cassis (9.5%), and Difflugia oblonga (9.2%) as well as five subdominants D. penardi (8.1%), Netzelia gramen (6.3%), N. oviformis (6.3%), D. petricola (4.8%), and D. linearis (4.3%) (Figure 5d). Shannon’s diversity index was 3.15.


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Assemblage | Taxa | Indicator Value | Probability |
|---|---|---|---|
| Littoral assemblage | Centropyxis discoides | 0.7522 | 0.001 |
| Galeripora discoides | 0.6857 | 0.001 | |
| Centropyxis aculeata | 0.6814 | 0.001 | |
| Cylindrifflugia acuminata | 0.5992 | 0.006 | |
| Difflugia lacustris | 0.5065 | 0.012 | |
| Arcella hemisphaerica | 0.4945 | 0.007 | |
| Cylindrifflugia lanceolata | 0.4745 | 0.013 | |
| Difflugia linearis | 0.4642 | 0.01 | |
| Centropyxis ecornis | 0.4148 | 0.017 | |
| Arcella vulgaris | 0.4074 | 0.017 | |
| Centropyxis cassis spinifera | 0.4058 | 0.04 | |
| Difflugia rubescens | 0.3865 | 0.015 | |
| Difflugia corona | 0.3616 | 0.011 | |
| Arcella rotundata | 0.3296 | 0.02 | |
| Sub-littoral assemblage | Pontigulasia rhumbleri | 0.75 | 0.001 |
| Difflugia claviformis | 0.6696 | 0.001 | |
| Difflugia urceolata | 0.5709 | 0.002 | |
| Golemanskia viscidula | 0.5294 | 0.001 | |
| Difflugia petricola | 0.5145 | 0.001 | |
| Zivkovicia spectabilis | 0.5046 | 0.005 | |
| Pontigulasia compressa | 0.477 | 0.002 | |
| Pontigulasia spiralis | 0.4091 | 0.005 | |
| Difflugia longicollis | 0.3913 | 0.027 | |
| Difflugia avellana | 0.3841 | 0.013 | |
| Difflugia sinuata | 0.3 | 0.021 | |
| Erugomicula bidens | 0.2927 | 0.02 | |
| Bottom slope assemblage | Difflugia oblonga | 0.4709 | 0.001 |
| Difflugia lithophila | 0.4018 | 0.012 | |
| Cyphoderia ampulla | 0.2917 | 0.04 | |
| Profundal assemblage | Netzelia gramen | 0.6727 | 0.001 |
| Difflugia minuta | 0.5354 | 0.001 | |
| Difflugia pristis | 0.5349 | 0.001 | |
| Difflugia penardi | 0.3925 | 0.001 | |
| Cylindrifflugia elegans | 0.3277 | 0.009 | |
| Netzelia oviformis | 0.682 | 0.001 |
3.4. Relative Abundance of Testate Amoeba Genera along the Depth Gradient
3.5. The Effect of Depth on the Species Diversity of Testate Amoebae
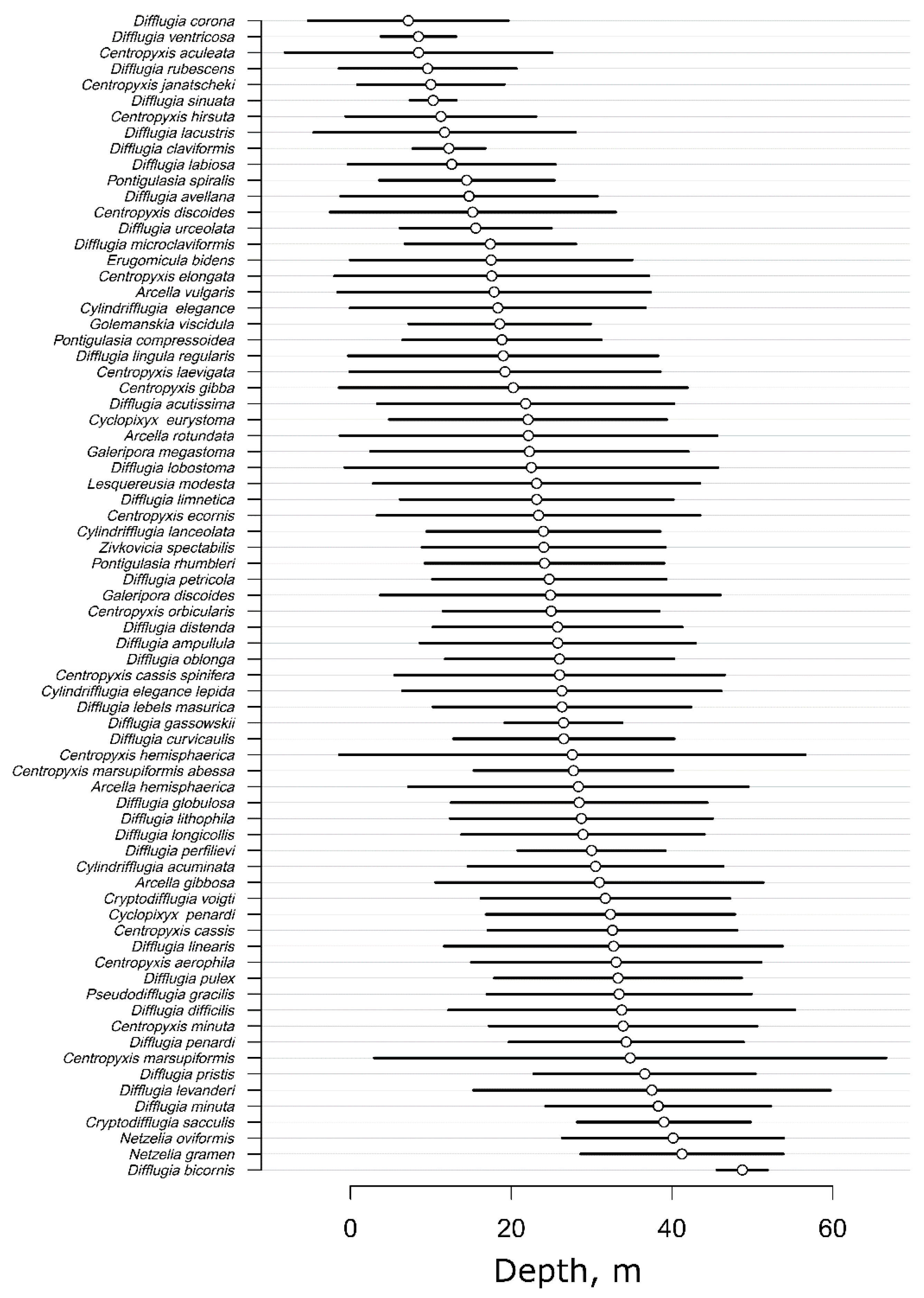
3.6. Optima and Tolerance of Testate Amoebae to Sampling Depth
4. Discussion
4.1. General Characteristics of Testate Amoeba Assemblages
4.2. Variation in Species Structure of Testate Amoeba Assemblages along the Depth Gradient
- Littoral Assemblages (LA). Our data indicate the dominance of species of the genus Centropyxis, Cylindrifflugia, and Arcella on the littoral that correlates to the observations in Ovcharitsa Reservoir [56], Smradlivo Ezero glacial lake [57], and the lake Dorian [58] in Bulgaria. In all these studies, the abundance of Centropyxis and Arcella at shallow depths was higher than at depths greater than 6 m. The high abundance of Centropyxis aculeata, C. discoides, and Galeripora discoides on the littoral corresponds well to the findings of Dabes & Velho [59], who showed the dominance of these species on the littoral in Lake São Francisco (Brazil). The confinement of Cylindrifflugia. acuminata and Arcella hemisphaerica to the conditions of the littoral were already shown in a study by Schönborn [60]. Sigala [61] reported the dominance of Centropyxis aculeata and Galeripora discoides in the sites with high oxygen content and warm water. Thus, the species structure of littoral assemblages in Lake Valdayskoe is similar to the littoral assemblages in other lakes.
- Sublittoral Assemblage (SA) considerably differs from the LA and is mainly formed by the species which were previously reported by many authors in oligo-, meso- and eutrophic lakes [49,62,63]. These authors also mention the high relative abundance of the genus Difflugia, the low relative abundance of Centropyxis, and the almost complete absence of Arcella at depths deeper than 9 m. D. oblonga was very abundant in the SA, but was not identified as indicator species for this assemblage. It may be explained by a high level of eutrophication and a thin sapropel layer in the sediments that create good conditions for the high reproduction of D. oblonga [11]. The list of identified indicator species may be divided into two groups. The first group is formed by Difflugia urceolata, D. petricola, and Pontigulasia rhumbleri which were previously observed only at depths greater than 5 m [13,57,64]. Another group of species includes Zivkovicia spectabilis, Difflugia claviformis, and Golemanskia viscidula which were previously reported in a depth range of 1.5–8 m [15,57] that also includes the shallow waters. Moreover, Torigai [65] claimed that G. viscidula depends on biotope productivity, but not on depth. Thus, the specimen structure of sublittoral assemblages consists of species earlier reported from the related or shallower depths.
- Bottom Slope Assemblage (BA) is described for the first time, so that there are no previous descriptions of assemblages of this type in the literature. Genus Difflugia dominates in BA, while genera Cylindrifflugia and Centropyxis occupy only 10–15% of the total abundance. There are three indicator species (Difflugia oblonga, D. lithophila and Cyphoderia ampulla) in this assemblage, however, Difflugia oblonga was usual in other assemblages and D. lithophila depends on high trophic status [66]. The most abundant species in the BA also have high relative abundance in sublittoral and profundal assemblages that might indicate a transitional nature of BA.
- Profundal assemblage (PA) was characterized by the presence of Netzelia oviformis, N. gramen, Difflugia minuta, and D. pristis. These findings correspond well with the results of Todorov [57] and Davidova [24], who described similar species structures in the profundal zone and mentioned Netzelia oviformis, N. gramen, and Difflugia minuta as typical species for profundal. Davidova [24] observed Netzelia species in benthic biotopes of the Rabisha reservoir (Bulgaria) with the maximal relative abundance at the deepest sites (12–15 m). Tsyganov et al. [17] mentioned N. gramen as a species-typical for sublittoral and profundal at depths from 4.5 to 20.5 m in Shatura lakes (Moscow region, Russia). Arriera [11] described the positive correlation between Netzelia oviformis and the sampling depth. Thus, the profundal assemblage species structure is typical for deep water lacustrine habitats.
4.3. Relative Abundance of Testate Amoeba Genera along the Depth Gradient
4.4. The Effect of Depth on the Species Diversity of Testate Amoebae
4.5. Optima and Tolerance of Testate Amoebae to Sampling Depth
- Eurybathic species. D. oblonga inhabits all depths with the greatest abundance in the biotopes with high eutrophication levels that corresponds well to the findings of Kihlman and Kauppila [76], and Arriera [11], who describe the positive correlation between D. oblonga and high levels of eutrophication. This suggests that D. oblonga is an indicator of high eutrophication, but not the depth, and is able to inhabit the depth gradient from 0 to 57 m. The absence of depth dependence for Cylindiflugia elegans was noted in the works of Davidova [15,24], who found C. elegans at all depths, from phytal to profundal. Tran [77] showed that C. elegans was a typical species in all studied aquatic habitats in Vietnam regardless of depth. We propose that D. elegans is a eurytopic species. The ability of C. cassis to inhabit all the depths also has been shown by Todorov [57] and Davidova [15].
- Conditionally stenobathic species:
- a.
- Littoral species (0–6 m depth). The high abundances of Cylindriflugia acuminata, Difflugia rubescens, and Arcella hemisphaerica in the littoral correlate well with the findings of Schönborn [60] who also mentioned that these species as typical for shallow waters. Even more evidence for such ecological preferences can be found for C. acuminata [60] and D. rubescens [78], so these two species can be considered reliable indicators for littoral conditions. At the same time, our results indicate that the relative abundance of Arcella hemisphaerica slightly increased in profundal. We associate this with the intensive accumulation of the dead shells of A. hemisphaerica from plankton, where this species is also abundant. Therefore, this species should be used as an indicator of shallow waters with caution. In our study, D. linearis has maximal abundance in the littoral (the habitat with high content of the sand in sediments) which correlate well with the results of Golemansky [79], who mentioned that D. linearis is a psammophilic species in marine habitats without depth sampling specification. Littoral habitats are generally characterized by a high sand content, but further studies might be needed for a better estimation of the ecological preferences of this species. The same is true for D. giganteacuminata which was observed at depths from 5 to 40 m by Davidova [15]; however, without specification of the exact depths. In our study, we observed high abundances of Centropyxis aculeata on the littoral that correlates with the findings of Patterson et al. [13] who discover C. aculeata in shallow water at depths of less than 10 m. However, a subspecies C. aculeata oblonga was reported in deep water assemblages in Shatura Lakes by Tsyganov et al. [17]. This inconsistency makes us cautions against straightforward interpretations of the ecological preferences of this taxon. For the other mentioned in results species, Centropyxis discoides, C. ecornis, C. laevigata and Galeripora discoides there is a lack of information about their ecological preference, so further studies are needed for fill the gap in our knowledge.
- b.
- Sub-littoral species (6–15 m depth). Four species have a maximal abundance or depth optima in this zone: D. ventricosa, D. claviformis, D. sinuata, and D. petricola. The maximum relative abundance of D. petricola was observed at depths of 9–15 m, which corresponds to the results of Todorov [57], who discovered D. petricola only at depths greater than 5 m in the Smradlivo Ezero lake. D. ventricosa and D. claviformis, in contrast to our findings, were previously registered in littoral biotopes [15,18] with eutrophic and hypertrophic conditions. We assume that these species are distributed at depths of less than 15 m and require biotopes with high organic content and thin sediments. There are no data about the depth preferences of D. sinuata, so we cannot include or exclude this species from the list. Thus, D. petricola can be used as an indicator of depths between 5 m and 15 m and D. ventricosa and D. claviformis occur in a depth range from 0 to 15 m.
- c.
- Deep-Water Species (>15 m). We found that six species Difflugia penardi, D. minuta, D. lithophila, D. pulex, Netzelia oviformis, and Difflugia longicollis had maximal abundance in the deep waters. Similar patterns were reported by Todorov [57] for D. minuta which was observed in the profundal zone; however, this study also reported that D. pulex inhabited not only the profundal zone, but also coastal mosses. This allows us to include D. minuta in the list of deep-depth indicators and excludes D. pulex from this list. Beyens et al. [80] reported D. penardi in biotopes with a pH of 6.52 ± 0.8, whereas in our study it was observed in a slightly acidic environment with a pH of 7.8–7.2 at depths of 24 m or deeper. Therefore, D. penardi may be considered an indicator of deep-water conditions. The maximum abundance of D. lithophila was observed at depths of 15–30 m. Macumber et al. [66] associated this species with the high trophic status of habitats but did not specify the sampling depth. There is no available data on the ecological preferences of D. longicollis. Thus, both species cannot now be set as deep-water indicators. The maximal relative abundance of N. oviformis in deep waters correspond well with the observations of Arrieira [11]. Although this species was also observed in plankton and periphyton, we believe that further researches allow us to prove the ability to use N. oviformis as a proxy of deep waters because the presence of testate amoebae in the plankton should not be considered stochastic [81]. Thus, D. minuta and D. penardi can be used as deep biotope indicators.
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Laminger, H. Quantitative Untersuchung Über Die Testaceenfauna (Protozoa, Rhizopoda) in Den Jüngsten Bodensee-Sedimenten. Biol. Jaarboek 1973, 41, 126–146. [Google Scholar]
- Schonborn, W. The Role of Protozoan Communities in Freshwater and Soil Ecosystems. Acta Protozool. 1992, 3, 11–18. [Google Scholar]
- Laminger, H. Untersuchungen Über Abundanz Und Biomasse Der Sedimentbewohnenden Testaceen (Protozoa, Rhizopoda) in Einem Hochgebirgssee (Vorderer Finstertaler See, Kühtai, Tirol). Int. Rev. Gesamten Hydrobiol. Hydrogr. 1973, 58, 543–568. [Google Scholar] [CrossRef]
- Tsyganov, A.N.; Komarov, A.A.; Mazei, N.G.; Borisova, T.V.; Novenko, E.Y.; Mazei, Y.A. Dynamics of the Species Structure of Testate Amoeba Assemblages in a Waterbody-to-Mire Succession in the Holocene: A Case Study of Mochulya Bog, Kaluga Oblast, Russia. Biol. Bull. 2021, 48, 938–949. [Google Scholar] [CrossRef]
- Mazei, Y.A.; Tsyganov, A.N.; Bobrovsky, M.V.; Mazei, N.G.; Kupriyanov, D.A.; Galka, M.; Rostanets, D.V.; Khazanova, K.P.; Stoiko, T.G.; Pastukhova, Y.A.; et al. Peatland Development, Vegetation History, Climate Change and Human Activity in the Valdai Uplands (Central European Russia) during the Holocene: A Multi-Proxy Palaeoecological Study. Diversity 2020, 12, 462. [Google Scholar] [CrossRef]
- Lotter, A.F. The palaeolimnology of Soppensee (Central Switzerlabnd), as evidenced by diatom, pollen, and fossil-pigment analyses. J. Paleolimnol. 2001, 25, 65–79. [Google Scholar] [CrossRef]
- Frey, D.G. Littoral and Offshore Communities of Diatoms, Cladocerans and Dipterous Larvae, and Their Interpretation in Paleolimnology. J. Paleolimnol. 1988, 1, 179–191. [Google Scholar] [CrossRef]
- Haberyan, K.A. Fossil Diatoms and the Paleolimnology of Lake Rukwa, Tanzania. Freshwater Biol. 1987, 17, 429–436. [Google Scholar] [CrossRef]
- Ndayishimiye, J.C.; Lin, T.; Nyirabuhoro, P.; Zhang, G.; Zhang, W.; Mazei, Y.; Ganjidoust, H.; Yang, J. Decade-Scale Change in Testate Amoebae Community Primarily Driven by Anthropogenic Disturbance than Natural Change in a Large Subtropical Reservoir. Sci. Total Environ. 2021, 784, 147026. [Google Scholar] [CrossRef]
- Beyens, L.; Meisterfeld, R. Protozoa: Testate Amoebae. In Tracking Environmental Changes Using Lake Sediments; Springer: Dordrecht, The Netherlands, 2001; Volume 3, pp. 121–153. [Google Scholar]
- Arrieira, R.L.; Alves, G.M.; Schwind, L.T.F.; Lansac-Tôha, F.A. Local Factors Affecting the Testate Amoeba Community (Protozoa: Arcellinida; Euglyphida) in a Neotropical Floodplain. J. Limnol. 2015, 73, 444–452. [Google Scholar] [CrossRef]
- Escobar, J.; Brenner, M.; Whitmore, T.J.; Kenney, W.F.; Curtis, J.H. Ecology of Testate Amoebae (Thecamoebians) in Subtropical Florida Lakes. J. Paleolimnol. 2008, 40, 715–731. [Google Scholar] [CrossRef]
- Patterson, R.T.; MacKinnon, K.D.; Scott, D.B.; Medioli, F.S. Arcellaceans (Thecamoebians) in Small Lakes of New Brunswick and Nova Scotia: Modern Distribution and Holocene Stratigraphic Changes. J. Foraminif. Res. 1985, 15, 114–137. [Google Scholar] [CrossRef]
- Tolonen, K.; Warner, B.G.; Vasander, H. Ecology of Testaceans (Protozoa: Rhizopoda) in Mires in Southern Finland: II. Multivariate Analysis. Archiv Protistenkunde 1994, 144, 97–112. [Google Scholar] [CrossRef]
- Davidova, R.; Golemansky, V.; Todorov, M. Diversity and Biotopic Distribution of Testate Amoebae (Arcellinida and Euglyphida) in Ticha Dam (Northeastern Bulgaria). Acta Zool. Bulg. 2008, 60, 7–18. [Google Scholar]
- Qin, Y.; Fournier, B.; Lara, E.; Gu, Y.; Wang, H.; Cui, Y.; Zhang, X.; Mitchell, E.A.D. Relationships between Testate Amoeba Communities and Water Quality in Lake Donghu, a Large Alkaline Lake in Wuhan, China. Front. Earth Sci. 2013, 7, 182–190. [Google Scholar] [CrossRef]
- Tsyganov, A.N.; Malysheva, E.A.; Zharov, A.A.; Sapelko, T.V.; Mazei, Y.A. Distribution of Benthic Testate Amoeba Assemblages along a Water Depth Gradient in Freshwater Lakes of the Meshchera Lowlands, Russia, and Utility of the Microfossils for Inferring Past Lake Water Level. J. Paleolimnol. 2019, 62, 137–150. [Google Scholar] [CrossRef]
- Davidova, R.; Vasilev, V. Seasonal Dynamics of the Testate Amoebae Fauna (Protozoa: Arcellinida and Euglyphida) in Durankulak Lake (Northeastern Bulgaria). Acta Zool. Bulg. 2013, 65, 27–36. [Google Scholar]
- Mazei, Y.A.; Tsyganov, A.N. Testate Amoebae from Freshwater Ecosystems of the Sura River Basin (Middle Volga Region). 1. Fauna and Morphological-Ecological Characteristics of Species. Zool. Zhurnal 2006, 85, 1267–1280. [Google Scholar]
- Dalby, A.P.; Kumar, A.; Moore, J.M.; Patterson, R.T. Preliminary Survey Of Arcellaceans (Thecamoebians) As Limnological Indicators In Tropical Lake Sentani, Irian Jaya, Indonesia. J. Foraminif. Res. 2000, 30, 135–142. [Google Scholar] [CrossRef]
- Mazei, Y.A.; Tsyganov, A.N. Testate Amoebae from Freshwater Ecosystems of the Sura River Basin (Middle Volga Region). 2. Community Structure. Zool. Zhurnal 2006, 85, 1395–1401. [Google Scholar]
- Legesse, D.; Gasse, F.; Radakovitch, O.; Vallet-Coulomb, C.; Bonnefille, R.; Verschuren, D.; Gibert, E.; Barker, P. Environmental Changes in a Tropical Lake (Lake Abiyata, Ethiopia) during Recent Centuries. Palaeogeogr. Palaeoclimatol. Palaeoecol. 2002, 187, 233–258. [Google Scholar] [CrossRef]
- Verschuren, D. Lake-Based Climate Reconstruction in Africa: Progress and Challenges. Hydrobiologia 2003, 500, 315–330. [Google Scholar] [CrossRef]
- Davidova, R. Testate Amoebae Communities (Protozoa: Arcellinida and Euglyphida) in the Rabisha Reservoir (Northwestern Bulgaria). Acta Zool. Bulg. 2010, 62, 259–269. [Google Scholar]
- Todorov, M. Testate Amoebae (Protozoa, Rhizopoda) of the Ribni Ezera Glacial Lakes in the Rila Mountains (South- West Bulgaria). Acta Zool. Bulg. 2004, 56, 243–252. [Google Scholar]
- Roe, H.M.; Patterson, R.T.; Swindles, G.T. Controls on the Contemporary Distribution of Lake Thecamoebians (Testate Amoebae) within the Greater Toronto Area and Their Potential as Water Quality Indicators. J. Paleolimnol. 2010, 43, 955–975. [Google Scholar] [CrossRef]
- Davidova, R.; Boycheva, M. Testate Amoebae Fauna (Amoebozoa, Rhizaria) from the Benthal of Kamchia Reservoir (Eastern Bulgaria). Acta Zool. Bulg. 2015, 67, 375–384. [Google Scholar]
- Barnett, R.L.; Dan J. Charman, D.J.; Roland, W.G.; Margot, H.; Saher, M.H.; Marshall, W.A. Testate Amoebae as Sea-Level Indicators in Northwestern Norway: Developments in Sample Preparation and Analysis. Acta Protozool. 2013, 52, 115–128. [Google Scholar]
- Nedogarko, I.V. (Ed.) Valdai Lakes (Review of Observation Results for 1946–2018), 1st ed.; Svoe Izdatelstvo: St Peterburg, Russia, 2021; p. 250. [Google Scholar]
- Kitaev, S.P. Ecological Bases of Bioproductivity of Lakes of Different Natural Zones (Tundra, Taiga, Mixed Forest); Nauka: Moscow, Russia, 1984; p. 529. [Google Scholar]
- Payne, R.J.; Mitchell, E.A.D. How Many Is Enough? Determining Optimal Count Totals for Ecological and Palaeoecological Studies of Testate Amoebae. J. Paleolimnol. 2009, 42, 483–495. [Google Scholar] [CrossRef]
- Todorov, M.; Bankov, N. An Atlas of Sphagnum-Dwelling Testate Amoebae in Bulgaria; Pensoft Publishers: Sofia, Bulgaria, 2019; Volume 1, p. e38685. [Google Scholar]
- Mazei, Y.A.; Tsyganov, A.N. Freshwater Testate Amoebae; KMK: Moscow, Russia, 2006. [Google Scholar]
- Core. R: A Language and Environment for Statistical Computing; R Foundation for Statistical Computing: Vienna, Austria, 2018.
- Vegan: Community Ecology Package; R Package Version 2.4-3; R Foundation for Statistical Computing: Vienna, Austria, 2022.
- Juggins, S. Rioja: Analysis of Quaternary Science Data; R Package Version (0.9-9); R Foundation for Statistical Computing: Vienna, Austria, 2020. [Google Scholar]
- Pielou, E.C. Mathematical Ecology; John Wiley & Sons Ltd.: Chichester, UK, 1977; p. 385. [Google Scholar]
- Grimm, E.C. CONISS: A FORTRAN 77 Program for Stratigraphically Constrained Cluster Analysis by the Method of Incremental Sum of Squares. Comp. Geosci. 1987, 13, 13–35. [Google Scholar] [CrossRef]
- Labdsv: Ordination and Multivariate Analysis for Ecology; R Foundation for Statistical Computing: Vienna, Austria, 2019.
- Clarke, K.R. Comparisons of Dominance Curves. J. Exp. Marine Biol. Ecol. 1990, 138, 143–157. [Google Scholar] [CrossRef]
- González-Miguéns, R.; Todorov, M.; Blandenier, Q.; Duckert, C.; Porfirio-Sousa, A.L.; Ribeiro, G.M.; Ramos, D.; Lahr, D.J.G.; Buckley, D.; Lara, E. Deconstructing Difflugia: The Tangled Evolution of Lobose Testate Amoebae Shells (Amoebozoa: Arcellinida) Illustrates the Importance of Convergent Evolution in Protist Phylogeny. Mol. Phylogenet. Evol. 2022, 175, 107557. [Google Scholar] [CrossRef] [PubMed]
- Nasser, N.A.; Gregory, B.R.B.; Singer, D.; Patterson, R.T.; Roe, H.M. Erugomicula, a New Genus of Arcellinida (Testate Lobose Amoebae). Palaeontol. Electr. 2021, 24, a16. [Google Scholar] [CrossRef]
- Neville, L.A.; McCarthy, F.M.G.; MacKinnon, M.D. Seasonal Environmental and Chemical Impact on Thecamoebian Community Composition in an Oil Sands Reclamation Wetland in Northern Alberta. Palaeontol. Electr. 2010, 13, 1–14. [Google Scholar]
- Ju, L.; Yang, J.; Liu, L.; Wilkinson, D.M. Diversity and Distribution of Freshwater Testate Amoebae (Protozoa) along Latitudinal and Trophic Gradients in China. Microb. Ecol. 2014, 68, 657–670. [Google Scholar] [CrossRef]
- Davidova, R.; Sevginov, S. Diversity and Distribution of Testate Amoebae (Amoebozoa, Rhizaria) in Reservoirs, Northeastern Bulgaria. Acta Sci. Nat. 2018, 5, 90–99. [Google Scholar] [CrossRef]
- Lorencova, M. Thecamoebians from Recent Lake Sediments from the ˇSumava Mts, Czech Republic. Bull. Geosci. 2009, 84, 359–376. [Google Scholar] [CrossRef][Green Version]
- Yang, J.; Zhang, W.; Feng, W.; Shen, Y. Freshwater Testate Amoebae of Nine Yunnan Plateau Lakes, China. J. Freshwater Ecol. 2005, 20, 743–750. [Google Scholar] [CrossRef][Green Version]
- Barnett, R.L.; Gehrels, W.R.; Charman, D.J.; Saher, M.H.; Marshall, W.A. Late Holocene Sea-Level Change in Arctic Norway. Quarter. Sci. Rev. 2015, 107, 214–230. [Google Scholar] [CrossRef]
- Cockburn, C.F.; Gregory, B.R.B.; Nasser, N.A.; Patterson, R.T. Intra-Lake Arcellinida (Testate Lobose Amoebae) Response to Winter de-Icing Contamination in an Eastern Canada Road-Side “Salt Belt” Lake. Microb. Ecol. 2020, 80, 366–383. [Google Scholar] [CrossRef]
- Nasser, N. Lacustrine Arcellinida (Testate Lobose Amoebae) as Bioindicators of Arsenic Contamination: A New Tool for Environmental Risk Assessment. Ph.D. Thesis, Carleton University, Ottawa, ON, Canada, 2020. [Google Scholar]
- Patterson, R.T.; Lamoureux, E.D.R.; Neville, L.A.; Macumber, A.L. Arcellacea (Testate Lobose Amoebae) as PH Indicators in a Pyrite Mine-Acidified Lake, Northeastern Ontario, Canada. Microb. Ecol. 2013, 65, 541–554. [Google Scholar] [CrossRef] [PubMed]
- Kumar, A.; Patterson, R.T. Arcellaceans (Thecamoebians): New Tools for Monitoring Long-and Short-Term Changes in Lake Bottom Acidity. Environ. Geol. 2000, 39, 689–697. [Google Scholar] [CrossRef]
- Azovsky, A.I.; Mazei, Y.A. Diversity and Distribution of Free-Living Ciliates from High-Arctic Kara Sea Sediments. Protist 2018, 169, 141–157. [Google Scholar] [CrossRef] [PubMed]
- Wlodarska-Kowalczuk, M.; Pearson, T.H. Soft-Bottom Macrobenthic Faunal Associations and Factors Affecting Species Distributions in an Arctic Glacial Fjord (Kongsfjord, Spitsbergen). Polar Biol. 2004, 27, 155–167. [Google Scholar] [CrossRef]
- Schallenberg, M.; Kalff, J. The Ecology of Sediment Bacteria in Lakes and Comparisons with Other Aquatic Ecosystems. Ecology 1993, 74, 919–934. [Google Scholar] [CrossRef]
- Davidova, R. Biotopic Distribution of Testate Amoebae (Protozoa: Arcellinida and Euglyphida) in Ovcharitsa Reservoir (Southeastern Bulgaria). Acta Zool. Bulg. 2012, 64, 13–22. [Google Scholar]
- Todorov, M. Testate Amoebae (Protozoa, Rhizopoda) of the Smradlivo Ezero Glacial Lake in the Rila National Park (Southwestern Bulgaria). Acta Zool. Bulg. 2005, 57, 13–23. [Google Scholar]
- Golemansky, V.; Todorov, M.; Temelkov, B. Biotopic Distribution of the Testate Amoebae (Rhizopoda: Arcellinida and Gromiida) in the Tectonic Lake Doiran (Southeast Europe). Acta Zool. Bulg. 2005, 57, 3–11. [Google Scholar]
- Dabés, M.B.G.S.; Velho, L.F.M. Assemblage of Testate Amoebae (Protozoa, Rhizopoda) Associated to Aquatic Macrophytes Stands in a Marginal Lake of the São Francisco River Floodplain, Brazil. Acta Sci. 2001, 23, 299–304. [Google Scholar]
- Schönborn, W. Die Ökologie Der Testaceen Im Oligotrophen See: Dargestellt Am Beispiel Des Großen Stechlinsees. Ph.D. Thesis, Friedrich-Schiller-Universität, Jena, Germany, 1961. [Google Scholar]
- Regalado, S.I.; García, L.S.; Alvarado, P.L.; Caballero, M.; Vázquez, L.A. Ecological Drivers of Testate Amoeba Diversity in Tropical Water Bodies of Central Mexico. J. Limnol. 2018, 77, 385–399. [Google Scholar] [CrossRef]
- Lisa, A.N.; David, G.C.; Francine, M.M.; Michael, D.M. Biogeographic Variation in Thecamoebian (Testate Amoeba) Assemblages in Lakes within Various Vegetation Zones of Alberta, Canada. Int. J. Biodivers. Conserv. 2010, 2, 215–224. [Google Scholar]
- Steele, R.E.; Nasser, N.A.; Patterson, R.T.; Gregory, B.R.B.; Roe, H.M.; Reinhardt, E.G. An Assessment of Sub-Meter Scale Spatial Variability of Arcellinida (Testate Lobose Amoebae) Assemblages in a Temperate Lake: Implications for Limnological Studies. Microb. Ecol. 2018, 76, 680–694. [Google Scholar] [CrossRef] [PubMed]
- Golemansky, V.G.; Todorov, M.T. Testate Amoebae (Protozoa: Rhizopoda) from Thailand. Acta Protozool. 2000, 39, 337–344. [Google Scholar]
- Torigai, K.; Schröder-Adams, C.J.; Burbidge, S.M. A Variable Lacustrine Environment in Lake Winnipeg, Manitoba: Evidence from Modern Thecamoebian Distribution. J. Paleolimnol. 2000, 23, 305–318. [Google Scholar] [CrossRef]
- Macumber, A.L.; Roe, H.M.; Prentice, S.V.; Sayer, C.D.; Bennion, H.; Salgado, J. Freshwater Testate Amoebae (Arcellinida) Response to Eutrophication as Revealed by Test Size and Shape Indices. Front. Ecol. Evol. 2020, 8, 568904. [Google Scholar] [CrossRef]
- Ivanova, K.O.; Kosheleva, T.N. Comparative analysis of structure of species dominance of marine Nematoda assemblages (Sevastopol Bays). Optimiz. Protect. Ecosyst. 2012, 7, 209–216. [Google Scholar]
- Nedogarko, I.V.; Kuznetsova, Y.N.; Reshetnikov, F.Y. Bottom Sediments of Lakes as an Indicator of Dynamic Factors. Meteorol. I Gidrol. 2005, 11, 86–94. [Google Scholar]
- Wiik, E.; Bennion, H.; Sayer, C.D.; Davidson, T.A.; Clarke, S.J.; McGowan, S.; Prentice, S.; Simpson, G.L.; Stone, L. The Coming and Going of a Marl Lake: Multi-Indicator Palaeolimnology Reveals Abrupt Ecological Change and Alternative Views of Reference Conditions. Front. Ecol. Evol. 2015, 3, 82. [Google Scholar] [CrossRef]
- Burdíková, Z.; Čapek, M.; Švindrych, Z.; Gryndler, M.; Kubínová, L.; Holcová, K. Ecology of testate amoebae in the Komořany Ponds in the Vltava Basin. Microb. Ecol. 2012, 64, 117–130. [Google Scholar] [CrossRef]
- Balik, V.; Song, B. Benthic freshwater testate amoebae assemblages (Protozoa: Rhizopoda) from Lake Dongting, People’s Republic of China, with description of a new species from the genus Collaripyxidia. Acta Protozoologica. 2000, 39, 149–156. [Google Scholar]
- Bonetti, C.H.C.; Eichler, B.B. Associações de Foraminíferos e Tecamebas Indicadoras de Sub-Ambientes Recentes Na Zona Estuarina Do Rio Itapitangui-Cananéia/SP. Master’s Thesis, Universidade de São Paulo, Brasil, Brasil, 1995. [Google Scholar]
- Alves, G.M.; Velho, L.F.M.; de Morais Costa, D.; Lansac-Tôha, F.A. Size Structure of Testate Amoebae (Arcellinida and Euglyphida) in Different Habitats from a Lake in the Upper Paraná River Floodplain. Eur. J. Protistol. 2012, 48, 169–177. [Google Scholar] [CrossRef] [PubMed]
- Jiménez, C.; Springer, M. Depth Related Distribution of Benthic Macrofauna in a Costa Rican Crater Lake. Rev. Biol. Trop. 1996, 44, 673–678. [Google Scholar] [PubMed]
- Celis, C.N.F.; Garibay, M.; Sigala, I.; Brenner, M.; Galindo, E.P.; García, L.S.; Massaferro, J.; Pérez, L. Testate Amoebae (Amoebozoa: Arcellinidae) as Indicators of Dissolved Oxygen Concentration and Water Depth in Lakes of the Lacandón Forest, Southern Mexico. J. Limnol. 2019, 79, 81–91. [Google Scholar] [CrossRef]
- Kihlman, S.; Kauppila, T. Effects of Mining on Testate Amoebae in a Finnish Lake. J. Paleolimnol. 2012, 47, 1–15. [Google Scholar] [CrossRef]
- Tran, H.Q. First Data on Testate Amoeba Composition in Tropical Karst Wetlands of Northern Vietnam in Relation to Type of Biotope and Season: New Bioindication Potentialities. Inland Water Biol. 2020, 13, 251–261. [Google Scholar] [CrossRef]
- Lamentowicz, L.; Gabka, M.; Lamentowicz, M. Species Composition of Testate Amoebae (Protists) and Environmental Parameters in a Sphagnum Peatland. Polish J. Ecol. 2007, 55, 749. [Google Scholar]
- Golemansky, V. Testate Amoebas and Monothalamous Foraminifera (Protozoa) from the Bulgarian Black Sea Coast. In Biogeography and Ecology of Bulgaria; Springer: Berlin/Heidelberg, Germany, 2007; pp. 555–570. [Google Scholar]
- Beyens, L.; Chardez, D.; de Baere, D.; Verbruggen, C. The Aquatic Testate Amoebae Fauna of the Strømness Bay Area, South Georgia. Antarc. Sci. 1995, 7, 3–8. [Google Scholar] [CrossRef]
- Alves, G.M.; Velho, L.F.M.; Simões, N.R.; Lansac-Tôha, F.A. Biodiversity of Testate Amoebae (Arcellinida and Euglyphida) in Different Habitats of a Lake in the Upper Paraná River Floodplain. Eur. J. Protistol. 2010, 46, 310–318. [Google Scholar]









| Genus. | Number of Species | Number of Shells | Species Share, % | Shell Share, % |
|---|---|---|---|---|
| Arcella Ehrenberg, 1830 | 4 | 295 | 4 | 3.1% |
| Centropyxis Stein, 1857 | 18 | 1803 | 18 | 18.7% |
| Cryptodifflugia Penard, 1890 | 5 | 10 | 5 | 0.1% |
| Cyclopyxix Deflandre, 1929 | 3 | 60 | 3 | 0.6% |
| Cylindrifflugia González-Miguéns et al., 2022 | 5 | 1567 | 5 | 16.2% |
| Cyphoderia Schlumberger, 1845 | 3 | 15 | 3 | 0.2% |
| Difflugia Leclerc, 1815 | 44 | 4817 | 44 | 49.8% |
| Erugomicula Nasser et al., 2022 [42] | 1 | 6 | 1 | 0.1% |
| Galeripora González-Miguéns et al., 2022 | 2 | 142 | 2 | 1.5% |
| Golemanskia González-Miguéns et al., 2022 | 1 | 91 | 1 | 0.9% |
| Hyalosphenia Stein, 1857 | 1 | 6 | 1 | 0.1% |
| Lagenodiffludia Medioli et Scott, 1983 | 1 | 1 | 1 | 0.0% |
| Lesquereusia Schlumberger, 1845 | 1 | 39 | 1 | 0.4% |
| Nebela Leidy, 1874 | 2 | 3 | 2 | 0.0% |
| Netzelia Ogden, 1979 | 2 | 619 | 2 | 6.4% |
| Oopyxis Jung, 1942 | 1 | 1 | 1 | 0.0% |
| Paraqudrula Deflandre, 1932 | 1 | 2 | 1 | 0.0% |
| Pontigulasia Rhumbler, 1896 | 3 | 113 | 3 | 1.2% |
| Pseudodifflugia Schlumberger, 1845 | 1 | 7 | 1 | 0.1% |
| Trigonopyxis Penard, 1912 | 1 | 1 | 1 | 0.0% |
| Zivkovicia Ogden, 1987 | 1 | 69 | 1 | 0.7% |
| Total | 101 | 9667 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Sysoev, V.V.; Tsyganov, A.N.; Reshetnikov, F.Y.; Mazei, Y.A. The Effects of Sampling Depth on Benthic Testate Amoeba Assemblages in Freshwater Lakes: A Case Study in Lake Valdayskoe (the East European Plain). Diversity 2022, 14, 974. https://doi.org/10.3390/d14110974
Sysoev VV, Tsyganov AN, Reshetnikov FY, Mazei YA. The Effects of Sampling Depth on Benthic Testate Amoeba Assemblages in Freshwater Lakes: A Case Study in Lake Valdayskoe (the East European Plain). Diversity. 2022; 14(11):974. https://doi.org/10.3390/d14110974
Chicago/Turabian StyleSysoev, Vlad V., Andrey N. Tsyganov, Fedor Y. Reshetnikov, and Yuri A. Mazei. 2022. "The Effects of Sampling Depth on Benthic Testate Amoeba Assemblages in Freshwater Lakes: A Case Study in Lake Valdayskoe (the East European Plain)" Diversity 14, no. 11: 974. https://doi.org/10.3390/d14110974
APA StyleSysoev, V. V., Tsyganov, A. N., Reshetnikov, F. Y., & Mazei, Y. A. (2022). The Effects of Sampling Depth on Benthic Testate Amoeba Assemblages in Freshwater Lakes: A Case Study in Lake Valdayskoe (the East European Plain). Diversity, 14(11), 974. https://doi.org/10.3390/d14110974