
Biodiversity and Phylogeny of North Atlantic Euphrosinidae (Annelida)



Abstract
:1. Introduction
2. Materials and Methods
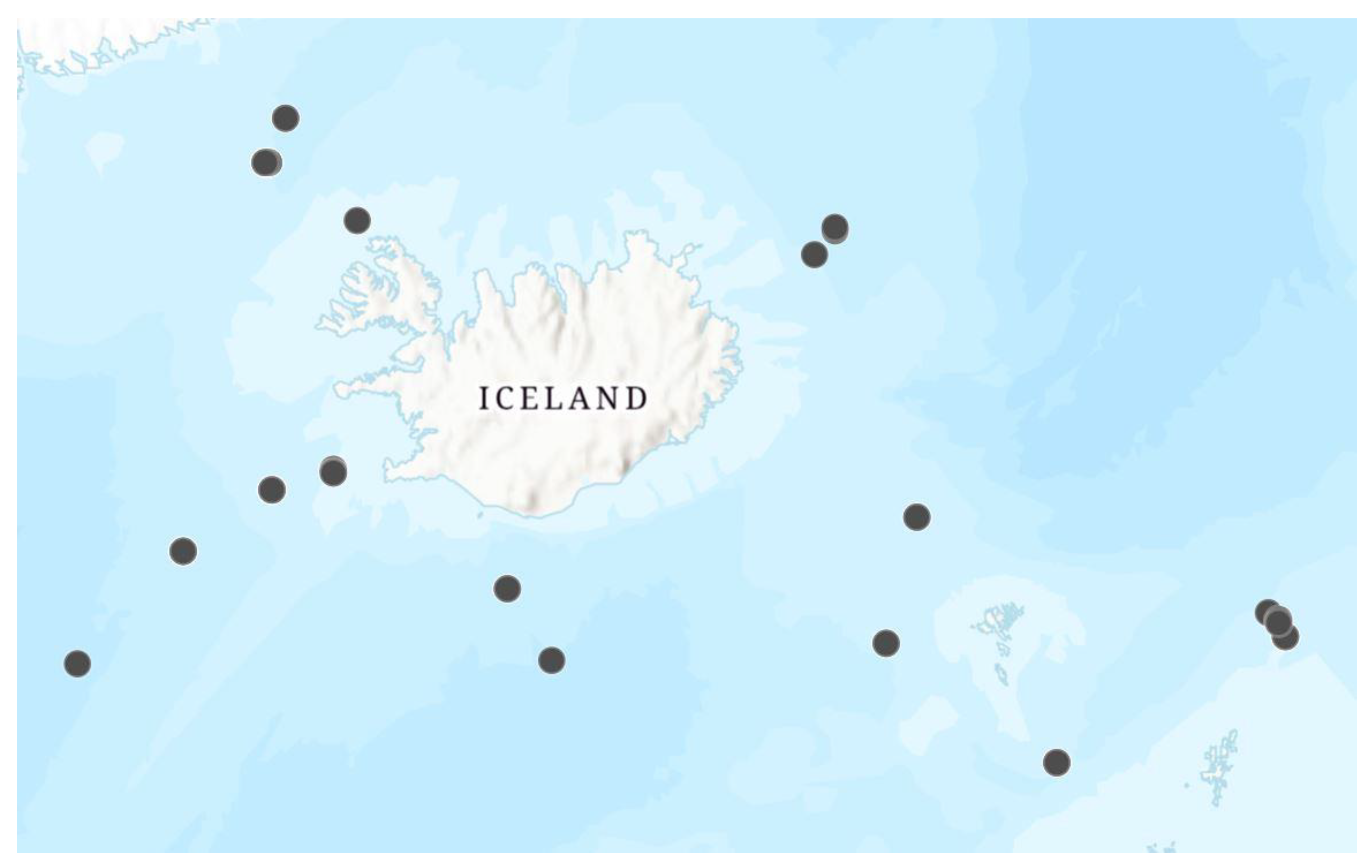
2.1. Sample Collection
2.2. Imaging
2.3. DNA Extraction, Amplification, and Sequencing
2.4. Phylogenetic Analysis of Euphrosinidae (16S + 28S)
3. Results and Discussion
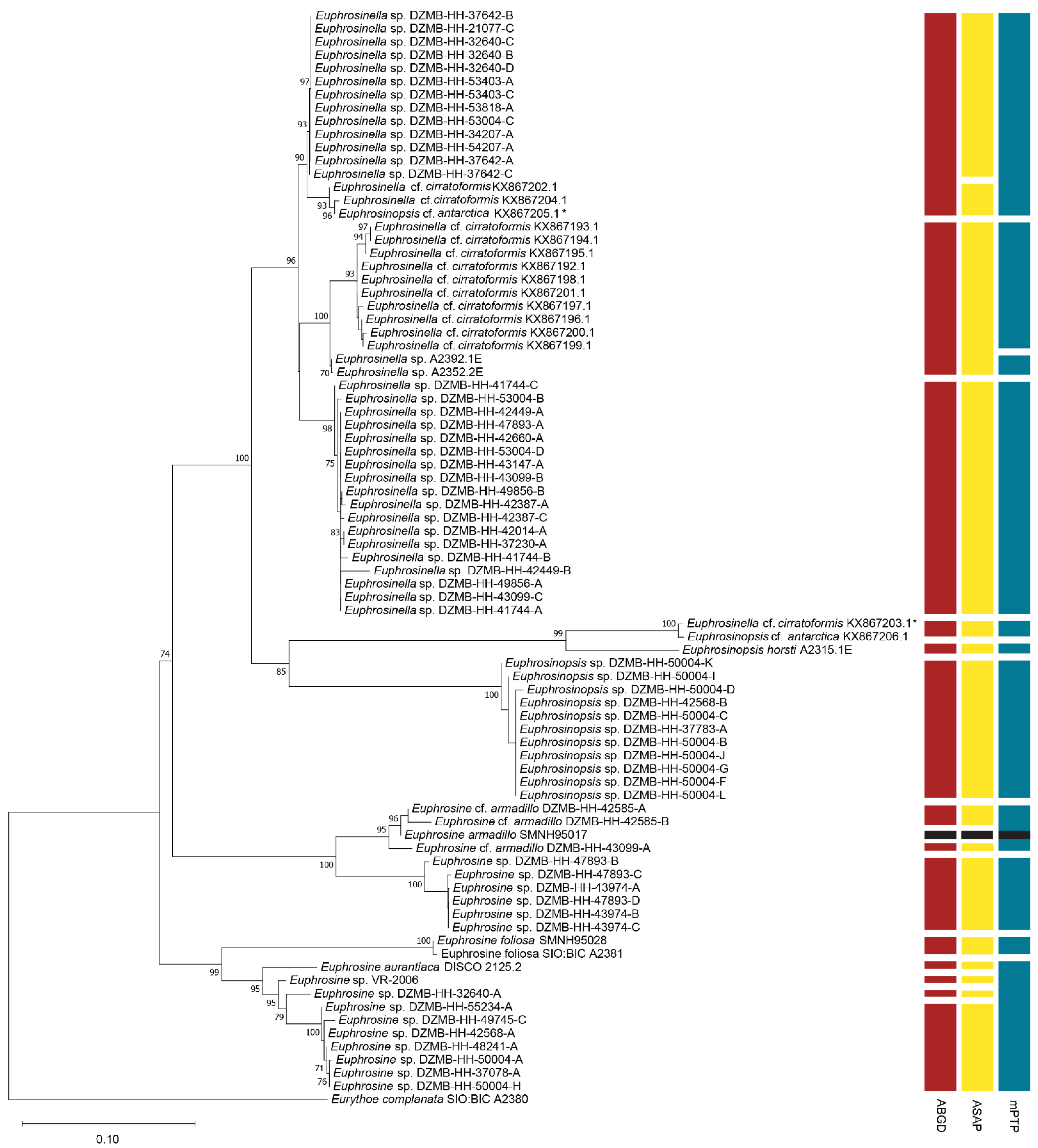
3.1. Diversity and Phylogeny of Euphrosinidae
3.2. Analysis of Contaminant Sequences
4. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Müller, J.; Schumacher, A.; Borda, E.; Rouse, G.W.; Bartolomaeus, T.; Tilic, E. “Brittleworms”: Ultrastructure and arrangement of the calcified chaetae of Euphrosine (Amphinomida, Annelida). Invertebr. Biol. 2021, 140, e12353. [Google Scholar] [CrossRef]
- Kudenov, J. Amphinomidae and Euphrosinidae (Annelida: Polychaeta) Principally from Antarctica, The Southern Ocean, and Subantarctic Regions. Ant. Acc. 1993, 58, 93–150. [Google Scholar] [CrossRef]
- Borda, E.; Kudenov, J.D. Euphrosinidae (Annelida: Amphinomida) Collected from Antarctica (R/V Polarstern, 1984, 1986) with Comments on the Generic Placement of Euphrosine Magellanica Ehlers, 1900. PBSW 2014, 126, 299–311. [Google Scholar] [CrossRef]
- Alalykina, I.L. Composition of Deep-Sea Polychaetes from the SokhoBio Expedition with a Description of a New Species of Labioleanira (Annelida: Sigalionidae) from the Sea of Okhotsk. Deep. Sea Res. Part II Top. Stud. Oceanogr. 2018, 154, 140–158. [Google Scholar] [CrossRef]
- WoRMS Editorial Board World Register of Marine Species. Available online: https://www.marinespecies.org (accessed on 9 October 2020).
- Gil, J.C.F. The European Fauna of Annelida Polychaeta. Ph.D. Thesis, Faculdade de Ciências, Universidade de Lisboa, Lisboa, Portugal, 2011. Available online: http://hdl.handle.net/10451/4600 (accessed on 7 September 2022).
- Carr, C.M.; Hardy, S.M.; Brown, T.M.; Macdonald, T.A.; Hebert, P.D.N. A Tri-Oceanic Perspective: DNA Barcoding Reveals Geographic Structure and Cryptic Diversity in Canadian Polychaetes. PLoS ONE 2011, 6, e22232. [Google Scholar] [CrossRef] [Green Version]
- Brasier, M.J.; Wiklund, H.; Neal, L.; Jeffreys, R.; Linse, K.; Ruhl, H.; Glover, A.G. DNA Barcoding Uncovers Cryptic Diversity in 50% of Deep-Sea Antarctic Polychaetes. R. Soc. Open Sci. 2016, 3, 160432. [Google Scholar] [CrossRef] [Green Version]
- Hartmann-Schröder, G.; Rosenfeldt, P. Die Polychaeten Der “Polarstern”-Reise ANT V/1 in Die Antarktis 1986. Teil 1: Euphrosinidae Bis Iphitimidae. Mitt. Hamb. Zool. Mus. Inst. 1992, 89, 85–124. [Google Scholar]
- Horst, R. New Species of the Euphrosyne from the Siboga-Expedition, with a Table of the Species Hitherto Known. Notes Leyden Mus. 1903, 23, 213–222. [Google Scholar]
- Johnson, H.P.; Johnson, H.P. A Preliminary Account of the Marine Annelids of the Pacific Coast, with Descriptions of New Species. Proc. Calif. Acad. Sci. 1897, 1, 153–198. [Google Scholar]
- Wiklund, H.; Nygren, A.; Pleijel, F.; Sundberg, P. The Phylogenetic Relationships between Amphinomidae, Archinomidae and Euphrosinidae (Amphinomida: Aciculata: Polychaeta), Inferred from Molecular Data. J. Mar. Biol. Assoc. U. K. 2008, 88, 509–513. [Google Scholar] [CrossRef]
- Halanych, K.M.; Janosik, A.M. A Review of Molecular Markers Used for Annelid Phylogenetics. Integr. Comp. Biol. 2006, 46, 533–543. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Brix, S.; Meißner, K.; Stransky, B.; Halanych, K.; Jennings, R.; Kocot, K.; Svavarsson, J. The IceAGE Project—A Follow up of BIOICE. Pol. Polar Res. 2014, 35, 141–150. [Google Scholar] [CrossRef]
- Bakken, T.; Kongsrud, J.A.; Oug, E.; Cochrane, S.; Moen, T.L.; Solbakken, B.E.B. Polychaetes from Jan Mayen (Annelida, Polychaeta). Polar Res. 2010, 29, 1–21. [Google Scholar] [CrossRef]
- Riehl, T.; Brenke, N.; Brix, S.; Driskell, A.; Kaiser, S.; Brandt, A. Field and Laboratory Methods for DNA Studies on Deep-Sea Isopod Crustaceans. Pol. Polar Res. 2014, 35, 203–224. [Google Scholar] [CrossRef] [Green Version]
- Palumbi, S. The Simple Fool’s Guide to PCR Version 2.0, Saturday, 27 July 1991; Department of Zoology and Kewalo Marine Laboratory: Honolulu, HI, USA, 1991; p. 47. [Google Scholar]
- Minh, B.Q.; Schmidt, H.A.; Chernomor, O.; Schrempf, D.; Woodhams, M.D.; von Haeseler, A.; Lanfear, R. IQ-TREE 2: New Models and Efficient Methods for Phylogenetic Inference in the Genomic Era. Mol. Biol. Evol. 2020, 37, 1530–1534. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Edgar, R.C. MUSCLE: Multiple Sequence Alignment with High Accuracy and High Throughput. Nucleic Acids Res. 2004, 32, 1792–1797. [Google Scholar] [CrossRef] [Green Version]
- Tamura, K.; Stecher, G.; Kumar, S. MEGA11: Molecular Evolutionary Genetics Analysis Version 11. Mol. Biol. Evol. 2021, 38, 3022–3027. [Google Scholar] [CrossRef]
- Kück, P.; Longo, G.C. FASconCAT-G: Extensive Functions for Multiple Sequence Alignment Preparations Concerning Phylogenetic Studies. Front. Zool. 2014, 11, 81. [Google Scholar] [CrossRef] [Green Version]
- Chernomor, O.; von Haeseler, A.; Minh, B.Q. Terrace Aware Data Structure for Phylogenomic Inference from Supermatrices. Syst. Biol. 2016, 65, 997–1008. [Google Scholar] [CrossRef] [Green Version]
- Puillandre, N.; Lambert, A.; Brouillet, S.; Achaz, G. ABGD, Automatic Barcode Gap Discovery for Primary Species Delimitation. Mol. Ecol. 2012, 21, 1864–1877. [Google Scholar] [CrossRef]
- Puillandre, N.; Brouillet, S.; Achaz, G. ASAP: Assemble species by automatic partitioning. Mol. Ecol. Resour. 2021, 21, 609–620. [Google Scholar] [CrossRef] [PubMed]
- Kapli, T.; Lutteropp, S.; Zhang, J.; Kobert, K.; Pavlidis, P.; Stamatakis, A.; Flouri, T. Multi-rate Poisson tree processes for single-locus species delimitation under maximum likelihood and Markov chain Monte Carlo. Bioinformatics 2016, 33, 1630–1638. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rousset, V.; Pleijel, F.; Rouse, G.W.; Erséus, C.; Siddall, M.E. A Molecular Phylogeny of Annelids. Cladistics 2007, 23, 41–63. [Google Scholar] [CrossRef] [PubMed]
- Gillet, P.; Dauvin, J.-C. Polychaetes from the Atlantic Seamounts of the Southern Azores: Biogeographical Distribution and Reproductive Patterns. J. Mar. Biol. Assoc. UK 2000, 80, 1019–1029. [Google Scholar] [CrossRef]
- Altschul, S.F.; Gish, W.; Miller, W.; Myers, E.W.; Lipman, D.J. Basic Local Alignment Search Tool. J. Mol. Biol. 1990, 215, 403–410. [Google Scholar] [CrossRef]
- Ramos, J.; Martín, G.; Sikorski, A. Syllidae (Polychaeta) from the Arctic and Sub-Arctic Regions. J. Mar. Biol. Assoc. UK 2010, 90, 1041–1050. [Google Scholar] [CrossRef]
- Blake, J.A. Bitentaculate Cirratulidae (Annelida, Polychaeta) Collected Chiefly during Cruises of the R/V Anton Bruun, USNS Eltanin, USCG Glacier, R/V Hero, RVIB Nathaniel B. Palmer, and R/V Polarstern from the Southern Ocean, Antarctica, and off Western South America. Zootaxa 2018, 4537, 1–130. [Google Scholar] [CrossRef] [Green Version]
- Arteaga-Flórez, C.; Bolaño-Lara, M.; Arteaga-Flórez, C.; Bolaño-Lara, M. Benthic Polychaetes Collected in the III Expedición “Admiral Padilla” Austral Summer 2016-2017, to the North of the Antarctic Peninsula. Boletín Investig. Mar. Costeras—INVEMAR 2021, 50, 221–226. [Google Scholar] [CrossRef]
- Würzberg, L.; Peters, J.; Schüller, M.; Brandt, A. Diet Insights of Deep-Sea Polychaetes Derived from Fatty Acid Analyses. Deep. Sea Res. Part II Top. Stud. Oceanogr. 2011, 58, 153–162. [Google Scholar] [CrossRef]
- Fauchald, K.; Jumars, P.A. The Diet of Worms: A Study of Polychaete Feeding Guilds. Oceanogr. Mar. Biol. Annu. Rev. 1979, 17, 193–284. [Google Scholar]




{kind=link}
{kind=link}
{kind=link}
| Genus | Eyes | Prostomium | Caruncle | Chaetae |
|---|---|---|---|---|
| Euphrosine | Two pairs of eyes | Prostomium with three appendages | Caruncle attached to body wall | Simple bifurcate chaetae; ringent chaetae and aciculae may be present |
| Euphrosinopsis | One large pair of eyes | Prostomium with five appendages | Caruncle not attached to the body wall for most of its length | Simple bifurcate chaetae; ringent chaetae may be present |
| Euphrosinella | Two pairs of eyes | Prostomium with five appendages | Caruncle not attached to the body wall for most of its length | Simple bifurcate chaetae |
| Primer Name | Sequence 5′ to 3′ |
|---|---|
| 16S arL | CGCCTGTTTATCAAAAACAT |
| 16S brH | CCGGTCTGAACTCAGATCACGT |
| 28S_Amph_F1 | ACCCGCTGAAYTTAAGCATATCAC |
| 28S_Amph_F2 | ACAAGTACCGTGAGGGAAAGTTG |
| 28S_Amph_R | CTTGGTCCGTGTTTCAAGACG |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Batts, R.A.; Whitman, K.N.; Meißner, K.; Kocot, K.M. Biodiversity and Phylogeny of North Atlantic Euphrosinidae (Annelida). Diversity 2022, 14, 996. https://doi.org/10.3390/d14110996
Batts RA, Whitman KN, Meißner K, Kocot KM. Biodiversity and Phylogeny of North Atlantic Euphrosinidae (Annelida). Diversity. 2022; 14(11):996. https://doi.org/10.3390/d14110996
Chicago/Turabian StyleBatts, Rowan A., Karsyn N. Whitman, Karin Meißner, and Kevin M. Kocot. 2022. "Biodiversity and Phylogeny of North Atlantic Euphrosinidae (Annelida)" Diversity 14, no. 11: 996. https://doi.org/10.3390/d14110996
APA StyleBatts, R. A., Whitman, K. N., Meißner, K., & Kocot, K. M. (2022). Biodiversity and Phylogeny of North Atlantic Euphrosinidae (Annelida). Diversity, 14(11), 996. https://doi.org/10.3390/d14110996